

Summary
Dette manuskriptet beskriver i vitro video mikroskopi protokoller for evaluering av vaskulær funksjon i rotte cerebral motstand arterier. Manuskriptet beskriver også teknikker for å vurdere microvessel tetthet fluorescently merket Lektiner og vev perfusjon bruker Laser Doppler Flowmetry.
Abstract
Denne protokollen beskriver bruken av i vitro TV mikroskopi evaluere vaskulær funksjon i isolerte cerebral motstand arteriene (og andre fartøy), og beskriver teknikker for å vurdere vevsperfusjon bruker Laser Doppler Flowmetry (LDF ) og microvessel tetthet benytte fluorescently merket Griffonia simplicifolia (GS1) Lektiner. Gjeldende metoder for å studere isolert motstand arterier på transmuralt press oppdaget i vivo og i fravær av parenchymal celle påvirkninger gir en viktig kobling mellom i vivo studier og informasjon fått fra molekylær reduksjonistiske tilnærminger som gir begrenset innsikt i integrerende svar på hele dyr nivå. LDF og teknikker for å identifisere arterioler og kapillærer med fluorescently-merket GS1 Lektiner gir praktiske løsninger for å aktivere etterforskerne å utvide kunnskapen fra studier av isolerte motstand arterier. Dette dokumentet beskriver bruk av disse teknikkene for å få grunnleggende kunnskap om vaskulær fysiologi og patologi i rotte som generell eksperimentell modell, og i en rekke spesialiserte genmodifiserte "designer" rotte stammer som kan gi viktig innsikt påvirker bestemte gener viktig vaskulær fenotyper. Utnytte disse verdifulle eksperimentelle tilnærminger i rotte stammer utviklet av selektiv formering strategier og ny teknologi for å produsere genet knockout modeller i rotte, utvide rigor av vitenskapelige lokale utviklet i knockout musen modeller og Utvid kunnskapen til en mer relevant dyr modell, med en godt forstått fysiologiske bakgrunn og egnethet for fysiologiske studier på grunn av sin større.
Introduction
De tidligste studiene av vaskulær funksjon i arteriene utnyttet kanal arterier og i mange tilfeller aorta. Styrkegenerering i store arterier var generelt studert ved å feste en ring segment av arterien til en force svinger i en vev bath. ved aorta, striper ved å kutte spiralformede av fartøyet slik at glatt muskel fibrene var orientert i lengderetningen mellom punktet for feste og kraft svingeren, å gi anslag for kraft generert av sammentrekning av glatt muskel langs den langsgående aksen. Standard teknikken for å kutte spiralformede strimler av aortas var å plassere glass stang i lumen av fartøyet, gjøre et kutt i fartøyet veggen ønsket og holde på slutten av den synlige kanten av fartøyet veggen som kuttet ble utvidet til å produsere en hel spiralformede stripe av fartøyet. På dette punktet, endothelial siden av fartøyet var generelt uten for å fjerne rusk før fartøyet stripen vedlegges force svingeren og submerging forberedelse i et oksygenrikt og temperatur kontrollert vev bad. Til slutt, avledet at tilnærming førte til en av de mest berømte og viktige funnene i historien til fysiologi av Furchgott og Zawadski1, nemlig rollen endotelet avslappende faktor (EDRF), senere identifisert som nitrogenoksid, i regulere vaskulær funksjon. Det avgjørende fører til at funn var en situasjon der etterforskerne opprettholdt en intakt endotelet ved å unngå kontakt med endotelial siden av arteria med utenlandske flater, og la merke til at aorta stripen ikke ha forventet sammentrekning til acetylkolin (ACh), men i stedet avslappet svar ACh. Basert på den observasjonen, etterforskerne utviklet en "sandwich" forberedelse de knyttet en aorta segmentet med en intakt endotelet (men ikke generere contractile kraften) til en standard spiralformede stripe av aorta og konvertert ACh-indusert sammentrekning i en avslapning.
To store fremskritt i dette området som brukes mye i dag er utviklingen av forberedelser for å måle aktiv contractile kraften i liten motstand arterier2,3 (som de i intestinal mesentery3 ) og cannulated motstand arterien forberedelser4,5,6. En av de tidligste rapportene, Mulvany og Halpern3 beskrevet bruk av wire myograph utarbeidelse å studere aktive contractile kraften i isolerte motstand arterier fra intestinal mesentery spontant Hypertensiv rotter (SHR) og normotensive WKY kontroller. Etter utviklingen av wire myograph systemet, ble cannulated motstand arterien forberedelsene utviklet for å tillate studier av fartøy nærmere i vivo forhold4,5,6. Begge metodene gir verdifulle resultater, har cannulated arterien utarbeidelse ekstra fordeler mer effektivt bevare iboende aktive tonen i arteriene; og slik at etterforskerne å studere aktive myogenic svar på endringer i transmuralt press og fartøy svar til endringer i flyt og endothelial skjæring stress (se Anmelde av Halpern og Kelley6).
Et hovedmål for denne utredningen er å beskrive hvordan du bruker hevdvunne teknikken av video mikroskopi bruke isolerte, cannulated motstand arterier for å få nøyaktig informasjon om mekanismer som regulerer aktive tonen i disse avgjørende fartøy, uavhengig av nevrale, humoral eller parenchymal celle påvirkninger. Denne grunnleggende informasjon, ansette en standard rotte modell og eksempler fra våre studier av nye genetisk konstruert rotte stammer, gir leseren et inntrykk av hvilke typer innsikt om vaskulær funksjon som kan oppnås med TV mikroskopi nærmer seg, og som kan brukes i studier som involverer noen kontroll og eksperimentelle gruppe(r) av etterforskerens valg, inkludert kraftige nye eksperimentelle rotte modeller produsert av selektiv inbreeding og nyutviklet genetisk engineering teknikker.
Takk til presisjon av TV mikroskopi tilnærminger gir måling av diameter endringer i cannulated arterien forberedelser svært verdifull informasjon om endotelet avhengige av og endotelet-uavhengig av vaskulære avslapning, samt viktige (og noen ganger uventet) endringer i vaskulær kontrollmekanismer oppstår med hypertensjon, høy salt diett og andre eksperimentelle tiltak. I tillegg måling av trykk-diameter relasjoner i isolert og cannulated motstand arteries som er maksimalt avslappet av behandling med Ca2 +-gratis løsning eller en farmakologisk vasodilator stoffet, kan investigator å vurdere strukturelle endringer i arteriene vaskulær remodeling og beregne passiv stress-belastning relasjoner7 som kan gi viktig innsikt i endringer i egenskapene for passiv mekanisk av arteriene som kan påvirke arteriell funksjon uavhengig av (eller i tillegg til) endringer i aktiv kontrollmekanismer. Det er også viktig å merke seg at informasjon fra studier av isolerte motstand arterier kan suppleres av anskaffet utnytte LDF, en praktisk metode for å vurdere vevsperfusjon i hele dyr nivå8,9 ,10, og av informasjon fra vurdere microvessel tetthet med fluorescently merket GS1 Lektiner, som spesifikt bindes til glykoprotein moieties i kjelleren membranen av små arterioler og kapillærer11 , 12. sistnevnte gir en svært nøyaktig anslag av microvessel som ikke er avhengig av klassiske vanskelighetene i estimering microvessel tetthet ved å telle fartøy i vivo, for eksempel manglende ikke-perfused fartøy der blodstrøm er stoppet på grunn av aktive nedleggelsen av arterioler. Når de brukes sammen, kan disse metodene gi viktig innsikt for å sammenligne funksjonelle endringer i isolerte motstand arteries til endringer i vevsperfusjon på microcirculatory nivå; og noen eksempler på bruk av de verdifulle nærmer seg sammen med cannulated arterien teknikker vil også bli gitt i stede manuskriptet.
Utredningen fokuserer på bruk av video mikroskopi teknikker å evaluere vaskulære endringene i arteriene outbred Sprague-Dawley rotter. Det er imidlertid viktig å merke seg at disse teknikkene har vist seg for å være svært verdifull i Klargjørende fenotypiske endringer i høyt spesialiserte genmodifiserte rotte stammer opprettet av selektiv formering eller genet redigering med teknikker. I dette manuskriptet gi vi eksempler på hvordan mikroskopi teknikker har gitt viktig informasjon vedrørende vaskulær funksjon i en rekke verdifulle rotte modeller, inkludert Dahl salt-følsomme (SS) rat-en innavlet rotte belastningen som er den mest utbredte brukt eksperimentell modell for å studere mekanismer for salt sensitive hypertenson18,19,20,21,22,23; og consomic rotter opprettet via selektiv formering SS rotter med salt-ufølsom Brown Norge (BN) rotte belastningen. Consomic rotten panelene vært hver kromosom fra Brown Norge rotta introgressed individuelt i Dahl SS24,25,26 genetisk bakgrunnen. Bruk av consomic rotten paneler har gitt verdifulle ledetråder om bestemte kromosomene som bidrar til salt følsomhet for blodtrykk og andre fenotyper, inkludert vaskulær reaktivitet24,25,26 ,27,28.
