

Summary
Udsættelse for miljøområdet toksiner kan akut påvirker udviklingslandene embryoner. Hormonsystemforstyrrende kemikalier såsom uvæsentligt er kendt for at påvirke nervesystemet. Her beskriver vi en protokol bruger en in vitro- hvirveldyr (kylling embryo) neuronal netværksmodel til at studere den funktionelle effekt af toksin eksponering på tidlige embryoner.
Abstract
BIS-phenoler, som bis-phenol A (BPA) og bis-phenol-S (BPS), polymeriserende agenter almindeligt anvendt i produktion af plast og mange dagligdags produkter. De klassificeres som endokrine forstyrre forbindelser (EDC) med østradiol-lignende egenskaber. Langvarig eksponering for EDC, selv ved lave doser, har været forbundet med forskellige sundhed defekter, herunder kræft, adfærdsmæssige forstyrrelser og barnløshed, med større sårbarhed i tidlige udviklingsmæssige perioder. For at studere virkningerne af BPA på udviklingen af neuronal funktion, brugte vi en in vitro- neuronal netværk stammer fra den tidlige chick embryonale hjerne som en model. Vi fandt, at eksponering for BPA påvirket udviklingen af netværksaktivitet, specielt spiking aktivitet og synkronisering. En ændring i netværksaktivitet er den afgørende sammenhæng mellem den molekylære mål af et lægemiddel eller sammensatte og dens virkning på adfærdsmæssige resultatet. Multi elektrode arrays i stigende grad nyttige værktøjer til at studere virkningerne af narkotika på netværk aktivitet in vitro. Der er flere systemer på markedet, og selv om der er variationer i antallet af elektroder, type og kvalitet af elektrode-array og analyse software, de grundlæggende principper, og data indhentet er den samme på tværs af de forskellige systemer. Selv i øjeblikket begrænset til analyse af to-dimensionelle i vitro kulturer, forbedres disse MEA systemer for at aktivere i vivo netværksaktivitet i hjernen skiver. Her give vi en detaljeret protokol for embryonale eksponering og optagelse neuronal netværksaktivitet og synchrony, sammen med repræsentative resultater.
Introduction
4, 4′-Isopropylidendiphenol, der almindeligvis omtales som Bisphenol-A eller BPA, findes en anti-oxidant tilsætningsstof i en bred vifte af polycarbonat og epoxy harpiks baseret produkter fra termisk papir, cd'er og splintres bevis glas, på indersiden belægning af drikkevarer dåser. Selv om BPA har været kendt for at være hormonforstyrrende forstyrre agent der efterligner østrogen1, har undersøgelser på dens skadelige virkninger som følge af hverdagens casual eksponering for BPA for det meste kommer ud i de sidste 10 – 15 år. Konsekvenserne af selv lave niveauer af BPA eksponering er mest dybtgående i tidlige udviklingsmæssige perioder, herunder under embryogenese gennem eksponering af mødre2,3. In utero eksponering af kvindelige embryoner til hormonforstyrrende kemikalier har også været forbundet med øget sygdom modtagelighed fra vaginal til breast cancer4,5. Dyreforsøg har vist eksponering for BPA fører til atypiske hjernens struktur og funktionelle abnormiteter manifesteret i adfærd6. Som følge heraf har været forbudt brugen af BPA i spædbarn sutteflasker af mest reguleringsorganer i Europa og Nordamerika, herunder FDA. For at overholde regler, har mange producenter skiftet til 4, 4'-sulfonyldiphenol eller bisphenol-S (BPS). Baseret på det faktum, at BPA og BPS er strukturelle analogstoffer deraf, og at de seneste rapporter viste sammenlignelige potens af BPS i østrogene transskription7, er det vigtigt at undersøge toksiciteten af denne sammensatte forhold til BPA.
Her beskriver vi en protokol for at teste effekten af BPA (og andre EDC) på netværk af neuroner ved hjælp af en hvirveldyr model, kylling embryo. In vitro kulturer af neuroner udgør synaptic kontakter og generere handling potentialer (også kaldet pigge). Spiking aktiviteten af disse kulturer kan registreres ved hjælp af multi elektrode array (MEA) systemer. Spikes er anses for at være i synchrony, når de opstår inden for 5 ms af hinanden. Indledende tilfældige spiking aktivitet, der i sidste ende synkroniserer er et centralt element for at udvikle neuronal netværk8,9. Synchrony kan måles ved hjælp af forskellige metoder og flere algoritmer er beskrevet i litteraturen9,10,11. I vores analyser bruger vi en algoritme udviklet af Paiva og kollegaer12 , som er integreret i den optagelse software, som drev MEA erhvervelse system. Den robuste spiking aktivitet af embryonale chick neuroner giver en prototype for at studere effekten af BPA på neurale netværk aktivitet13. Brug af multi elektrode arrays til at optage spiking aktivitet, konstaterede vi, at BPA eksponering hæmmer udviklingen af neuronal spiking synchrony9,14. Her give vi en detaljeret metode til at studere BPA eksponering på kylling embryo neuroner i kultur sammen med repræsentative resultater på udviklingen af neuronal spiking synchrony i chick embryonale kulturer.
Protocol
Protokollen nedenfor er blevet standardiseret for at afprøve virkningerne af BPA eksponering under (tidlig) embryogenese, og kan blive modificeret til brug med BPS, BPF eller andre EDC i chick embryonale neuroner.
Denne protokol følger de institutionelle politikker i Delaware State University og NIH politik for aviær embryoner. Protokollen er også i bekræftelse med DSUS materiale og kemisk sikkerhedsretningslinjer.
1. materielle Setup
- Opløse BPA (m.w. 228.29) i 10% ethanol (v/v) i vand for at lave en 1 mM stamopløsning (0,23 mg / mL 10% ethanol). Gøre efterfølgende fortyndinger (0,1 µM, 0,5 µM, 1 µM, 5 µM og 10 µM) i neurobasal medium.
