

Introduction
Простейшим паразитом, Trypanosoma brucei, является возбудителем заболеваний , которые влияют на людей (через Trypanosoma brucei gambiense и Trypanosoma brucei rhodesiense) и животных (через Trypanosoma brucei brucei) во всей Африке к югу от Сахары. Он передается хозяину млекопитающего через слюну цеце вектора мухи. Оба человека и животных Африканский трипаносомоз вызывают серьезное экономическое бремя в эндемичных районах, а также несколько препаратов доступны или в развитии для лечения этих болезней. Понимание механизмов иммунной уклонения имеет решающее значение для разработки лекарств для трипаносомозу. Антигенная изменчивость плотного вариантные покрытия поверхности гликопротеина (VSG) , который покрывает паразита является одним из основных средств , с помощью которых Т. brucei уклоняется реакции антитела млекопитающих. Около 2000 вариантов гена VSG существуют в геноме трипаносом, но только один транскрибируется в любой момент времени от одного из ~ 15 Expresсайты Sion. "Переключение" выраженного VSG может происходить либо путем копирования гена VSG в сайт активного выражения, или путем транскрипционной активации ранее немого сайта выражения (обзор 1).
Хотя большая работа посвящена исследованию антигенной изменчивости T. brucei, факторы, влияющие на частоту переключения до сих пор плохо изучены. Исследования на сегодняшний день было затруднено тем , что в то время как переключение происходит стохастически в пробирке, частоты переключения очень низкие, порядка 1 в приблизительно 10 6 клеток 2. Это делает его трудно измерить ли уменьшение заданного возрастает фактор или частоту переключения, так как переключение трудно обнаружить в первую очередь. До 2009 года, методы выделения коммутаторами в данной популяции были длительными и трудоемкими. К ним относятся пассажей трипаносом через мышей, иммунизированных против доминирующего VSG, а затем сбора клетокчерез день 3, или считая сотни клеток с помощью иммунофлуоресценции 4,5. Другая стратегия зависит от отбора на устойчивость к лекарственным препаратам для выделения коммутаторами 6. Поскольку африканские трипаносомы , выращенные в пробирке , как правило , состоят из большого населения , выражающего один крупный вариант, и гораздо меньшее население коммутаторами , выражающих альтернативные варианты, мы имеем в виду основной вариант в этой статье в качестве доминирующей, начиная VSG. При этом мы ни в коей мере хотим сказать, что этот крупный вариант имеет большую пригодность по сравнению с другими вариантами в популяции.
Здесь мы опишем метод, который может надежно оценить число трипаносом, выражающую недоминирующую VSG в данной популяции в 3 - 4 ч. Этот метод особенно полезен, когда кто-то хочет, чтобы установить, увеличивает ли данное генетическое манипулирование или медикаментозное лечение количество коммутируемых клеток в популяции. Вместо того, чтобы избавить население клеток, экспрессирующих Д.О.СМОРОДИНСКИЙ, начиная VSG через наркотики или иммунологическими средствами, эти клетки уничтожаются путем первого нанесения на них магнитными гранулами, соединенными с антителом против доминирующего VSG, а затем изолировать их на магнитной колонке. Коммутируемая население затем собирают в проточные и вновь окрашивали Флуорофор меченными анти-VSG антитела для выявления загрязняющих веществ. Количественное достигается путем добавления определенного числа абсолютных счетных бус для каждого образца так , что отношение бусинок к клеткам могут быть определены и использованы для количественного определения количества коммутаторами в популяции 7.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Примечание: В ходе технологического процесса, необходимо сохранить клетки на льду. Холодные носители также должны использоваться на всем протяжении.
1. Образец урожая
- Grow Lister 427 штамм трипаносом кровотока до плотности 0,5 - 1 миллион / мл. Лучше всего, чтобы начать культур с небольшим количеством паразитов. Спин вниз 50 х 10 6 клеток / образец в течение 10 мин при 1500 х г. Не забудьте оставить 1 × 10 6 клеток в культуре для последующего использования в качестве положительного и отрицательного контроля антител.
