

Summary
暴露在环境中的毒素会对发育中的胚胎产生剧烈影响。内分泌干扰的化学物质, 如 bisphenols, 对神经系统有负面影响。在这里, 我们描述了一个协议使用体外脊椎动物 (小鸡胚) 神经元网络模型来研究毒素暴露对早期胚胎的功能影响。
Abstract
双酚, 如双酚 a (BPA) 和双酚 S (BPS), 是聚合剂广泛用于生产塑料和许多日常产品。它们被归类为内分泌干扰化合物 (EDC) 与雌二醇样的性质。长期暴露于 EDCs, 即使在低剂量, 已与各种健康缺陷, 包括癌症, 行为紊乱, 和不孕, 在早期发育期更大的脆弱性。为了研究 BPA 对神经元功能发育的影响, 我们采用了从早期鸡胚脑中提取的体外神经网络作为模型。我们发现, 对 BPA 的接触影响了网络活动的发展, 特别是峰值活动和同步。网络活动的变化是药物或化合物分子靶的关键环节及其对行为结果的影响。多电极阵列越来越成为研究药物对网络活动的影响的有用工具,体外。市场上有几个可用的系统, 尽管电极数量有变化, 电极阵列的类型和质量以及分析软件, 基本的基本原理, 以及获得的数据在整个不同的系统。虽然目前仅限于对二维体外区域性进行分析, 但正在改进这些多边环境协定系统, 以便在脑切片中启用体内网络活动。在这里, 我们提供了一个详细的协议, 胚胎暴露和记录神经元网络活动和同步, 以及具有代表性的结果。
Introduction
4,4′--Isopropylidenediphenol, 通常称为双酚 a 或 BPA, 是一种抗氧化剂添加剂, 发现广泛的聚碳酸酯和环氧树脂为基础的产品, 从热纸, CDs, 粉碎玻璃, 到内涂层饮料罐。虽然 bpa 已经被认为是一种模拟雌激素1的内分泌干扰剂, 但由于每天对 bpa 的偶然接触而导致的不良影响的研究大多在过去10–15年中出现。甚至低水平的 BPA 暴露的后果是最深刻的早期发育期, 包括在胚胎发生期间通过暴露母亲2,3。在子宫内暴露女性胚胎对内分泌干扰化学物质也与增加的疾病易感性从阴道到乳腺癌4,5。动物研究表明, 暴露于 BPA 导致非典型的大脑结构和功能异常表现在行为6。因此, 欧洲和北美洲的大多数监管机构, 包括 FDA, 都禁止在婴儿奶瓶中使用 BPA。为了遵守法规, 许多制造商已改用 44 '-sulfonyldiphenol 或双酚 S (BPS)。根据 bpa 和 bps 是结构类似物的事实, 最近的报告显示, 在雌激素转录7中 bps 的可比效力, 研究这种化合物相对于 BPA 的毒性是很重要的。
