

Summary
합성 생물학 비정규 아미노산의 co-translational 삽입을 사용 하 여 전례 없는 속성으로 단백질의 엔지니어링을 수 있습니다. 여기, 우리는 어떻게 소설 형광 분 광 속성, "금" 형광 성 단백질 (GdFP), 불리는 GFP 형 fluorophore의 spectrally 레드 이동 변종 생산 대장균 에서 선택적 압력 관 (SPI)를 통해 제시.
Abstract
형광 단백질의 살아있는 세포 형광 현미경 검사 법에 대 한 특히 생명 과학에 대 한 기본적인 도구 이다. 야생-타입 및 설계 변형 Aequorea 빅토리아 (avGFP)에서 녹색 형광 단백질의 다른 종에서 homologs 이미 광학 스펙트럼의 큰 부분을 커버, 스펙트럼 갭 남아 근처-적외선 영역에 대 한 어떤 avGFP 기반 fluorophores 사용할 수 없습니다. 형광 단백질 레드 이동 (FP) 이체 실질적으로 여러 분자 종의 스펙트럼 unmixing 위한 툴킷 확장 하지만 산호에서 파생 자연스럽 게 발생 레드 이동 FPs 또는 말미 낮은 형광 양자 수율 및 열 등 한 avGFP 변종에 비해 사진-안정성. 더 조작 및 빨강 스펙트럼 영역 쪽으로 발 색 단의 활용된 시스템의 가능한 확장 또한 유전자 코드를 정한 20 표준 아미노산의 레 퍼 토리에 의해 제한 됩니다. 이러한 한계를 극복 하기 위해 합성 생물학 더 스펙트럼 적색 이동 발 색 단 깡패로 비 정식 아미노산의 삽입을 통해 얻을 수 있습니다. SPI의 응용 프로그램 엔지니어 avGFP 변종 소설 스펙트럼 속성을 설명합니다. 트립토판 auxotrophic 대장균 에서 단백질 표정 수행 긴장 및 적당 한 indole 선구자와 성장 매체를 보완 하 여. 셀 안에이 일어나 해당 트립토판 아날로그로 변환 하 고 응답 UGG codons ribosomal 기계에 의해 단백질으로 통합. AvGFP (ECFP)는 전자 기증 4 aminotryptophan에 의해 향상 된 "청록색" 변종에서 Trp-66의 교체 GdFP 108 nm Stokes shift와 강하게 레드 이동 방출 최대 결과 (574 nm), 보다는 열역학으로 안정 되어 있는 동안 그것의 전임자 ECFP입니다. 정식이 아닌 아미노산의 잔류물 관련 법인 질량 분석에 의해 분석 된다. GdFP의 광 속성 생명 과학에서 유전으로 인코딩된 프레임의 중요 한 응용 프로그램 중 하나로 시간 해결 형광 분광학에 의해 특징.
Introduction
형광 단백질 GFP 가족의 해파리 Aequorea 빅토리아 19621 (avGFP)에서 녹색 형광 성 단백질 및 다른 진 핵 세포에서 19942 에 첫 번째 분리 식의 발견 이후 되고있다 매우 귀중 한 도구 및 생명 과학에서 목표입니다. 광범위 한 유전과 분자 공학 포함 종의 코 돈 사용의 조정, 접는, 향상 된 성숙, 증가 광도, oligomerization의 예방과 스펙트럼 및 광화학 속성의 조정의 가속 역 photoswitch3,,45,6기능을 포함 하 여. GFP 빚에서 그것의 4-(p-hydroxybenzylidene)의 형광 발 색 단 imidazolidin-5-1 (HBDI). 후자는 autocatalytically에서 형성 아미노산 (Ser-65/Tyr-66/Gly-67 avGFP에) 소위 발 색 단 깡패 분자 산소7의 영향 아래 펩 티 드 등뼈에 내에서 추가 공유 결합의 형성. Resonantly 안정된 활용된 시스템 표시 범위와 특성 녹색 형광이이 단백질의 흡수에 대 한 수 있도록 분자 환경으로 동적으로 작용 합니다.