Forsigtig: BPA er en miljømæssig toksin og bør udvises forsigtighed ved håndtering af pulveret. - Inkuber kylling æg i en kommercielt tilgængelig bordplade æg inkubator ved 37 ° C. Inkuber æg i 7 dage for E7 embryoner.
- Forberede ventetid Sterilize det nødvendige antal MEAs ved opblødning i 70% ethanol i 3-4 timer og derefter skylles tre gange i ca. 10 mL sterilt, destilleret vand i BSLII hætte.
Bemærk: MEAs indeholder 64 nanoporous platinum elektroder arrangeret i en 8 x 8-gitter. Elektrode diameter er 30 µm og afstanden mellem elektroderne er 200 µm. Brug ikke denatureret ethanol til sterilisering af multilaterale miljøaftaler, da dette vil føre til korrosion af MEA. - Placer MEAs inde i en steril beholder med låg (for at opretholde sterile forhold) og flytte til en bordplade inkubator. Bages i mindst 6 timer ved 55 ° C. Dette trin er afgørende for korrekt sterilisation og temperaturen bør ikke overstige 60 ° C. Vedligeholdelse af steriliteten, flytte fadet indeholdende MEAs BSLII hætte til opbevaring indtil videre brug.
Bemærk: Vi bruger en ovn sikker glas bagning parabol med plastlåg og overflade sterilisere det med 70% ethanol og sted i BSLII hætte til tørring. - Alikvot ekstra cellulære matrix ECM-løsningen. Tø hætteglas ved 4 ° C natten over og fryse 100 µL alikvoter ved-20 ° C.
Bemærk: ECM leveres frosset. ECM bør ikke bringes til stuetemperatur indtil klar til pels som det polymeriserer ved stuetemperatur og når polymeriserede, det er umuligt at pels. ECM uden vækstfaktorer er foretrukket at forhindre yderligere virkninger på grund af ukendte faktorer. - Sterilisere alle dissektion instrumenter (Dumont #5 pincet, buet pincet, lille saks og foråret saks) og selvstændig forsegling materiale belagt dissektion retter med 70% ethanol og lad dem tørre i dissektion hætte.
2. chick embryonale Neuron kultur og netværksaktivitet
- På dagen for plating, fjerne et hætteglas af ECM fra-20 ° C fryser, spray med 70% ethanol, og Anbring på is. Fortyndet til 25% ved at tilføje 300 µL koldt neurobasal medium inde BSLII hætte.
- Ved hjælp af en P200 pipetteman tilføje 100 µL af 25% ECM til midten af MEA pas på ikke at røre ved elektroderne og fjerne øjeblikkeligt forlader en tynd film på overfladen. Dække MEA og placere i CO2 inkubator (37 ° C og 5% CO2) indtil klar til pladen af neuroner.
- Sterilisere yderstof af et E7 æg (embryonale dag 7, inkuberes ved 37 ° C i 7 dage) med 70% ethanol. Hug hovedet af fosteret i et Sylgard nederste dissektion skål indeholdende kolde sterile Hank afbalanceret salte løsning (HBSS) uden calcium. Skær omkring øjnene og Fjern øjne. Ved hjælp af DuMont #5 fine pincet og foråret saks lave et snit på den ventrale side og fjerne de ydre lag af huden til at udsætte den forhjernen og optiske tectum. Skræl og fjern den pial membran omhyggeligt. Overføre forebrain til en anden petriskål og skær den i små stykker på ca. 2 mm med fjeder saks.
Bemærk: Bør ikke være nogen blodkar, der er knyttet til forebrain efter fjernelse af pial membranen. Hvis den er tilgængelig, er det at foretrække at udføre dissektion i en steril dissektion hætte, selvom vi har opnået ganske gode resultater, når dissektion er udført en hætte, så længe de instrumenter og dissektion område er grundigt steriliseres med 70% ethanol . - Ved hjælp af en steril overførsel pipette, indsamle stykker af forhjernen i en 15 mL-centrifugerør og fjerne så meget af HBSS som muligt efter at lade stykker af forhjernen synker til bunden i centrifugeglasset.
- Der tilsættes 1 mL 0,05% Trypsin/EDTA (pre-hjertelig til 37 ° C) og inkuberes ved 37 ° C i 15 minutter.
- Bruger en Pasteur pipette, omhyggeligt fjerne trypsin uden at forstyrre stykkerne af væv og tilsæt 1 mL neurobasal medium. Lade stykker af væv synker til bunds og fjerne medium. Gentag denne vask en gang mere. Dette trin er at vaske af trypsin/EDTA.
- Der tilsættes 2 mL neurobasal medium og starte triturating. Ændring indebærer at tage en steril brand-poleret Pasteur pipette og passerer væv gennem det flere gange forsigtigt indtil ingen flere stykker af væv er set. Hvis nogen stykker stadig efter gentagne afleveringer, bare lade dem at bilægge til bunden.
Bemærk: Undgå skumdannelse mens triturating - hvis skum danne, stoppe og fjerne ved aspiration. - Fortynd re suspenderede celler 1:10 med neurobasal medium og tælle levedygtige celler benytter Trypan blå farvestof og en hemocytometer. Mix 50 µL af de fortyndede celler med 50 µL af Trypan blå opløsning i et 0,5 mL centrifugeglas. Tilføje 10 µL til hemacytometer efter markedsføring af coverslip og tæller de lyse klare celler (blå celler er døde og bør ikke medregnes).
Bemærk: Typisk celle numre fra et enkelt E7 optiske tectum interval mellem 1 og 5 x 107 celler/mL. - Plade de dissocierede celler på ECM belagt MEAs med en tæthed på 2.200 celler/square mm. For vores MEA system svarer dette til omkring 130.000 celler pr. MEA. Tilføj neurobasal medium for at bringe volumen i MEA til 1 mL og placere MEAs i CO2 inkubator natten for celle vedhæftet fil og neurite forlængelse (fig. 1).