在这里, 我们描述了一个协议, 以测试 BPA (和其他 EDCs) 对神经元网络的影响, 使用脊椎动物模型, 小鸡胚胎。体外神经元的培养形成突触接触并产生动作电位 (也称为峰值)。这些文化的峰值活动可以用多电极阵列 (多边环境协定) 系统记录下来。当它们在5毫秒内发生时, 峰值被认为是同步的。最终同步的初始随机峰值活动是开发神经网络8、9的关键特征。同步可以使用各种方法进行测量, 并且在文献9、10、11中描述了几种算法。在我们的分析中, 我们使用由 Paiva 和同事12开发的算法, 它集成到了驱动多边环境协定采集系统的记录软件中。胚胎小鸡神经元的鲁棒峰值活动为研究 BPA 对神经网络活动的影响提供了一个原型, 即13。利用多电极阵列记录峰值活动, 我们观察到 BPA 暴露抑制神经元峰值同步的发展9,14。在这里, 我们提供了一个详细的方法来研究 BPA 暴露在鸡胚神经元的文化, 并具有代表性的结果, 发展的神经元峰值同步性的鸡胚培养。
Protocol
下面给出的协议已经标准化, 以测试 BPA 暴露在 (早期) 胚胎发生期间的影响, 并可能被修改, 用于与 BPS, BPF, 或其他在小鸡胚胎神经元的 EDC。
该议定书遵循特拉华州立大学的体制政策和国家卫生研究院关于禽类胚胎的政策。《议定书》还确认了争端谅解书的材料和化学品安全准则。
1. 材料设置
- 将 BPA (m.w. 228.29) 溶于水中10% 乙醇 (v/v), 使1毫米的库存溶液 (0.23 毫克每毫升10% 乙醇)。在µM 介质中进行后续稀释 (0.1 µM、0.5 µM、1µM、5µM 和 10 neurobasal)。
注意: BPA 是一种环境毒素, 在处理粉末时应注意。 - 在37摄氏度的商用台式鸡蛋孵化器孵化小鸡卵。为 E7 胚胎孵化7天的卵。
- 准备多边环境协定. 用70% 乙醇浸泡3到4小时, 然后在 BSLII 罩中用大约10毫升的不育蒸馏水冲洗三次, 以杀菌所需的多边环境协定。