발 색 단 깡패 내 방향족 아미노산의 존재는 필수입니다. 그러나, 산 성 아미노산 표준 레 퍼 토리 4 향기로운 잔류물 (그의 페, Trp, Tyr) 구성 되어 있습니다. 이 실질적으로 더 많은 레드 이동 avGFP 변종 Discosoma 풀 coralimorphs에서 DsRed8 등 mKate/mNeptune9 에서 가장 레드 이동 자연 FPs 기준으로 달성 하기 위해 기존의 mutagenesis 접근 제한 바다 곰 Entacmaea quadricolor 따라서, 600 이상 광학 스펙트럼의 빨강 및 가까운 적외선 부분 nm 띄엄띄엄 GFP 변종으로 덮여 있다. 이것은, 물론, 스펙트럼 디 멀티플렉싱 여러 fluorophore 종의 동시에 필요로 하는 형광 현미경 접근에 대 한 심한 제한. 예를 들어 긴 파장 마커는 또한 만들 필요가10을 이미징 하는 깊은 조직에 대 한 설정에서 700-1000 nm 사이 피부 조직의 낮은 흡수 정권의 사용.
AvGFP에서 파생 된 형광 단백질 분 광 속성 및 그들의 chromophores11의 화학 특성에 따라 여러 가지 종류로 나누어집니다. 야생-타입 발 색 단 중립, 페 놀 폼 사이 equilibrated 혼합물으로 존재 하는 그것의 깡패 Ser-65/Tyr-66/Gly-67와 (λ최대 = 395 nm, ε = 21000 M-1c m-1)와 음이온 phenolate 양식 (λ최대 = 475 nm, ε 7100 M = -1c m-1), 그리고 방출 스펙트럼 전시 508에서 단일 피크 nm. Ser-65의 수 산 기 그룹은 중요 한 중요성의 Glu-222 발 색 단 주변에는 H-본드 기부 (거리: 3.7 Å),이 카복실산의 이온화를 승진 시키는. 클래스는 음이온 phenolate 발을 색 단, EGFP 같이 의해 특징입니다 (페-64-레이/Ser-65-Thr; λ최대 = 488 nm, ε = 35600 M-1c m-1, λem = 509 nm). Ser-65-Thr(Ala,Gly) 대체 때문에 중립 페 놀 폼의 395 nm 여기 피크 억제와 음이온 phenolate의 470-475 nm 피크는 5-에 six 배 향상 및 490 이동 nm. 클래스 II에는 중성 페 놀 발 색 단, 사파이어 GFP로 단백질 구성 되어 있습니다. Thr-203-일 대체 399에만 피크를 떠나 475 nm 여기를 억제 하는 여기에 거의 완전히 nm. 음이온 발 색 단 solvated 제대로 될 수 없습니다, 때문에 중립적인 형태로 선호 됩니다. 클래스 III 구성 "노란색" 형광 변종 (EYFP; Ser-65-Gly/Val-68-Leu/Ser-72-Ala/Thr-203-Tyr; Λ최대 ε = 514 nm, ε = 84, 600 M-1c m-1, λem = 527 nm) 상호 π 스태킹 향기로운 측면 체인과 phenolate, Thr-203-His(Trp,Phe,Tyr) 대체에 의해 초래로, 이끌어 내는 최대 20 nm 레드 이동 방출 년 맥시 마 (Thr-203-Tyr) 추가 대체 (Gln-69-리스) 결과 다른 1-2 nm 적색 이동 529 nm,11알려진 가장 레드 이동 avGFP 변종. Indole (Tyr-66-Trp)에 대 한 페 놀의 교환을 만듭니다 청록색 형광 ECFP 처럼 클래스 IV, (Ser-65-세/Tyr-66-Trp; λmax1 = 434 nm, ε = 24,800 M-1c m-1; λmax2 452 = nm, ε = 23,600 M-1c m-1 ; Λem1 477 = nm, λem2 = 504 nm). 부피가 큰 indole 숙박 아마 다른, 보상 돌연변이 의해 활성화 됩니다. ECFP 여기 및 방출 맥시 마가 inbetween 중립 또는 음이온 chromophores 단백질의. 클래스 V 단백질 항구는 석탄 대신 이미 (Tyr-66-그의), 예를들면., 블루 형광 단백질 같은 EBFP. 클래스 VI 따라서 가장 블루 이동 여기 및 방출 피크 위치에 이르게 한 페 놀-페 닐 exchange 독점적으로, 중립 발 색 단 양식 선호에 의해 생산 됩니다 (360 nm와 442 nm, 각각).