Bemærk: For multilaterale miljøaftaler på forskellige områder og elektrode numre, forskellige celle tætheder kunne blive prøvet - generelt højere celle tæthed resulterer i større netværksaktivitet, men også fører til kortere levede kulturer. - Den næste dag, tilføje BPA til en slutkoncentration på 10 µM i neurobasal medium fra 1 mM lager foretaget i trin 1.1 til de behandlede MEA. Kontrolelementet MEA, tilføje den samme mængde 10% ethanol i vand (v/v) opløsning.
- Erstat brugt medium med neurobasal medium indeholdende 10 µM BPA hver anden dag indtil 7 dage in vitro- (7DIV) og hver dag derefter, som netværk aktivitet stiger.
Bemærk: Netværksaktivitet stiger, metaboliske aktivitet øger og medieændringer skal være mere hyppige. Lad ikke medium vende orange.
3. registrering Neuronal netværksaktivitet
- På dagen for erhvervelse, udveksle næringssubstratet med friske neurobasal medium og vende tilbage de multilaterale miljøaftaler til CO2 inkubator for mindst 2 h før optagelse. Start optagelse software og Indstil temperaturen af MEA til 37 ° C ved at klikke på ikonet temperatur (supplerende figur 3.1a-b).
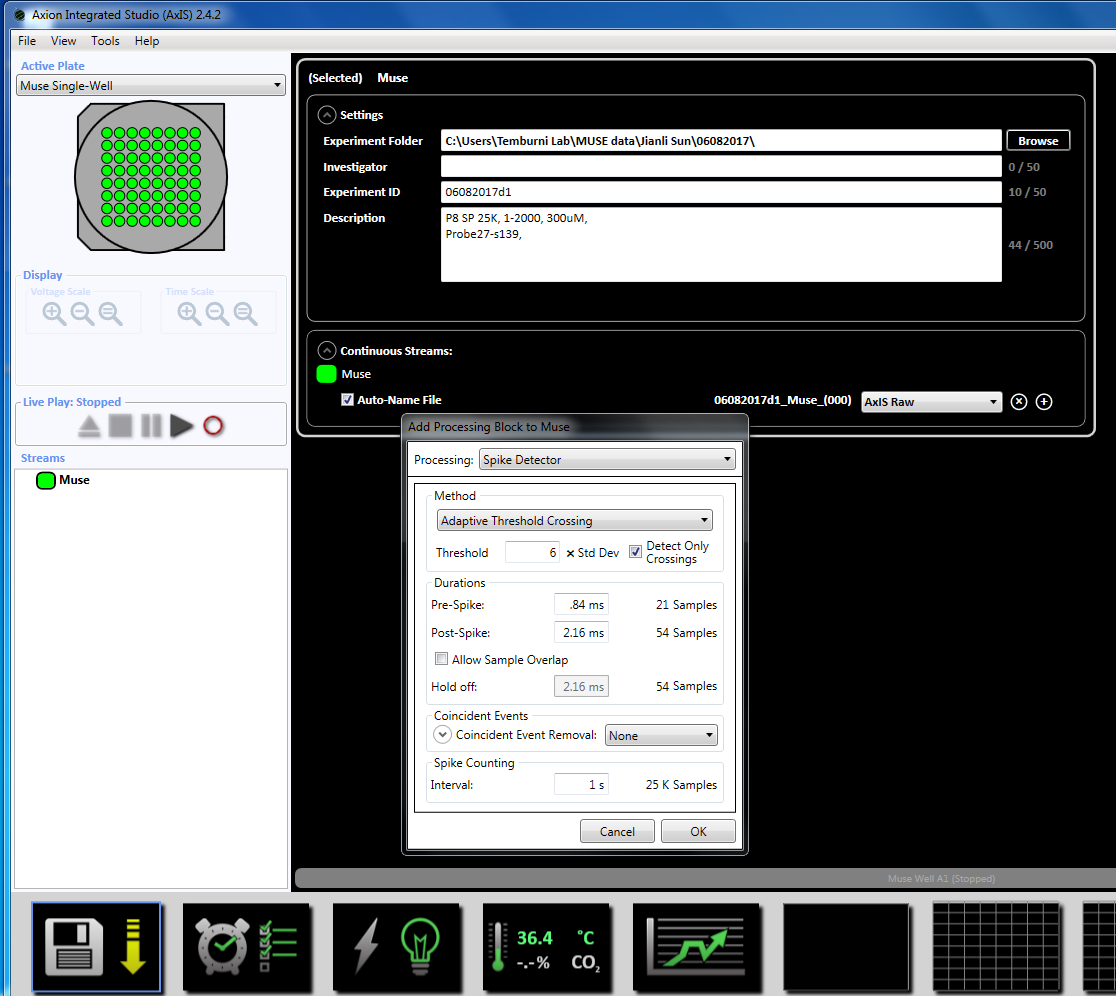
Bemærk: Registrering kan begynde så tidligt som i dag 3 i yderklædningen og kan fortsætte så længe kulturer er i live. - Konfigurere parametrene erhvervelse ved at højreklikke på ikonet "Muse" (under strømme i det venstre vindue panel) og vælg "Tilføj behandling" og "Spike detektor" og klik på OK i pop op-vinduet. "Spike detektor (6 x STD)" vises under filnavnet (supplerende figur 3.2a-b).