Selektiv formering forsvarsstrategier utnytte SS rotter og consomic rotter bærer individuelle BN kromosomene har aktivert generasjonen av avgrensede congenic stammer med små segmenter av personlige Brown Norge kromosomene introgressed i Dahl SS genetisk bakgrunn22,29. Dette kan gi svært verdifulle innspill til bestemte gener eller begrense regioner av kromosomer som kan påvirke avgjørende fysiologiske variabler, som blodtrykk, nyre skader og vaskulær reaktivitet22,29. Et kraftig tillegg til rotte genetisk verktøykassen er utviklingen av rotte genet knockout modeller bruker avansert genet redigering teknikker som ZFNs, transcriptional aktivator-lignende-effektor nucleases (TALENS), og nylig CRISPR-Cas913 ,14,15,16,17. Ankomsten av disse kraftige teknikker som gener å bli slått i rotte er en uhyre viktig utvikling fordi genet knockout studier hittil har brukt (og fortsette å bruke) mus nesten utelukkende. En annen eksperimentell komponent i dagens papir viser verdien av cannulated arterien teknikker og video mikroskopi evaluere fysiologiske kontrollmekanismer i knockout rotter mangler master antioksidant og celle beskyttende transkripsjon faktor, kjernefysiske faktor (erythroid-avledet 2) - som - 2 (NRF2)30,31, som ble utviklet ved hjelp av TALEN teknologi i Sprague-Dawley genetisk bakgrunn17. I disse eksperimentene, ble i vitro video mikroskopi teknikker brukt til å gi funksjonelle verifisering av tap av NRF2 genet og teste en potensielt verdifulle terapeutisk tilnærming basert på direkte oppregulering av NRF2-mediert antioksidant forsvar. NRF-2 er betydelig terapeutiske betydning i kampen mot vaskulær oksidativt stress hos mennesker, i lys av skuffende resultatene av kliniske forsøk med direkte administrasjon av antioksidanter som vitamin C og E32.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Medical College i Wisconsin institusjonelle Animal Care og bruk Committee (IACUC) godkjent alle protokoller beskrevet i dette dokumentet og alle prosedyrer er i samsvar med National Institutes of Health (NIH) Office for laboratoriet dyr velferd (OLAW) forskrifter.
1. forberedelse av løsninger og fartøy kammer
- Før gjennomfører en rekke eksperimenter, forberede 2 L 20 x konsentrert salt lagerløsning består av 278 finans NaCl; 14 finans KCl; 11.52 finans MgSO4. 7T2O; og 9,4 finans CaCl2. 2H2O. Også forberede 2 L 20 x konsentrert reservelager 80.8 finans NaHCO3 og 0,4 finans EDTA, og 2 L 20 x konsentrert Ca2 +-gratis lager løsning med 281.6 finans NaCl; 14 finans KCl og 11.52 finans MgSO4. 7T2O.
Merk: 20 X lager løsninger kan lagres i kjøleskapet før bruk. - På dagen for eksperimentet, forberede 2 L fysiologisk saltløsning (PSS) fra 20 x konsentrerte lager løsninger som følger: legge til 100 mL 20 x salt lager 1800 mL deionisert vann i 2 L Erlenmeyer kolbe eller kanne på en motorisert gripende plate. Legge til 100 mL 20 x reservelager mens kontinuerlig equilibrating løsningen med en gassblanding som inneholder 21% O2, 5% CO2, saldo N2og rører med magnetiske gripende bar. Sakte legge 0,28 g NaH2PO4 mens overvåking pH; Juster etter behov til pH 7.4 ved å legge dråper 6 N HCl eller 6.5 N NaOH løsning fra en Pasteur pipette. Når PSS er forberedt og pH justeres, Legg 1,98 g av glukose til PSS.
Merk: Det er viktig å sakte legge NaH2PO4 siste mens overvåking pH i PSS fordi tillegg av NaH2PO4 til en alkalisk (pH > 7.4) kunne danne en kalsium fosfat bunnfall, som angitt av den utseendet til en skyet løsning eller en hvit bunnfall i bunnen av beholderen.
Merk: Den endelige sammensetningen av PSS er 119 mM/L NaCl; 4.7 mM/L KCl; 1,17 mM/L Mg2så4; 1,6 mM/L CaCl2; 1,18 mM/L NaH2PO4; 24 mM/L NaHCO3; 0,03 mM/L EDTA; og 5,5 finans glukose. Selv om sammensetningen av PSS kan variere mellom laboratorier, er denne oppskriften velegnet for å opprettholde vaskulær tone, endothelial funksjon og Svar å vasoactive agenter i isolerte motstand arterier. - Bestemme maksimale diameter og vurdere aktive tonen i arterien ved å produsere maksimal utvidelse av fartøyet, forberede 500 mL Ca2 +-gratis PSS ved å legge til 25 mL av den 20 X Ca2 +-gratis salt lager til 450 mL deionisert vann , etterfulgt av 25 mL av 20 x reservelager i Erlenmeyer kolbe eller kanne lignende gå 1.2 over. Legge til 0,07 g NaH2PO4 løsningen mens overvåke og justere pH av løsningen. Ca2 +-gratis PSS legges til PSS reservoaret og fartøy chamber på slutten av å unngå tappe intracellulær Ca2 + butikker som kan påvirke fartøyet svar i normal PSS. Fordi Ca2 +-gratis løsning er lagt til på slutten av å produsere maksimal avslapning av arteries, det er ikke nødvendig å legge til glukose i PSS.
Merk: Når fullfører, den endelige sammensetningen av Ca2 +-gratis PSS er 120.6 mM/L NaCl; 4.7 mM/L KCl; 1,17 mM/L Mg2så4; 1,18 mM/L NaH2PO4; 24 mM/L NaHCO3; 0 mM/L CaCl2; og 0,03 mM/L EDTA.
Merk: For mange studier som krever maksimal utvidelse av arterien, en kalsium oppføring blokker som verapamil (1 µM) og/eller nitrogenoksid giver som natrium nitroprusside (10 µM) kan legges til løsningen, i tillegg til fjerne Ca2 + fra PSS. - Opprettholde den PO2, PCO2og pH av PSS ved kontinuerlig equilibrating PSS strømmer inn i fartøyet kammeret i en standard orgel bad brukes til å studere isolert aorta ringer, intestinal glatt muskel eller andre vev (figur 1). Bruke syntetisk fluoropolymer tetrafluoroethylene rør koble gasstanken til orgel bad fordi denne slangen er gass ugjennomtrengelige, i motsetning til mange andre former for rør, f.eks, latex.
- Plass en lite luft stein koblet til balanse gass blandingen i fartøyet kammeret for å opprettholde PSS gass sammensetningen.
Merk: Fartøyet svar til endringer i PO2 kan testes ved equilibrating PSS i fartøyet kammer og luminal perfusate med gass blandinger som inneholder ulike prosenter av O2, f.eks, 21% O2, 10% O2, 5% O2 , og 0% O2, med 5% CO2 og balanse N233,34,35. For større arterier med tykke vegger, hvor spredningen av oksygen i sentrum av fartøyet veggen kan være en begrensning, brukes en høyere andel av oksygen, f.eks, 95% O2 . - Overvåke temperaturen i fartøyet kammeret tett, mens enkelte kammer kan variere i deres varme overføring egenskaper.
Merk: Mange kommersielt forberedt fartøyet kammer brukes for studier av cannulated motstand arterier utnytte en peristaltiske pumpen å levere oksygenrikt PSS fra et gass-equilibrated reservoar og gir svært nøyaktig kontroll over badevann og oksygenering av PSS. - Plass PSS i en stor (2 L) Mariotte flaske, stopper og sentrale røret skal fungere som et reservoar kontinuerlig levere PSS i orgel badekaret som varmer og gass-equilibrates PSS strømmer inn i fartøyet kammeret (figur 1A).
- Stedet åpningen av det sentrale røret i Mariotte flaske på samme nivå som toppen av PSS i orgel badekaret, for å opprettholde en konstant hydrostatisk trykk leder for levering av PSS i orgel badekaret. Bruk polyetylen rør koblet til en J-formet glass eller plast røret å levere PSS fra Mariotte flasken i orgel badekaret.
- Luminal perfusjon (figur 1A), bruke polyetylen rør koble tilsig Pipetter til et PSS reservoar består av en 60 cc plastsprøyte opphøyet til en posisjon som opprettholder ønsket tilsig trykket (vanligvis 80 mmHg for studier av rotte cerebral arteriene), målt med et trykk svinger koblet til systemet via en stopcock.
- Koble strøm pipette slik polyetylen tillate PSS å strømme gjennom fartøyet som svar på en trykkgradient og koble ut linjen til et reservoar ligner tilsig reservoaret. Bruke en lignende stopcock og press svinger tilkobling for å måle strøm press.
Merk: Prosedyrene for å sette transmuralt press og kontrollere flyt gjennom fartøyet er beskrevet i § 2, nedenfor. - På slutten av eksperimentet, rense kammer, levering linjene og reservoaret systemer med destillert vann. Jevnlig, erstatte rør og levering linjene, ren eller erstatte stopcocks i systemet og regelmessig underlagt noen glass PSS reservoarer en syre vask å hindre vekst av bakterier og andre mikroorganismer som forårsaker forurensning og påvirke fartøyet reaktivitet.