클래식 사이트 감독 mutagenesis 변종의 소설 avGFP 발 색 단, 65-67 tripeptide 및 20 표준 아미노산의 프레임에 상호 작용 잔류물의 순열에 의해 생산을 위해 특히 적당 하다. 향기로운 아미노산의 비정규 변종 ribosomal 단백질 합성12동안 소개 될 때 이러한 가능성은 더욱 확장할 수 있습니다. 원칙적으로,이 두 가지 방법이 있다. 첫 번째 전략 단백질 번역 기계, 특히 관련된 아미노산 아날로그 쪽으로 aminoacyl-tRNA synthetases (aaRSs)의 기판 허용 오차에 의존합니다. 이를 위해 고효율, auxotrophic 대장균 식 변종 해당 천연 아미노산을 합성할 수 있는 고용. 문화 매체에 그 적당 한 정식이 아닌 아미노산 (ncAAs) 또는 전조를 추가 하 여 후자의 교체 수 있습니다. 이 전략로 알려진 또한 선택적 압력 관 (SPI)13,14, 잔류물 특정 교체를, ncAA의 글로벌 관에 결과 수 있습니다. 두 번째 전략 사용 정지 codon에 의해 ncAA 혐의로 억압 tRNAs aaRS 효소 설계. 이 프레임에서 정지 codons의 쓰루가 발생 하며 사이트별 ncAA 설립 따라서, 정지 codon 억제 (SCS)의이 방법은 유전자 코드15의 확장에 지도 한다. 통해 mutagenesis, 정지 codon 원하는 사이트에서 대상 유전자에 배치 됩니다. 원칙적으로, SPI 또한 메트로 등 Trp 드문 정식 아미노산 대체에 대 한 선택은 그에 주어진 재조합 펩 티 드 및 단백질 베어링 독특한 미식 축구 설치를 만드는 데 사용할 수 있습니다. Trp와 SPI 접근 아날로그 4-F-,-F-및 6-F-Trp, 7-아 자-Trp, 4-오-그리고 5-오-Trp, 뿐 아니라 4-, 5, 5-NH2등의 큰 다양 한와 함께 작동 하도록 표시 되었습니다-Trp 또는 심지어 β (thienopyrrolyl) 알라닌 파생 상품16 17,18,,1920. 따라서, SPI는 정식이 아닌 변종 맞게 더 스펙트럼 및이 FPs의 스톡 변화 가능성을 탐구 하 여 GFP chromophores의 향기로운 아미노산을 교체에 대 한 매우 유리한 될 수 있습니다. 모든 단백질 시퀀스 수정에 관해서는 FP 폴딩 및 발 색 단 성숙과 호환성은 실험적으로 테스트 되어야 한다.
이 작품에서는, 우리는 IV ECFP21, 야생-타입 avGFP Tyr, Trp 잔류물의 발 색 단 깡패 내 대신 운반 클래스를 이용 한다. SPI를 사용 하 여이 Trp-66 (및 Trp-57만 다른 Trp 잔류물 ECFP에) 4-아미노-Trp.에 의해 대체 됩니다. 4-아미노-Trp는 발 색 단 내에서 전자 기부 아미노 그룹의 존재 상태까지 레드 이동 양성자 전송 (ESPT) 108 nm Stokes shift 부여의 공명 안정화를 하시 더군요. 이 "금" 형광 성 단백질 (GdFP) 구성 최대 형광의 가장 큰 레드 시프트와 변종 (574 nm) 모든 avGFP 파생 단백질 사이. 우리 SPI에 의해 GdFP 단백질 생산 방법에 설명 하 고 질량 분광학에 의해 결과 수정된 단백질의 필수 분석을 위한 프로토콜을 제공 합니다. 또한, 우리 GdFP 수 활용 및 시간 해결 형광 분광학 접근에 분석 하는 방법을 보여줍니다.
Protocol
1. Trp auxotrophic 대장균 의 변화
- 화학적으로 변형 또는 electrocompetent Trp auxotrophic E. 콜라이 긴장, 예의 (50 µ L) 세포. ATTC 49980 (WP2, 돌연변이 E. 콜라이 긴장 B/R22에서 파생), 열 충격 또는 electroporation, 각각 사용 하 여 pQE-80 L His6 ECFP 플라스 미드의 1 ng / µ L 수성 해결책의 1 µ L와 함께. 자세한 내용은 정돈 과학 교육 데이터베이스23,24 를 참조 하십시오.