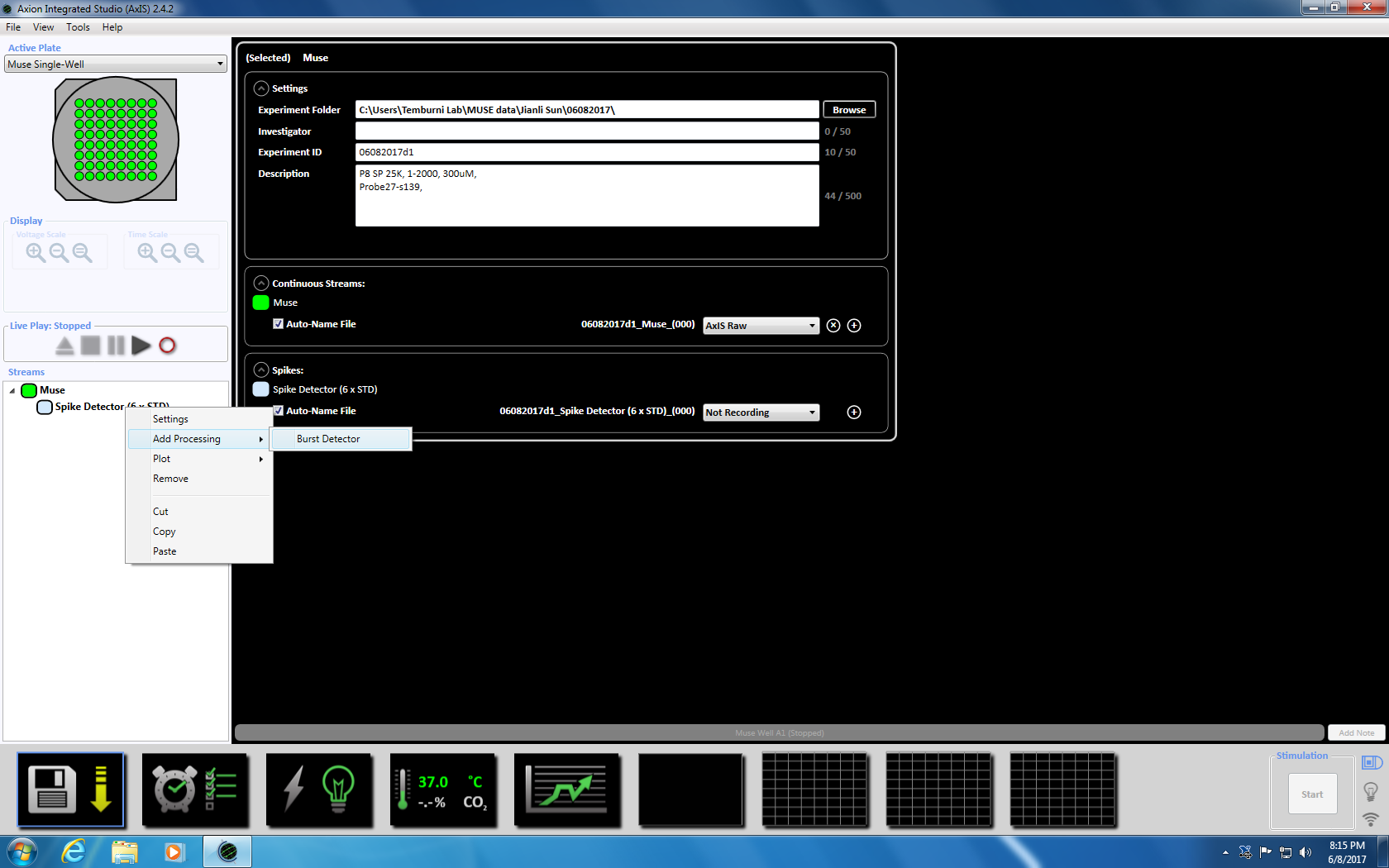
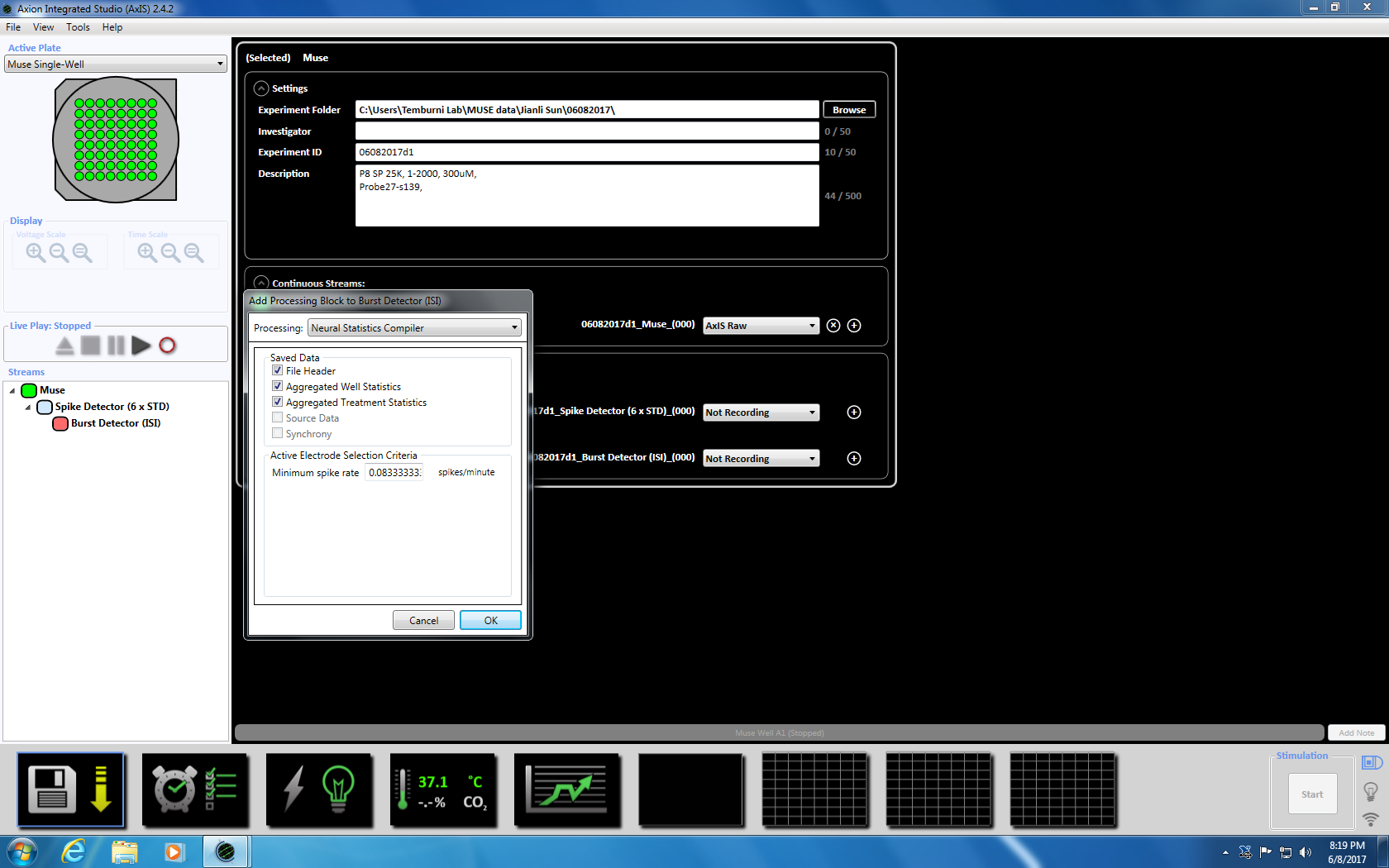
- Næste Højreklik på Spike detektor, Vælg "Tilføj behandling" og "Burst detektor" og klik på OK i pop op-vinduet. "Burst detektor" (ISI) vises under ikonet Muse (supplerende figur 3.3a-b).
