

Summary
환경 독 소에 노출 수 있습니다 심하게 발전 태아에 영향을. 내 분 비 혼란 시키는 화학 제품 bisphenols 부정적인 알려져 있습니다와 같은 신 경계에 영향을. 여기는 초기 배아에 독 소 노출의 기능에 미치는 영향을 연구 하는 생체 외에서 (병아리 태아) 척추 신경 네트워크 모델을 사용 하는 프로토콜에 설명 합니다.
Abstract
두번째-페 놀, 비스 페 놀 A (BPA)와 두번째-페 놀-S (BPS), 플라스틱 및 수많은 일상적인 제품의 생산에서 널리 이용 되는 대리인 polymerizing 있습니다. 그들은 내 분 비 혼란 시키는 화합물 (EDC) estradiol 같은 속성으로 분류 됩니다. 장기 노출 EDCs, 낮은 복용량 에서도 다양 한 건강 결함 등 암, 행동 장애, 불 임, 초기 발달 기간 동안 큰 취약점으로 연결 되었습니다. 신경 기능의 개발에 대 한 BPA의 효과 연구, 우리는 체 외에서 신경 네트워크 모델 초기 배아 여자 뇌에서 파생 된 사용. 우리는에 노출 발견 BPA 특히 급상승 활동 및 동기화 네트워크 활동의 개발에 영향을. 네트워크 활동에 변화는 분자 화합물 또는 약의 대상과 행동 결과에 미치는 영향 사이의 중요 한 링크입니다. 다중 전극 배열 점점 네트워크 활동 체 외에 약물의 효과 연구에 유용한 도구가 되고있다. 여러 시스템 사용할 수 시장에서 있으며, 전극, 유형 및 전극 배열 및 기본 기본 원리, 분석 소프트웨어의 품질의 수에 있는 변이 그리고 데이터 취득은 걸쳐 동일 합니다 다른 시스템입니다. 현재 2 차원 생체 외에서 문화 분석에 국한, 하지만 이러한 MEA 시스템 네트워크 활동 두뇌 조각에서 vivo에서 사용할 수 있도록 향상 되 고 된다. 여기, 우리는 배아 노출 및 신경 네트워크 활동 및 대표 결과 함께 synchrony에 대 한 상세한 프로토콜을 제공 합니다.
Introduction
4, 4 '-Isopropylidenediphenol, 일반적으로 언급으로 Bisphenol-A 또는 BPA, 산화 첨가제에에서 있으면 다양 한 폴 리 카보 네이트 및 열 종이에서 배열 하는 에폭시 수 지 기반 제품, Cd, 및 균열 증거 유리 안쪽에의 코팅 음료 캔. BPA 에스트로겐1을 내 분 비 방해 대리인 것으로 알려져 있다, 비록 그것의 병에 효과에 BPA 매일 우연한 노출 때문에 대 한 연구는 주로 마지막 10-15 년에 밖으로 나와. BPA 노출도 낮은 레벨의 결과 어머니2,3의 노출을 통해 embryogenesis 동안 포함 한 초기 발달 기간에 가장 심오한. Utero에서 노출 여성 태아의 내 분 비 방해 화학 물질에는 또한 유 방 암4,5질에서 증가 질병 민감성에 연결 되었다. 동물 연구는 BPA에 노출 리드 비정형 뇌 구조와 기능 이상이 동작6에 설명 했다. 결과적으로, 유아 젖 병에 BPA 사용 하 여 유럽과 북미 지역, FDA 등에서 가장 규제 기관에 의해 금지 되었습니다. 규정 준수, 많은 제조 업체 4, 4'-sulfonyldiphenol 또는 bisphenol-S (BPS)로 전환 합니다. BPA와 BPS 인 구조 아날로그와 최근 보고서 estrogenic 전사7BPS의 유사한 효능을 보였다는 사실에 근거한 BPA 기준으로이 화합물의 독성 연구 중요 하다.
여기는 병아리 태아 척추 모델을 사용 하 여 신경 세포의 네트워크에 BPA (및 다른 EDCs)의 효과 테스트 하는 프로토콜에 설명 합니다. 생체 외에서 문화 뉴런의 시 냅 스 연락처를 형성 하 고 (스파이크 라고도 함) 하는 활동 전위를 생성. 이러한 문화의 급격히 활동 다중 전극 배열 (MEA) 시스템을 사용 하 여 기록할 수 있습니다. 스파이크는 synchrony에 있을 때 그들은 서로 5 ms 이내 발생으로 간주 됩니다. 결국 동기화 초기 랜덤 급격히 활동 신경 네트워크8,9개발의 핵심 기능입니다. Synchrony 다양 한 방법을 사용 하 여 측정 될 수 있다 그리고 몇 가지 알고리즘 문학9,,1011에 설명 되어 있습니다. 우리의 분석에서 우리는 MEA 수집 시스템 드라이브 레코딩 소프트웨어에 통합 되어 있는 동료와 Paiva12 에 의해 개발 된 알고리즘을 사용 합니다. 미 발달 병아리 뉴런의 강력한 스파이크 활동 신경 네트워크 활동13에 BPA의 효과 공부에 대 한 프로토 타입을 제공 합니다. 관찰 기록 급격히 활동 다중 전극 배열 사용 하 여, BPA 노출 신경 스파이크 synchrony9,14의 개발을 억제 합니다. 여기, 우리는 병아리 배아 문화에 신경 스파이크 synchrony의 개발에 대표적인 결과 함께 문화에 병아리 태아 신경에 BPA 노출 공부에 대 한 상세한 방법론을 제공 합니다.
Protocol
아래에 주어진 프로토콜 (초기) embryogenesis, 동안 BPA 노출의 효과 테스트를 표준화 하 고 BPS, BPF, 또는 병아리 배아 뉴런에서 다른 EDC와 함께 사용 하기 위해 수정할 수 있습니다.
이 프로토콜에는 델라웨어 주립 대학의 제도적 정책 및 조류 배아에 대 한 NIH 정책 다음과 같습니다. 프로토콜은 DSU의 재료와 화학 안전 지침 확인입니다.