注: 多边环境协定包含 8 x 8 网格中的64个纳米孔铂电极。电极直径为30µm, 电极间距为200µm。不要使用变性乙醇杀菌, 因为这将导致对多边环境协定的腐蚀。 - 将多边环境协定放在一个无菌容器内, 盖上 (以保持无菌条件), 并移动到工作台顶部孵化器。烘烤至少6小时在55°c。这一步骤对正确的灭菌至关重要, 温度不应超过摄氏60摄氏度。保持不孕, 将含有多边环境协定的碟子移至 BSLII 罩内存放, 直至进一步使用。
注: 我们使用烤箱安全玻璃烤盘与塑料盖子和表面消毒它与70% 乙醇和地方在 BSLII 敞篷为烘干。 - 整除了额外的蜂窝矩阵 ECM 解决方案。解冻的瓶子在4°c 隔夜和冻结100µL 整除数在-20 摄氏度。
注: ECM 已被发运冻结。ECM 不应该被带到室温, 直到它在室温下聚合, 一旦聚合, 就不可能涂上涂层。没有生长因子的 ECM 更倾向于防止由于未知因素造成的额外影响。 - 消毒所有解剖器械 (#5 钳、弯钳、小剪刀、弹簧剪刀) 和自密封材料, 涂上70% 乙醇, 让它们在解剖罩中干燥。
2. 鸡胚神经元培养与网络活动
- 电镀当天, 从-20 °c 冰箱中取出一小瓶 ECM, 用70% 乙醇喷雾, 然后放在冰上。在 BSLII 罩内加入300µL 冷 neurobasal 培养基稀释至25%。
- 使用 P200 pipetteman 将100µL 25% ECM 添加到多边环境协定中心, 注意不要触摸电极, 并立即去除表面上的薄膜。在 CO2孵化器 (37 ° C 和 5% CO2) 中覆盖多边环境协定和位置, 直到准备好对神经元进行板。
- 对 E7 卵的外壳进行消毒 (胚胎日 7, 在37摄氏度孵化7天), 70% 乙醇。将胚胎斩首成一个 Sylgard 的底部解剖皿, 含有冷不育的汉克平衡盐溶液 (HBSS) 而不含钙。切开眼睛, 去掉眼球。使用 #5 细钳和弹簧剪刀在腹侧进行切口, 去除外层皮肤, 露出前脑和视神经顶盖。仔细剥离和去除脑膜膜。将前脑转移到另一个培养皿中, 用弹簧剪刀把它切成2毫米的小块。
