

Summary
Eksponering for miljøgifter kan akutt påvirke utvikling embryo. Endokrine forstyrre kjemikalier som bisphenols er kjent for å påvirke nervesystemet. Her beskriver vi en protokoll med en i vitro virveldyr (chick embryoet) nevrale nettverk-modell for å studere funksjonelle virkningen av gift eksponering på tidlig embryo.
Abstract
BIS-fenoler, som bis-fenol A (BPA) og bis-fenol-S (BPS), er polymerizing agenter som er mye brukt i produksjon av plast og mange dagligdagse produkter. De er klassifisert som endokrine forstyrre forbindelser (EDC) med østradiol-lignende egenskaper. Langvarig eksponering EDCs, selv ved lave doser, har vært knyttet til forskjellige sunnhet feilene inkludert kreft og atferdsforstyrrelser infertilitet, med større sårbarhet i tidlig utviklingsmessige perioder. For å studere virkningene av BPA på utviklingen av nevrale funksjon, brukte vi et i vitro nevrale nettverk avledet fra tidlig chick embryonale hjernen som modell. Vi fant at eksponering for BPA påvirket utviklingen av nettverksaktivitet, spesielt skyter aktivitet og synkronisering. En endring i nettverksaktivitet er avgjørende koblingen mellom molekylære mål av et medikament eller sammensatte og dens effekt på atferdsdata utfallet. Flere elektrode matriser blir stadig mer nyttig verktøy for å studere effekten av legemidler på nettverk aktivitet i vitro. Det er flere systemer tilgjengelig i markedet, og selv om det er variasjoner i antall elektroder, typen og kvaliteten på elektroden matrisen og analyseprogramvare, de grunnleggende underliggende prinsippene, og dataene innhentet er den samme på tvers av forskjellige systemer. Foreløpig begrenset til analyse av todimensjonal i vitro kulturer, blir disse MEA systemer forbedret for å aktivere i vivo nettverksaktivitet i hjernen skiver. Her gir vi en detaljert protokoll for embryonale eksponering og nevrale nettverksaktivitet og synchrony, sammen med representant resultater.
Introduction
4, 4-Isopropylidenediphenol, ofte referert til som bisfenol-A eller BPA, finnes en antioksidant additiv i en rekke polykarbonat og epoxy harpiks basert produkter fra thermal papir, CDer og knuse bevis glass, på innsiden belegg av drikke bokser. Selv om BPA er kjent for å være en endokrine forstyrre agent som etterligner østrogen1, har studier på dens effekter skyldes daglige casual eksponering for BPA hovedsakelig kommet ut de siste 10-15 årene. Konsekvensene av med lave nivåer av BPA eksponering er dypeste i tidlig utviklingsmessige perioder, inkludert under embryogenesis gjennom eksponering mødre2,3. In utero eksponering for kvinnelige embryo endokrine forstyrre kjemikalier har også vært knyttet til økt sykdom mottakelighet fra vaginal til brystet kreft4,5. Dyrestudier har vist eksponering for BPA fører til atypisk hjernens struktur og funksjonelle unormalt manifestert i atferd6. Som en konsekvens, har bruk av BPA i spedbarn fôring flasker blitt svartelistet av mest regulerende instanser i Europa og Nord-Amerika, inkludert FDA. For å overholde forskrifter, har mange produsenter byttet til 4, 4'-sulfonyldiphenol eller bisfenol-S (BPS). Basert på det faktum at BPA og BPS er strukturelle analoger og at siste rapportene viste sammenlignbar styrken på BPS i estrogenic transkripsjon7, er det viktig å studere toksisitet av denne sammensatte forhold til BPA.
Her beskriver vi en protokoll for å teste effekten av BPA (og andre EDCs) på nettverk av neurons bruker virveldyr modell, chick fosteret. In vitro kulturer av neurons form synaptic kontakter og generere handling potensialer (også kalt pigger). Spiking aktiviteten av disse kulturene kan registreres med flere elektrode matrise (MEA) systemer. Toppene anses å være i synkronisering når de oppstår innen 5 ms av hverandre. Første tilfeldig skyter aktivitet som til slutt synkroniserer er et viktig trekk for å utvikle nevrale nettverk8,9. Synkronisering kan måles ved hjelp av ulike metoder og flere algoritmer er beskrevet i litteraturen9,10,11. I våre analyser bruker vi en algoritme utviklet av Paiva og kolleger12 som er integrert i innspillingen programvare som kjører MEA oppkjøpet systemet. Den robuste spiking aktiviteten av embryonale chick neurons gir en prototype for å studere effekten av BPA på nettverk aktivitet13. Bruker multi elektrode matriser til skyter aktiviteten, observerte vi at BPA eksponering hemmer utviklingen av nevrale skyter synkronisering9,14. Her gir vi en detaljert metodikk for å studere BPA eksponering på chick embryoet nerveceller i kultur med representant resultater på utviklingen av nevrale skyter synkronisering i chick embryonale kulturer.
Protocol
Protokollen nedenfor blitt standardisert for å teste effekten av BPA eksponering under (tidlig) embryogenesis, og kan bli endret for bruk med BPS, BPF eller andre EDC i chick embryonale neurons.
Denne protokollen følger institusjonelle politikken til Delaware State University og NIH policy for avian embryoer. Protokollen er også i bekreftelse med DSUS materiale og kjemiske sikkerhetsretningslinjer.
1. materialet oppsett
- Løses BPA (m.w. 228.29) i 10% etanol (v/v) i vann for å lage en lagerløsning med 1 mM (0,23 mg per mL 10% etanol). Gjør etterfølgende fortynninger (0,1 µM, 0,5 µM, 1 µM, 5 µM og 10 µM) i neurobasal medium.
Forsiktig: BPA er en miljømessig gift og forsiktighet bør utvises ved håndtering av pulver. - Ruge kylling egg i en kommersielt tilgjengelig tabletop egg inkubator på 37 ° C. Ruge egg for 7 dager for E7 embryoer.
- Forberede MEAs. Sterilize antall tiltak av soaking i 70% etanol 3 til 4 h og skyll tre ganger i ca 10 mL steril destillert vann i BSLII panseret.