1. 소재 설정
- 1 m m 재고 솔루션 (10% 에타 놀의 mL 당 0.23 m g)을 물에 10%의 에탄올 (v/v)에서 BPA (m.w. 228.29)을 분해. Neurobasal 매체에 후속 희석 (0.1 µ M, 0.5 µ M, 1 µ M, 5 µ M 그리고 10 µ M)를 확인 합니다.
주의: BPA는 환경 독 소 그리고 분말을 처리할 때 주의 행사 해야 합니다. - 37 ° c.에 상용 탁상 계란 인큐베이터에서 병아리 알 품 어 E7 배아에 대 한 일 동안 알을 품 어.
- 3 ~ 4 h 70% 에탄올에 몸을 담글 하 여 측정 Sterilize MEAs의 필요한 수를 준비 하 고 세 번 약 10 mL BSLII 후드에 멸 균 증류수에 씻어.
참고:는 MEAs는 8 x 8 표에 배열 64 성공 백 금 전극 포함. 전극 직경 30 µ m 이며, 전극 사이의 간격 200 µ m 이다. 로이 MEA의 부식으로 이어질 것입니다 MEAs는 살 균에 대 한 변성된 에탄올을 사용 하지 마십시오. - (무 균 상태를 유지)에 뚜껑 살 균 컨테이너 내부 MEAs 장소 테이블 상단 인큐베이터로 이동. 55 ° c.에 적어도 6 시간 동안 구워 이 단계는 적절 한 살 균을 위해 중요 하 고 온도 60 ° c.를 초과 하지 않아야 합니다. 무 균 유지, 추가 사용까지 MEAs 스토리지에 대 한 BSLII 후드를 포함 하는 접시를 이동 합니다.
참고: 우리는 플라스틱 뚜껑 오븐 안전한 유리 베이킹 접시 사용 하 고 표면 70% 에탄올과 건조에 대 한 BSLII 후드에서 소독. - 약 수 엑스트라 세포 매트릭스 ECM 솔루션입니다. 밤새 4 ° C에서 유리병을 녹여 및 동결-20 ° c.에 100 µ L aliquots
참고: ECM 냉동 배송입니다. ECM 해야 하지 주어질 실내 온도에 준비까지 코트와 실내 온도에 polymerizes, 그것 생산, 일단 코트 수는 없습니다. ECM 성장 인자 없이 알 수 없는 요인으로 인해 추가 효과 방지 하기 위해 선호 된다. - 모든 해 부 악기 (Dumont #5 집게, 곡선된 겸 자, 작은 위, 및 봄이 위) 소독 자체 씰링 소재 70% 에탄올과 해 부 요리를 코팅 하 고 절 개 후드에 말리.
2. 병아리 배아 신경 문화와 네트워크 활동
- 도금의 날, 제거 된 유리병-20 ° C 냉동 고에서 ECM의 70% 에탄올 스프레이 얼음에. BSLII 후드 안쪽에 300 µ L 찬 neurobasal 매체를 추가 하 여 25% 희석.
- p 200를 사용 하 여 pipetteman을 터치 전극 표면에 박막을 떠나 즉시 제거 돌보는 MEA의 중심에 25 %ECM 100 µ L를 추가 합니다. MEA를 커버 하 고 신경 세포를 접시에 준비까지 CO2 배양 기 (37 ° C, 5% CO2)에 배치.
- 70% 에탄올과 E7 달걀 (배아 7 일, 7 일 동안 37 ° C에서 incubated)의 외피를 소독. 감기 균 행 크의 균형 염 솔루션 (HBSS) 칼슘 없이 포함 된 Sylgard 하단 해 부 접시에 배아를 목을 벨. 눈 주위 고 눈알을 제거. 집게를 잘 몬 트 # 5를 사용 하 여 봄이 위 복 부에 절 개를 확인 하 고 개 및 광학 tectum 폭로 피부의 바깥 층을 제거. 껍질 하 고 신중 하 게 pial 멤브레인을 제거 합니다. 다른 페 트리 접시에는 개를 전송 하 고 봄이 위 약 2 m m의 작은 조각으로 잘라.
참고: 거기 해서는 안됩니다 pial 막의 제거 후에 개에 연결 된 어떤 혈관. 비록 우리 악기와 해 부 지역 70% 에탄올으로 소독 철저 하 게은으로 절 개 후드 밖에 서 수행 하는 경우 상당히 좋은 결과 얻은 그것은 메 마른 절 개 후드, 해 부를 수행 하는 것이 좋습니다 가능한 경우, . - 살 균 전송 피 펫을 사용 하 여, 15 mL 원심 분리기 관으로 개 조각을 수집 하 고 제거 가능한 HBSS 개 싱크 분리기 튜브의 바닥의 조각을 시키는 후.
- 0.05%의 1 mL을 추가 트립 신/EDTA (37 ° C에 미리 예 열)와 15 분 동안 37 ° C에서 품 어.
- 파스퇴르 피 펫을 사용 하 여 신중 하 게 조직 조각을 방해 하지 않고 트립 신을 제거 하 고 1 mL neurobasal 매체를 추가. 조직 싱크대 바닥에 조각 하 게 하 고 매체를 제거 합니다. 이 세척을 한 번 더 반복 합니다. 이 단계는 트립 신/EDTA를 씻어.
- Neurobasal 매체의 2 개 mL를 추가 하 고 분쇄 시작. 분쇄는 메 마른 불 광택 파스퇴르 피 펫을 복용 하 고 그것을 통해 조직의 여러 번 통과 부드럽게 조직의 아니 더 많은 조각을 볼 수 있습니다 때까지 포함 한다. 어떤 부분이 반복된 패스 후 남아 있으면 그냥 두고 바닥에 정착.
참고: 분쇄-거품은 형성 중지 하 고 열망 하 여 제거 하는 동안 볼만 하지 마십시오. - Neurobasal 매체와 다시 중단된 셀 1시 10분 희석 하 고 Trypan Blue 염료와는 hemocytometer를 사용 하 여 가능한 셀. 믹스 50 µ L 희석된 셀의 0.5 mL 원심 분리기 튜브에서 Trypan 블루 솔루션의 50 µ L로. hemacytometer에는 coverslip에 배치 후 10 µ L을 추가 하 고 밝은 명확한 셀 (파란색 셀 죽 었 고 계산 한다).