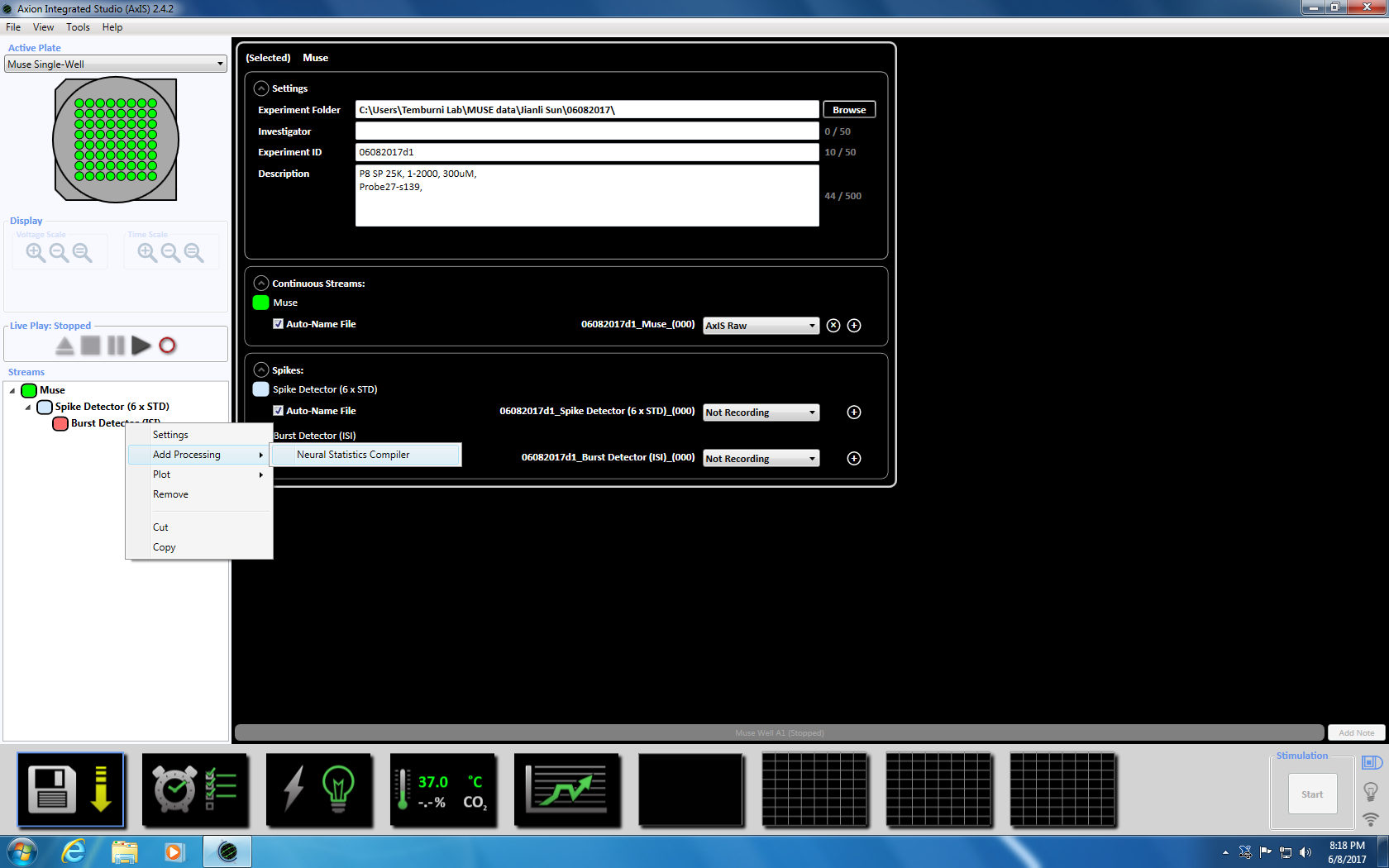
- Næste Højreklik på Burst detektor, Vælg "Tilføj behandling" og "Neurale statistik Compiler." I pop-up-vinduet, sikre at filens Header, aggregeres godt statistikker og Synchrony er markeret. Klik på OK. "Statistik Compiler" vises nedenfor (supplerende figur 3.4.a-b).