Merk: Tiltak inneholde 64 nanoporous platina elektroder ordnet i en 8 x 8-rutenett. Elektroden diameter er 30 µm og avstanden mellom elektrodene er 200 µm. Bruk ikke denaturert etanol for sterilisering av tiltak som dette vil føre til korrosjon av MEA. - Plasser tiltak inne en bakteriefri beholderen med lokk (for å opprettholde sterile forhold) og flytte til en bordplaten inkubator. Stek i minst 6 timer på 55 ° C. Dette trinnet er avgjørende for riktig sterilisering og temperaturen skal ikke overstige 60 ° C. Opprettholde sterilitet, flytte parabolen inneholder tiltak til BSLII panseret for lagring til videre bruk.
Merk: Vi bruker en ovn trygt glass bakervarer parabol med plast lokk og overflaten steriliseres med 70% etanol og plass i BSLII panseret for tørking. - Aliquot ekstra mobil matrix ECM løsningen. Tine ampullen på 4 ° C over natten og fryse 100 µL dele på 20 ° C.
Merk: ECM leveres frosset. ECM bør ikke bli brakt til romtemperatur før klar til coat den polymerizes i romtemperatur og, når polymerized, er det umulig å belegge. ECM uten vekst faktorer er foretrukket å forhindre flere effekter ukjent faktorene. - Sterilisere alle disseksjon instrumenter (Dumont #5 tang, buede tang, liten saks og våren saks) og selvtettende belagt disseksjon retter med 70% etanol og la dem tørke i disseksjon panseret.
2. chick embryonale Nevron kultur og nettverksaktivitet
- Ved plating, fjerne ampuller av ECM fra 20 ° C fryseren, spray med 70% etanol, og plasser på is. Fortynne 25% av tilføyer 300 µL kaldt neurobasal medium innenfor BSLII hette.
- Bruke en P200 pipetteman legge til 100 µL av 25% ECM til midten av MEA ta vare ikke å berøre elektrodene og fjerne umiddelbart forlater en tynn film på overflaten. Dekker MEA og plasser i CO2 inkubator (37 ° C og 5% CO2) til klar til plate neurons.
- Sterilisere den ytre skallet av E7 egg (embryonale dag 7 ruges på 37 ° C i 7 dager) med 70% etanol. Halshugge embryoet i en Sylgard bunnen disseksjon tallerken med kaldt sterilt Hanks balansert salter løsning (HBSS) uten kalsium. Skjær rundt øynene og fjern eyeballs. Bruke DuMont #5 fine tang og våren gjør en snitt på ventral side og fjerne det ytterste lag av huden til å utsette den forebrain og optikk tectum. Skrell og fjerne pial membranen nøye. Overføre forebrain til en annen Petriskål og skjær den i små biter på ca 2 mm med våren saks.
Merk: Det bør ikke være noen blodkar knyttet til forebrain etter fjerning av pial membranen. Hvis tilgjengelig, er det best å utføre dissection i sterilt disseksjon hette, selv om vi har oppnådd ganske gode resultater når disseksjon utføres utenfor hette som instrumenter og disseksjon området er grundig sterilisert med 70% etanol . - Bruker en steril overføring pipette, samle bitene i forebrain til en 15 mL sentrifuge rør og fjerne så mye av HBSS som mulig etter la biter av forebrain synke til bunns av sentrifuger røret.
- Legge 1 mL av 0,05% Trypsin/EDTA (pre oppvarmet til 37 ° C) og Inkuber på 37 ° C i 15 minutter.
- Bruker en Pasteur pipette, nøye fjerne trypsin uten å forstyrre biter av vev og legger 1 mL neurobasal medium. La biter av vev synke til bunns og fjerne mediet. Gjenta denne vask igjen. Dette trinnet er å vaske av trypsin/EDTA.
- Legg 2 mL neurobasal medium og starte triturating. Føden innebærer å ta en steril brann-polert Pasteur pipette og passerer vevet gjennom den flere ganger forsiktig til ingen flere biter av vev er sett. Hvis noen brikker igjen etter gjentatte passerer, bare la dem bosette til bunnen.
Merk: Unngå skumming mens triturating - hvis skum gjør danne, stoppe og fjerne av aspirasjon. - Fortynne re suspendert celler 1:10 med neurobasal medium og telle levedyktige cellers Trypan blått fargestoff og en hemocytometer. Mix 50 µL utvannet cellene med 50 µL Trypan blå løsning i et 0,5 mL sentrifuge rør. Legge til 10 µL av hemacytometer etter plassering på dekkglassvæske og telle lyse klart celler (blå celler er døde og bør ikke bli regnet).
Merk: Typisk cellen tallene fra en enkelt E7 fiberoptisk tectum varierer mellom 1 og 5 x 107 celler/mL. - Plate på avstand celler i ECM belagt tiltak på en tetthet på 2200 celler/kvadrat mm. For våre MEA system betyr dette ca 130 000 celler per MEA. Legg neurobasal medium for å bringe volumet i MEA til 1 mL og plassere tiltak i CO2 inkubator over natten for cellen vedlegg og neurite forlengelsen (figur 1).
Merk: For tiltak av ulike områder og elektroden tall, ulike celle tettheter kan prøves - generelt høyere celle tetthet resulterer i større nettverksaktivitet, men også fører til kortere bodde kulturer. - Neste dag, legge BPA til en siste konsentrasjon av 10 µM i neurobasal medium fra 1 mM lager gjort i trinn 1.1 behandlet MEA. Legge til samme volum av 10% etanol i (v/v) og vann kontroll MEA.
- Erstatt brukte medium med neurobasal medium som inneholder 10 µM BPA annenhver dag inntil 7 dager i vitro (7DIV) og hver dag etter som nettverket aktivitet øker.
Merk: Som nettverksaktivitet øker, metabolsk aktivitet øker og media endringer må være hyppigere. Ikke la mediet slå oransje.