참고: 단일 E7 광섬유 tectum 범위 1와 5 x 107 셀/mL 사이에서 일반적인 셀 번호. - 천연 접시 ECM에 세포 코팅 MEAs 2200 셀/평방 mm의 밀도. 우리의 나 시스템에 대 한이 MEA 당 약 130000 셀 변환 됩니다. 밤새 셀 첨부 파일 및 neurite 연장 (그림 1)에 대 한 CO2 인큐베이터에서 MEAs을 1 ml는 MEA에 볼륨을가지고 neurobasal 매체를 추가 합니다.
참고: 다른 분야와 전극 숫자의 MEAs에 대 한 다른 셀 밀도 수 시도-일반적으로, 더 높은 세포 조밀도 결과 더 큰 네트워크 활동, 하지만 또한 짧은 살린된 문화. - 다음 날, neurobasal 매체에 10 µ M의 최종 농도에 대우 MEA 단계 1.1에서에서 만든 1 m m 재고에서 BPA를 추가 합니다. 제어 MEA 물 (v/v) 솔루션에서 10% 에탄올의 동일한 볼륨을 추가 합니다.
- 대체 보냈다 매체 neurobasal 매체 포함 10 µ M BPA 매일까지 7 일에서 생체 외에서 (7DIV) 그 후, 매일 네트워크 활동 증가.
참고: 네트워크 활동 증가, 신진 대사 활동 증가 하 고 미디어 변경 내용을 더 자주 필요. 중간을 오렌지를 설정 하지 마십시오.
3. 신경 네트워크 활동을 기록
- 수집의 날, 신선한 neurobasal 매체와 문화 매체를 교환 하 고 녹음 하기 전에 2 h의 최소 CO2 배양 기에는 MEAs를 반환 합니다. 레코딩 소프트웨어를 시작 하 고 37 ° C에 온도 아이콘을 클릭 하 여는 MEA의 온도 설정 (보충 그림 3.1a-b).
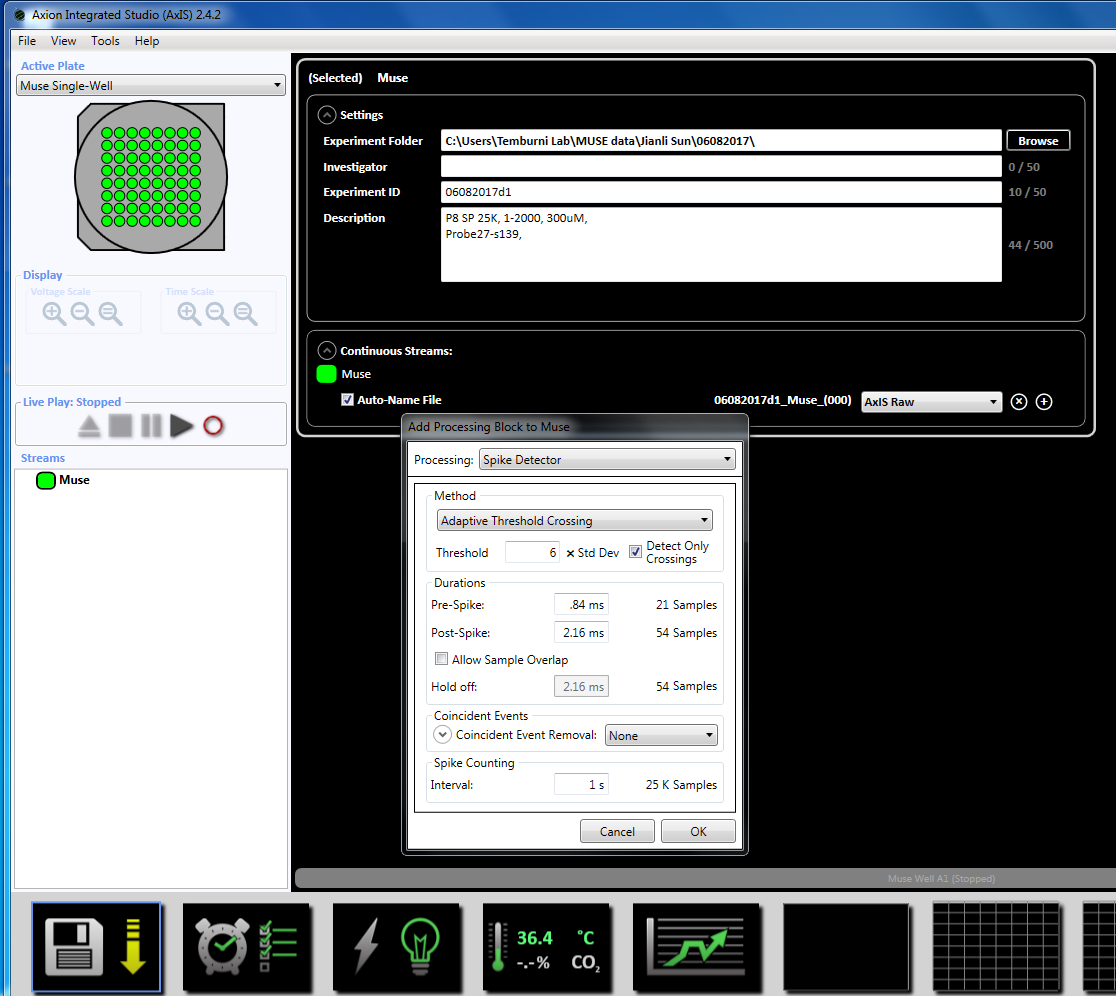
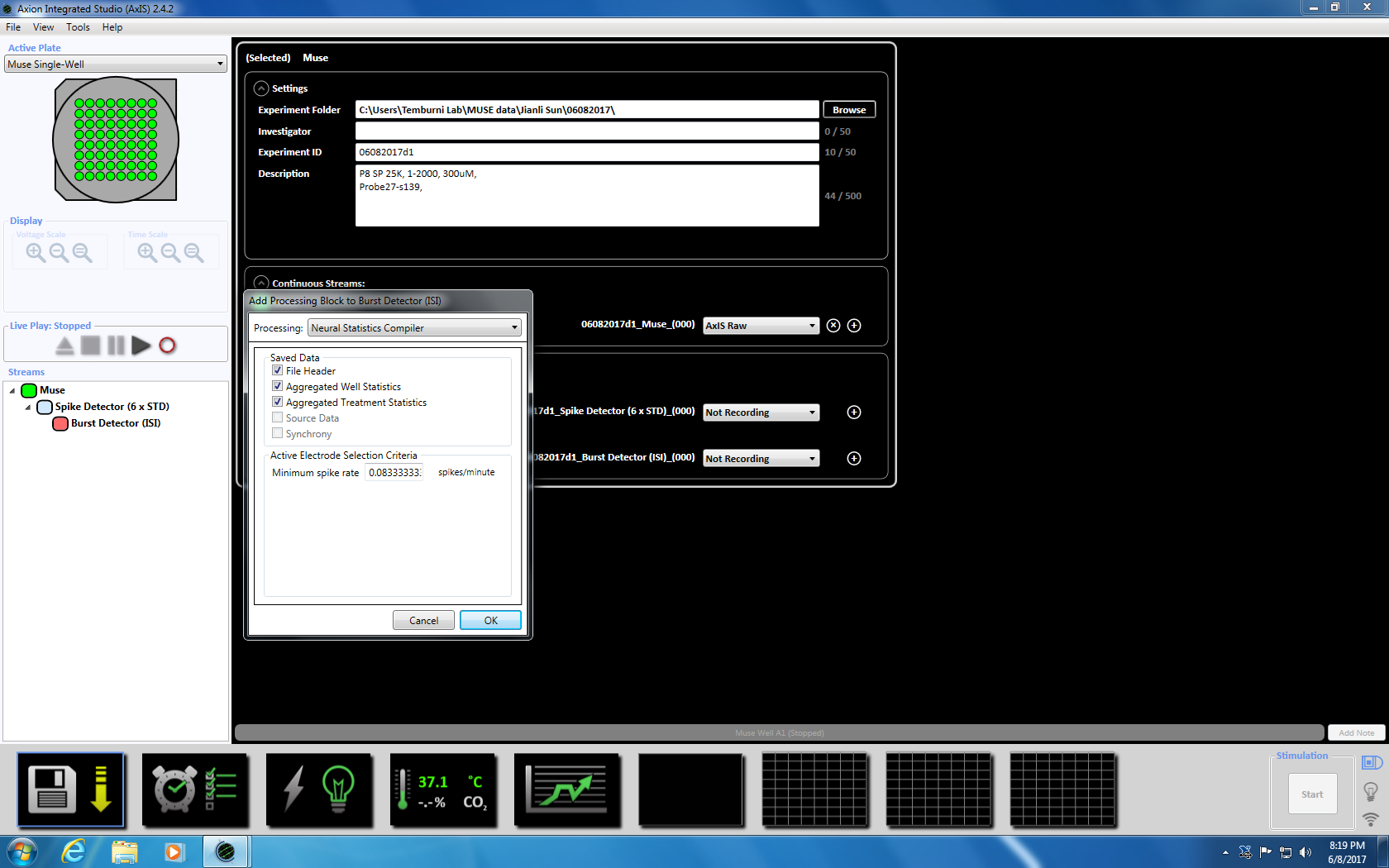
참고: 녹음 도금의 3 일 일찍 시작할 수 있습니다 및 문화 살아 있는 계속할 수 있습니다. - (아래 왼쪽된 창 패널에서 스트림) "뮤 즈" 아이콘을 마우스 오른쪽 