注: 脑膜膜切除后, 前脑不应有任何血管附着。如果有的话, 最好是在无菌解剖罩进行解剖, 虽然我们取得了相当好的结果, 当解剖是在罩外进行, 只要仪器和解剖区彻底消毒70% 乙醇. - 使用无菌转移吸管, 将前脑片收集到15毫升离心管中, 并在让前脑片下沉到离心管底部后尽可能 HBSS。
- 添加1毫升0.05% 胰蛋白酶/EDTA (预热到37°c) 和孵化在37°c 15 分钟。
- 使用巴斯德吸管, 小心地去除胰蛋白酶, 不扰乱组织和添加1毫升 neurobasal 培养基。让组织的片断下沉到底部并且去除媒介。再重复一次这个洗涤。这一步是洗掉胰蛋白酶/EDTA。
- 加入2毫升的 neurobasal 培养基, 开始研制过程。研磨包括服用无菌的火抛光的巴斯德吸管, 并通过它轻轻地通过它几次, 直到没有更多的组织被看到。如果在反复传球后留下任何碎片, 只要让它们沉淀到底部即可。
注: 避免起泡, 而研制过程-如果泡沫形成, 停止和消除的愿望。 - 用 neurobasal 培养基稀释重悬浮细胞 1:10, 用台盼蓝染料和 hemocytometer 计数活细胞。在0.5 毫升离心管中混合50µL 的稀释细胞与50µL 的台盼蓝溶液。添加10µL 到 hemacytometer 后放置在盖玻片和计数明亮的透明细胞 (蓝色细胞死了, 不应该计数)。
注意: 从单个 E7 光学顶盖的典型细胞数介于1和 5 x 10 7 细胞/mL 之间. - 将 ECM 上的离解细胞涂在2200细胞/平方毫米的密度上。对于我们的多边环境协定系统, 这相当于每个多边环境协定的大约13万个单元。添加 neurobasal 介质以将多边环境协定中的体积增加到1毫升, 并在 CO2孵化器中过夜以实现单元格附件和突起扩展 (图 1)。
注意: 对于不同区域和电极数的多边环境, 可以尝试不同的细胞密度--一般情况下, 较高的细胞密度会导致网络活动的增加, 但也会导致更短的生存文化。 - 第二天, 添加 BPA 到最后浓度的10µM 在 neurobasal 培养基从1毫米的股票在步骤1.1 所处理的多边协定。对控制多边环境协定, 在水 (v/v) 溶液中加入相同体积的10% 乙醇。
- 每隔一天用含有10µM BPA 的 neurobasal 培养基替换所用介质, 直到7天的体外(7DIV) 和此后的每一天, 随着网络活动的增加。