Примечание: Этот протокол также может быть использован на трипаносом, выделенных из крови животных (обсуждение см для более подробной информации). - Пипеткой или слейте большую часть супернатанта (оставьте около 750 мкл). Передача клетки 1,5 мл трубки микроцентрифужных.
- Спин клетки при 4 ° С в течение 4 мин при 5,200 XG в микроцентрифуге и удаления надосадочной жидкости.
2. Магнитный Этикетировочное
- Ресуспендируют клеток в 150 мкл культуральной среды (HMI-9 Wiй сыворотки, например) + первичное антитело против ВСГ в соответствующем разведении.
Примечание: анти-ВСГ антитело производится в доме и использовали в разведении 1:50. Важно использовать первичный анти-VSG антитело, которое не меченного флуорофором. - Вихревые клетки, используя вихревой адаптер-60 при скорости 6 - 8 в холодном помещении при температуре 4 ° С в течение 10 мин. Мытье с 800 - 1000 мкл холодного HMI-9 с сывороткой.
- Спин при 4 ° С в течение 4 мин при 5,200 XG и аспирата супернатант. Мытье с 1 мл HMI-9 с сывороткой и спином, как указано выше. Аспирируйте супернатант. Ресуспендируют осадок в 100 мкл HMI-9 с сывороткой.
- Добавьте 110 мкл магнитных активированных сортировки клеток (MSC) микрогранулы. С помощью анти-мышь, анти-кроличий или анти-биотин шарики в надлежащих случаях для первичного антитела анти-VSG. Хорошо перемешать. Вихревые клетки в холодном помещении при 4 ° С в течение 10 мин.
- В то время как клетки встряхиванием, установить одну магнитную активированные разделения колонок / образец на магнит разделения. Обязательно иметь повторноceptacle (мы используем центрифужную пробирку 15 мл) в колонке, чтобы собрать проточные. Настройка устройства в холодном помещении.
- Добавляют 2 мл HMI-9 с сывороткой к прайм колонке. После того, как заливка место 15 мл центрифужные пробирки под колонну для сбора проточный из каждого образца.
- После 10-минутной инкубации на стадии 2.4, добавьте 800-1000 мкл холодной HMI-9 с сывороткой. Отжима при температуре 4 ° С в течение 4 мин при 5,200 XG и удалить супернатант для удаления несвязанного антитела. Промыть 1 мл HMI-9 с сывороткой и повторите оставляя 100 мкл супернатанта.
- Флик центрифужную пробирку энергично и визуально проверить трубку, чтобы убедиться, что осадок полностью ресуспендировали. Добавьте 1 мл HMI-9 с сывороткой и пипеткой вверх и вниз, чтобы убедиться, что клетки хорошо ресуспендировали.
3. Магнитное разделение
- Убедитесь, что столбцы загрунтованы, как описано выше. Применить клетки к колонке. Сбор проточные с клетками, экспрессирующими недоминантную В.С.G. 1 мл среды + трипаносомы обычно занимает 6 - 7 мин, чтобы протекать через колонку.
- Мытье 2x 1 мл HMI-9 с сывороткой и собирать проточного в той же трубе, как в шаге 3.1.
- Разделить проточный (3 мл) в две 1,5 мл микропробирок.
Примечание: это могут быть объединены позже или половина может быть использована для выделения РНК и т.д. - Удалить столбец из магнита. Добавьте 3 мл HMI-9 с сывороткой в колонну и погрузить колонку в 15 мл центрифужную пробирку, чтобы получить клетки, которые выражают начальную, доминирующую VSG. Удалить 300 мкл элюированного материала для последующего анализа. Держите эти клетки на льду.
- Для использования в качестве положительного и отрицательного контроля, занимает приблизительно 1 миллион клеток из исходной культуры контрольных клеток, растущих при 37 ° C (те, как ожидается, не часто переключаться) и пипетку их в 1,5 мл трубки микроцентрифужных.
Примечание: положительные контрольные клетки будут окрашивали флуорофора меченных антител на этапе 4.1. Отрицательные контрольные клетки РЭМАв неокрашенными в остальной части протокола. - Спин проточные образцы и положительный контрольный образец антител при 4 ° С в течение 4 мин при 5,200 XG и удалить супернатант.