단추로 클릭 하 여 인수 매개 변수를 놓으십시오, "추가 처리" 및 "스파이크 탐지기"를 선택 하 고 팝업 창에서에서 확인을 클릭 합니다. "탐지기 스파이크 (6 x 표준)" 파일 이름 아래에 나타납니다 (보충 그림 3.2a-b).
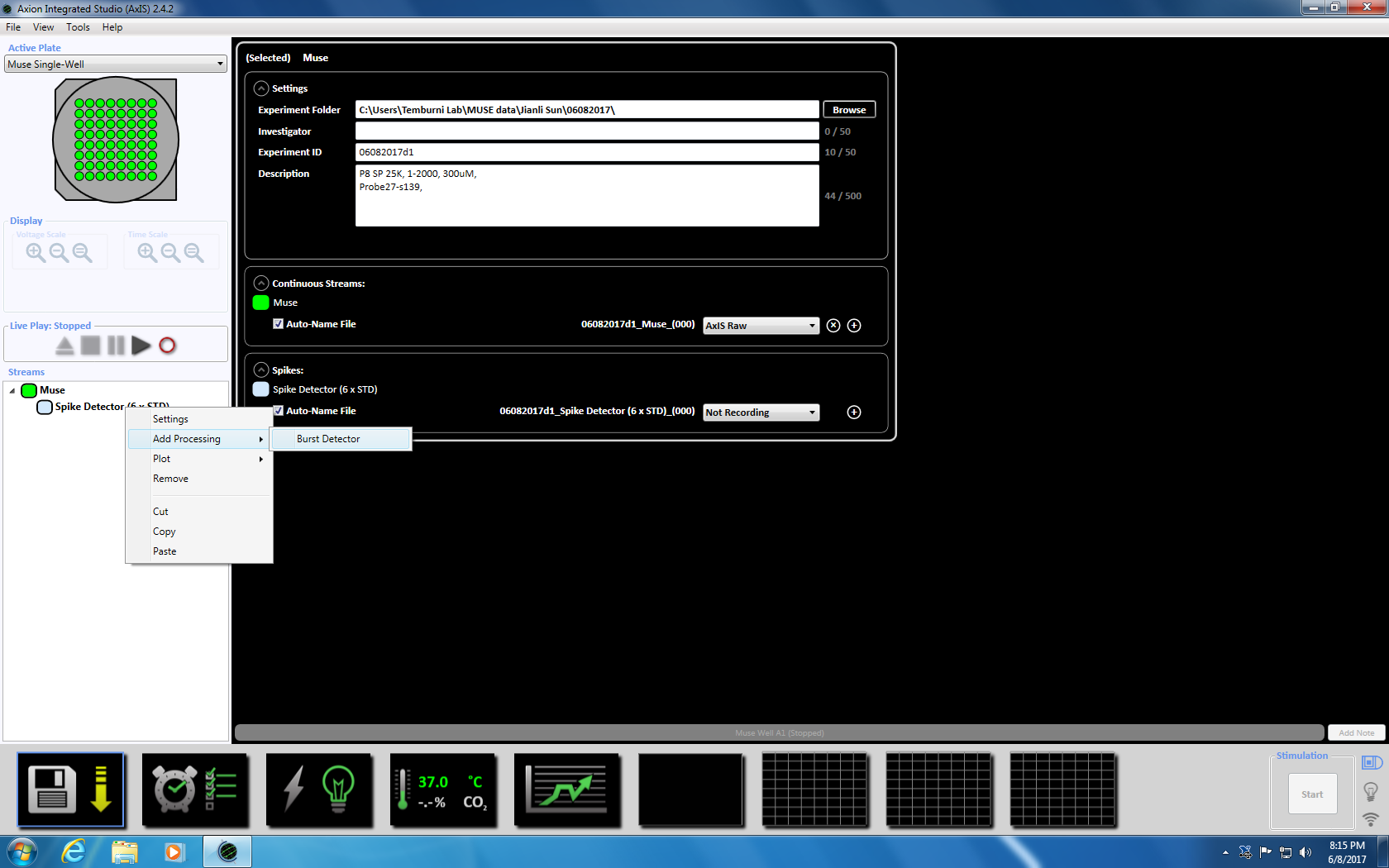
- 다음 스파이크 검출기에 오른쪽 클릭, "추가 처리" 및 "파열 탐지기"를 선택 하 고 팝업 창에서에서 확인을 클릭 합니다. "탐지기 버스트" (ISI) 뮤 즈 아이콘 아래에 나타납니다 (보충 그림 3.3 a-b).
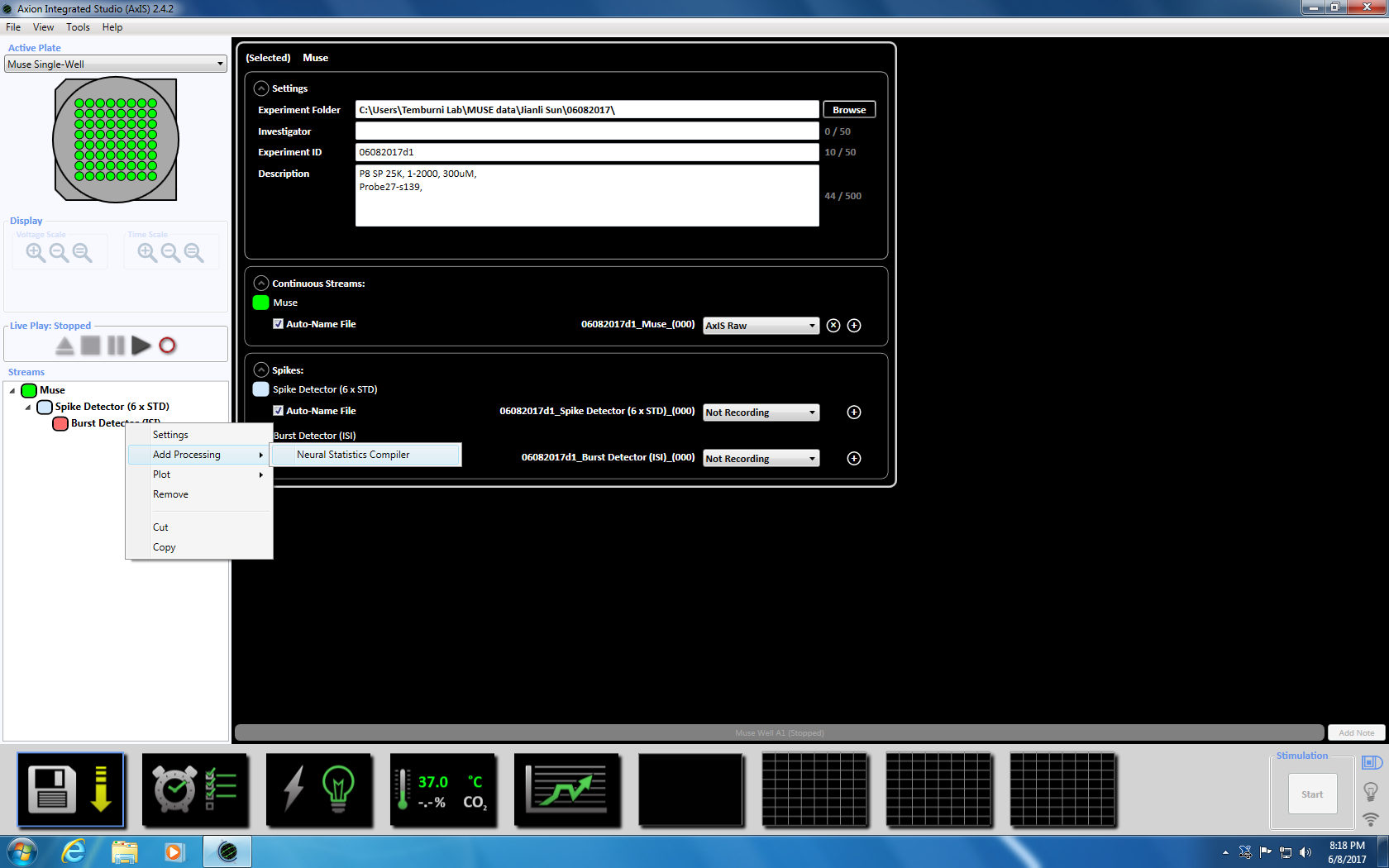
- 다음 버스트 검출기에 오른쪽 클릭, "추가 처리"와 "신경 통계 컴파일러." 선택 팝업 창에서에서 파일 헤더, 잘 통계 집계 및 Synchrony 선택 되었는지 확인 합니다. 클릭 확인입니다. "통계 컴파일러" 아래에 표시 됩니다 (보충 그림 3.4 a-b).