2. cannulated arterien forberedelse
- Bedøve rotte Sprague-Dawley med 5% isoflurane og vedlikehold av bedøvelsen bruke 1.5-2.5% medisinsk karakter oksygen36. Du kan også administrere intramuskulær injeksjon inneholder ketamin (75.0 mg/kg), acepromazine (2,5 mg/kg) og xylazine (10,0 mg/kg); en intraperitoneal injeksjon av pentobarbital (50-60 mg/kg); eller en annen godkjent metode for anestesi, avhengig av protokoller og/eller etterforsker innstillinger.
- Halshugge rotta under dyp anestesi og fjerne hjernen for studier av cerebral motstand arterier.
- Etter fjerning av hjernen, forsiktig isolere arterien cerebri (MCA) (eller andre arterier severdigheter, f.eks, basilar arterien eller bakre arteria cerebri)37,38. Å isolere MCAs, plasserer hjernen ryggen i et glass Petriskål fylt med iskald PSS (figur 2).
- Bruk Vannas saks og en Dumont #5 fin spiss tang til å avgiftsdirektoratet MCA fra hjernen. Rengjør alle gjenværende hjernevevet fra MCA ved hjelp av tang og overføre arterien til en temperatur kontrollert fartøyet kammer som inneholder PSS som beskrevet tidligere. 33 , 34 .
- Slik overfører arterien til fartøyet kammeret, hold forbrukeravgift fartøyet av ACA eller bakre kommuniserende arterien segmentet og forsiktig plassere den i kammeret.
Merk: I tillegg til MCA, cannulated fartøy systemet er egnet for en rekke små fartøy preparater, inkludert skjelettlidelser muskel motstand arterier33,39,40, hvem motstand arterier 38 , 41 , 42og stor (første ordre) arterioler av den cremaster muskel43, samt menneskelige koronar arterioler og menneskelige arterioler fikk fra subkutan fettvev under gluteal biopsi44,45, 46,47. - Fest arterien til tilsig brønnene ved å trekke det mot pipette basen til spissen fremskritt innenfor lumen av MCA. Sikre arterien på tilsig pipette ved å knytte en sløyfe utarbeidet av en enkelt strand fiber tidligere ertet fra 10-0 suturer rundt arteria (figur 1B). Sikre den motsatte enden av MCA til utstrømming pipette ved å stramme en andre Sutur sløyfe rundt fartøyet (figur 1B).
Merk: Sutur løkkene på Mikropipetter før montering fartøyet og plassere dem inntil det siste punktet for vedlegg, slik at de kan være lett gled over arterien og fort sikret når fartøyet er i posisjon, som minimerer risikoen for den arterie glir av pipetter.
Merk: Mikropipetter tilberedes fra Borosilikatglass kapillær rør (2 mm ytre diameter, 1 mm indre diameter, 10 cm lang) med en loddrett brønnene avtrekker. Før knytte arterien, samsvarer med tips diameter på Mikropipetter så langt som mulig å forhindre uoverensstemmelser tilsig og utløp motstand i perfusjon systemet. - Når arterien er trygt bundet til Mikropipetter, bruke mikrometer koblet til tilsig pipette innehaveren for å strekke arterien til i situ lengden.
- Tie av alle sidegreiner med singelen tråder ertet fra 10-0 suturer for å opprettholde et konstant trykk i arterien.
- Kontroller at lekkasjer ved å sørge for at intraluminal (transmuralt) trykket forblir konstant etter midlertidig stenge tilsig pipette. Tie av grenene eller sjekk for hull i fartøyet Hvis trykket faller. Gjenopprette perfusjon etter bekrefter at transmuralt trykket forblir konstant.
Merk: Cannulation av isolerte motstand arterier krever fingerferdighet og praksis. De viktigste forholdsreglene følges er å unngå å bryte Pipetter og sørge for at arteria ikke skyver på pipetter. Det er viktig å være forsiktig med isolert arteriene gjennom hele prosedyren som traume til fartøyet kan skade endotelet og/eller forstyrrer normal funksjon av vaskulær glatt muskel. - Måle interne diameter av arterien bruker en video mikroskopi oppsett (figur 1A) som består av et videokamera festet til dissecting mikroskop og koblet til en video mikrometer og TV-skjerm (tall 1B, 1 C). Dette gjør observatøren måle fartøyet diameter manuelt ved å plassere bevegelige Referanselinjer på indre veggen av arterien og, hvis ønskelig, på ytterveggen av arterien, for å måle fartøyet veggtykkelse.
Merk: Noen video mikrometer tilbyr automatisk sporing av fartøyet dimensjoner.
Merk: Kalibrere det video mikrometer med et mikroskop scenen mikrometer og tilsig og utløp press transdusere med en kvikksølv manometer (0 mmHg, 50 mmHg, 100 mmHg, 150 mmHg og 200 mmHg) mellom eksperimenter for å sikre nøyaktig måling av fartøyet diameter og intraluminal press.
Merk: Standardkontrollen transmuralt trykket for rotte MCA eksperimenter er 80 mmHg. Kalibrering for høyere og lavere nivåer sikrer nøyaktigheten for studier av myogenic svar på endringer i transmuralt trykk og passiv press-diameter kurver i maksimalt utvidede blodårer. - Juste høyden på tilsig og utløp reservoarene å opprettholde ønsket transmuralt trykket på et konstant nivå. Øke tilsig reservoaret et lite (< 5 mmHg) og senke utløp reservoaret like mye opprettholder mener transmuralt trykket og skaper perfusjon flyt i fartøyet lumen6.
Merk: Øke tilsig reservoaret og senke utløp reservoaret ved like mengder opprettholder den samme mener transmuralt distending trykket i arterien, men genererer hydrostatisk trykk gradering som forårsaker flyt og skjæring stress i fartøyet, slik at den etterforsker evaluere endotelet avhengige av svar til endringer i intraluminal skjæring stress i ulike eksperimentelle grupper48. - For å vurdere myogenic svar og fartøy svar på vasodilator stimuli, kontroller at arteria viser et passende nivå av aktive tone (ca 40%) før eksperimentet. Forkaste alle arterier mangler aktive tone på resten unntatt fartøy, f.eks, liten hvem arteriene som ikke vanligvis forårsaker aktiv hvile tone.
Merk: For skip som ikke vanligvis forårsaker spontan tone, må du pre constrict arteriene av tilsvarende beløp utnytte en vasoconstrictor Agonistiske som noradrenalin eller phenylephrine. En god tilnærming til å velge dosen av Agonistiske brukes til pre constrict arterien er å benytte en EF50 dose av vasoconstrictor agent, f.eksnoradrenalin41,42. Imidlertid bør farmakologiske vasoconstrictor agenter ikke brukes til arteriene som viser spontan aktive tone under hvile forhold. - Teste endotelet avhengige av reaktivitet av arteria cannulated motstand ved å måle fartøyet diameter lagt økende konsentrasjoner av ACh (10-10 M-10-5 M) til fartøyet kammeret. Teste endotelet-uavhengig nitrogenoksid følsomhet av vaskulær glatt muskel ved å måle fartøyet diameter lagt økende konsentrasjoner (10-10 M-10-5 M) av nitrogenoksid donor natrium nitroprusside til den fartøyet kammeret.
Merk: Følsomhet for vasoconstrictor agenter og andre vasodilator agonister kan testes på en lignende måte ved å legge til økende konsentrasjoner av Agonistiske fartøyet kammeret.
Merk: I tillegg til vasoactive agenter, ulike stoffer, hemmere og andre farmakologiske agenter kan legges til vev badekaret og/eller luminal perfusate. Endringer i PO2 (samt PCO2 og pH) kan selektivt administreres endothelial siden eller ekstra-luminal siden av arterien av separate balanse av den luminal perfusate med en luften sten koblet til en kalibrert gassblanding forskjellig fra blandingen til equilibrate PSS i vev bad33,34,49. Myogenic svar på endringer i transmuralt press kan studeres ved å lukke ut pipette og øke (eller senke) høyden av PSS reservoaret koblet til tilsig pipette49,50,51 til øke eller redusere intraluminal trykk. - For å teste rollen endotelet i formidling av fartøyet svar på bestemte stimuli, Fjern vaskulære endotelet og sammenligne fartøyet svar på disse stimuli i tilstedeværelse eller fravær av endotelet. For å fjerne endotelet, nøye løse arteria fra utløp pipette og langsomt perfuse lumen av arterien med en luft bolus (0,5-1.0 mL). Etter luft perfusjon, kan du gjenopprette PSS perfusjon Fjern mobilnettet rusk før re binde fartøyet til utstrømming pipette43.
Merk: Etter endotelial denudation prosedyren er det viktig å kontrollere at endotelet fjernes ved testing vaskulær Svar å en Agonistiske (vanligvis ACh) kjent for å produsere endotelet avhengige vasodilatasjon i at beholdertype. Men i enkelte patologisk slippes endotelet avhengige av vasoconstrictors, da, er det viktig å kontrollere at constrictor svaret er eliminert etter endothelial. - På slutten av eksperimentet, bestemmer den maksimale diameteren Arterien ved å legge til Ca2 +-gratis PSS til perfusate og superfusate. Beregne aktiv hvile tone (%) som ((DMaks-Dresten) /Dmax) x 100, der Dmax er maksimale diameter i nærvær av Ca2 +-gratis løsning og Dresten er hvile kontroll diameter.