Примечание: Другой 300 мкл аликвоты элюированной клеток может быть использован в качестве положительного контроля для анти-VSG окрашивания.
4. Окрашивание выявлять загрязняющие вещества и коммутаторами
- Ресуспендируют проточные образцов и положительных контрольных клеток в 100 мкл HMI-9 с сывороткой + первичным, флуорофора меченных, анти-VSG-антитело в соответствующем разведении.
Примечание: меченными анти-ВСГ антител производится в доме и использовали в разведении 1: 1000. Флуорофор меченных анти-ВСГ может быть из того же источника, что и немеченых антител при разделении MACS step.There должно быть 3 мл проточного образца в двух микропробирок следующие стадии 3.5. - Для того, чтобы проанализировать все проточные с помощью проточной цитометрии, объединить окатышей в двух микропробирок при повторном Сусв ожидании образцов в 100 мкл HMI-9 с сывороткой + анти-VSG антителом. Хорошо перемешать.
- Вихревые клетки в холодном помещении при 4 ° С в течение 10 мин. Добавить 800 - 1000 мкл холодной HMI-9 с сывороткой. Образцы отжима при температуре 4 ° С в течение 4 мин при 5,200 XG и удалить супернатант для удаления несвязанного антитела.
- Вымойте клеток с 1 мл HMI-9 с сывороткой. Образцы отжима при температуре 4 ° С в течение 4 мин при 5,200 XG и удаляют супернатант. В течение этого последнего спина, спина вниз элюированный-образцы со стадии 3.4 и отрицательного контрольного образца.
- Ресуспендируют все проточные и элюированные образцы в: 148,5 мкл HMI-9 с сывороткой, 25 мкл абсолютного подсчета шарики и 1,5 мкл Пропидиум йодида окрашивание раствора (175 мкл общего объема). Ресуспендируют положительные и отрицательные контрольные образцы антитела в 175 мкл HMI-9 с сывороткой. Обязательно запишите число абсолютных подсчета шариков / 25 мкл для конкретной партии используется.
- Запуск всех образцов через поток цитометр и сбор данных. Мертвые клетки пятно, как PI
- Рассчитать частоты коммутации с использованием проточной цитометрии данных.
ПРИМЕЧАНИЕ: Для получения инструкций о том , как рассчитать частоты переключения, пожалуйста , обратитесь к дополнительному файлу потока цитометрии Calculations.docx и в таблице 1.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Описанный здесь метод был использован для демонстрации того, что двунитевых разрывов внутри сайта экспрессии VSG увеличилось количество коммутаторами в популяции 8. Здесь мы покажем, репрезентативные результаты из популяции трипаносом, которые были аналогичным образом индуцированными для генерации двухцепочечной разрыв на участке экспрессии. Мы сравниваем эти трипаносом к тем, которые не были индуцированная , чтобы генерировать двухцепочечной перерыв. На рисунке 1 показаны репрезентативные проточной цитометрии участков из не-индуцированных и индуцированных клеток , собранных в проточных от магнитной колонки сортировки активированных клеток. Для данного эксперимента доминирующим началом VSG был VSG2. Левую панели фиг.1А показывают вперед и боковое рассеивание, что позволяет ворота быть нарисовав вокруг абсолютных шариков подсчета и клеток , которые показывают прямой и боковой разброс подписи , что характерно для живых клеток. Обратите внимание, что ворота могутбыть обращено, чтобы включить больше клеток, потому что мертвые клетки могут быть устранены с PI окрашивания; Тем не менее, ворота должны быть одинаковыми для всех образцов. Правые панели фиг.1А показывают только те клетки , которые попадают в "Live" ворота на левой панели. Клетки, которые испачкать положительно для ПИ-регулятора (Q1 и Q2) являются мертвые клетки. Клетки, которые испачкать положительно для доминантной VSG и отрицательно для PI в Q3 являются загрязняющие вещества. Клетки, которые отрицательно испачкать как для ПИ и доминирующего VSG в 4-м квартале живые клетки, перешедшие. Можно заметить, что процент загрязняющих веществ в Q3 ниже для тех ячеек, где была индуцировали двухручьевой перерыв. Тем не менее, проценты в Q3 не могут быть использованы, чтобы сделать вывод, есть ли еще коммутаторами в данной популяции. Только путем сравнения отношения количества клеток в Q4 числу шариков , собранных , что можно вычислить тактовую частоту. В таблице 1 приведены номера , полученные от FloW цитометрии участков и метод расчета процентов коммутаторами в этих популяциях. Эти расчеты проводились на 3 -х биологических образцах , чтобы получить наблюдаемые частоты переключения , отображаемые на рисунке 1В. Уровень стохастического переключения в пробирке является довольно низким, как рассчитано здесь в среднем 9.53 × 10 -5, в то время индуцирования результаты перерыв в 1 - 6 × 10 -3 клеток, перешедших к недоминантной VSG.