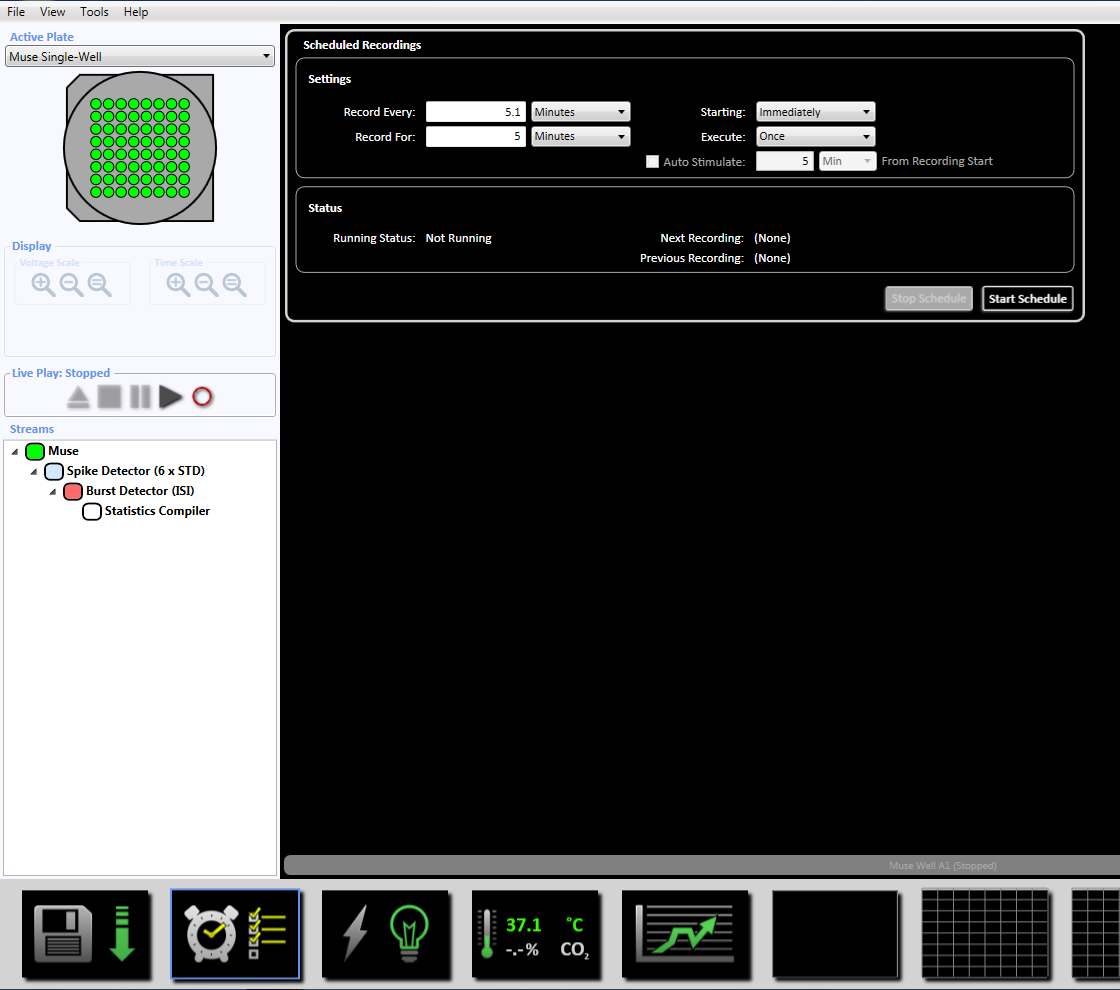
참고:이 적응형 임계값 (6 x 표준 필터), 5, 평균 발사 속도 감지 10에 스파이크의 최소한 100 ms에 간 스파이크 간격을 설정 합니다 synchrony 매개 변수 20 ms에서와 최소 스파이크 속도 분 당 0.083333 스파이크에서 s. - 하단에 시계 아이콘을 클릭 하 여 녹음 시간 5 분 (300, 000 밀리초)에 대 한 예약된 녹화를 설정 합니다. 이 모든 5.1 분을 기록 하 고 5 분 동안 기록에 대 한 설정 섹션에서 정보를 변경 하 여 이루어집니다. 이 즉시 시작 될 것 이다 하 고 한 번 실행 될 것 이다 (보충 그림 3.5).
- MEA의 온도 MEA를 이동 하기 전에 37 ° C를 도달 했습니다 확인 하십시오. 인큐베이터에서는 MEA를 제거 하 고 기록 단위에 배치는 MEA를 잠글. 시작 예정 된 녹음 섹션에서 시작 레코드 또는 "파일 재생" 창에서 기록 아이콘을 클릭 하 여 네트워크 활동을 기록 합니다. 일반적으로, 5 분 (300, 000 밀리초)의 녹화 시간, 충분 하다 비록 더 이상 녹음 최대 10 분의 얻을 수 있습니다.
- 녹음 후 인큐베이터에는 MEA를 반환 합니다.
참고: 합니다 주의 불 임을 유지 하 고는 MEA가 인큐베이터 밖에 시간을 최소화 합니다.
4. 데이터 분석
- 오프 라인 레코딩 소프트웨어를 사용 하 여 원시 데이터의 분석을 수행할 수 있습니다. 파일 풀 다운 메뉴 클릭 하 고 녹음을 엽니다. 원시 데이터 파일 분석을 선택 합니다. 필요한 급격히 매개 변수 캡처 원시 녹음 재생.
- 스파이크의 평균 수를 얻으려면 스파이크 검출기 모듈을 사용 하 고 Synchrony 인덱스를 얻기위한 신경 통계 컴파일러 모듈을 사용 합니다.
- 스파이크 검출기, 버스트 검출기 및 (제 3) 하기 전에 통계 컴파일러 모듈 로드 합니다.
- 스파이크 검출기, 버스트 검출기 및 통계 컴파일러 모듈 창 내에서 풀 다운 메뉴에서 스파이크 목록, 네트워크 버스트 목록 및 고급 통계, 각각 선택 합니다.