Merk: Diameter målinger av maksimalt avslappet arterier i forskjellige eksperimentelle grupper er verdifull i å sammenligne aktiv hvile tone, strukturelle remodeling (dvs.endringer i veggen tykkelse og lumen diameter) og passiv mekaniske egenskaper (stress-belastning relasjoner beregnet fra passiv diameter på ulike nivåer av transmuralt press).
3. evaluering av hjerne blod flyte svar med LDF
- Bedøve dyret med 5% isoflurane og sikre rotta inne en stereotaxic apparat9,10.
- Opprettholde dyret under konstant anestesi mens overvåking puste frekvens, slutten tidevanns CO2og dybden av anestesi med en tå knipe36.
- Nøye tynne skallen til gjennomskinnelighet bruker en lav hastighet tannlege bore og mineralolje for å gi optisk kopling10. Forsiktig å unngå å generere sterk varme og unngå gjennomtrengende benet.
Merk: Fortynning av skallen lar laserlys å nå underliggende vev og reflekteres tilbake til proben for å måle Doppler skiftet, omfanget av som bestemmes av hvor mange bevegelige partikler (dvs., røde blodlegemer) og deres hastighet. - Sikre LDF sonden i en micromanipulator og plasser den rett over tynnet området av skallen. Under eksperimentet er det svært viktig å hindre bevegelse LDF sonden eller forberedelse, LDF er utformet for å måle flyt i et begrenset område av vev og er svært følsom for bevegelse.
Merk: Enhver bevegelse av sonden fra sin opprinnelige posisjon vil gi et estimat av blodstrøm i et annet område i vevet, slik at sammenligninger. Mens LDF gir ikke absolutt strømnings verdiene og er ikke egnet for sammenligning mellom fag, er det en utmerket måte å noninvasively vurdere endringer i vevsperfusjon svar på eksperimentell intervensjoner i enkelte fag; og relativ endringer i LDF signalet fra kontrollverdier kan gjennomsnitt og forhold til endringer i LDF signalet fra kontrollen i andre eksperimentelle grupper.
Merk: LDF er en praktisk tilnærming til innblikk i faktorer regulerer blodstrømmen nivået av hele vaskulær sengen i ulike eksperimentelle grupper8,9,10. Evaluering av vevsperfusjon med LDF gir en praktisk tilnærming for å assimilere kunnskapen fra isolert fartøyet studier i en hel seng perspektiv. Sperring regionale forskjeller i vaskulær kontrollmekanismer mellom motstand arterier og blodsirkulasjonen gir målinger med LDF en god indikasjon på vev blod flytkontroll som vanligvis samsvarer med resultatene med cannulated arterie forberedelser.
4. evaluering av skjelettlidelser muskel Microvessel tetthet med GS1 Lektiner
- Fjern cremaster muskelen fra en mannlig rotten ved kutte åpne pungen med standard fine kirurgisk saks og deretter bruke en Dumont #5 tang for å forstå muskelen.
Merk: Tynn muskler (f.eks, cremaster muskelen, samt extensor digitorum longus og tibialis fremre musklene som finnes i både mannlig og kvinner rotter) er ideell for bruk som helhet mounts for Lektiner studier, selv om histologiske inndelinger kan brukes for tykkere vev. - Fjern cremaster muskelen fra testicle med et enkelt snitt. Legg den i iskalde PSS og feste det ut i en Petriskål med en silikon-elastomer fôr nederst i overflaten. Forsiktig erte bort bindevev bruker Dumont #5 tang.
- Skyll muskel prøvene med 2 mL bufrede PSS og deretter dyppe dem i rhodamine-merket GS1 Lektiner (20 µg/mL PSS) for 50 min i en 12-vel celle kultur plate med 2 mL/godt som er pakket i aluminiumsfolie utelate lys.
- Fjerne vev fra Lektiner løsning, skyll tre ganger i PSS med 5 min "vaske" incubations på en rocker og Monter dem på objektglass. Pass på å ruge vev i mørket og lagre lysbilder i mørket for å hindre tap av fluorescens.
Merk: Hvis lysbildene kan brukes umiddelbart, kan de holdes i kjøleskapet uten tap av fluorescens. For langvarig lagring, kan lysbildene oppbevares i en fryser for å hindre forverring11. - Evaluere microvessel tetthet av opptellingen antallet av kryss av merket microvessels datagenerert rutenettlinjer lagt over bildet52eller et klart rutenettet overlegg lagt over skjermen brukes til å vise lysbildene.
Merk: GS1 Lektiner tilnærminger har blitt brukt til å demonstrere: salt-indusert mikrovaskulær rarefaction53, beskyttende effekten av hindre salt indusert angiotensin II undertrykkelse i å gjenopprette microvessel tetthet i salt-matet dyr17 ,53,54; rollen NRF2 formidling beskyttende effekten av lav dose angiotensin II infusjon å forhindre microvessel rarefaction i salt-matet rotter17; og også for å vurdere rollen av angiotensin II opprettholde angiogenic Svar å kronisk muskelstimulasjon salt matet rotter54,55. En fordel av GS1 Lektiner teknikken er at det kan brukes å vurdere microvessel tetthet i de samme dyrene som brukes for studier av cannulated motstand arterier eller LDF.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
In vitro mikroskopi av cannulated motstand arterier tillater for studier av faktorer som påvirker aktive tonen i liten motstand arteriene (og større arterioler) ved normal i vivo transmuralt trykk og i fravær av parenchymal celle påvirker. I tillegg til å vurdere reaktivitet av fartøyene til ulike vasodilator og vasoconstrictor stimuli og myogenic svar på transmuralt press høyde i normal PSS, Ca2 +-gratis PSS kan legges til perfusate og superfusate på slutten av eksperimentet å angi maksimal fartøyet diameter og veggen tykkelsen. Siste målingene er svært verdifull i vurderingen vaskulær remodeling, dvs.endringer i veggen/lumen forholdet som kan oppstå i respons på endringer i blodtrykk eller administrasjon av farmakologiske agenter. Målinger av diameteren på maksimalt strekte fartøy i Ca2 +-gratis PSS er nyttig for å beregne iboende tone som ((DMaks-Dresten) /DMaks) x 100, der Dmax og Dresten er maksimalt (Ca2 +- Gratis PSS) og hvile diameter (PSS), henholdsvis på kontroll balanse presset (vanligvis 80 mmHg for MCA). I vår erfaring gjennomsnitt aktive tonen i MCA rundt 40% ved transmuralt trykk av 80 mmHg. Måle diameteren på maksimalt utvidede blodårer er også en praktisk måte å vurdere passiv mekanisk egenskapene til fartøyene måle diameteren av arteries på ulike nivåer av transmuralt distending press og bruke resultatene til beregne stress-belastning relasjoner og andre mekaniske egenskaper av fartøy7,56.
Figur 3 er en skjematisk fremstilling av tre avgrensede congenic stammer fra SS.13BN rotta utnytte flere selektiv formering tilnærminger. I disse rotte stammer, chromosomal segmentene inkluderer enten Brown Norge renin allelet (Ren1-BN) eller avskåret like over (Ren1-SSA) eller bare under (Ren1-SSB) renin genet locus (dermed beholder SS renin allelet). Som studere29, chromosomal segmentet som inneholder Brown Norge renin allelet inneholdt bare 25 gener.
Figur 4 illustrerer svaret endotelet avhengige av vasodilator ACh (1 µM) i isolerte MCAs fra Dahl SS hannrotter og tre falt congenic stammer vises i Figur 3. Kombinert bruk av disse tre congenic stammer som inkludere (Ren1-BN) eller kuttes av like over (Ren1-SSA) eller under (Ren1-SSB) av renin genet begrenset regionen rundt (dvs., området rundt renin genet locus) til ca. 25 gener. Disse eksperimentene, dyrene var normotensive og matet en lav salt (0,4% NaCl) diett. I denne studien29 endotelet avhengige av dilatasjon til ACh var fraværende i SS foreldrenes belastningen og i begge de congenic stammene beholde SS renin allelet, men ble restaurert i den congenic stammen bærer BN renin allelet i SS genetisk bakgrunnen. Resultatene av denne studien støttet resultatene av tidligere studier utnytte SS.13BN consomic rotter57,58 og gitt sterke bevis for at endotelial dysfunksjon finnes i SS rotter, selv når de er normotensive og Lei en lav salt diett. Tatt er sammen resultatene av disse studiene29,57,58 støtter hypotesen om at endotelial dysfunksjon i SS rotter på grunn av nedsatt regulering av renin genet, resulterer i kroniske eksponering til lav nivåer av angiotensin II i blodet.