3. registrere nevrale nettverksaktivitet
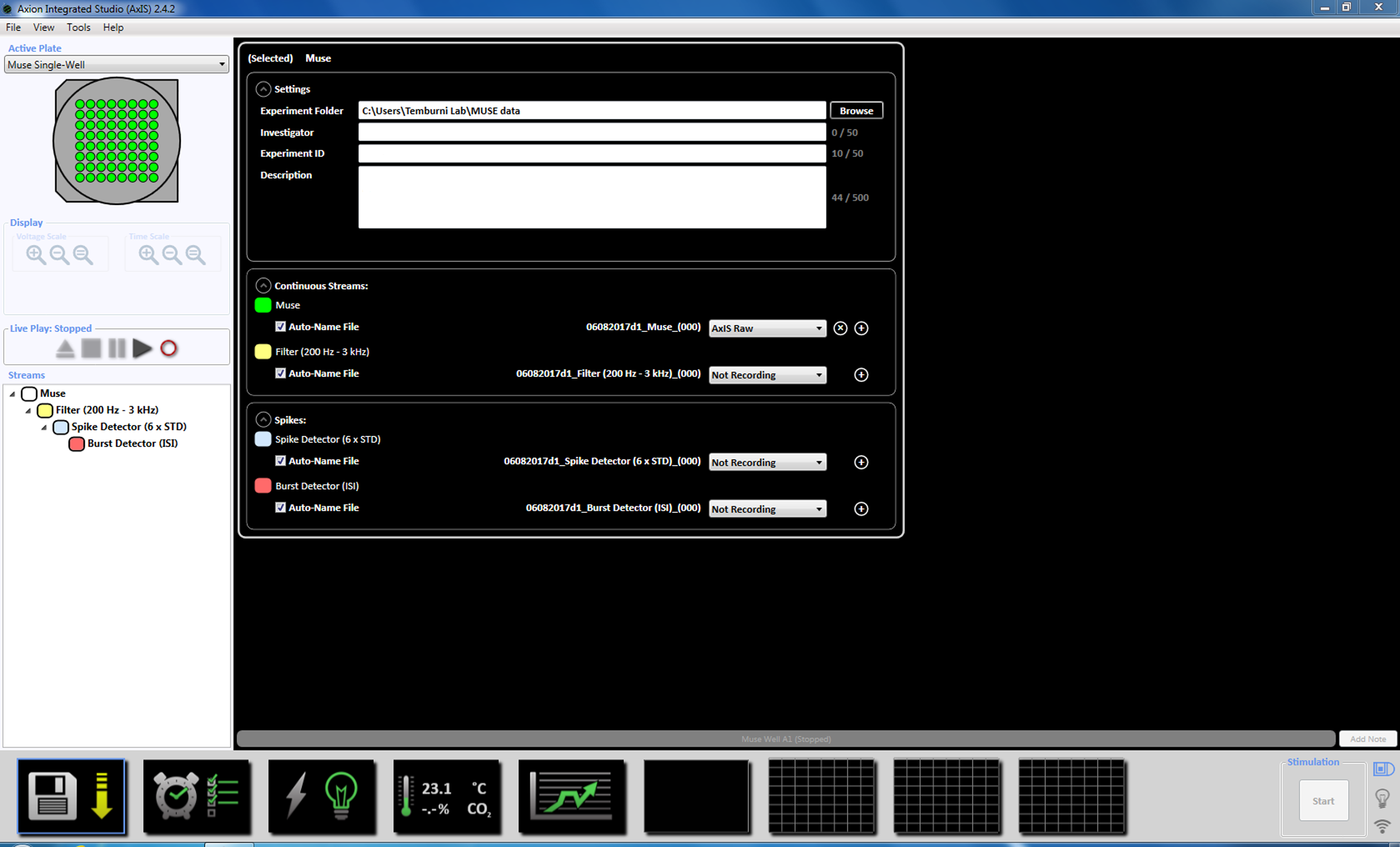
- Ved oppkjøp, utveksle kultur medium med fersk neurobasal medium og tilbake til tiltak til CO2 inkubator i minst 2 timer før opptak. Start innspillingen programvare og angi temperaturen på MEA 37 ° C ved å klikke på ikonet temperatur (supplerende figur 3.1a-b).
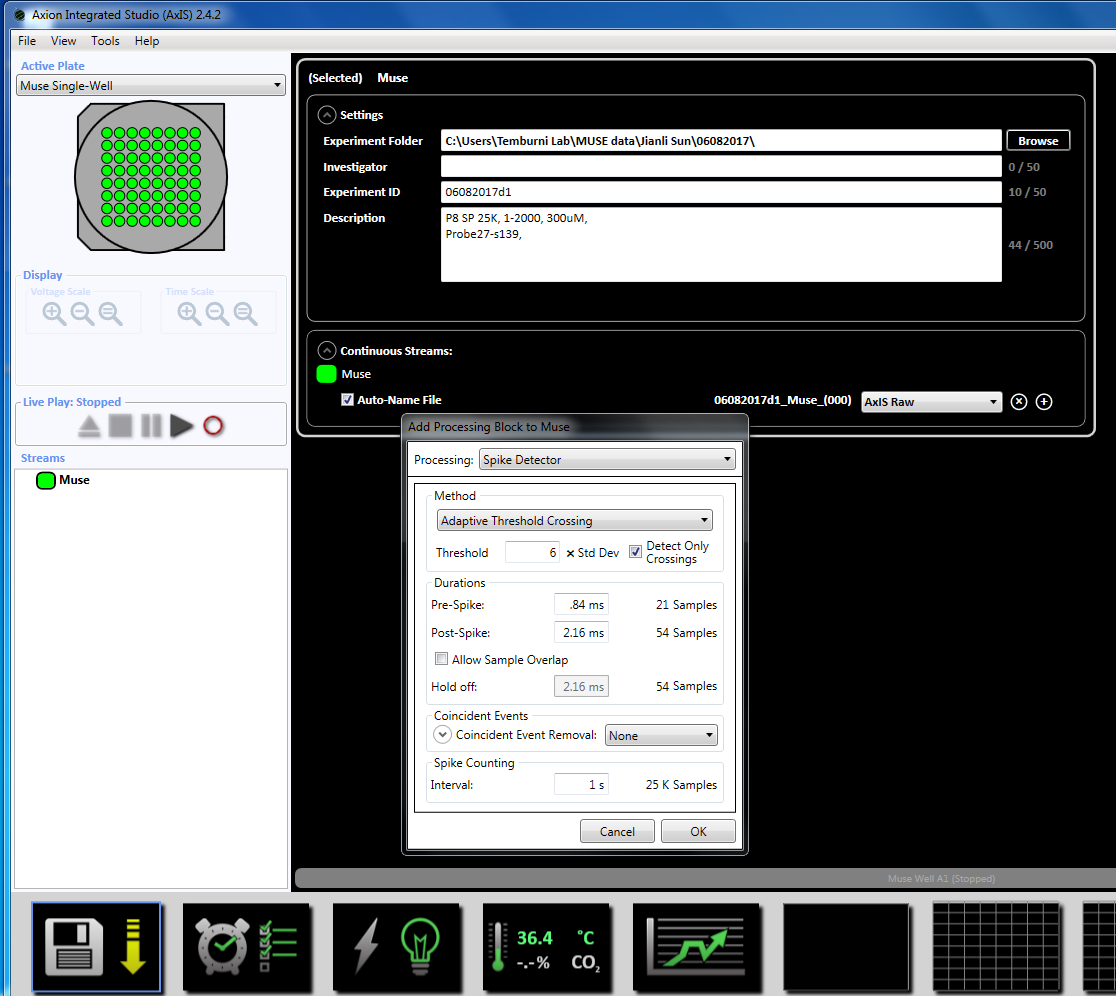
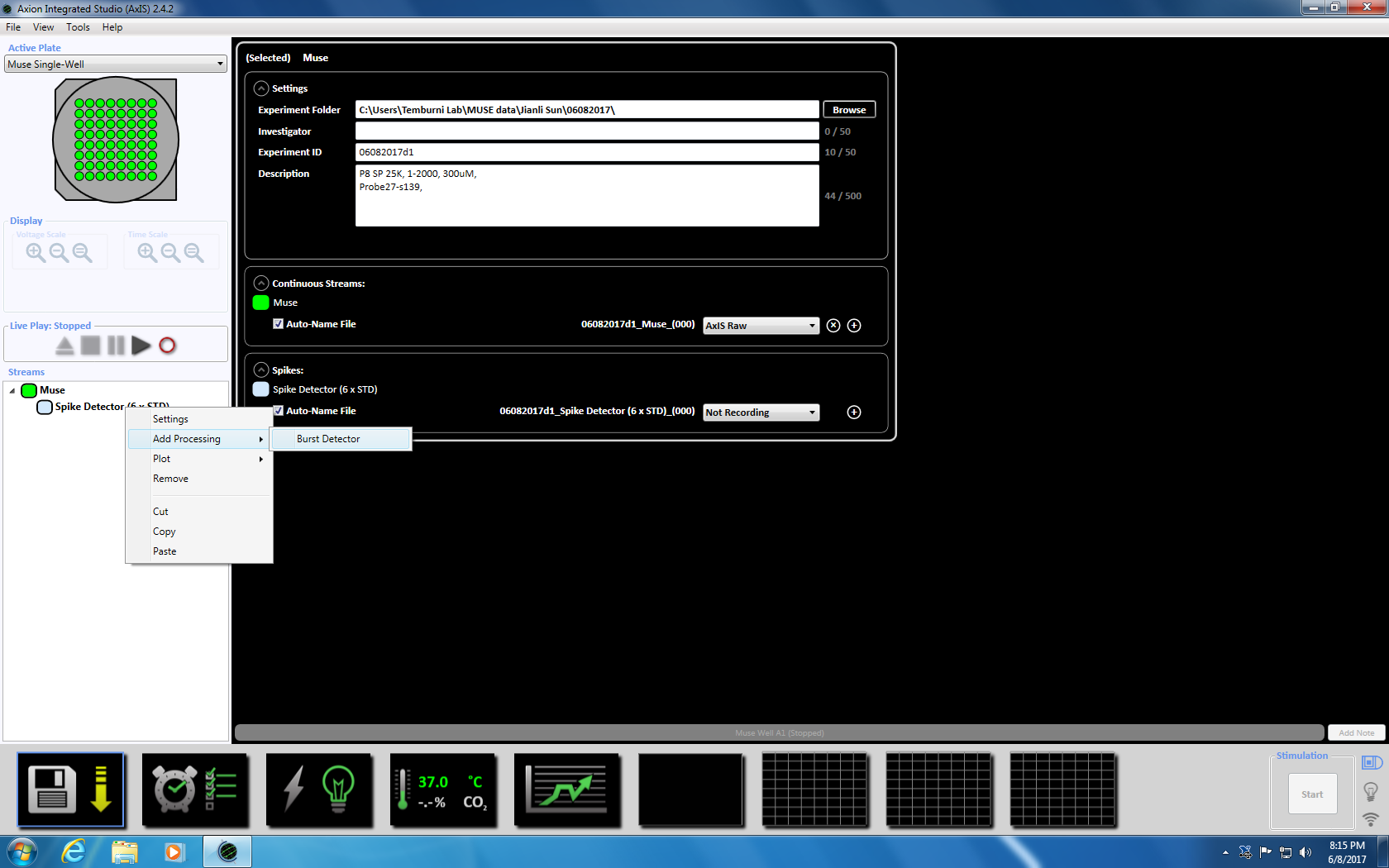
Merk: Opptak kan starte så tidlig som i dag 3 av plating og kan fortsette så lenge kulturer er i live. - Angi parametrene oppkjøpet ved å høyreklikke på ikonet "Muse" (under bekker i venstre vinduet panelet) og velg "Legg til behandling" og "Spike detektor" og klikk OK i popup-vinduet. "Spike detektor (6 x STD)" vises under filnavnet (supplerende figur 3.2a-b).
- Deretter høyreklikk på Spike detektoren, velg "Legg til behandling" og "Burst detektor" og klikk OK i popup-vinduet. "Burst detektor" (ISI) vises under ikonet Muse (supplerende figur 3.3a-b).