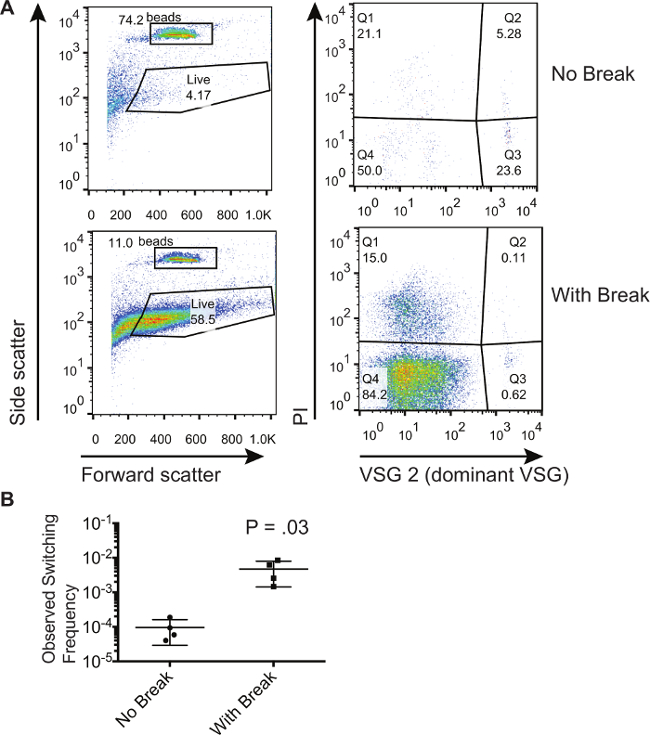
Рисунок 1. Выделение и Количественная коммутируемого трипаносом населения. (А) методом проточной цитометрии , показывающий фракции трипаносомы во фракции проточном после выбора не-коммутаторам на магнитной колонке сортировочного активированных клеток. Популяция клеток с индуцированной I-SCEI двухцепочечной перерыв сравнивается с популяции клеток без индуцированного BREак. (В) Рассчитано частоты коммутации для ячеек , которые имеют или не наведенных с I-SCEI двухцепочечной перерыва. Номер на графике представлены результаты двух хвостатых, непарный тест T. Усы представляют SD. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Справочная файл 1. Расчеты проточной цитометрии. Пожалуйста , нажмите здесь , чтобы загрузить этот файл.

Таблица 1. Расчет частоты переключения. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой таблицы.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Что касается экспериментальной техники, наиболее важный компонент протокола держит все образцы холодной. Трипаносомы очень быстро усваивают антитела , связанные с их поверхностью 9, но этот процесс зависит моторику, и не оказывает влияния на анализ тех пор , пока клетки выдерживали при 4 ° С. Все образцы должны всегда находиться на льду, и пипеткой должно быть сделано быстро, чтобы свести к минимуму воздействие на окружающую среду C лаборатории 25 °. Холодный ЧМИ-9 с сывороткой должны быть доступны в начале эксперимента и должны храниться на льду во всем. Выделение трипаносом, запуская их через колонку должно быть сделано в холодном помещении, так как она занимает 5 - 7 минут, чтобы запустить образец 1 мл через колонку. Если имеется больше выборок, чем единицы магнитной сепарации, доступных, магнитная изоляция может быть сделано последовательно, до тех пор, как образцы активно не отделенные держат на льду.