- 감독된 폴더에.csv 파일을 만듭니다 있는 데이터 파일을 기록 하려면 기록 단추를 클릭 합니다. 스파이크 매개 변수-스파이크와 Synchrony 인덱스의 총 수-.csv 파일에 기록 됩니다.
참고:이 프로세서 시간과 컴퓨터 리소스를 사용할 수 원시 데이터를 기록 하는 동안 실시간으로 분석 권장 하지 않습니다.
Representative Results
우리 병아리 태아 신경 문화에서 synchrony 개발에 BPA의 효과 검사합니다. 급상승 활동의 동기화는 신경 네트워크 체 외의 정상적인 발전의 지표 이다. 서로 5 ms 이내 두 스파이크 발생 동기 간주 됩니다. 처음 무작위 급격히 활동을 표시 하는 체 외에서 신경 문화-스파이크 무작위로 발생 하 고 간 스파이크 간격 표시 정규 분포. 문화는 성숙, 급격히 활동 더 동기화 된다 고 간 스파이크 간격 일정. 문화의 동기 급격히 활동 Synchrony 인덱스 (SI) 파생 소프트웨어12 기록 사용 하 여 시간 스파이크의 교차 상관 분석으로 quantitated 했다. 우리는 BPA 노출 부정적인 영향을 미치는 네트워크 활동 개발 급격히 활동과 synchrony 인덱스 (그림 2는 및 2b)를 줄임으로써 발견. 요약 하자면, 우리는 배아에 BPA 노출의 해로운 효과 미치는 영향 문화에 병아리 뉴런에서 synchrony 개발을 통해 평가 될 수 있다 신경 기능을 포함 하 여 그것의 발전 나타났습니다.

그림 1: 여자 개 절 개에서 분리, 도금, 및 네트워크 활동의 기록 단계를 묘사 하는 프로토콜의 도식. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 2: (A) 스파이크의 평균 수는 상당히 낮은 BPA 취급 E7 여자 개 문화 제어 문화에 비교 될 때. 스파이크의 평균 수는 레코딩 소프트웨어를 사용 하 여 추출 되었다. 수단 (제어, 14,890 ± 949.0, N = 15, BPA, 5,624 ± 465.9, N = 22) 크게 달랐다 (짝이 없는 t-검정, p < 0.0001). (B) 평균 synchrony 인덱스 (SI)는 크게 낮은 BPA 취급 E7 여자 개 문화 제어 문화에 비교 될 때. SI는 레코딩 소프트웨어 내에서 신경 통계 컴파일러 모듈을 사용 하 여 추출 했다. 시 수단 (제어, 0.5159 ± 0.06547 N = 15, BPA, 0.1140 ± 0.01840 N = 22) 크게 달랐다 (짝이 없는 t-검정, p < 0.0001). 오차 막대는 표준 편차를 묘사. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
Discussion
급속 한 성장 및 embryogenesis 등 개발의 단계는 BPA를 포함 한 다양 일 분 비 방해 화합물의 해로운 효과에 특히 취약 합니다. 우리 생체 외에서 척추 신경 네트워크 모델에 BPA 노출의 효과 연구에 대 한 상세한 프로토콜을 제공 합니다. 이 프로토콜을 사용 하 여, 우리는 BPA synchrony 인덱스 개발 신경 네트워크 (그림 2-b)로 서 스파이크 속도 낮추고 설립.
우리의 방법론은 BPA 노출 초기 embryogenesis 동안 공부 고 다른 EDCs의 효과 연구를 쉽게 적용할 수 있습니다. 여자에서 배아 신경 문화 쥐 또는 마우스를 포함 하 여 다른 척 추가 있는 모델에 비해 상대적으로 쉽게 설정 수 있습니다. 또한, 필요는 없습니다 별도 동물 방-실험실 벤치에 간단한 보육은 이러한 분석을 수행 하기에 충분 한입니다. 우리는 네트워크 활동에 EDC, BPA의 영향을 평가 하기 위한 멀티 전극 시스템 (MEA)의 사용을 설명 합니다. 여기에 설명 된 프로토콜은 다른 시스템에 적용할 수 있습니다. 이 프로토콜의 중요 한 측면은 무 균의 유지 보수. 이 포함 하는 무 균 조건 하에서 배아의 해 부-70% 에탄올과 살 균 행 크의 균형 염 솔루션의 사용 악기를 살 균 하는 것입니다 충분 한. 데이터가 주 기간 동안 수집, 그것은 살 균 처리는 MEAs의 유지에 중요 한입니다. 일단 설립 신경 문화 수 수 경작 장기-최대 3 개월-고 따라서 EDCs 및 신경 네트워킹에 다른 화합물에 장기 노출 공부 하는 데 유용. 이 프로토콜의 또 다른 중요 한 측면은 도금을 해 부에서 시간. 최적의 시간은 trypsin으로 직물의 외피를 포함 하 여 30 분입니다. 더 이상 시간을, 셀의 가난한 생존.
여기 어떤 화학 또는 행동과 신경 기능에 영향을 주는 약물의 평가에 적용할 수 있는 기본 프로토콜을 설명 합니다. 여기, 우리는 m m2당 2200 셀의 셀 밀도 사용 그러나,이 수정할 수 및 다른 셀 밀도 사용 될 수 있다. 일반적으로, 우리는 셀 밀도 증가 네트워크 활동-짧은 시간에 더 많은 스파이크 나타났습니다. 방법 설명, 네트워크 활동에 화학 물질의 효과 사정에 매우 유용 하지만 한계는. 방법의 가장 큰 한계 중 하나입니다 이러한 생체 외에서 문화 2 차원 척추 뇌의 3 차원 구조를 반영 하지 않을 수 있습니다. 이 슬라이스 기록을 사용 하 여 극복할 수 있습니다. 또 다른 대안은 이다, 발전 병아리 태아 치료를 적용할 의미 계란15의 광범위 한 끝에 작은 창, 치료 처방의 끝에 뇌를 해 부하는 vibratome에 두꺼운 부분 고 대 한 MEA에 배치 네트워크 활동을 기록합니다.
우리의 프로토콜 네트워크 활동의 개발에 EDCs의 효력의 검사를 가능 하 게 하 고 이러한 화학 물질의 효과 대 한 기계적으로의 탐험을 제공 한다는 점에서 중요 하다.