참고: 표정 벡터 pQE-80 L His6-ECFP 인코딩합니다는 N-말기 6 그의 태그 ECFP21 x 락 연산자 세균성 T5 발기인에 의해 구동. 복제의 colE1 유래와 앰프R 선택 마커 추가 운반 (pQE-80 L 벡터 백본 시퀀스에서 찾을 수 있습니다: https://www.qiagen.com/mx/resources/resourcedetail?id=c3b71572-4d82-4671-a79b-96357fe926d1&lang=en & 자동 제안 = true). (후 발 색 단 성숙25) His6 ECFG 야생-타입 단백질의 이론적인 분자량은 28303.92 다. 번역된 대상 단백질 순서는 (그의 태그 밑줄, 굵게 벡터 파생 시퀀스) 다음과 같이: MRGSHHHHHHGSMVSKGEELFTGVVPILVELDGDVNGHKFSVSGEGEGDATYGKLTLKFICTTGKLPVPWPTLVTTLTWGVQCFSRYPDHMK
QHDFFKSAMPEGYVQERTIFFKDDGNYKTRAEVKFEGDTLVNRIELKGIDFKEDGNILGHKLEYNYISHNVYITADKQKNGIKANFKIRHNIEDGS
VQLADHYQQNTPIGDGPVLLPDNHYLSTQSALSKDPNEKRDHMVLLEFVTAAGITLGMDELYK입니다. - 10 g/L 포도 당, 100 µ g/mL 암 피 실린 보충 파운드-한 천 배지 (표 1)에 변환 된 셀 접시 하 고 37 ° C에서 접시를 밤새 품 어.
2. 재조합 형 단백질 표정
-
대장균 ATCC 49980 pQE-80 L His6-ECFP의 숙박 문화
- 파운드 매체의 5 mL을 준비 (표 1; 10 g/L 포도 당, 100 µ g/mL 암 피 실린 보충) 살 균 14 mL에 폴리스 티 렌 에어로빅 성장을 위한 튜브 문화와 함께 접종 한 메 마른 피 펫 팁 이나 접종 루프를 사용 하 여 한 천 배지에서 홀로 식민지.
참고: 갓 변환 된 셀에서 식민지를 사용 하 여 것이 좋습니다. (단계 1.2.)에서 세균성 식민지와 접시 4 ° C에서 며칠 동안 저장할 수 있습니다. - 200-250 rpm에서 궤도 셰이 커에서 37 ° C에서 세포를 밤새 품 어.
- 파운드 매체의 5 mL을 준비 (표 1; 10 g/L 포도 당, 100 µ g/mL 암 피 실린 보충) 살 균 14 mL에 폴리스 티 렌 에어로빅 성장을 위한 튜브 문화와 함께 접종 한 메 마른 피 펫 팁 이나 접종 루프를 사용 하 여 한 천 배지에서 홀로 식민지.
-
야생-타입 ECFP의 표현
- 10 mL 신선한 파운드 매체 접종 (표 1; 10 g/L 포도 당, 100 µ g/mL 암 피 실린 보충) 100 µ L 100 mL 삼각 플라스 크에에서 하룻밤 문화의와 함께. 37 ° c 200 rpm에서 궤도 셰이 커에서 플라스 크를 품 어.
참고: 필요에 따라이 단계에서에서 수행할 수 10 mL NMM19 매체 (표 1) 100 µ g/L 암 피 실린과 0.5 m m L-트립토판 보충 (또는, indole 사용할 수 있습니다). - 600에서 광학 밀도 측정 nm (OD600) 20 분 마다 우선적으로 측정 셀 밀도 600에서 멸종 확인 하 여 nm (OD600) 항상 경로 길이가 1 cm.의 베트를 사용 하 여 분 광 광도 계에 대 한 참조를 수행 해당 문화 매체를 사용 하 여 측정입니다. 샘플 및 샘플 0.1-0.8의 측정 값을 가져오기 위해 그럼 OD600 희석 요소를 사용 하 여 계산 혼합 희석. 자세한 내용은 이전 게시 26를 참조 하십시오.
- OD600 값이 0.5-0.8 (는 접종 후 약 2-3 h)에 도달, SDS 페이지 (나트륨 라우릴 황산 polyacrylamide 젤 전기 이동 법, 4 단계)에 대 한 "유도" 전에 샘플을 가져가 라.
- 0.5 m m IPTG (이소프로필 β-D-1-thiogalactopyranoside, 1 M 재고 솔루션에서) 액체 문화를 조정 하 여 대상 단백질 발현을 유도 하 고 4-8 h 200 rpm에서 궤도 통에 30 ° C에서 품 어.
참고: 청록색 형광 단백질2737 ° C 이하의 온도에서 일반적으로 표현 됩니다. - SDS 페이지 (단계 4)에 대 한 "후 식" 샘플을 가져가 라.