注意: 随着网络活动的增加, 新陈代谢活动增加, 媒体的变化需要更加频繁。不要让中号变成橙色。
3. 记录神经元网络活动
- 在购置日, 在录制前将培养基与新鲜的 neurobasal 培养基交换, 并将多边环境协定返回 CO2孵化器, 至少2小时。通过单击温度图标 (补充图 3.1a-b), 启动录制软件并将多边环境协定的温度设置为37°c。
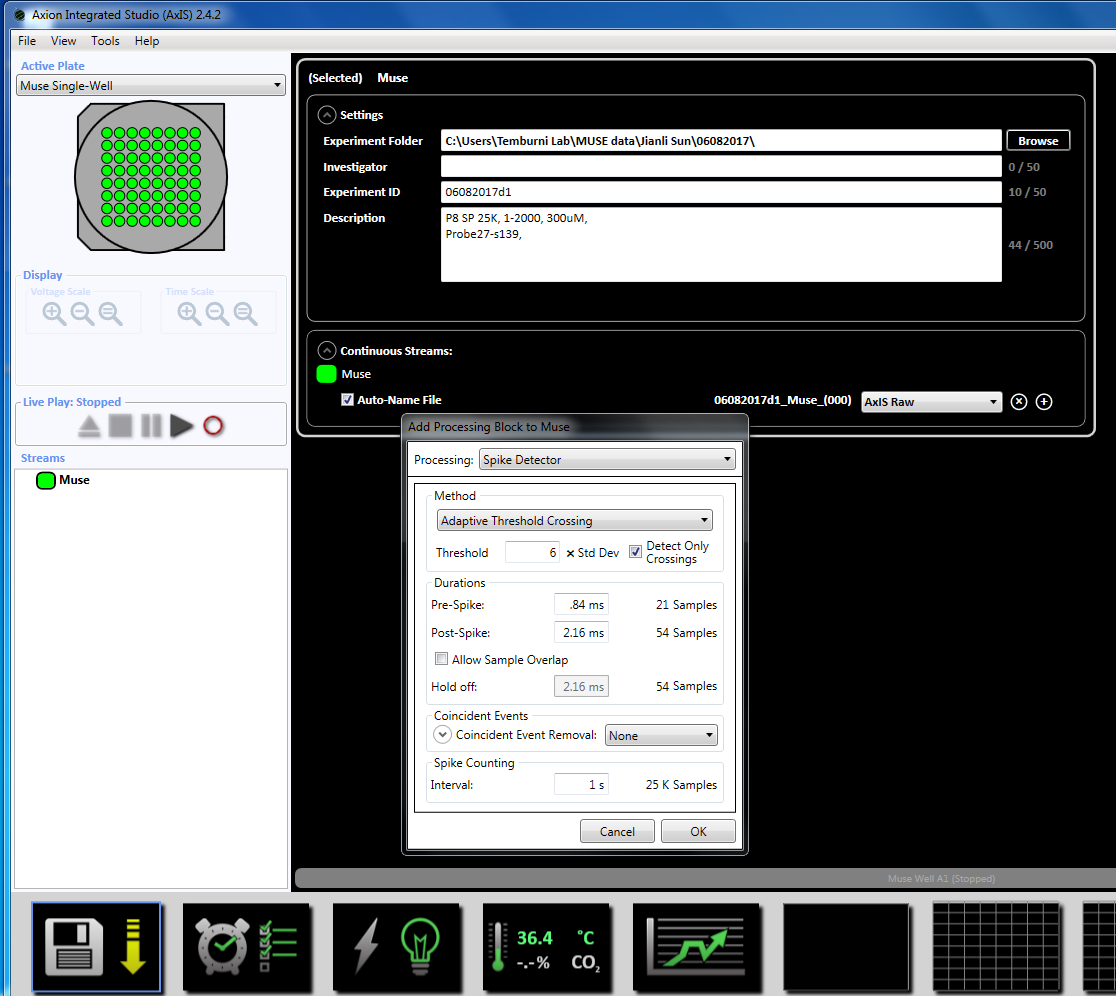
注: 记录可以早在3天开始电镀, 只要文化还活着就可以继续。 - 通过右键单击 "缪斯" 图标 (在左侧窗口面板中的 "流" 下), 然后选择 "添加处理" 和 "尖峰检测器", 然后在弹出窗口中单击 "确定", 设置采集参数。"尖峰探测器 (6 x STD)" 将出现在文件名下面 (补充图 3.2a b)。
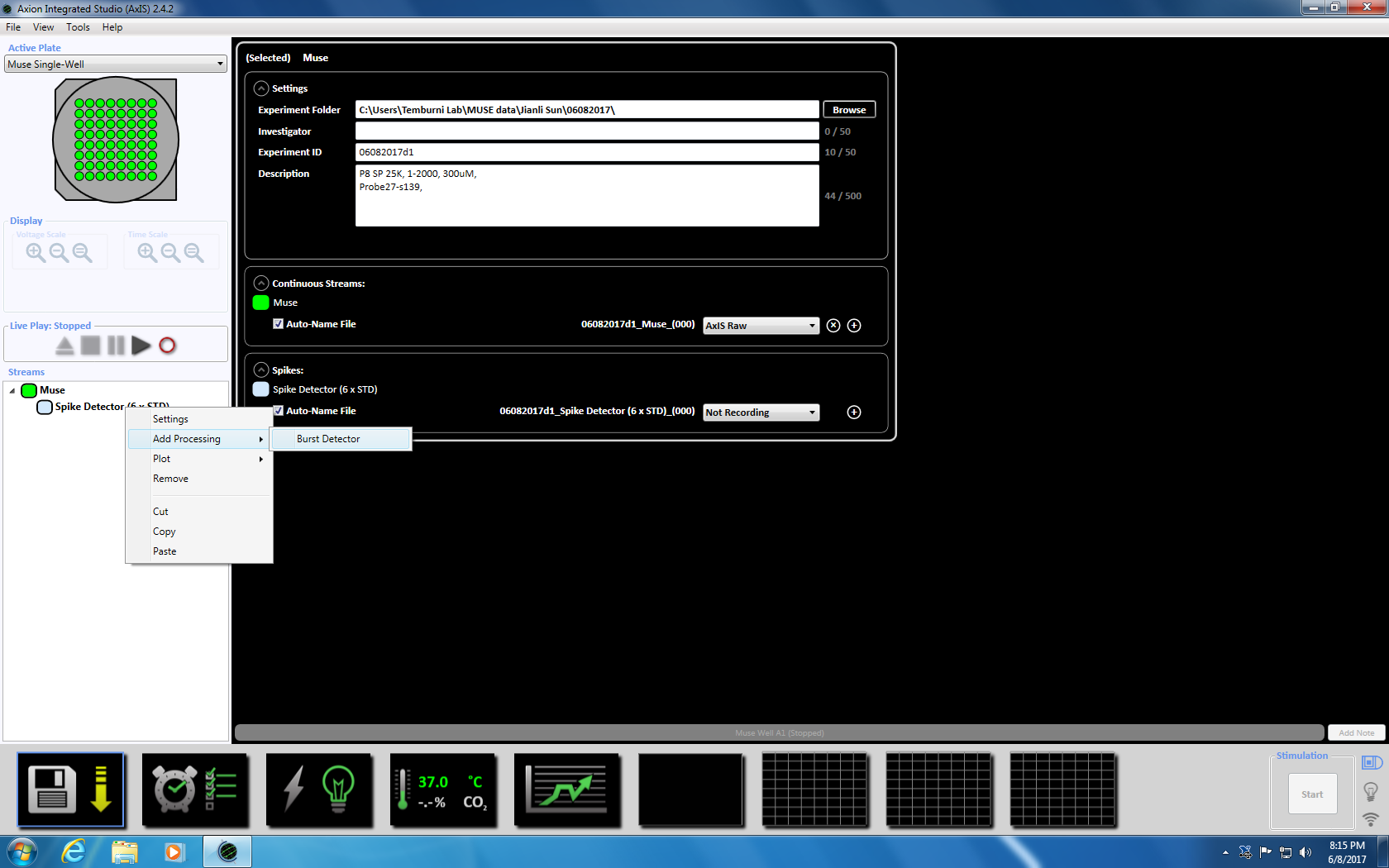
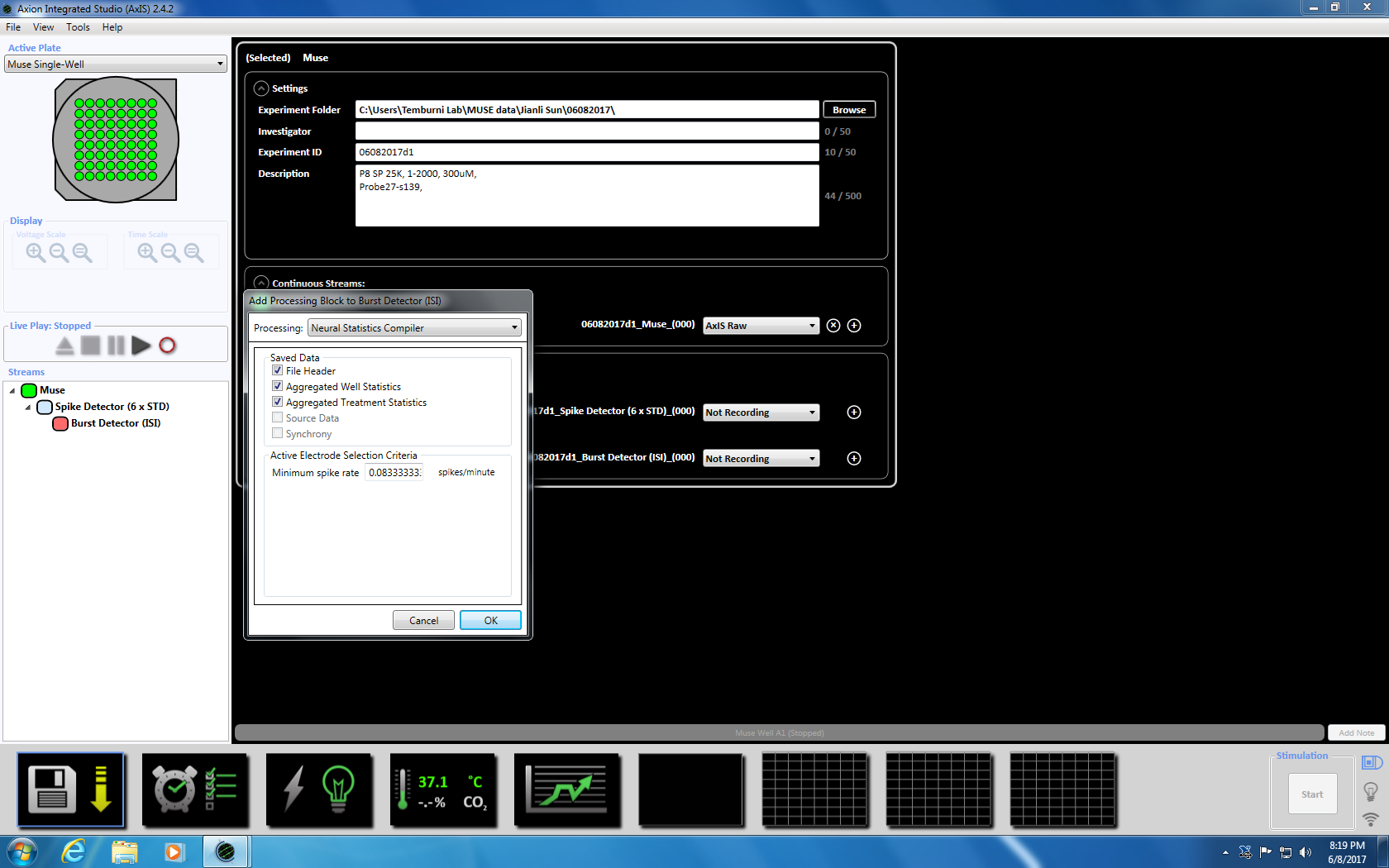
- 下一步右击尖峰探测器, 选择 "添加处理" 和 "突发检测器", 并在弹出窗口中单击 "确定"。"爆裂探测器" (ISI) 将出现在缪斯图标的下面 (补充图 3.3a-b)。
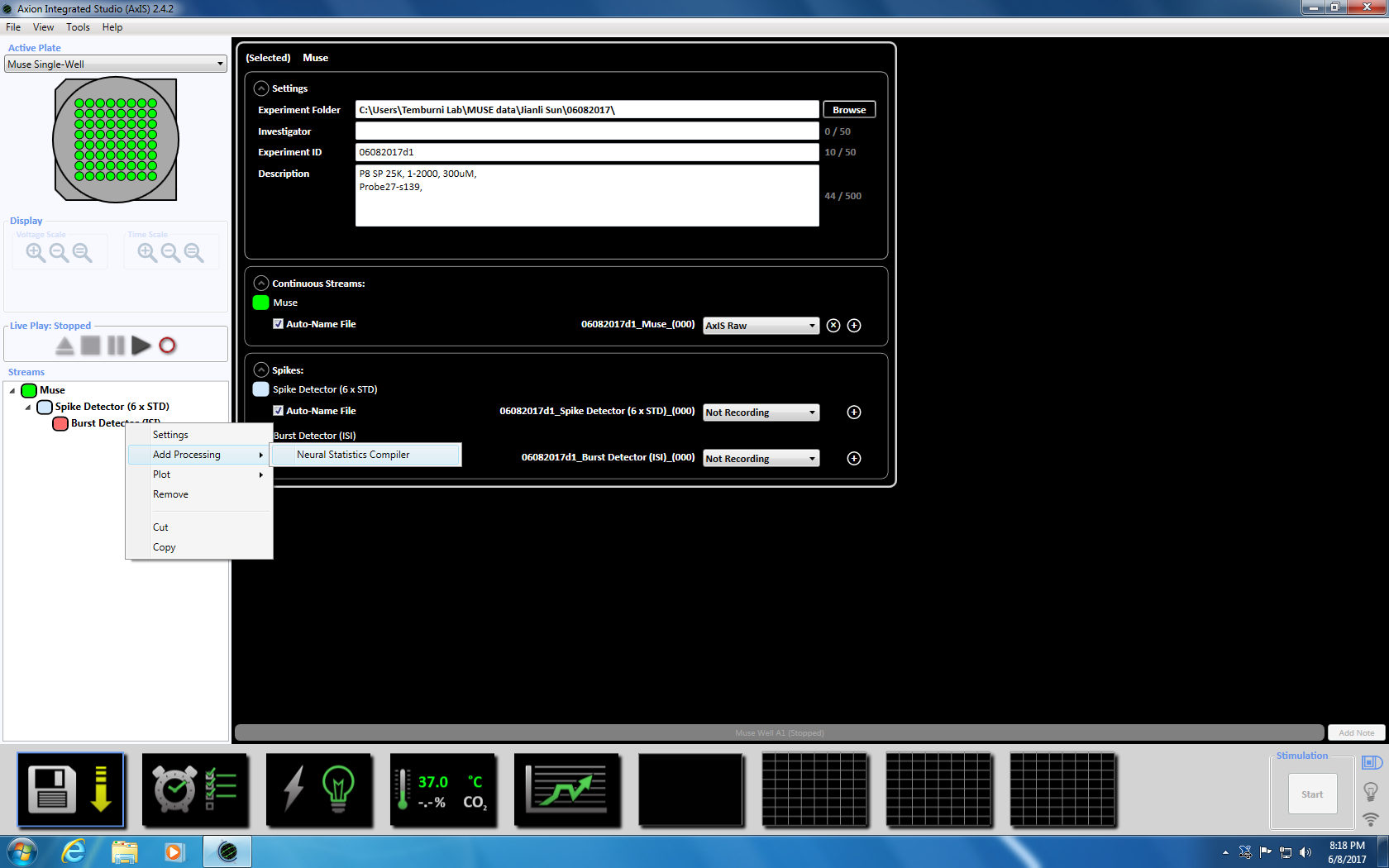
- 下一步右击突发检测器, 选择 "添加处理" 和 "神经统计编译器"。在弹出窗口中, 确保选择了文件头、聚合井统计信息和同步。单击 "确定"。"统计编译器" 将出现在下面 (补充图 3.4a-b)。
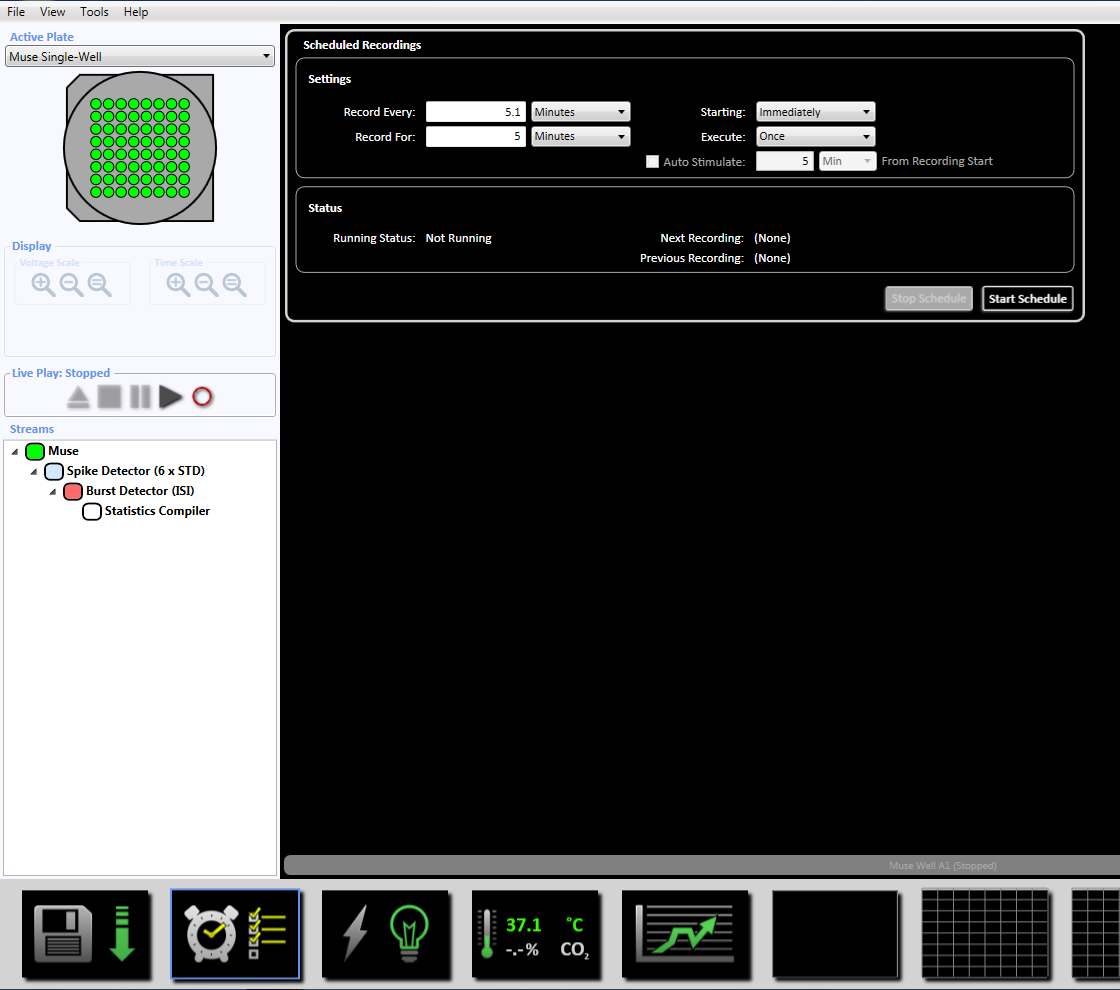
注意: 这将设置自适应阈值交叉 (6x STD 过滤器), 100 毫秒之间的穗间间隔, 最小峰值为 5, 平均发射率检测在十年代, 同步参数为20毫秒, 最小穗速率为每分钟0.083333 个峰值。 - 通过单击底部的时钟图标, 设置录制时间为5分钟 (30万毫秒) 的定时录制。这是通过将设置节中的信息更改为每5.1 分钟记录一次并记录5分钟来实现的。这将立即启动, 并将执行一次 (补充图 3.5)。