Disclosures
저자 들은 아무 경쟁 관심사를 선언 합니다.
Acknowledgments
이 연구는 NSF (HBCU 업 연구 개시 수상, HRD 1401426 및 EPSCoR EPS-0814251)와 NIH (COBRE 1P20GM103653-01A1)에 의해 지원 됩니다. 캔사스에서 델라웨어 INBRE III 6404 친교에 의해 지원 됩니다.
Materials
| Name | Company | Catalog Number | Comments |
| #5 foreceps | Fine Science Tools | 11251-10 | |
| Axion Muse MEA | Axion Biosystems | M64-GL1-30Pt200 | Will be called MEA system in manuscript |
| Axis Software | Axion Biosystems | Will be called recording software in the manuscript | |
| BPA | Sigma-Aldrich | 239658-250g | |
| curved forceps | Fine Science Tools | 11272-50 | |
| EtOH | Sigma-Aldrich | 64-17-5 | |
| Fertilized chicken eggs | from any local farm or Spafas | ||
| HBSS | Fisher | 14170112 | |
| Hemacytometer | Fisher | 02-671-6 | |
| Matrigel Growth Factor Reduced, Phenol Red-Free | BD Biosciences | 356231 | Will be called Extra Cellular Matrix (ECM) in the manuscript |
| Neurobasal medium | BrainBits | Nb4-500 | |
| Neuroexplorer statistical software | Nex Technologies | Neuroexplorer version 5 | |
| Pasteur pipettes | Fisher | 13-678-20A | |
| spring scissors | Fine Science Tools | 15514-12 | |
| Sylgard bottom dissection dishes | Living Systems Instrumentaion | DD-90-S-BLK-3PK | |
| Trypan Blue dye | Fisher | 15-250-061 | |
| Trypsin-EDTA | Fisher | 15400054 |
References
- Anahara, R., Yoshida, M., Toyama, Y., Maekawa, M., Masayuki, K., Ishino, F., Toshimori, K., et al. Estrogen agonists, 17 beta-estradiol, bisphenol A, and diethylstilbestrol, decrease cortactin expression in the mouse testis. Arch. of Histol. Cytol. 69 (2), 101-107 (2006).
- Dodds, C. Synthetic oestrogens. Br. Med. Bull. 11 (2), 131-134 (1955).
- Grignard, E., Lapenna, S., Bremer, S. Weak estrogenic transcriptional activities of Bisphenol A and Bisphenol S. Toxicology In Vitro. 26 (5), 727-731 (2012).
- Herbst, A. L., Ulfelder, H., Poskanzer, D. C. Adenocarcinoma of the vagina: association of maternal stilbestrol therapy with tumor appearance in young women. New Eng. J. Med. 284, 878-881 (1971).
- Jenkins, S., Raghuraman, N., Eltoum, I., Carpenter, M., Russo, J., Lamartiniere, C. A. Oral exposure to bisphenol A increases dimethylbenzanthracene-induced mammary cancer in rats. Environ Health Persp. 117 (6), 910-915 (2009).
- Okada, A., Kai, O. Effects of estradiol-17beta and bisphenol A administered chronically to mice throughout pregnancy and lactation on the male pups' reproductive system. Asian J Androl. 10 (2), 271-276 (2008).
- Palanza, P., Gioiosa, L., Vom Saal, F. S., Parmigiani, S. Effects of developmental exposure to bisphenol A on brain and behavior in mice. Environ. Res. 108 (2), 150-157 (2008).
- Mimoto, A., Fujii, M., Usami, M., Shimamura, M., Hirabayashi, N., Kaneko, T., et al. Identification of an estrogenic hormone receptor in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 364 (4), 883-888 (2007).
- Allard, P., Colaiacovo, M. Bisphenol A impairs the double-strand break repair machinery in the germLine and causes chromosome abnormalities. Proceedings Natl. Acad. Sci. 107 (47), 20405-20410 (2010).
- Mersha, M. D., Patel, B. M., Patel, D., Richardson, B. N., Dhillon, H. S. Effects of BPA and BPS exposure limited to early embryogenesis persist to impair non-associative learning in adults. Behav. Brain Funct. : BBF. 11, 27 (2015).
- Chen, Y., Shu, L., Qiu, Z., Lee, D. Y., Settle, S. J., et al. Exposure to the BPA-Substitute Bisphenol S Causes Unique Alterations of GermLine Function. PLOS Genetics. 12 (7), 1006223 (2016).
- Shein Idelson, M., Ben-Jacob, E., Hanein, Y. Innate Synchronous Oscillations in Freely-Organized Small Neuronal Circuits. PLoS ONE. 5 (12), 14443 (2010).
- Baltz, T., Herzog, A., Voigt, T. Slow Oscillating Population Activity in Developing Cortical Networks: Models and Experimental Results. J. Neurophysiol. 106 (3), 1500-1514 (2011).
- Pettmann, B., Louis, J. C., Sensenbrenner, M. Morphological and Biochemical Maturation of Neurones Cultured in the Absence of Glial Cells. Nature. 281 (5730), 378-380 (1979).
- Zhang, H., Wu, C. Y., Wang, W., Harrington, M. A. Interneuronal Synapses Formed by Motor Neurons Appear to Be Glutamatergic. NeuroReport. 22 (16), 809-813 (2011).
- Paiva, A. R. C., Park, I., Príncipe, J. C. A Comparison of Binless Spike Train Measures. Neural Comp. and Appl. 19 (3), 405-419 (2010).
- Cretu, A., Fotos, J. S., Little, B. W., Galileo, D. S. Human and Rat Glioma Growth, Invasion, and Vascularization in a Novel Chick Embryo Brain Tumor Model. Clin Exp Metastasis. 22, 225-236 (2005).
- Chiappalone, M., et al. Dissociated cortical networks show spontaneously correlated activity patterns during in vitro development. Brain Res. 93, (2006).
- Li, X., et al. Long-term recording on multi-electrode array reveals degraded inhibitory connection in neuronal network development. Biosens Bioelectron. 22 (7), 1538-1543 (2007).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}