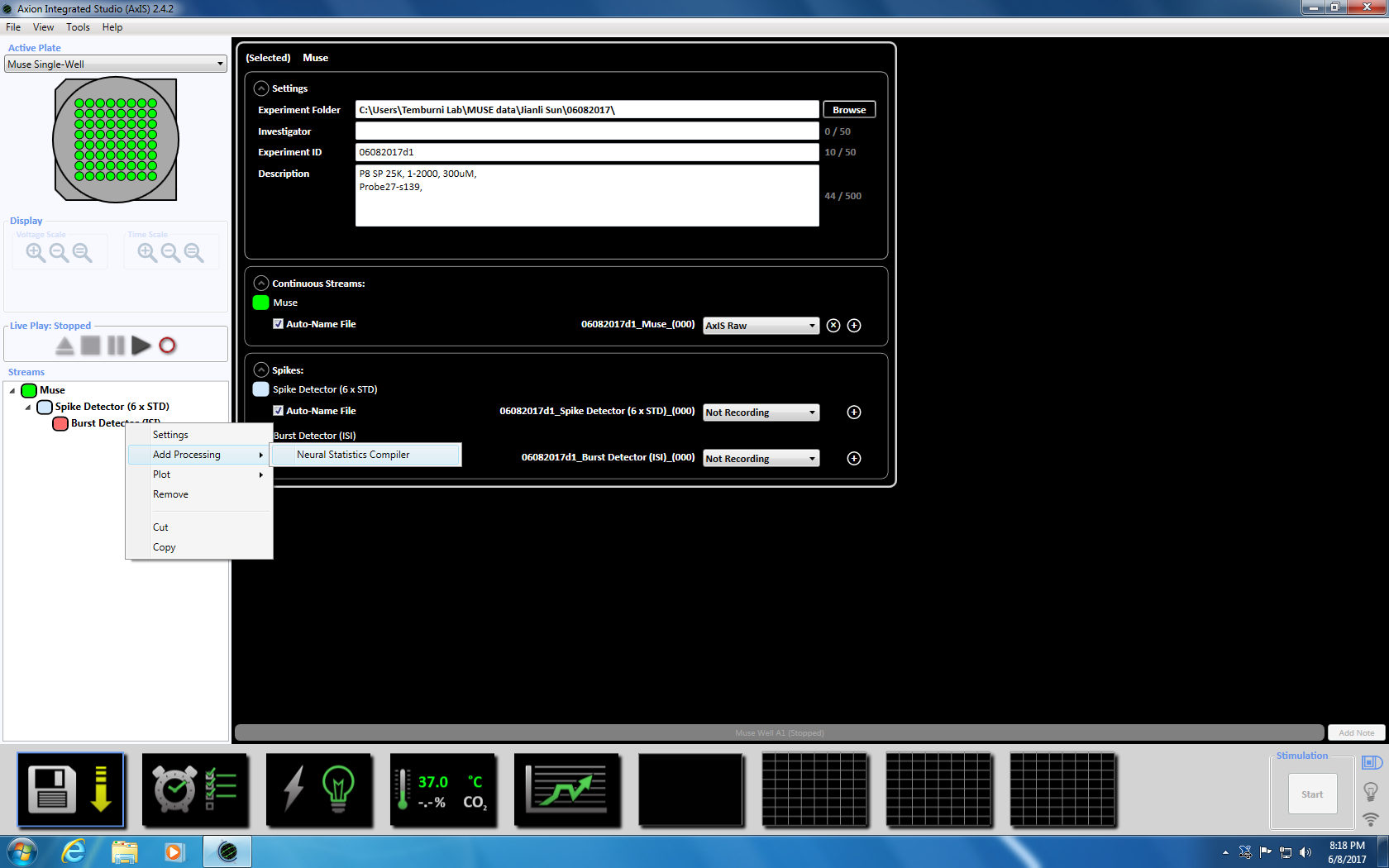
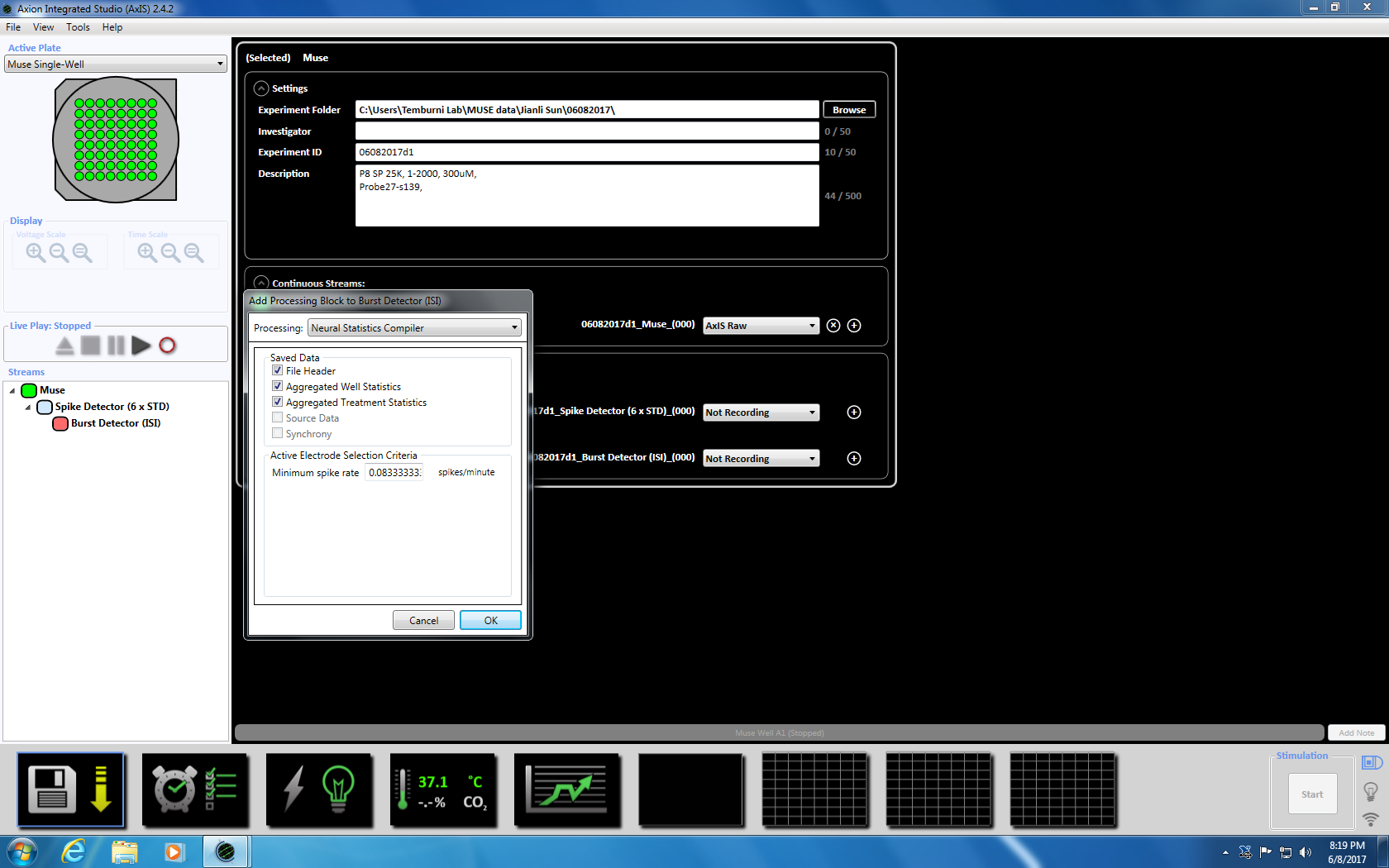
- Neste Høyreklikk Burst detektor, velg "Legg til behandling" og "Nevrale statistikk kompilatoren." I popup-vinduet, kan du sikre at filhodet, samles også statistikk og synkronisering er valgt. Klikk på OK. «Statistikk kompilatoren» vises under (supplerende figur 3.4a-b).