- 在移动多边环境协定之前, 确保多边环境协定的温度达到37摄氏度。从孵化器中移除多边环境协定, 将其放在记录单元上, 并锁定协定。通过单击 "文件播放" 窗口中 "计划录制" 部分或 "记录" 图标中的 "开始" 记录, 开始记录网络活动。通常情况下, 记录时间为5分钟 (30万毫秒) 是足够的, 虽然可以获得长达10分钟的更长的记录。
- 录音后, 将多边环境协定归还孵化器。
注意: 必须注意保持不孕, 并尽量减少环境协定在孵化器之外的时间。
4. 数据分析
- 对原始数据的分析可以使用记录软件离线进行。单击 "文件" 下拉菜单并打开录制。选择要分析的原始数据文件。重放原始录制以捕获所需的峰值参数。
- 为了获得峰值的平均数, 使用尖峰检测模块和获取同步索引, 使用神经统计编译器模块。
- 像以前一样加载峰值检测器、爆裂探测器和统计编译模块 (第3节)。
- 从尖峰检测器、爆裂探测器和统计编译器模块窗口中的下拉菜单中, 分别选择 "尖峰列表"、"网络突发列表" 和 "高级度量"。
- 单击 "记录" 按钮开始录制将在定向文件夹中创建. csv 文件的数据文件。峰值参数 (峰值和同步索引的总数) 将记录在. csv 文件中。
注意: 不建议在记录原始数据时进行实时分析, 因为这可能会消耗处理器时间和计算资源。
Representative Results
我们研究了 BPA 对鸡胚神经元培养同步发育的影响。峰值活动同步是神经网络体外正常发育的指标。当两个峰值发生在5毫秒之间时, 它们被认为是同步的。体外神经元培养最初显示随机峰值活动-峰值随机发生, 穗间间隔显示正常分布。随着培养的成熟, 峰值活动变得更加同步, 穗间间隔是恒定的。通过使用记录软件12生成同步指数 (SI), 量化的同步峰值活动, 通过交叉相关分析, 对某一区域性进行了脉冲峰值时间的相互相关分析。我们发现 BPA 暴露通过减少峰值活动和同步索引 (图 2a和2b) 对网络活动开发产生不利影响。总之, 我们已经表明, BPA 暴露对胚胎的有害影响它的发展, 包括神经元功能, 可以评估通过同步发展的小鸡神经元的文化。

图 1:描述从小鸡前脑解剖到离解、电镀和记录网络活动步骤的协议示意图.请单击此处查看此图的较大版本.