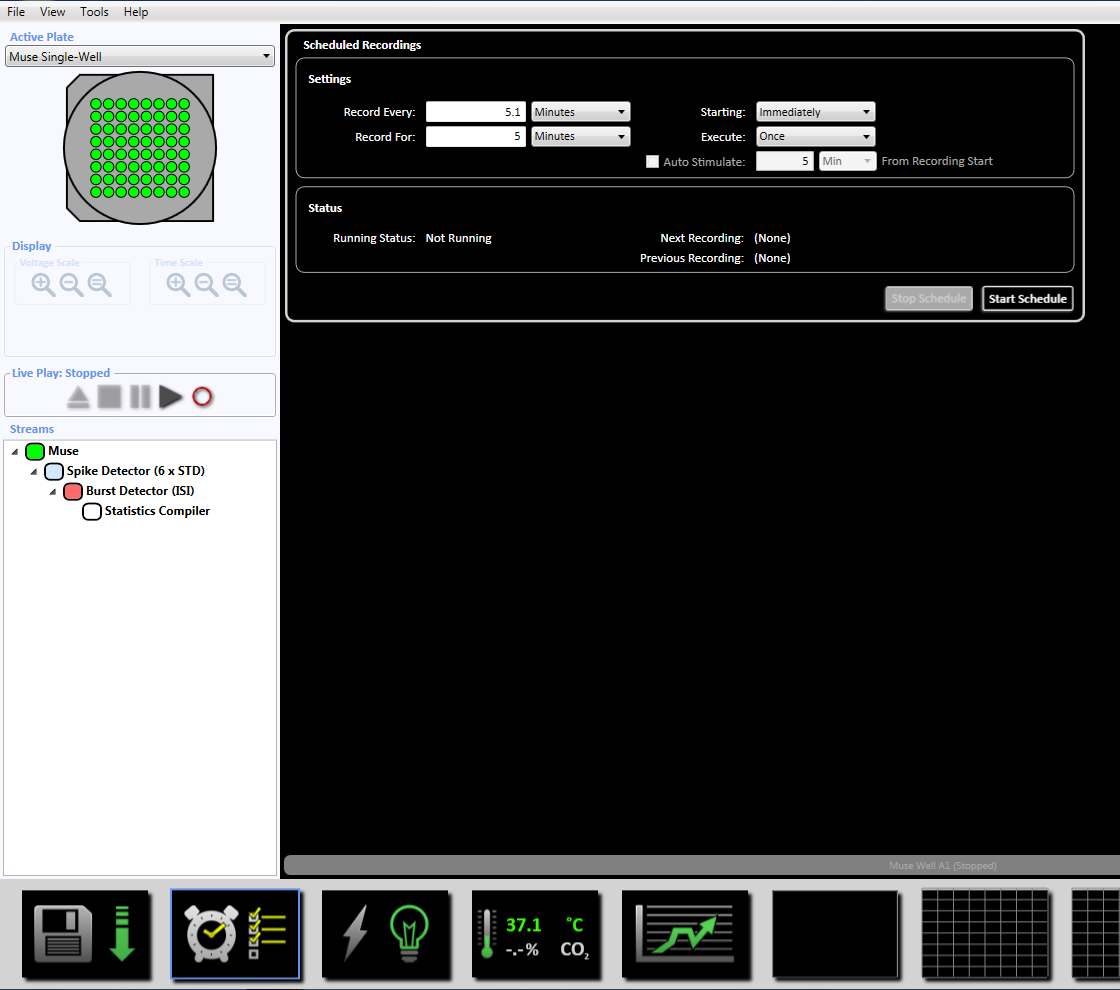
Bemærk: Dette vil indstille adaptive tærskel passage (6 x STD filter), Inter spike interval på 100 ms med et minimum af pigge på 5, betyder fyring sats opdagelse på 10 s med en synchrony parameter på 20 ms, og den mindste spike hastighed 0.083333 pigge pr. min. - Oprettet den planlagte optagelse for optagetid på 5 minutter (300.000 ms) ved at klikke på ur-ikonet nederst. Dette opnås ved at ændre oplysningerne i afsnittet indstilling til at registrere alle 5.1 minutter og optage i 5 minutter. Dette vil blive indledt umiddelbart og vil blive udført en gang (supplerende figur 3.5).
- Sikre, at temperaturen i MEA har nået 37 ° C før du flytter MEA. Fjerne MEA fra rugemaskinen, placere den på optagelse enhed og låse MEA. Starte optagelsen netværksaktivitet ved at klikke på start-post i afsnittet planlagt optagelse eller ikonet post i vinduet "Fil afspilles". Typisk, en optagetid på 5 minutter (300.000 ms) er tilstrækkelig, selv om længere optagelser af op til 10 minutter kan fås.
- Efter optagelse, skal du returnere MEA til rugemaskine.
Bemærk: Pleje skal tages for at opretholde sterilitet og at minimere tiden MEA er udenfor rugemaskine.
4. dataanalyse
- Analyse af de rå data kan gøres offline ved hjælp af optagelse software. Klik på fil rage ned menu og åbne optagelse. Vælg rå datafilen skal analyseres. Replay de rå optagelser til at fange de nødvendige spiking parametre.
- For at få den gennemsnitlige antallet af pigge, bruge modulet Spike detektor og for at opnå Synchrony indeks, bruge modulet neurale statistik Compiler.
- Indlæse Spike detektor, brast detektor og statistik Compiler moduler som før (afsnit 3).
- Fra rullemenuerne i Spike detektor, brast detektor og statistik Compiler modul windows vælge Spike liste, brast netværksliste og avancerede Metrics, henholdsvis.
- Klik på knappen Optag for at starte optagelsen den datafil, som vil skabe en .csv-fil i mappen styret. Spike parametre - antallet af pigge og Synchrony indeks - vil blive registreret i .csv-filen.