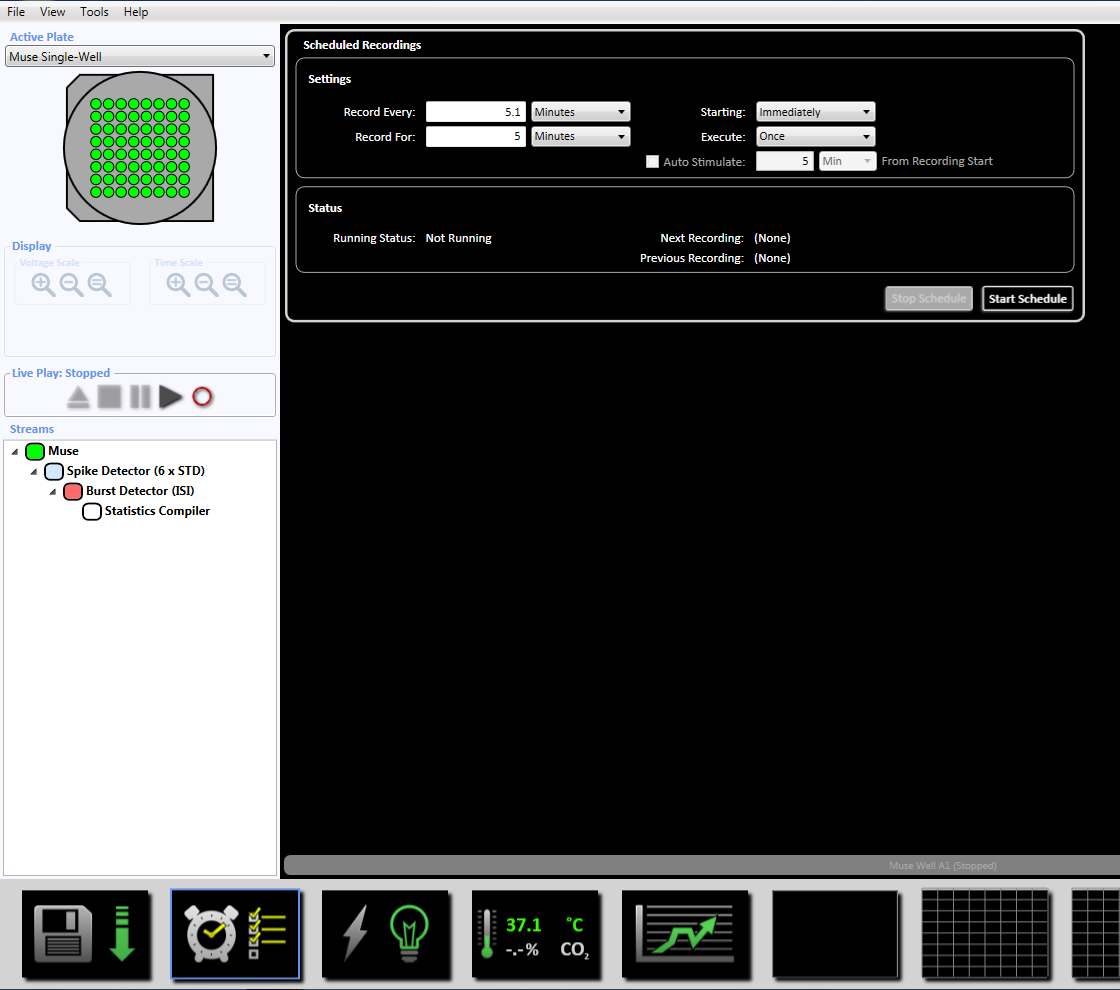
Merk: Dette setter dynamisk terskel krysset (6 x STD filter), mellom spike intervallet på 100 ms med et minimum antall pigger på 5, gjennomsnittet skyte rate påvisning på 10 s med en synkronisering parameter på 20 ms og minimum spike hastigheten 0.083333 spikes per min. - Definere det planlagte opptaket for tid på 5 minutter (300.000 ms) ved å klikke på klokkeikonet nederst. Dette oppnås ved å endre informasjonen i avsnittet innstillingen å registrere hver 5.1 minutter og registrere i 5 minutter. Dette starter umiddelbart og kjøres en gang (supplerende figur 3,5).
- Kontroller at temperaturen i MEA har nådd 37 ° C før MEA. Fjerne MEA settefiskanlegg, plassere den på innspillingen enheten og låse MEA. Starte innspillingen av nettverksaktivitet ved å klikke på start posten i delen planlagt opptak eller post-ikonet i vinduet "Filen Play". En opptakstid på 5 minutter (300.000 ms) er vanligvis tilstrekkelig, men lengre opptak av opptil 10 minutter kan oppnås.
- Etter opptak, tilbake MEA til inkubator.
Merk: Omsorg må tas for å opprettholde sterilitet og minimere tiden MEA utenfor inkubator.