En annen studie59, oppsummert i figur 5forhold svar på endotelet avhengige av vasodilator ACh i isolerte MCAs (MCA) fra Dahl SS hannrotter og SS.5BN consomic rotter bære kromosom 5 (med Brown Norge gener for ulike isoformene av cytochrome P450-4A ω-hydroksylase). I denne studien, dyr ble matet enten en lav salt (0,4% NaCl) eller høy salt (4% NaCl) diett. I motsetning til MCA fra SS rotter, ble endotelet avhengige dilatasjon til ACh opprettholdt i MCA fra SS.5BN consomic rotter innmatede lav salt diett. I tillegg (og i motsetning til resultatene fra SS.13BN consomic rotter57,58, Sprague-Dawley, rotter,33,,39,,40,,60og golden hamstere 61), en høy salt diett kunne eliminere ACh-indusert dilatasjon i SS.5BN rotter, indikerer at CYP450-4A ω-hydroksylase og 20-hydroxyeicosatetraenoic syre (20-HETE) er en viktig bidragsyter til vaskulær oksiderende stress og endothelial dysfunksjon i SS rotter og under høyder i salt inntak i andre stammer. Fra en eksperimentell perspektiv, svikt i de høye salt diett å eliminere endotelet avhengige av dilatasjon til ACh gir et eksempel på hvordan studier av cannulated motstand arterier kan produsere uventede funn som går fra konvensjonell visdom og kan føre til en ny forståelse av komplekse kontrollmekanismer påvirke funksjonen til disse avgjørende fartøy.
Figur 6 viser responsen av isolerte MCA ACh (1 µM) i vill type og Nrf2(- / -) knockout rotter matet en lav salt diett (0,4% NaCl), høy salt diett (4% NaCl), eller en høy salt diett som inneholder en kjent NRF2 aktivator30,62 . I fravær av salt-indusert oksidativt stress var ACh-indusert dilatasjon lignende MCA fra vill type og Nrf2(- / -) knockout rotter. I samsvar med resultatene av tidligere studier av Sprague-Dawley rotter, eliminert høy salt diett ACh-indusert dilatasjon i vill type og Nrf2(- / -) knockout rotter. Tillegg av NRF2 induser til de høye salt diett restaurert endotelet avhengige av dilatasjon til ACh i vill type rotter, men ikke i Nrf2(- / -) rotter, demonstrere vellykket eliminering av Nrf2 genet i knockout rotter, og støtter bruk av NRF2 opp-regulatorer som et terapeutisk tilnærming til å forbedre vaskulær oksidativt stress.
Figur 7 sammenfatte resultatet av LDF målinger sammenligne svar av pial mikrosirkulasjonen progressiv reduksjoner i arterial blodtrykket produsert av etterfølgende blod volum uttak i Sprague-Dawley rotter på enten en vanlig salt (0,4% NaCl) diett eller en høy salt (4% NaCl) diett i fire uker. I motsetning til rotter foret med en lav salt diett, matet rotter høy salt diett utstillingen en svekket evne til å opprettholde en konstant blodstrøm under reduksjoner i perfusjon press, indikerer at mekanismene ansvarlig for hjerne blod flyte autoregulation er kompromittert av langvarig eksponering for høy salt diett. Resultatene av disse eksperimentene er konsekvent med tilstedeværelse av nedsatt vaskulær avslapning mekanismer i isolerte cerebral arterier salt-matet rotter60 og gir et godt eksempel på hvordan LDF målinger kan benyttes til å støtte og forlenge den resultatene av studier ansette isolerte motstand arteries til nivået av hele vaskulær sengen.
I tillegg til å evaluere motstand arterien funksjonen bruker isolert arterier og vevsperfusjon bruker laser-Doppler flowmetry, microvessel tetthet og angiogenic svar på ulike stimuli som kronisk muskel stimulering54, 55 kan evalueres i de samme dyrene som bruker fluorescently-merket GS1 Lektiner, som selektivt bindes til glykoprotein moieties i kjelleren membranen av arterioler og kapillærer. Figur 8 viser typisk Lektiner farging i cremaster muskelen Sprague-Dawley rotter fôret en lav salt diett eller høy salt diett for 2 uker (Merk lavere tetthet av microvessels i høy salt matet rotter). Microvessel tetthet vurderes ved å telle kryss av Lektiner-merket microvessels med en computer-genererte referanse rutenettet52, og gir en utmerket metode for å vurdere angiogenic svar54,55 eller oppdage og kvantifisere tapet av microvessels i forhold som hypertensjon og forhøyet salt inntak12,17,53.

Figur 1: In vitro video mikroskopi oppsett er ansatt å studere isolerte, cannulated motstand arteriene (A). Cannulated motstand arterie knyttet til glass Mikropipetter (B) og video mikrometer displayet viser måling av lumen diameter i cannulated motstand arterie (C). Skala bar i panelet B representerer den interne diameteren arteria cannulated (µm), og 125 µm i panelet C er skjermen av avstanden mellom de flyttbare Referanselinjer plassert manuelt på de indre veggene av arterien av observatøren gjennomføre eksperimentet . Klikk her for å se en større versjon av dette tallet.

Figur 2: Skjematisk diagram av fremgangsmåten for å isolere arterien cerebri fra hjernen og forbereder det for cannulation med Mikropipetter. Klikk her for å se en større versjon av dette tallet.

Figur 3: Skjematisk diagram av avgrensede congenic rotten stammer har små segmenter av Brown Norge kromosom 13 introgressed i Dahl salt følsom genetisk bakgrunnen. Chromosomal segmenter inneholdt Brown Norge renin allelet (Ren1-BN) eller kuttes av like over (Ren1-SSA) eller like nedenfor (Ren1-SSB) renin genet locus og dermed beholde SS renin allelet. Dette tallet har blitt endret og gjengitt fra Durand et al. 29 med tillatelse fra American fysiologiske Society. Klikk her for å se en større versjon av dette tallet.

Figur 4: Response av isolert, cannulated MCAs av SS rotter og congenic rotten stammer (illustrert i figur 2) til 1 µM acetylkolin. Uttrykkes som gjennomsnittlig diameter endring (Δ µm) ±SEM etter hvile kontroll diameter før acetylkolin, og er replotted fra en opprinnelige studie av Durand et al. 29 Klikk her for å se en større versjon av dette tallet.

Figur 5: Svar av isolerte, cannulated MCAs av SS rotter og SS.5BN consomic rotter på 1 µM acetylcholine i dyr matet lav salt (0,4% NaCl) eller høy salt (4% NaCl) diett. Uttrykkes som gjennomsnittlig diameter endring (Δ µm) ±SEM etter hvile kontroll diameter før acetylkolin, og er replotted fra en opprinnelige studie av Lukaszewicz et al. 64 Klikk her for å se en større versjon av dette tallet.

Figur 6: Svar på 1 µM acetylcholine i isolerte, cannulated MCAs Nrf2(- / -) knockout rotter og wild kontrollerer matet lav salt diett (0,4% NaCl), høy salt diett (4% NaCl), eller høy salt diett som inneholder en kjent NRF2 aktivator30, 62. data uttrykkes som gjennomsnittlig diameter endring (Δ µm) ±SEM etter hvile kontroll diameter før acetylkolin. Klikk her for å se en større versjon av dette tallet.

Figur 7: Cerebral blodgjennomstrømning vurdert av LDF i pial blodsirkulasjonen. Sprague-Dawley rotter vedlikeholdes på en høy salt diett i fire uker utstilt en betydelig svekkelse i deres evne til å opprettholde blod flyte konstant som arterielt blodtrykk ble redusert svar på etterfølgende blod volum uttak. Dataene tegnes som mener prosent av kontroll ±SEM * p < 0,05 vs lavt saltinnhold diett på samme mener arteriell press. Klikk her for å se en større versjon av dette tallet.
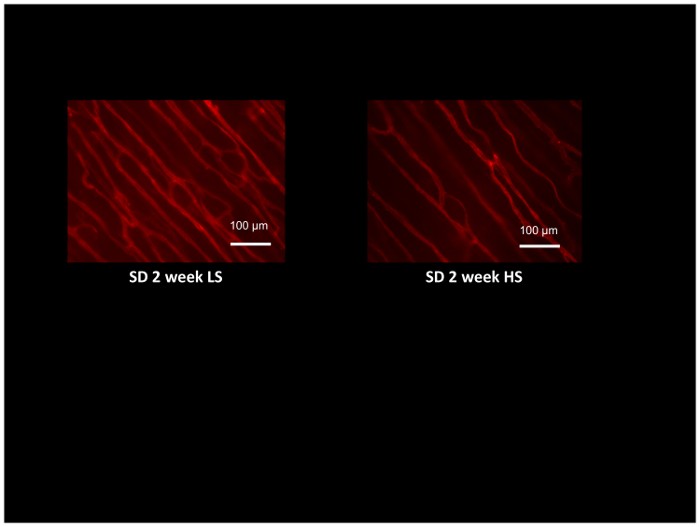
Figur 8: Mikroskop-bilde av rotte cremaster musklene merket med rhodamine-merket GS1 Lektiner å identifisere arterioler og kapillærer for vurdering av microvessel. Cremaster musklene Hentet fra Sprague-Dawley rotter fôret lav salt (LS, 0,4% NaCl) eller høy salt (HS:4% NaCl) diett i 2 uker, og demonstrere mikrovaskulær rarefaction i HS-matet dyr sammenlignet med LS kontroller. Skala bar = 100 µm. Klikk her for å se en større versjon av dette tallet.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Som nevnt i innledningen, dette dokumentet beskriver bruk av TV mikroskopi og isolert motstand arterien tilnærmingsmåter for å evaluere vaskulær funksjonen ikke bare i standard rotte modeller (som ansatt i videoen), men også i høyt spesialiserte genetisk foretatt rotte stammer, som viser romanen og kraftige innsikt som kan oppnås utnytte disse metodene. Bruk av disse kraftige teknikker å evaluere aktive tone og passiv mekaniske egenskaper av liten motstand arterier kan gi viktig informasjon om et bredt spekter av vaskulær kontrollmekanismer inkludert endotelet avhengige av regulering aktive tone i motstand arteriene, vaskulære glatt muskel funksjon under normale og patofysiologiske forhold og passiv egenskaper av arteries knyttet til vaskulær remodeling og endringer i vaskulære veggen mekanikk. Endotelial dysfunksjon har vist seg å være en kraftig prognostiske indikator på flere kardiovaskulære bivirkninger hos mennesker inkludert hjerte-relaterte død63og cannulated motstand arterien forberedelsene er spesielt nyttig i oppdage endotelial dysfunksjon og øke vår forståelse av mekanismer av endotelial dysfunksjon.