Bemærk: realtid analyse mens du optager rådata anbefales ikke, da dette kunne forbruge Processortid og it-ressourcer.
Representative Results
Vi undersøgte effekten af BPA på synchrony udvikling i kylling embryo neuronal kulturer. Synkronisering af spiking aktivitet er en indikator for den normale udvikling af neuronal netværk in vitro. Når to pigge forekomme inden for 5 ms af hinanden, betragtes de som synkrone. In vitro neuronal kulturer i første omgang vise tilfældige spiking aktivitet — pigge opstår tilfældigt og Inter spike intervallerne viser en normalfordeling. Som kulturer modne, spiking aktiviteten bliver mere synkroniseret og Inter spike intervaller er konstant. Synkron spiking aktivitet af en kultur var quantitated af cross-korrelation analyse af spike gange benytter optagelse software12 for at udlede en Synchrony indeks (SI). Vi fandt, at BPA eksponering negativt påvirker netværk aktivitet udvikling ved at reducere spiking aktivitet og synchrony indeks (figur 2en og 2b). Sammenfattende har vi vist, at de skadelige effekter af BPA eksponering på et foster påvirker dens udvikling, herunder neuronal funktion, der kan vurderes gennem synchrony udvikling i chick neuroner i kultur.

Figur 1: Skematisk af protokollen skildrer skridt fra dissektion af chick forhjernen til dissociation, plating, og optagelse af netværksaktivitet. Venligst klik her for at se en større version af dette tal.

Figur 2: (A) det gennemsnitlige antal pigge er betydeligt lavere i BPA behandlet E7 chick forhjernen kulturer sammenlignet med kontrol kulturer. Det gennemsnitlige antal af pigge blev udvundet ved hjælp af optagelse software. Betyder (kontrol, 14,890 ± 949.0, N = 15 og BPA, 5,624 ± 465.9, N = 22) var signifikant forskellig (uparret t-test, p < 0,0001). (B) det gennemsnitlige synchrony indeks (SI) er betydeligt lavere i BPA behandlet E7 chick forhjernen kulturer sammenlignet med kontrol kulturer. SI blev udvundet ved hjælp af modulet neurale statistik Compiler inden for optagelse software. SI midler (kontrol, 0.5159 ± 0.06547 N = 15 og BPA, 0.1140 ± 0.01840 N = 22) var signifikant forskellig (uparret t-test, p < 0,0001). Fejllinjer skildrer standardafvigelse. Venligst klik her for at se en større version af dette tal.
Discussion
Faser af hurtig vækst og udvikling såsom embryogenese er særligt sårbare over for de skadelige virkninger af forskellige hormonforstyrrende stoffer herunder BPA. Vi leverer detaljerede protokoller for at studere virkningerne af BPA eksponering i en in vitro- hvirveldyr neuronal netværksmodel. Ved hjælp af denne protokol, etableret vi at BPA sænker spike sats samt synchrony indekset i de udviklende neuronal netværk (figur 2a-b).
Vores metode var designet til at studere BPA eksponering under tidlige embryogenese og nemt kan tilpasses for at studere virkningerne af andre EDC. Embryonale neuronal kulturer fra chick er relativt let at etablere i forhold til andre hvirveldyr modeller, herunder rotte eller mus. Desuden er der ingen krav om en separat animalske rum — en simpel inkubator på et laboratoriebænk er tilstrækkelige til at udføre disse assays. Vi beskriver brugen af en multi elektrode system (MEA) for at vurdere effekten af EDC, BPA, på netværksaktivitet. De protokoller er beskrevet her kan anvendes på andre systemer. Et kritisk aspekt i denne protokol er vedligeholdelse af sterilitet. Dette omfatter dissektion af embryo under sterile forhold — sterilisering af instrumenter med 70% ethanol og brug af sterile Hank afbalanceret salte løsning er tilstrækkelig. Da data er indsamlet over en periode på uger, er det afgørende at opretholde steril håndtering af de multilaterale miljøaftaler. Neuronal kulturer en gang etableret kan være kulturperler langvarig - op til tre måneder – og er derfor nyttig for at studere langtidseksponering EDC og andre forbindelser på neuronal netværk. Et andet vigtigt aspekt af denne protokol er tiden fra dissektion til plating. Det optimale tidspunkt er 30 min, herunder inkubation af væv med trypsin. Jo længere den tid, det tager, jo dårligere overlevelse af celler.
Vi beskriver her en grundlæggende protokol, der kan anvendes til vurdering af eventuelle kemiske eller stof, der påvirker adfærd og neurale funktion. Her, har vi brugt en celle tæthed af 2.200 celler pr. mm2; men dette kan ændres og andre celle tætheder kan bruges. Generelt har vi fundet at øge celle tæthed øger netværksaktivitet - flere pigge på kortere tid. Metoden beskrevet, selvom meget nyttigt i vurderingen af virkningerne af kemikalier på netværksaktivitet har begrænsninger. En af de største begrænsninger af metoden er, at disse in vitro kulturer er to dimensionelle ikke måske afspejler den tre-dimensionelle arkitektur af hvirveldyr hjernen. Dette kan løses ved hjælp af skive optagelser. Et andet alternativ er at anvende behandlingerne til at udvikle kylling embryo af betyder i et lille vindue skåret på den brede ende af æg15, dissekere hjernen i slutningen af behandlingsregime, gøre tykke sektioner på en vibratome og sted på MEA for optagelse netværksaktivitet.
Vores protokol er væsentlig, idet det giver mulighed for undersøgelse af effekten af EDC på udviklingen af netværksaktivitet og tilbyder udforskning af en mekanistisk grundlag for virkningerne af disse kemikalier.
Disclosures
Forfatterne erklærer, at de har ingen konkurrerende interesser.