- 5000 x g와 4 ° C에서 10 분 동안 원심 분리 하 여 세균 세포를 수확
- Decanting 여는 상쾌한을 삭제 하 고 대상 단백질 정화까지-20 ° C 또는-80 ° C에서 셀 펠 릿을 동결.
- 10 mL 신선한 파운드 매체 접종 (표 1; 10 g/L 포도 당, 100 µ g/mL 암 피 실린 보충) 100 µ L 100 mL 삼각 플라스 크에에서 하룻밤 문화의와 함께. 37 ° c 200 rpm에서 궤도 셰이 커에서 플라스 크를 품 어.
-
SPI GdFP 생산
- 10 µ L 100 mL 삼각 플라스 크에에서 하룻밤 문화의와 100 µ g/mL 암 피 실린, 15 µ M 트립토판 보충 NMM19 매체 (표 1)의 10 mL를 접종 하 고 하룻밤 200 rpm에서 궤도 통에 30 ° C에서 문화 플라스 크를 품 어.
참고: 대장균 및 SPI의 재배에 대 한 화학적으로 정의 된 미디어의 다양 한 사용할 수입니다. NMM 여기서, 뿐만 아니라 MOPS 중간28, 포도 당-미네랄 소금 중간29, 데이비스 최소 중간30, M9 최소 중간31또는 GMML32 를 사용할 수 있습니다. - 다음 날, 값만 30 분 이상 0.05 미만으로 변경 될 때까지 세600 매 30 분을 측정 합니다. 대 지 값 약 1 이어야 한다입니다.
참고: 편차 ± 0.3 단위로 사용할 수 있습니다. 세균성 긴장 매체 사용에 따라, 초기 트립토판 농도 (단계. 2.3.1) 조정을 할 수 있습니다. - SDS 페이지 (단계 4)에 대 한 "유도" 전에 샘플을 가져가 라.
- 5000 x g와 4 ° C에서 10 분 동안 원심 분리 하 여 세균 세포를 수확 Decanting 여는 상쾌한을 삭제 합니다.
- 100 mL 삼각 플라스 크에 10 ml 100 µ g/mL 암 피 실린을 NMM19 매체의 셀 resuspend와 50 m m 재고 솔루션을 사용 하 여 1 m m의 최종 농도 4-아미노-indole 추가. 200 rpm에서 궤도 통에 30 ° C에서 30 분 동안 부 화를 계속 합니다.
참고:이 단계는 암 피 실린의 낮은 화학적 안정성 권장 하 고 4-아미노-indole의 세포질 통풍 관을 보장. - IPTG의 0.5 m m 1 M 재고를 사용 하 여 최종 농도를 추가 하 여 대상 단백질 발현을 유도 하 고 200 rpm에서 궤도 통에 30 ° C에서 하룻밤 샘플을 품 어.
참고: 청록색 형광 단백질2737 ° C 이하의 온도에서 일반적으로 표현 됩니다. - 다음 날, OD600을 측정 합니다.
- SDS 페이지 (단계 4)에 대 한 "후 식" 샘플을 가져가 라.
- 5000 x g와 4 ° C에서 10 분 동안 원심 분리 하 여 세균 세포를 수확 하 고 상쾌한 decanting 의해 삭제.
- 이러한 선박 원심 분리에 사용 하지 않은 경우에 주걱을 사용 하 여 50 mL 원뿔 폴리스 티 렌 튜브에 셀 펠 릿을 전송 합니다. 대상 단백질 정화까지-20 ° C 또는-80 ° C에서 셀 펠 릿을 고정 합니다.
- 10 µ L 100 mL 삼각 플라스 크에에서 하룻밤 문화의와 100 µ g/mL 암 피 실린, 15 µ M 트립토판 보충 NMM19 매체 (표 1)의 10 mL를 접종 하 고 하룻밤 200 rpm에서 궤도 통에 30 ° C에서 문화 플라스 크를 품 어.
3. 대상 단백질 정화 고정된 금속 이온 친화성 크로마토그래피 (IMAC)을 통해
-
세균성 세포 세포의 용 해
- 10-20 분 동안 얼음에 셀 펠 릿 녹여
- 얼음에 얼음 처럼 차가운 바인딩 버퍼 (표 1)의 5 mL을 사용 하 여 50 mL 원뿔 폴리스 티 렌 튜브에 셀 펠 릿을 resuspend.