图 2:(A)与控制文化相比, BPA 治疗 E7 鸡前脑培养物的平均穗数明显低。使用记录软件提取峰值的平均数量。方法 (控制, 14890 949.0, N = 15 和 BPA, 5624 @ 465.9, N = 22) 有显著差异 (配对 t 检验, p < 0.0001)。(B)与控制文化相比, BPA 治疗 E7 鸡前脑培养物的平均同步指数 (SI) 显著降低。使用记录软件中的神经统计编译器模块提取 SI。SI 手段 (控制, 0.5159 0.06547 N = 15 和 BPA, 0.1140 @ 0.01840 N = 22) 是明显地不同的 (未配对的 t 试验, p < 0.0001)。误差线描述标准偏差。请单击此处查看此图的较大版本.
Discussion
快速生长和发育的阶段, 如胚胎发生特别容易受到各种内分泌干扰化合物, 包括 BPA 的有害影响。我们提供详细的协议, 以研究 BPA 暴露在一个体外脊椎动物神经网络模型的影响。通过使用该协议, 我们建立了 BPA 降低了发展中神经网络 (图 2a b) 的峰值速率和同步索引。
我们的方法是设计来研究 BPA 暴露在早期胚胎发生, 可以很容易地适应研究其他 EDCs 的影响。与其他脊椎动物模型 (包括老鼠或老鼠) 相比, 小鸡的胚胎神经元培养相对容易建立。此外, 没有要求单独的动物室-一个简单的孵化器在实验室的长凳上足以进行这些化验。我们描述了使用多电极系统 (多边环境协定), 以评估的影响, 该公司, BPA, 对网络活动。此处描述的协议可应用于其他系统。这项议定书的一个关键方面是维持不孕。这包括在无菌条件下对胚胎进行解剖--用70% 乙醇杀菌的仪器和使用无菌汉克的平衡盐溶液是足够的。由于这些数据是在几周内收集的, 因此保持对多边环境协定的无菌处理至关重要。曾经建立的神经元培养可以长期培养到三个月, 因此对于研究神经网络上的 EDCs 和其他化合物的长期接触是有用的。该协议的另一个重要方面是从解剖到电镀的时间。最佳时间为30分钟, 包括用胰蛋白酶培养组织。所花的时间越长, 细胞的存活率就越差。
我们这里描述了一个基本的协议, 可用于评估任何影响行为和神经功能的化学药品或药物。在这里, 我们使用了2200个细胞密度每毫米2;然而, 这可以修改和其他细胞密度可以使用。总的来说, 我们发现, 细胞密度的增加增加了网络活动-在较短的时间内更多的峰值。所描述的方法虽然在评估化学品对网络活动的影响方面非常有用, 但也有局限性。该方法的最大限制之一是这些体外培养是二维的, 可能无法反映脊椎动物大脑的三维结构。这可以通过使用切片录制来克服。另一种选择是将治疗方法应用于发育中的小鸡胚胎, 方法是在卵子的宽端切开一个小窗口, 在治疗方案的末尾解剖大脑, 在 vibratome 上做厚的切片, 并将其放置到多边环境协定中, 以记录网络活动。
我们的议定书具有重要意义, 因为它能够检查 EDCs 对网络活动发展的影响, 并为这些化学物质的作用提供了一个机械基础的探索。
Disclosures
作者声明他们没有竞争的兴趣。
Acknowledgments
这项研究得到了 NSF (HBCU 研究发起奖, 人力资源开发1401426和 EPSCoR EPS-0814251) 和 NIH (COBRE 1P20GM103653-01A1) 的支持。K.S. 得到特拉华 INBRE-III. 6404 的奖学金支持。
Materials
| Name | Company | Catalog Number | Comments |
| #5 foreceps | Fine Science Tools | 11251-10 | |
| Axion Muse MEA | Axion Biosystems | M64-GL1-30Pt200 | Will be called MEA system in manuscript |
| Axis Software | Axion Biosystems | Will be called recording software in the manuscript | |
| BPA | Sigma-Aldrich | 239658-250g | |
| curved forceps | Fine Science Tools | 11272-50 | |
| EtOH | Sigma-Aldrich | 64-17-5 | |
| Fertilized chicken eggs | from any local farm or Spafas | ||
| HBSS | Fisher | 14170112 | |
| Hemacytometer | Fisher | 02-671-6 | |
| Matrigel Growth Factor Reduced, Phenol Red-Free | BD Biosciences | 356231 | Will be called Extra Cellular Matrix (ECM) in the manuscript |
| Neurobasal medium | BrainBits | Nb4-500 | |
| Neuroexplorer statistical software | Nex Technologies | Neuroexplorer version 5 | |
| Pasteur pipettes | Fisher | 13-678-20A | |
| spring scissors | Fine Science Tools | 15514-12 | |
| Sylgard bottom dissection dishes | Living Systems Instrumentaion | DD-90-S-BLK-3PK | |
| Trypan Blue dye | Fisher | 15-250-061 | |
| Trypsin-EDTA | Fisher | 15400054 |
References
- Anahara, R., Yoshida, M., Toyama, Y., Maekawa, M., Masayuki, K., Ishino, F., Toshimori, K., et al. Estrogen agonists, 17 beta-estradiol, bisphenol A, and diethylstilbestrol, decrease cortactin expression in the mouse testis. Arch. of Histol. Cytol. 69 (2), 101-107 (2006).