4. dataanalyse
- Analyse av rådata kan gjøres frakoblet ved hjelp av innspillingen programvare. Klikk på rullegardinmenyen for filen og åpne opptak. Velg filen rådataene som skal analyseres. Spill rå innspillinger for å ta de nødvendige spiking parametrene.
- For å oppnå gjennomsnittlig antall av toppene, bruke modulen Spike detektor og for å oppnå synkronisering indeksen, bruke modulen Neural statistikk kompilatoren.
- Laste Spike detektor, Burst detektor og statistikk kompilatoren moduler som før (del 3).
- Rullegardinmenyene i Spike detektor, brast detektor og statistikk kompilatoren modulen windows Velg Spike liste, brast nettverkslisten og avanserte beregninger, henholdsvis.
- Klikk på opptaksknappen for å starte innspillingen datafilen som vil skape en CSV-fil i mappen rettet. Parameterne spike - totalt antall spisser og Synchrony indeks - registreres i CSV-filen.
Merk: sanntids analyse mens rådata anbefales ikke som dette kan oppta prosessortid og beregningsressurser.
Representative Results
Vi undersøkte effekten av BPA på synkronisering utvikling i chick embryoet neuronal kulturer. Synkronisering av skyter aktivitet er en indikator for normal utvikling av nevrale nettverk i vitro. Når to spikes oppstår innen 5 ms av hverandre, kan det anses de synkrone. In vitro neuronal kulturer i utgangspunktet vise tilfeldig skyter aktivitet-toppene oppstår tilfeldig og intervallene mellom spike viser en normalfordeling. Som kulturer eldre, spiking aktiviteten blir mer synkronisert og intervallene mellom pigg er konstant. Synkron skyter aktiviteten til en kultur var quantitated av kryss-korrelasjon analyse av spike timene benytter innspillingen programvare12 for å utlede en synkronisering indeks (SI). Vi fant at BPA eksponering negativt påvirker nettverk aktivitet utvikling ved å redusere skyter aktivitet og synchrony indeksen (figur 2en og 2b). I sammendraget, har vi vist at de skadelige virkningene av BPA eksponering på et embryo påvirker utviklingen, inkludert neuronal funksjon som kan vurderes gjennom synkronisering utvikling i chick nerveceller i kultur.

Figur 1: Skjematisk av protokollen som viser Disseksjon av chick forebrain trinnene til dissosiasjon, plating, og opptak av nettverksaktivitet. Klikk her for å se en større versjon av dette tallet.

Figur 2: (A) gjennomsnittlig antall toppene er betydelig lavere i BPA behandlet E7 chick forebrain kulturer sammenlignet med kontrollen kulturer. Gjennomsnittlig antall toppene ble hentet ved hjelp av innspillingen programvare. Midler (kontroll, 14,890 ± 949.0, N = 15 og BPA, 5,624 ± 465.9, N = 22) var signifikant forskjellig (t-test for kortet, p < 0,0001). (B) gjennomsnittlig synkronisering indeksen (SI) er betydelig lavere i BPA behandlet E7 chick forebrain kulturer sammenlignet med kontrollen kulturer. SI ble hentet ved hjelp av Neural statistikk kompilatoren modulen i innspillingen programvare. SI midler (kontroll, 0.5159 ± 0.06547 N = 15 og BPA, 0.1140 ± 0.01840 N = 22) var signifikant forskjellig (t-test for kortet, p < 0,0001). Feilfelt viser standardavviket. Klikk her for å se en større versjon av dette tallet.
Discussion
Rask vekst og utvikling som embryogenesis er særlig sårbare for de skadelige virkningene av ulike endokrine-forstyrre forbindelser inkludert BPA. Vi gir detaljert protokoller for å studere virkningene av BPA eksponering i en i vitro virveldyr nevrale nettverksmodell. Bruker denne protokollen, etablerte vi at BPA senker spike hastigheten som synkronisering indeksen i utviklingsland nevrale nettverk (figur 2AB).
Vår metode ble utviklet for å studere BPA eksponering under tidlig embryogenesis og lett kan tilpasses for å studere virkningene av andre EDCs. Embryonale neuronal kulturer fra dama er relativt enkelt å etablere forhold til andre virveldyr modeller, inkludert rotte eller mus. Dessuten, det er ingen krav til et eget dyr rom-en enkel inkubator på en laboratoriebenk er tilstrekkelig til å gjennomføre disse analyser. Vi beskrive bruken av et multi elektrode system (MEA) for å vurdere effekten av EDC, BPA, på nettverksaktivitet. Protokollene beskrevet her kan brukes på andre systemer. En viktig del av denne protokollen er opprettholdelse av sterilitet. Dette inkluderer Disseksjon av fosteret under sterile forhold-sterilisering instrumenter med 70% etanol og bruk av steril Hanks balansert salter løsningen er tilstrekkelig. Som dataene samles løpet av uker, er det viktig å opprettholde steril håndtering av tiltak. Neuronal kulturer når etablert kan kultivert langsiktig - opp til tre måneder, og er derfor nyttig for studerer langsiktige eksponering for EDCs og andre forbindelser på nevrale nettverk. En annen viktig del av denne protokollen er tiden fra disseksjon til plating. Det optimale tidspunktet er 30 min, inkludert inkubasjonstiden for vevet med trypsin. Lengre tid tatt, jo dårligere overlevelse av cellene.
Her beskriver vi en grunnleggende protokoll som kan bli brukt til vurdering av alle kjemiske eller stoff som påvirker virkemåten og nevrale funksjon. Her har vi brukt en celle tetthet av 2200 celler per mm2; men dette kan endres og andre cellen tettheter kan brukes. Generelt, har vi funnet at økende celle tetthet øker nettverksaktivitet - mer toppene på kortere tid. Metoden beskrevet, svært nyttige i vurdering effekter av kjemikalier på nettverksaktivitet har begrensninger. En av de største begrensningene av metoden er at disse i vitro kulturer er to dimensjonale ikke gjenspeiler den tredimensjonale arkitekturen av virveldyr hjernen. Dette kan løses ved hjelp av skive innspillinger. Et annet alternativ er gjelder behandlinger for utvikle chick fosteret ved betyr av et lite vindu kuttet i den brede enden av egget15, dissekere hjernen på slutten av behandlingsregime gjøre tykke snitt på en vibratome og plasser på MEA for opptak nettverksaktivitet.