Также очень важно, что образцы быть очень хорошо перемешана афтер последней отмывки после инкубации с микрошариков и до изоляции на магнит. Это может быть достигнуто путем агрессивного стряхивая центрифужного трубки или легкого встряхивания. Ни одна гранула или сгустки клеток не должны быть видны перед добавлением образца в разделительную колонну, которую можно проверить, посмотрев на уровне света микроскопии. Если сгустки присутствуют, уровень загрязнения в проточном образце клеток, выражающая доминирующую VSG, как правило, гораздо выше, чем хотелось бы. При выполнении вычисления для числа коммутаторами в образце в конце протокола, важно принимать во внимание, был ли использован весь проточный для анализа методом проточной цитометрии, или использовали ли наполовину. Если только половина была использована, общее количество коммутаторами, вычисленных следует умножить на 2.
При удалении супернатант в течение стадий промывки, то нет необходимости отбирают пипеткой все до последней капли, и, как правило, 15 - 25 мкл являютсялевый окружающий осадок клеток. Это особенно важно во время завершающей стадии окрашивания и стирки, потому что в этот момент гранулы редко встречаются в проточных образцов, потому что есть так мало переключился клеток.
В то время как окончательное анти-ВСГ пятно, как правило, выполняется с использованием очищенной, флуорофора меченных анти-VSG антителом, первый анти-ВСГ пятно перед магнитной изоляции может быть сделано с очищенным антителом или сыворотки крови. Мы также использовали биореактор супернатант, полученный из гибридомы, экспрессирующих анти-VSG антител. Независимо от того, материал, используемый для исходного окрашивания необходимо титровать, чтобы обеспечить хорошее разделение элементов, или не экспрессируют доминирующий, начиная VSG.
Это характерно для популяции клеток в проточных быть загрязнена клеток, экспрессирующих доминирующую VSG. В то время как это загрязнение должно быть минимальным, в нашем опыте это редко для населения, чтобы быть полностью лишена клеток выразитьИНГ доминирующую VSG. Важно также отметить, что клетки, которые прошли через процедуры выделения не окрашивают в ярко с анти-VSG антитела, как те, которые не прошли через процедуру. Мы предполагаем, что две вещи, которые могли бы объяснить эту разницу. Во-первых, спиннинг трипаносом приводит к пролития VSG, и есть много вращений, участвующих в процедуре выделения. Это можно было бы ожидать, чтобы уменьшить интенсивность сигнала для окончательного анти-VSG пятно. Во-вторых, что первый шаг протокола предполагает окрашивание с первичным антителом против VSG. Если эпитопы, распознаваемые антителами, используемого на этой первой стадии такие же, как и закупоривать эпитопы, распознаваемые антителами в конечном красителем, можно было бы ожидать, что интенсивность сигнала будет ниже, чем при использовании только одно антитело. По этой причине важно, чтобы титровать антитело, которое используется на конечной стадии, таким образом, что сигнал от положительно окрашенной клетки примерно на двапорядков выше, чем для отрицательно окрашивали клетки. Таким образом, даже если интенсивность сигнала от положительно окрашивания клетки уменьшается в клетках, которые подверглись процедуре изоляции, эти положительные клетки будут по-прежнему хорошо отделены друг от отрицательных клеток на участке проточной цитометрии. Если функция автоматической компенсации используется, одиночные контрольные образцы пятна должны быть подготовлены. Антитела, которые мы сгенерировали доступны для покупки из Memorial Sloan Онкологический центр monocolonal объекта антитела. Мы имели лучший успех с использованием немеченного моноклональных антител IgG. Мы провели протокол с поликлональными антителами VSG, но поликлональные антитела могут иногда распознавать другие VSGs являются предпочтительными, так моноклональные антитела. ВСГ идентичность может быть определена путем секвенирования VSG кДНК, полученных при амплификации законсервированных последовательностей в VSG 3'-UTR и сращенных лидера. Особым антитела могут быть протестированы на специфичность по ранее выделенным переключилсяКлетки, которые VSG идентичность'S была определена, как описано выше.