- 20 µ L의 50 mg/mL lysozyme, 1 mg/mL DNase I 및 1 mg/ml RNase A. 20 µ L 튜브, 5 번, 반전으로 부드럽게 혼합 닫고 30 분 동안 얼음에 그것을 유지의 20 µ L를 추가 합니다.
참고: 일부 셀 중단 lysozyme에 의해 촉매로 발생 합니다. - Sonification 15 mL 폴리스 티 렌 튜브 2 비 자금 얼음으로 냉각의 3 분의 3 주기를 사용 하 여 초음파 균질 화기 팁을 사용 하 여 세포를 lyse s 펄스, 4의 일시 중지 및 45% 진폭의 s.
참고: 또는, 고압 균질 사용할 수 있습니다, 예., 14000 psi에서 20 주기. 필요한 경우 최소한의 악기 볼륨에 도달 바인딩 버퍼를 사용 하 여 희석. 또한, 단백질 추출 시 약은 세포 파쇄에 대 한 사용할 수 있습니다. 예제를 보려면 참조 자료 테이블. - 15000-18000 x g, 4 ° c.에 30 분 동안 원심 분리기 샘플
- 신선한 튜브와 액체의 볼륨 노트는 상쾌한을 전송 합니다.
- 5 mL 플라스틱 Luer 잠금 주사기와 polyvinylidene 불 소 (PVDF) 주사기 필터를 사용 하 여 0.45 μ m 주사기 필터를 통해 솔루션을 필터링 합니다.
- SDS 페이지 (단계 4)에 대 한 "lysate" 샘플을 가져가 라.
- 세포 파편 펠 릿 ddH2O (동일한 볼륨으로 전 lysate) resuspend
- SDS 페이지 (단계 4)에 대 한 샘플 "펠 릿"을 가져가 라.
-
아이맥 정화
- Prepacked 또는 제조업체의 지침에 따라 아이맥 FPLC (고속 단백질 액체 크로마토그래피) 열 자동 포장 1 mL을 사용 합니다. 바인딩 버퍼 (표 1)를 사용 하 여 열에 세포 lysate를 적용 한 후 다음과 같이 세척 단계 뿐만 아니라 열 평형에 대 한.
- 수집 하 고 표시 빛 황금 색깔에 의해 확인 될 수 있다 GdFP와 eluate 분수 수영장.
참고: 필요에 따라 대상 단백질 수 수 eluted 이미 선형 그라디언트를 사용 하 여 (0-250 m m) 자동된 FPLC 시스템을 사용 하 여. - 466에서 소멸 계수에 대 한 문학 값을 사용 하 여 단백질 농도 결정 nm (ɛ466 nm = 23,700 M-1 c m-1)33 참조로 차입 버퍼. 에 대 한 내용은 절차, 이전 게시26를 참조 하십시오.
- SDS 페이지 샘플 "eluate" 고 Coomassie 얼룩 경우 레인 당 단백질의 1-10 µ g를 사용 하 여.
참고: SDS 샘플 금액 착 방법 및 염료 감도 따라 달라질 수 있습니다. - 투 석 버퍼 또는 분자 무게 컷오프 (MWCO) 5000-10, 000의 멤브레인을 사용 하 여 MS 버퍼 eluate 분수의 약 수 dialyze 제조업체의 지침에 따라 투 석 막 준비. 3 회 2 시간 이상에 대 한 100 mL 버퍼에 대 한 1 mL 샘플 dialyze 대 한 자세한 내용은이 절차, 이전 게시34를 참조 하십시오.
- 저장용, 동결-80 ° c.에 투 석 버퍼에 단백질 샘플
참고: Aliquots 6 개월 이상 안정 해야 합니다.
4. SDS 페이지 샘플 준비 대장균 전체 세포 추출 물
- OD600 의 1 mL에 해당 셀 정지 전송 = 1 정지 (예. OD600500 µ L = 2) 1.5 mL microcentrifuge 관으로.
- 5000 x g, 실 온에서 10 분 동안 원심 분리 하 여 세포를 수확. 상쾌한 pipetting으로 삭제 합니다.
- DdH2O의 80 µ L를 더하고 20 µ L 5 배 sds pipetting으로 셀 펠 릿을 혼합 염료 버퍼 (표 1)를 로드.
- 물 목욕 또는 열 블록에서 5 분 동안 95 ° C에가 열에 의해 세포 변성. 그 후, 실내 온도에 샘플 멋진.
- 10 µ L를 사용 하 여 Coomassie 스테인드 SDS-페이지 이전 게시35에 따르면.