Acknowledgments
Denne undersøgelse er støttet af NSF (HBCU-UP forskning indledning Award, HRD 1401426 og EPSCoR EPS-0814251) og NIH (COBRE 1P20GM103653-01A1). K.S. understøttes af et stipendium fra Delaware INBRE-III 6404.
Materials
| Name | Company | Catalog Number | Comments |
| #5 foreceps | Fine Science Tools | 11251-10 | |
| Axion Muse MEA | Axion Biosystems | M64-GL1-30Pt200 | Will be called MEA system in manuscript |
| Axis Software | Axion Biosystems | Will be called recording software in the manuscript | |
| BPA | Sigma-Aldrich | 239658-250g | |
| curved forceps | Fine Science Tools | 11272-50 | |
| EtOH | Sigma-Aldrich | 64-17-5 | |
| Fertilized chicken eggs | from any local farm or Spafas | ||
| HBSS | Fisher | 14170112 | |
| Hemacytometer | Fisher | 02-671-6 | |
| Matrigel Growth Factor Reduced, Phenol Red-Free | BD Biosciences | 356231 | Will be called Extra Cellular Matrix (ECM) in the manuscript |
| Neurobasal medium | BrainBits | Nb4-500 | |
| Neuroexplorer statistical software | Nex Technologies | Neuroexplorer version 5 | |
| Pasteur pipettes | Fisher | 13-678-20A | |
| spring scissors | Fine Science Tools | 15514-12 | |
| Sylgard bottom dissection dishes | Living Systems Instrumentaion | DD-90-S-BLK-3PK | |
| Trypan Blue dye | Fisher | 15-250-061 | |
| Trypsin-EDTA | Fisher | 15400054 |
References
- Anahara, R., Yoshida, M., Toyama, Y., Maekawa, M., Masayuki, K., Ishino, F., Toshimori, K., et al. Estrogen agonists, 17 beta-estradiol, bisphenol A, and diethylstilbestrol, decrease cortactin expression in the mouse testis. Arch. of Histol. Cytol. 69 (2), 101-107 (2006).
- Dodds, C. Synthetic oestrogens. Br. Med. Bull. 11 (2), 131-134 (1955).
- Grignard, E., Lapenna, S., Bremer, S. Weak estrogenic transcriptional activities of Bisphenol A and Bisphenol S. Toxicology In Vitro. 26 (5), 727-731 (2012).
- Herbst, A. L., Ulfelder, H., Poskanzer, D. C. Adenocarcinoma of the vagina: association of maternal stilbestrol therapy with tumor appearance in young women. New Eng. J. Med. 284, 878-881 (1971).
- Jenkins, S., Raghuraman, N., Eltoum, I., Carpenter, M., Russo, J., Lamartiniere, C. A. Oral exposure to bisphenol A increases dimethylbenzanthracene-induced mammary cancer in rats. Environ Health Persp. 117 (6), 910-915 (2009).
- Okada, A., Kai, O. Effects of estradiol-17beta and bisphenol A administered chronically to mice throughout pregnancy and lactation on the male pups' reproductive system. Asian J Androl. 10 (2), 271-276 (2008).
- Palanza, P., Gioiosa, L., Vom Saal, F. S., Parmigiani, S. Effects of developmental exposure to bisphenol A on brain and behavior in mice. Environ. Res. 108 (2), 150-157 (2008).
- Mimoto, A., Fujii, M., Usami, M., Shimamura, M., Hirabayashi, N., Kaneko, T., et al. Identification of an estrogenic hormone receptor in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 364 (4), 883-888 (2007).
- Allard, P., Colaiacovo, M. Bisphenol A impairs the double-strand break repair machinery in the germLine and causes chromosome abnormalities. Proceedings Natl. Acad. Sci. 107 (47), 20405-20410 (2010).
- Mersha, M. D., Patel, B. M., Patel, D., Richardson, B. N., Dhillon, H. S. Effects of BPA and BPS exposure limited to early embryogenesis persist to impair non-associative learning in adults. Behav. Brain Funct. : BBF. 11, 27 (2015).
- Chen, Y., Shu, L., Qiu, Z., Lee, D. Y., Settle, S. J., et al. Exposure to the BPA-Substitute Bisphenol S Causes Unique Alterations of GermLine Function. PLOS Genetics. 12 (7), 1006223 (2016).
- Shein Idelson, M., Ben-Jacob, E., Hanein, Y. Innate Synchronous Oscillations in Freely-Organized Small Neuronal Circuits. PLoS ONE. 5 (12), 14443 (2010).
- Baltz, T., Herzog, A., Voigt, T. Slow Oscillating Population Activity in Developing Cortical Networks: Models and Experimental Results. J. Neurophysiol. 106 (3), 1500-1514 (2011).
- Pettmann, B., Louis, J. C., Sensenbrenner, M. Morphological and Biochemical Maturation of Neurones Cultured in the Absence of Glial Cells. Nature. 281 (5730), 378-380 (1979).
- Zhang, H., Wu, C. Y., Wang, W., Harrington, M. A. Interneuronal Synapses Formed by Motor Neurons Appear to Be Glutamatergic. NeuroReport. 22 (16), 809-813 (2011).
- Paiva, A. R. C., Park, I., Príncipe, J. C. A Comparison of Binless Spike Train Measures. Neural Comp. and Appl. 19 (3), 405-419 (2010).
- Cretu, A., Fotos, J. S., Little, B. W., Galileo, D. S. Human and Rat Glioma Growth, Invasion, and Vascularization in a Novel Chick Embryo Brain Tumor Model. Clin Exp Metastasis. 22, 225-236 (2005).
- Chiappalone, M., et al. Dissociated cortical networks show spontaneously correlated activity patterns during in vitro development. Brain Res. 93, (2006).
- Li, X., et al. Long-term recording on multi-electrode array reveals degraded inhibitory connection in neuronal network development. Biosens Bioelectron. 22 (7), 1538-1543 (2007).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}