For å illustrere effekten av i vitro video mikroskopi teknikker ansette isolert motstand arteriene, gitt vi eksempler på bruk av disse teknikkene Dahl salt sensitive (SS) rotter og romanen consomic rotten stammer som viser en redusert salt følsomhet for blodtrykk sammenlignet med SS foreldrenes stamme24. Disse studiene undersøkt vaskulær kontrollmekanismer knyttet til gener på to kromosomene som er spesielt interessert i å bidra til salt følsomhet for blodtrykk og vaskulær endringer i SS rotte. Disse kromosomene er kromosom 13, bærer det renin genet18,22,29,57,58, og kromosom 5, bærer gener for isoformene av CYP450-4A ω-hydroksylase64 ,65, enzymet som syntetiserer 20-HETE, og har store innvirkninger på nyrefunksjonen og vaskulær reaktivitet66,67,68. En annen siste og kraftig tillegg til rotte genetisk verktøykassen er utviklingen av rotte genet knockout modeller bruker avansert genet redigeringsteknikker inkludert: ZFNs; transcriptional aktivator-lignende-effektor nucleases (TALENS), og nylig CRISPR-Cas9,13,,14,,15,,16,,17. In vitro video mikroskopi bruker disse teknikkene for å studere isolert MCAs fra en Nrf2(- / -) knockout rotte modell som mangler viktig antioksidant og celle beskyttende transkripsjon faktor NRF2, har gitt viktige og tidligere ukjent innsikt mekanismer av salt-indusert endotelial dysfunksjon i fravær av en opphøyd blod trykk17. Bestemte resultater eksperimenter utnytte disse spesialiserte rotte modeller er beskrevet i flere tidligere rapporter17,29,57,58,59, 60 , 64 , 65 .
Mens det er klart at studier av isolerte, cannulated motstand arterier er svært verdifulle i forstå mekanismene regulere funksjon av disse kritisk viktig fartøy under en rekke forhold, er det svært viktig å utøve en antall forholdsregler for å sikre at får nøyaktige og pålitelige resultater. Mens cerebral arterier og motstand arterieene av mange andre vaskulær senger viser iboende tone, må noen arteriene (spesielt hvem motstand arteriene) være pre kontrakt med vasoconstrictors som noradrenalin for å evaluere Svar å vasodilator stimuli, og mer presist simulere i vivo forhold, der båtene er under påvirkning av nevrale og humorale vasoconstrictor stimuli, som noradrenalin løslatt fra adrenerge nerve terminaler. Som sådan, er det viktig å bli kjent med de grunnleggende egenskapene av arteries studier, enten fra litteratur eller fra nøye gjennomført foreløpige eksperimenter. Fordi endotelet spiller en stor rolle i å regulere store arteriene, liten arterier og arterioler i blodsirkulasjonen, er det viktig å være forsiktig å unngå å skade endotelet under isolasjon og cannulation av arterien. Den klassiske testen for endotelet integritet er demonstrasjon at ACh forårsaker fartøyet dilatasjon. En påminnelse er at tilfeller av oksidativt stress fartøyet endotelet kan være intakt, men vasodilatation til ACh er fraværende fordi store mengder superoxide renovere nitrogenoksid hindre vasodilator svaret. I slike tilfeller kan endothelial integritet verifiseres ved å gjenta ACh programmet i nærvær av en superoxide åtseldyr som tempol, som skal gjenopprette vasodilatasjon svar ACh og andre endotelet avhengige av vasodilator stimuli. Også i en rekke pathological betingelser, kan endotelet frigi stoffer som forårsaker sammentrekning av vaskulær glatt muskelceller og innsnevring av arterien; og i noen tilfeller endotelet avhengige av dilatasjon (eller innsnevring) er formidlet via andre stoffer som cyclooxygenase metabolitter, H2O2, epoxygenase metabolitter, etc. Klassisk testen for endotelet avhengige av vasodilator eller vasoconstrictor stoff er å vise at dilation eller innsnevring av arterien er eliminert av endothelial fjerning. Til slutt, identiteten til forskjellige endotelet avhengige av vasodilator og vasoconstrictor stoffer kan vanligvis fastslås ved administrasjon av bestemte hemmere eller fjerner, for eksempel L-navn å hemme nitrogenoksid syntase, indomethacin å hemme dannelsen av cyclooxygenase metabolitter, catalase åtseleter H2O2, thromboxane syntase hemmere, epoxygenase hemmere, og motstanderne av den CYP450-4A/20-HETE veien.
Det er også viktig (og lærerikt) for å kvantifisere mengden aktive tone på slutten av eksperimentet av perfusing og superfusing arterien med en Ca2 +-gratis PSS, eller administrere en maksimal dose av en kraftige vasodilator agent som papaverine. Et typisk nivå av aktiv hvile tonen i MCA, beregnet som ((DMaks-Dresten) /Dmax) x 100, er ca 40%, der Dmax og Dresten er maksimum (Ca2 +-gratis PSS) og diameter (PSS), henholdsvis på kontroll balanse presset (vanligvis 80 mmHg for MCA). Fartøyer med betydelig mindre aktive tone eller arterier viser Segmentinformasjon constrictions eller dilations, ekskluderes fra analyse, som de tegnene er veiledende traume til fartøyene. Måling av diameter på maksimalt utvidede blodårer i Ca2 +-gratis løsning kan også etterforskeren å vurdere passiv mekanisk egenskapene til fartøyene måling arteriell diameter og beregning av stress-belastning relasjoner under sammenhengende fasader i transmuralt trykket i maksimalt strekte fartøy7,56. Disse passiv stress-belastning relasjoner hentes lett, og en verdifull indikasjon om eventuelle endringer i mekanisk egenskapene til fartøyene.
Renslighet av Pipetter, koblinger og rør leverer reservoarene er helt avgjørende for vellykket eksperimenter. I denne forbindelse er det viktig å tømme alle løsninger av slangen etter eksperimentet er fullført, og til å rense og rense vevet bad, levering slangen og alle reservoarer brukes til å lagre, varme, og gass-equilibrate PSS før nå fartøyet kammeret. Stopcocks og ventiler i levering systemet bør også rengjøres og endres med jevne mellomrom som skal noen rør bærer PSS. Et klassisk tegn på forurenset slangen er en grå dis generert av mugg og bakterier; og endringene ledsages av tap av normal reaktivitet i blodårene skyldes andre stoffer produsert av bakteriell forurensning. Forurensning av bakterier og andre mikroorganismer kan imidlertid fortsatt være tilstede i fravær av noen synlig bevis.
Vi tror at dagens papir gir et nyttig eksempel for bruk av tid-æret teknikker som er eksepsjonelt velegnet for studier av viktige liten motstand arteries av ulike vaskulær senger. Kombinert med standard tilnærmingsmåter for å evaluere vevsperfusjon, som LDF og GS1 Lektiner metode for å vurdere microvessel tetthet, i vitro video mikroskopi av cannulated motstand arterier gir svært verdifull innsikt i den faktorer som kontrollerer vevsperfusjon og hvordan disse kan endres i sykdomstilstander. I tillegg gir en kraftig måte å studere grunnleggende mekanismer for vaskulær glatt muskel og endothelial funksjon i standard rotte modeller, kan bruk av video mikroskopi å studere individuell arterier brukes til andre dyr modeller og menneskelig motstand arterier. Bruk av video mikroskopi av isolerte motstand arterier romanen genmodifiserte rotte modeller åpner nye dører å forstå fenotypiske endringer som oppstår som svar på endrede funksjon av et mangfold (og stadig voksende) av gener.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingen konkurrerende økonomiske interesser.
Acknowledgments
Forfatterne uttrykker deres oppriktige takk til Katie Fink og Lynn Dondlinger for deres uvurderlig hjelp i utarbeidelsen av dette manuskriptet.
Gi støtte: NIH #R21-OD018309; #R56-HL065289; og #R01-HL128242.