Протокол, представленные здесь, имеет ряд преимуществ. Он может быть использован как для выделения коммутаторами для последующего анализа и количественной оценки количества коммутаторами в данной популяции. Он также может быть использован для выделения конкретного варианта интерес, до тех пор, как антитело доступно для данного варианта. Кроме того, можно осуществить этот протокол, используя трипаносом, выделенные из животных, при условии, что эритроциты устраняются с использованием анти-Ter119 покрытием-магнитных гранул в соответствии с протоколом производителя. Мы не тестировали нижний предел для количества трипаносом необходимых, но мы успешно выполнили протокол, используя 2,5 - 10 миллионов трипаносом из 250 мкл крови. И, наконец, протокол может быть использован в сочетании с анализом колебаний, чтобы получить частоту переключения в данной популяции.
Процедура изоляции вполне еAST, принимая 3 - 4 ч от начала до конца в зависимости от количества образцов, и, таким образом, является более эффективным, чем предыдущие методы, которые требуют предварительного иммунизации мышей или специализированных штаммов, содержащих маркеры лекарственной устойчивости. Метод ограничена тем фактом, что антитела против исходного VSG требуются, однако. Без таких реагентов, альтернативный способ , такой как VSG-SEQ может быть более подходящим выбором для оценки которого VSGs высказываются в данной популяции 10.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Мы хотели бы иметь в виду Георгиевский крест общих рекомендаций по трипаносом биологии. Эта работа также была поддержана грантом GCE Билла и Мелинды Гейтс, DS в НФС Graduate Fellowship (Research DGE-1325261) для управления записями и NIH / NIAID (грант № AI085973) в ФНП. Мы благодарим Галадриэль Hövel-Miner для использования штамма, содержащего ген и распознавания сайта I-SCEI.
Materials
| Name | Company | Catalog Number | Comments |
| unlabeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| fluorophore labeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| HMI-9 media | n/a | n/a | HMI-9 is made in house |
| Propidium Iodide | BD Pharmingen | 556463 | |
| CountBright absolute counting beads | Thermofisher Scientific | C36950 | |
| LD columns | Miltenyi Biotech | 130-042-901 | |
| MACS magnet | Miltenyi Biotech | 130-042-303 | |
| MACS magnetic separator | Miltenyi Biotech | 130-042-302 | |
| vortex adapter-60 | ThermoFisher scientific | AM10014 | |
| flow cytometer | Coulter | n/a | |
| flow cytometer analysis software | FloJo | n/a |
References
- Horn, D. Antigenic variation in African trypanosomes. Molecular & Biochemical Parasitology. 195 (2), 123-129 (2014).
- Lamont, G. S., Tucker, R. S., Cross, G. A. Analysis of antigen switching rates in Trypanosoma brucei. Parasitology. 92 (Pt 2), 355-367 (1986).
- Rudenko, G., McCulloch, R., Dirks-Mulder, A., Borst, P. Telomere exchange can be an important mechanism of variant surface glycoprotein gene switching in Trypanosoma brucei. Mol Biochem Parasitol. 80 (1), 65-75 (1996).
- Turner, C. M., Barry, J. D. High frequency of antigenic variation in Trypanosoma brucei rhodesiense infections. Parasitology. 99 (Pt 1), 67-75 (1989).
- Miller, E. N., Turner, M. J. Analysis of antigenic types appearing in first relapse populations of clones of Trypanosoma brucei. Parasitology. 82 (1), 63-80 (1981).
- Horn, D., Cross, G. A. Analysis of Trypanosoma brucei vsg expression site switching in vitro. Mol Biochem Parasitol. 84 (2), 189-201 (1997).
- Figueiredo, L. M., Janzen, C. J., Cross, G. A. M. A histone methyltransferase modulates antigenic variation in African trypanosomes. PLoS Biol. 6 (7), e161 (2008).
- Boothroyd, C. E., Dreesen, O., et al. A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature. 459 (7244), 278-281 (2009).
- Engstler, M., Pfohl, T., et al. Hydrodynamic Flow-Mediated Protein Sorting on the Cell Surface of Trypanosomes. Cell. 131 (3), 505-515 (2007).
- Mugnier, M. R., Cross, G. A. M., Papavasiliou, F. N. The in vivo dynamics of antigenic variation in Trypanosoma brucei. Science. 347 (6229), 1470-1473 (2015).