참고: SDS 샘플 금액 착 방법 및 염료 감도 따라 달라질 수 있습니다.
5. 그대로 단백질 질량 분석-고성능 액체 크로마토그래피 (HPLC) 분무 이온화 시간의 비행 질량 분석 (LC-ESI-TOF-MS)을 결합
참고: HPLC 그라데이션, 설정 및 버퍼 분리 및 사용 하는 악기에 따라 달라질 수 있습니다. 모범적인 장비에 대 한 참조 자료 테이블.
- 확인 (단계 3.2.3.)를 사용 하 여 위에서 설명한 대로 MS 버퍼에 대 한 샘플에서 단백질 농도 dialyzed MS 버퍼 (자료 테이블 참조) 참고로.
- 80 µ L의 최종 볼륨에 대 한 MS 버퍼를 사용 하 여 0.1 mg/ml 단백질 샘플을 희석, 혼합 주의 pipetting 함으로써, 전송 솔루션 유리 삽입과 함께 MS autosampler 유리병으로 고 모자와 함께 그것을 닫습니다. 유리병을 터치 하 여 기포를 제거 합니다.
- MS 버퍼의 1 mL에 유리 삽입 (버퍼 공백) 없이 두 번째 autosampler 유리병을 채우십시오.
- 따 뜻 해 악기를 허용 합니다. 계기 캘리브레이션을 수행 합니다. 액체 착 색 인쇄기-학년의 충분 한 양을 사용할 수 있는지 확인 (> 100 mL).
- 80% 버퍼는 (0.1% 개미 산 ddH2O), 버퍼 B와 결합 된 5%에서 선형 20 분 HPLC 그라디언트 프로그램 (이기에 0.1% 포 름 산).
- 0.3 mL/min의 흐름에서 HPLC를 시작 하 고 열 압력 안정 될 때까지 기다립니다.
- LC ESI TOF MS 방법 5 µ L의 autosampler 주입 볼륨을 설정 하 고 실행 빈 다음 샘플 실행 한 작업을 만들 할당 해당 autosampler 유리병 위치. 작업을 실행 합니다.
- 작업 완료 후, 생성 된 샘플 데이터 파일을 엽니다. Deconvolution에 대 한 총 이온 전류 (TIC) 음모에서 범위를 선택 하 고 최대 엔트로피 deconvolution 알고리즘을 사용 하 여 MS 스펙트럼을 deconvolute.
참고: 실험 조건에 따라 추가 종 발생할 수 있습니다 비 성숙 FP에서 또는 버퍼 이온 adducts.
6. 형광 수명 측정 및 부패 관련 (DAS) 스펙트럼 GdFP
참고: 시간 해결 형광 분광학의 계측에 대 한 참조 하십시오 테이블의 자료 를 모범적인 장비. 흡 광도 형광 여기 및 형광 단백질의 방출 스펙트럼 또한 실험실 UV/Vis와 형광 분 광을 사용 하 여 기록할 수 있습니다.
-
GdFP의 파장 해결 형광 수명 측정
- 2 mL PBS 버퍼 pH 7에서 (표 1)로 희석 하 여 GdFP의 1 µ M 솔루션의 준비. 1 cm 석 영 cuvette로 솔루션을 채우십시오.
- 샘플 여기 ps 펄스 470 nm 레이저 및 488 nm 긴 패스 방출 필터를 설치 하 고 600 L/m m36 (TWCSPC) 검출기의 파장 정권 인수에 대 한 계산 시간 및 파장 상관 단일 광자의 격자를 조정합니다 500-700 nm입니다.
- 약 103 건의 소프트웨어를 계산 하는 단일 광자와 형광 감퇴 곡선의 인수 최대에서 누적 됩니다 때까지 약 200 x 103 광자/s의 수 속도에서 형광 방출을 얻을.
-
경 음악 응답 기능37 (IRF)의 측정
- PH 7에서 PBS 버퍼에 1 g/L 콜 로이드 실리 카 (~ 220 m2/g)으로 가득 1 cm 석 영 cuvette로 샘플 베트를 교체 합니다.
참고: 실리 카 서 스 펜 션 400 g/L 수성 현 탁 액을 사용 하 여 준비가 되어 있습니다. - 488 nm 긴 패스 방출 필터 및 100 x 103 카운트/s 아래에 TWCSPC 검출기에서 수 속도를 조정 삽입 회색 필터를 제거 합니다.