Materials
| Name | Company | Catalog Number | Comments |
| SS Rat | Medical College of Wisconsin | SS/JHsd/Mcwi strain | Contact Dr. Aron Geurts (ageurts@mcw.edu) |
| SS.5BN Consomic Rat | Medical College of Wisconsin | SS-Chr 5BN/Mcwi strain | Contact Dr. Aron Geurts (ageurts@mcw.edu) |
| SS.13BN Consomic Rat | Medical College of Wisconsin | SS-Chr 13BN/Mcwi strain | Contact Dr. Aron Geurts (ageurts@mcw.edu) |
| Ren1-BN Congenic Rat | Medical College of Wisconsin | SS.BN-(D13hmgc41-D13)hmgc23/Mcwi strain | Contact Dr. Aron Geurts (ageurts@mcw.edu) |
| Ren1-SSA Congenic Rat | Medical College of Wisconsin | SS.BN-(D13rat77-D13rat105/Mcwi strain | Contact Dr. Aron Geurts (ageurts@mcw.edu) |
| Ren1-SSB Congenic Rat | Medical College of Wisconsin | SS.BN-(D13rat124-D13rat101/Mcwi strain | Contact Dr. Aron Geurts (ageurts@mcw.edu) |
| Nrf2(-/-) Knockout Rat and Wild Type Littermates | Medical College of Wisconsin | SD-Nfe212em1Mcwi strain | Contact Dr. Aron Geurts (ageurts@mcw.edu) |
| Low Salt Rat Chow (0.4% NaCl)-AIN-76A | Dyets, Inc. | 113755 | |
| High Salt Rat Chow (4% NaCl)-AIN-76A | Dyets, Inc. | 113756 | |
| Colorado Video Caliper | Colorado Video, Inc. | Model 308 | |
| Video Camera | Hitachi | KPM1AN | |
| Microscope | Olympus Life Science | CKX41 | |
| Television Monitor | Panasonic | WVBM1410 | |
| Pressure Transducers | Stoelting | 56360 | |
| Blood Pressure Display Unit | Stoelting | 50115 | |
| Cannulated Artery Chamber | Living Systems Instrumentation | CH-1 | Single vessel chamber for general use |
| Temperature Controller for Single Chamber | Living Systems Instrumentation | TC-09S | |
| Gas Dispersion Tube, Miniature,Straight | Living Systems Instrumentation | GD-MS | Provides aeration in the vessel bath |
| Gas Exchange Oxygenator, Miniature | Living Systems Instrumentation | OX | Allows gas exchange with perfusate |
| Laser-Doppler Flowmeter | Perimed | PeriFlux 5000 LDPM | |
| GS1 Lectin | Vector Labs | RL-1102 | |
| Glass Capillary Tubes for Micropipettes | Fredrich Haer Co. | 27-33-1 | 2 mm ODX1 mm ID |
| Verticle Pipette Puller | David Kopf Instruments | Model 700C | |
| Nylon suture material (10/0)-3 PLY | Ashaway Line and Twine Manufacturing Co. | 114-ANM-10 | Single strands of 3 ply nylon suture teased out for use on vessels |
| Dumont #5 Forceps-Inox | Fine Science Tools | 11254-20 | |
| Vannas Scissors | Fine Science Tools | 15003-08 | |
| Protandim | Protandim | NRF2 Inducer: Contact Dr. Joe McCord (JOE.MCCORD@UCDENVER.EDU) | |
| Sodium Chloride | Fisher Bioreagents | BP358-212 | |
| Sodium Bicarbonate | Fisher Chemical | S233-3 | |
| Dextrose (d-glucose) anhydrous | Fisher Chemical | D16-500 | |
| Magnesium Sulfate (MgSO4-7H2O) | Sigma Aldrich | M1880-500 G | |
| Calcium Chloride (CaCl2-2 H2O) | Sigma | C5080-500G | |
| Sodium Phosphate-Monobasic (NaH2PO4) | Sigma | S0751-500G | |
| Potassium Chloride (KCl) | Fisher Chemical | P217-500G | |
| Ethylenediaminetetraacetic acid disodium salt dihydrate (EDTA) | Sigma | ED255-500G |
References
- Furchgott, R. F., Zawadzki, J. V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature. 288, 373-376 (1980).
- Bevan, J. A., Osher, J. V. A direct method for recording tension changes in the wall of small blood vessels in vitro. Agents Actions. 2, 257-260 (1972).
- Mulvany, M. J., Halpern, W. Contractile properties of small arterial resistance vessels in spontaneously hypertensive and normotensive rats. Circ. Res. 41, 19-26 (1977).
- Speden, R. N. The use of excised, pressurized blood vessels to study the physiology of vascular smooth muscle. Experientia. 41, 1026-1028 (1985).
- Osol, G., Halpern, W. Myogenic properties of cerebral blood vessels from normotensive and hypertensive rats. Am. J. Physiol. 249, H914-H921 (1985).
- Halpern, W., Kelley, M. In vitro methodology for resistance arteries. Blood Vessels. 28, 245-251 (1991).
- Feihl, F., Liaudet, L., Waeber, B. The macrocirculation and microcirculation of hypertension. Curr Hypertens Rep. 11, 182-189 (2009).
- Smits, G. J., Roman, R. J., Lombard, J. H. Evaluation of laser-Doppler flowmetry as a measure of tissue blood flow. J Appl Physiol. 61, 666-672 (1985).
- Hudetz, A. G., Roman, R. J., Harder, D. R. Spontaneous flow oscillations in the cerebral cortex during acute changes in mean arterial pressure. J Cereb Blood Flow Metab. 12, 491-499 (1992).
- Hudetz, A. G., Smith, J. J., Lee, J. G., Bosnjak, Z. J., Kampine, J. P. Modification of cerebral laser-Doppler flow oscillations by halothane, PCO2, and nitric oxide synthase blockade. Am J Physiol. 269, H114-H120 (1995).
- Hansen-Smith, F. M., Watson, L., Lu, D. Y., Goldstein, I. Griffonia simplicifolia I: fluorescent tracer for microcirculatory vessels in nonperfused thin muscles and sectioned muscle. Microvasc Res. 36, 199-215 (1988).
- Greene, A. S., Lombard, J. H., Cowley, A. W., Hansen-Smith, F. M. Microvessel changes in hypertension measured by Griffonia simplicifolia I lectin. Hypertension. 15, 779-783 (1990).
- Aitman, T., Dhillon, P., Geurts, A. M. A RATional choice for translational research? Dis Model Mech. 9, 1069-1072 (2016).
- Geurts, A. M., et al. Knockout rats via embryo microinjection of zinc-finger nucleases. Science. 325, 433 (2009).
- Geurts, A. M., et al. Generation of gene-specific mutated rats using zinc-finger nucleases. Methods Mol Biol. 597, 211-225 (2010).
- Geurts, A. M., Moreno, C. Zinc-finger nucleases: new strategies to target the rat genome. Clin Sci (Lond). 119, 303-311 (2010).
- Priestley, J. R., Kautenburg, K. E., Casati, M. C., Endres, B. T., Geurts, A. M., Lombard, J. H. The NRF2 knockout rat: a new animal model to study endothelial dysfunction, oxidant stress, and microvascular rarefaction. Am J Physiol Heart Circ Physiol. 310, H478-H487 (2016).
- Cowley, A. W., et al. Brown Norway chromosome 13 confers protection from high salt to consomic Dahl S rat. Hypertension. 37, 456-461 (2001).
- Rapp, J. P. Dahl salt-susceptible and salt-resistant rats. A review. Hypertension. 4, 753-763 (1982).
- Rapp, J. P., Wang, S. M., Dene, H. A genetic polymorphism in the renin gene of Dahl rats cosegregates with blood pressure. Science. 243, 542-544 (1989).
- Manning, R. D. Jr, Meng, S., Tian, N. Renal and vascular oxidative stress and salt-sensitivity of arterial pressure. Acta Physiol Scand. 179, 243-250 (2003).
- Moreno, C., et al. Multiple blood pressure loci on rat chromosome 13 attenuate development of hypertension in the Dahl S hypertensive rat. Physiol Genomics. 31, 228-235 (2007).
- Tobian, L., Lange, J., Iwai, J., Hiller, K., Johnson, M. A., Goossens, P. Prevention with thiazide of NaCl-induced hypertension in Dahl "S" rats. Evidence for a Na-retaining humoral agent in "S" rats. Hypertension. 1, 316-323 (1979).
- Mattson, D. L., et al. Chromosome substitution reveals the genetic basis of Dahl salt-sensitive hypertension and renal disease. Am J Physiol Renal Physiol. 295, F837-F842 (2008).
- Kunert, M. P., et al. Consomic strategies to localize genomic regions related to vascular reactivity in the Dahl salt-sensitive rat. Physiol Genomics. 26, 218-225 (2006).
- Cowley, A. W., Liang, M., Roman, R. J., Greene, A. S., Jacob, H. J. Consomic rat model systems for physiological genomics. Acta Physiol Scand. 181, 585-592 (2004).
- Kunert, M. P., Dwinell, M. R., Lombard, J. H. Vascular responses in aortic rings of a consomic rat panel derived from the Fawn Hooded Hypertensive strain. Physiol Genomics. 42A, 244-258 (2010).
- Liang, M., et al. Renal medullary genes in salt-sensitive hypertension: a chromosomal substitution and cDNA microarray study. Physiol Genomics. 8, 139-149 (2002).