Våre protokollen er viktig fordi det gir undersøkelse av effekten av EDCs på utviklingen av nettverksaktivitet og tilbyr utforskningen av mekanistisk grunnlag for effekten av disse kjemikaliene.
Disclosures
Forfatterne erklærer at de har ingen konkurrerende interesser.
Acknowledgments
Denne studien støttes av NSF (HBCU-UP forskerpris innvielsen, HRD 1401426 og EPSCoR EPS-0814251) og NIH (COBRE 1P20GM103653-01A1). K.S. støttes av et fellesskap fra Delaware INBRE-III 6404.
Materials
| Name | Company | Catalog Number | Comments |
| #5 foreceps | Fine Science Tools | 11251-10 | |
| Axion Muse MEA | Axion Biosystems | M64-GL1-30Pt200 | Will be called MEA system in manuscript |
| Axis Software | Axion Biosystems | Will be called recording software in the manuscript | |
| BPA | Sigma-Aldrich | 239658-250g | |
| curved forceps | Fine Science Tools | 11272-50 | |
| EtOH | Sigma-Aldrich | 64-17-5 | |
| Fertilized chicken eggs | from any local farm or Spafas | ||
| HBSS | Fisher | 14170112 | |
| Hemacytometer | Fisher | 02-671-6 | |
| Matrigel Growth Factor Reduced, Phenol Red-Free | BD Biosciences | 356231 | Will be called Extra Cellular Matrix (ECM) in the manuscript |
| Neurobasal medium | BrainBits | Nb4-500 | |
| Neuroexplorer statistical software | Nex Technologies | Neuroexplorer version 5 | |
| Pasteur pipettes | Fisher | 13-678-20A | |
| spring scissors | Fine Science Tools | 15514-12 | |
| Sylgard bottom dissection dishes | Living Systems Instrumentaion | DD-90-S-BLK-3PK | |
| Trypan Blue dye | Fisher | 15-250-061 | |
| Trypsin-EDTA | Fisher | 15400054 |
References
- Anahara, R., Yoshida, M., Toyama, Y., Maekawa, M., Masayuki, K., Ishino, F., Toshimori, K., et al. Estrogen agonists, 17 beta-estradiol, bisphenol A, and diethylstilbestrol, decrease cortactin expression in the mouse testis. Arch. of Histol. Cytol. 69 (2), 101-107 (2006).
- Dodds, C. Synthetic oestrogens. Br. Med. Bull. 11 (2), 131-134 (1955).
- Grignard, E., Lapenna, S., Bremer, S. Weak estrogenic transcriptional activities of Bisphenol A and Bisphenol S. Toxicology In Vitro. 26 (5), 727-731 (2012).
- Herbst, A. L., Ulfelder, H., Poskanzer, D. C. Adenocarcinoma of the vagina: association of maternal stilbestrol therapy with tumor appearance in young women. New Eng. J. Med. 284, 878-881 (1971).
- Jenkins, S., Raghuraman, N., Eltoum, I., Carpenter, M., Russo, J., Lamartiniere, C. A. Oral exposure to bisphenol A increases dimethylbenzanthracene-induced mammary cancer in rats. Environ Health Persp. 117 (6), 910-915 (2009).
- Okada, A., Kai, O. Effects of estradiol-17beta and bisphenol A administered chronically to mice throughout pregnancy and lactation on the male pups' reproductive system. Asian J Androl. 10 (2), 271-276 (2008).
- Palanza, P., Gioiosa, L., Vom Saal, F. S., Parmigiani, S. Effects of developmental exposure to bisphenol A on brain and behavior in mice. Environ. Res. 108 (2), 150-157 (2008).
- Mimoto, A., Fujii, M., Usami, M., Shimamura, M., Hirabayashi, N., Kaneko, T., et al. Identification of an estrogenic hormone receptor in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 364 (4), 883-888 (2007).
- Allard, P., Colaiacovo, M. Bisphenol A impairs the double-strand break repair machinery in the germLine and causes chromosome abnormalities. Proceedings Natl. Acad. Sci. 107 (47), 20405-20410 (2010).
- Mersha, M. D., Patel, B. M., Patel, D., Richardson, B. N., Dhillon, H. S. Effects of BPA and BPS exposure limited to early embryogenesis persist to impair non-associative learning in adults. Behav. Brain Funct. : BBF. 11, 27 (2015).
- Chen, Y., Shu, L., Qiu, Z., Lee, D. Y., Settle, S. J., et al. Exposure to the BPA-Substitute Bisphenol S Causes Unique Alterations of GermLine Function. PLOS Genetics. 12 (7), 1006223 (2016).
- Shein Idelson, M., Ben-Jacob, E., Hanein, Y. Innate Synchronous Oscillations in Freely-Organized Small Neuronal Circuits. PLoS ONE. 5 (12), 14443 (2010).
- Baltz, T., Herzog, A., Voigt, T. Slow Oscillating Population Activity in Developing Cortical Networks: Models and Experimental Results. J. Neurophysiol. 106 (3), 1500-1514 (2011).
- Pettmann, B., Louis, J. C., Sensenbrenner, M. Morphological and Biochemical Maturation of Neurones Cultured in the Absence of Glial Cells. Nature. 281 (5730), 378-380 (1979).
- Zhang, H., Wu, C. Y., Wang, W., Harrington, M. A. Interneuronal Synapses Formed by Motor Neurons Appear to Be Glutamatergic. NeuroReport. 22 (16), 809-813 (2011).
- Paiva, A. R. C., Park, I., Príncipe, J. C. A Comparison of Binless Spike Train Measures. Neural Comp. and Appl. 19 (3), 405-419 (2010).
- Cretu, A., Fotos, J. S., Little, B. W., Galileo, D. S. Human and Rat Glioma Growth, Invasion, and Vascularization in a Novel Chick Embryo Brain Tumor Model. Clin Exp Metastasis. 22, 225-236 (2005).
- Chiappalone, M., et al. Dissociated cortical networks show spontaneously correlated activity patterns during in vitro development. Brain Res. 93, (2006).
- Li, X., et al. Long-term recording on multi-electrode array reveals degraded inhibitory connection in neuronal network development. Biosens Bioelectron. 22 (7), 1538-1543 (2007).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}