- Dodds, C. Synthetic oestrogens. Br. Med. Bull. 11 (2), 131-134 (1955).
- Grignard, E., Lapenna, S., Bremer, S. Weak estrogenic transcriptional activities of Bisphenol A and Bisphenol S. Toxicology In Vitro. 26 (5), 727-731 (2012).
- Herbst, A. L., Ulfelder, H., Poskanzer, D. C. Adenocarcinoma of the vagina: association of maternal stilbestrol therapy with tumor appearance in young women. New Eng. J. Med. 284, 878-881 (1971).
- Jenkins, S., Raghuraman, N., Eltoum, I., Carpenter, M., Russo, J., Lamartiniere, C. A. Oral exposure to bisphenol A increases dimethylbenzanthracene-induced mammary cancer in rats. Environ Health Persp. 117 (6), 910-915 (2009).
- Okada, A., Kai, O. Effects of estradiol-17beta and bisphenol A administered chronically to mice throughout pregnancy and lactation on the male pups' reproductive system. Asian J Androl. 10 (2), 271-276 (2008).
- Palanza, P., Gioiosa, L., Vom Saal, F. S., Parmigiani, S. Effects of developmental exposure to bisphenol A on brain and behavior in mice. Environ. Res. 108 (2), 150-157 (2008).
- Mimoto, A., Fujii, M., Usami, M., Shimamura, M., Hirabayashi, N., Kaneko, T., et al. Identification of an estrogenic hormone receptor in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 364 (4), 883-888 (2007).
- Allard, P., Colaiacovo, M. Bisphenol A impairs the double-strand break repair machinery in the germLine and causes chromosome abnormalities. Proceedings Natl. Acad. Sci. 107 (47), 20405-20410 (2010).
- Mersha, M. D., Patel, B. M., Patel, D., Richardson, B. N., Dhillon, H. S. Effects of BPA and BPS exposure limited to early embryogenesis persist to impair non-associative learning in adults. Behav. Brain Funct. : BBF. 11, 27 (2015).
- Chen, Y., Shu, L., Qiu, Z., Lee, D. Y., Settle, S. J., et al. Exposure to the BPA-Substitute Bisphenol S Causes Unique Alterations of GermLine Function. PLOS Genetics. 12 (7), 1006223 (2016).
- Shein Idelson, M., Ben-Jacob, E., Hanein, Y. Innate Synchronous Oscillations in Freely-Organized Small Neuronal Circuits. PLoS ONE. 5 (12), 14443 (2010).
- Baltz, T., Herzog, A., Voigt, T. Slow Oscillating Population Activity in Developing Cortical Networks: Models and Experimental Results. J. Neurophysiol. 106 (3), 1500-1514 (2011).
- Pettmann, B., Louis, J. C., Sensenbrenner, M. Morphological and Biochemical Maturation of Neurones Cultured in the Absence of Glial Cells. Nature. 281 (5730), 378-380 (1979).
- Zhang, H., Wu, C. Y., Wang, W., Harrington, M. A. Interneuronal Synapses Formed by Motor Neurons Appear to Be Glutamatergic. NeuroReport. 22 (16), 809-813 (2011).
- Paiva, A. R. C., Park, I., Príncipe, J. C. A Comparison of Binless Spike Train Measures. Neural Comp. and Appl. 19 (3), 405-419 (2010).
- Cretu, A., Fotos, J. S., Little, B. W., Galileo, D. S. Human and Rat Glioma Growth, Invasion, and Vascularization in a Novel Chick Embryo Brain Tumor Model. Clin Exp Metastasis. 22, 225-236 (2005).
- Chiappalone, M., et al. Dissociated cortical networks show spontaneously correlated activity patterns during in vitro development. Brain Res. 93, (2006).
- Li, X., et al. Long-term recording on multi-electrode array reveals degraded inhibitory connection in neuronal network development. Biosens Bioelectron. 22 (7), 1538-1543 (2007).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}