- Durand, M. J., Moreno, C., Greene, A. S., Lombard, J. H. Impaired relaxation of cerebral arteries in the absence of elevated salt intake in normotensive congenic rats carrying the Dahl salt-sensitive renin gene. Am J Physiol Heart Circ Physiol. 299, H1865-H1874 (2010).
- Hybertson, B. M., Gao, B., Bose, S. K., McCord, J. M. Oxidative stress in health and disease: the therapeutic potential of Nrf2 activation. Mol Aspects Med. 32, 234-246 (2011).
- Itoh, K., et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem Biophys Res Commun. 236, 313-322 (1997).
- Myung, S. K., et al. Efficacy of vitamin and antioxidant supplements in prevention of cardiovascular disease: systematic review and meta-analysis of randomised controlled trials. BMJ. 346, f10 (2013).
- Fredricks, K. T., Liu, Y., Lombard, J. H. Response of extraparenchymal resistance arteries of rat skeletal muscle to reduced PO2. Am J Physiol. 267, H706-H715 (1994).
- Fredricks, K. T., Liu, Y., Rusch, N. J., Lombard, J. H. Role of endothelium and arterial K+ channels in mediating hypoxic dilation of middle cerebral arteries. Am J Physiol. 267, H580-H586 (1994).
- Frisbee, J. C., Maier, K. G., Falck, J. R., Roman, R. J., Lombard, J. H. Integration of hypoxic dilation signaling pathways for skeletal muscle resistance arteries. Am J Physiol Regul Integr Comp Physiol. 283, R309-R319 (2002).
- Pavlov, T. S., Ilatovskaya, D. V., Palygin, O., Levchenko, V., Pochynyuk, O., Staruschenko, A. Implementing Patch Clamp and Live Fluorescence Microscopy to Monitor Functional Properties of Freshly Isolated PKD Epithelium. J Vis Exp. (103), (2015).
- Nelson, M. T., Conway, M. A., Knot, H. J., Brayden, J. E. Chloride channel blockers inhibit myogenic tone in rat cerebral arteries. J Physiol. 502 (Pt 2), 259-264 (1997).
- Brayden, J. E., Halpern, W., Brann, L. R. Biochemical and mechanical properties of resistance arteries from normotensive and hypertensive rats. Hypertension. 5, 17-25 (1983).
- Weber, D. S., Lombard, J. H. Elevated salt intake impairs dilation of rat skeletal muscle resistance arteries via ANG II suppression. Am J Physiol Heart Circ Physiol. 278, H500-H506 (2000).
- Weber, D. S., Lombard, J. H. Angiotensin II AT1 receptors preserve vasodilator reactivity in skeletal muscle resistance arteries. Am J Physiol Heart Circ Physiol. 280, H2196-H2202 (2001).
- Wang, J., Roman, R. J., Falck, J. R., de la Cruz, L., Lombard, J. H. Effects of high-salt diet on CYP450-4A omega-hydroxylase expression and active tone in mesenteric resistance arteries. Am J Physiol Heart Circ Physiol. 288, H1557-H1565 (2005).
- Raffai, G., et al. Modulation by cytochrome P450-4A omega-hydroxylase enzymes of adrenergic vasoconstriction and response to reduced PO2 in mesenteric resistance arteries of Dahl salt-sensitive rats. Microcirculation. 17, 525-535 (2010).
- Mishra, R. C., Wulff, H., Hill, M. A., Braun, A. P. Inhibition of Myogenic Tone in Rat Cremaster and Cerebral Arteries by SKA-31, an Activator of Endothelial KCa2.3 and KCa3.1 Channels. J Cardiovasc Pharmacol. 66, 118-127 (2015).
- Freed, J. K., Beyer, A. M., LoGiudice, J. A., Hockenberry, J. C., Gutterman, D. D. Ceramide changes the mediator of flow-induced vasodilation from nitric oxide to hydrogen peroxide in the human microcirculation. Circ Res. 115, 525-532 (2014).
- Beyer, A. M., Durand, M. J., Hockenberry, J., Gamblin, T. C., Phillips, S. A., Gutterman, D. D. An acute rise in intraluminal pressure shifts the mediator of flow-mediated dilation from nitric oxide to hydrogen peroxide in human arterioles. Am J Physiol Heart Circ Physiol. 307, H1587-H1593 (2014).
- Durand, M. J., et al. Vascular actions of angiotensin 1-7 in the human microcirculation: novel role for telomerase. Arterioscler Thromb Vasc Biol. 36, 1254-1262 (2016).
- Beyer, A. M., et al. Transition in the mechanism of flow-mediated dilation with aging and development of coronary artery disease. Basic Res Cardiol. 112, 5 (2017).
- Muller, J. M., Chilian, W. M., Davis, M. J. Integrin signaling transduces shear stress--dependent vasodilation of coronary arterioles. Circ Res. 80, 320-326 (1997).
- Liu, Y., Harder, D. R., Lombard, J. H. Interaction of myogenic mechanisms and hypoxic dilation in rat middle cerebral arteries. Am J Physiol Heart Circ Physiol. 283, H2276-H2281 (2002).
- Potocnik, S. J., et al. Endothelium-dependent vasodilation in myogenically active mouse skeletal muscle arterioles: role of EDH and K+ channels. Microcirculation. 16, 377-390 (2009).
- Harder, D. R. Pressure-dependent membrane depolarization in cat middle cerebral artery. Circ Res. 55, 197-202 (1984).
- Greene, A. S., Rieder, M. J. Measurement of vascular density. Methods Mol. Med. 51, 489-496 (2001).
- Hernandez, I., Cowley, A. W., Lombard, J. H., Greene, A. S. Salt intake and angiotensin II alter microvessel density in the cremaster muscle of normal rats. Am J Physiol. 263, H664-H667 (1992).
- Resende, M. M., Amaral, S. L., Moreno, C., Greene, A. S. Congenic strains reveal the effect of the renin gene on skeletal muscle angiogenesis induced by electrical stimulation. Physiol Genomics. 33, 33-40 (2008).
- Petersen, M. C., Munzenmaier, D. H., Greene, A. S. Angiotensin II infusion restores stimulated angiogenesis in the skeletal muscle of rats on a high-salt diet. Am J Physiol Heart Circ Physiol. 291, H114-H120 (2006).
- Frisbee, J. C., Weber, D. S., Liu, Y., DeBruin, J. A., Lombard, J. H. Altered structure and mechanics of skeletal muscle arteries with high-salt diet and reduced renal mass hypertension. Microvasc Res. 59, 323-328 (2000).
- Drenjancevic-Peric, I., Lombard, J. H. Introgression of chromosome 13 in Dahl salt-sensitive genetic background restores cerebral vascular relaxation. Am J Physiol Heart Circ Physiol. 287, H957-H962 (2004).
- Drenjancevic-Peric, I., Phillips, S. A., Falck, J. R., Lombard, J. H. Restoration of normal vascular relaxation mechanisms in cerebral arteries by chromosomal substitution in consomic SS.13BN rats. Am J Physiol Heart Circ Physiol. 289, H188-H195 (2005).
- Lukaszewicz, K. M., Paudyal, M. P., Falck, J. R., Lombard, J. H. Role of vascular reactive oxygen species in regulating cytochrome P450-4A enzyme expression in Dahl salt-sensitive rats. Microcirculation. 23, 540-548 (2016).
- Lombard, J. H., Sylvester, F. A., Phillips, S. A., Frisbee, J. C. High-salt diet impairs vascular relaxation mechanisms in rat middle cerebral arteries. Am J Physiol Heart Circ Physiol. 284, H1124-H1133 (2003).
- Priestley, J. R., et al. Reduced angiotensin II levels cause generalized vascular dysfunction via oxidant stress in hamster cheek pouch arterioles. Microvasc Res. 89, 134-145 (2013).
- Velmurugan, K., Alam, J., McCord, J. M., Pugazhenthi, S. Synergistic induction of heme oxygenase-1 by the components of the antioxidant supplement Protandim. Free Radic Biol Med. 46, 430-440 (2009).
- Widlansky, M. E., Gokce, N., Keaney, J. F., Vita, J. A. The clinical implications of endothelial dysfunction. J Am Coll Cardiol. 42, 1149-1160 (2003).
- Lukaszewicz, K. M., Falck, J. R., Manthati, V. L., Lombard, J. H. Introgression of Brown Norway CYP4A genes on to the Dahl salt-sensitive background restores vascular function in SS-5BN consomic rats. Clin Sci (Lond). 124, 333-342 (2013).
- Lukaszewicz, K. M., Lombard, J. H. Role of the CYP4A/20-HETE pathway in vascular dysfunction of the Dahl salt-sensitive rat. Clin Sci (Lond). 124, 695-700 (2013).
- Roman, R. J. P-450 metabolites of arachidonic acid in the control of cardiovascular function. Physiol Rev. 82, 131-185 (2002).
- Roman, R. J., Maier, K. G., Sun, C. W., Harder, D. R., Alonso-Galicia, M. Renal and cardiovascular actions of 20-hydroxyeicosatetraenoic acid and epoxyeicosatrienoic acids. Clin Exp Pharmacol. 27, 855-865 (2000).
- Roman, R. J., Alonso-Galicia, M. P-450 eicosanoids: A novel signaling pathway regulating renal function. News Physiol Sci. 14, 238-242 (1999).