- 16 채널 TWCSPC 감지기의 채널 8에서에서 470 nm 광자의 수집에 대 한 격자를 조정 합니다.
- 때까지 약 10 x 103 건의 최대 방출에 누적 되는 IRF를 취득 합니다.
- 변환 형광 감퇴 곡선 및 ASCII 데이터 파일을 글로벌 IRF 피팅38 프로그램.
- 3 지 수 구성 요소와 연결 된 매개 변수로 수명의 합계의 모델에 맞는 행위 글로벌.
- 플롯 부패와 관련 된 스펙트럼 (DAS) 개별 감퇴 컴포넌트의 진폭 배포판으로 파장의 종속성에 데이터 분석 소프트웨어.
- PH 7에서 PBS 버퍼에 1 g/L 콜 로이드 실리 카 (~ 220 m2/g)으로 가득 1 cm 석 영 cuvette로 샘플 베트를 교체 합니다.
Representative Results
선택적 압력 관의 기술을 사용 하 여, Trp-66 ECFP (및 Trp-57만 다른 Trp 잔류물에 ECFP)의 발 색 단 깡패에 수로 대체 4-아미노-Trp, 그로 인하여 뚜렷한 스펙트럼 특성을 가진 레드 이동 GdFP 생성. 질량 분석은 그림 1에 표시 된 결과 함께 원하는 화학 량 론 통합 정식이 아닌 아미노산의 단백질를 설명 하기 위해 사용 되어야 한다. 나중에, 정상 상태 및 시간 및 파장 해결 형광 분광학의 pH 의존에 초점을 맞춘 GdFP fluorophore의 속성을 특성 뿐만 아니라 현미경 검사 법, 대 한 UV 흡수 분광학에서 데이터 제공은 스펙트럼입니다.
4-아미노-Trp로 ECFP에서 두 Trp 잔류물의 교환 확인, 대량 spectrometric 분석 수행 됩니다. 그림 1 GdFP의 대표 deconvoluted ESI MS 스펙트럼을 보여준다. 야생-타입 ECFP가 28,283.9 다 대량의 계산된 단백질 발 색 단 성숙 후 GdFP의 해당 질량은 28,313.9 다. GdFP의 deconvoluted ESI MS 스펙트럼 전시 주요 대량 피크 28,314.1 ± 0.1에서 10 ppm 미만으로 이론적인 값에서 성적이 상자 다. 이 SPI 통해 ncAA의 설립을 확인 분석25의이 유형에 대 한 일반적인 정확도 범위 내에서 되 고, (실험 값 야생-타입 ECFP: 28,283.7 다).
그림 2 보여줍니다 confocal 형광 이미징 현미경 검사 법 (CFIM) ECFP, EGFP, EYFP 및 GdFP PBS 버퍼에 박테리아의 물의 resuspension 따라 표현 하는 세균성 세포의. 모든 이미지는 UV 객관적이 고 레이저 여기에서 각 샘플에 대 한 동일한 에너지에 대 한 장비는 현미경에 인수 했다.
그림 3A GdFP, 항상 매우 비슷한 여기 에너지 (파장 그림2)와 함께 모니터링을 포함 한 다양 한 프레임을 표현 하는 대장균 박테리아의 CFIM 이미지의 오버레이로 표시 합니다. 그림 3B 표시 FP 이체의 발 색 단 구조를 보여 줍니다. ECFP에 비해 GdFP의 밝기에 대해서 (형광 양자 수율 φ = 0.4), EGFP (φ = 0.6) 및 EYFP (φ = 0.6) GdFP, 형광 빛의 광범위 한 수집 범위에 대 한 주의 하는 것이 중요 하다 (30 nm) 20 달리 사용 되었다 nm 다른 spe에 대 한 사용 cies, 비슷한 값을 이미지의 강도 조정 하기 위하여. 약간 낮은 소멸 계수와 독특한 photophysical 속성의 결과로 감소 양자 수율 GdFP의 밝기 표시 다른 FPs에 비해 낮은입니다.
ECFP (그림 3C)의 흡수 스펙트럼 434에서 두 특성 맥시 마는 nm와 452 nm. 반면, GdFP 466에 최대 하나의 광범위 한 레드 이동 흡수 밴드 특징 이다 nm. EGFP의 흡수는 더 레드-이동 488 nm. 그러나 GdFP의 훨씬 더 큰 Stokes shift 때문에, (108 nm) ECFP에 비해 (41 nm) 및 EGFP (20 nm), GdFP의 방출 스펙트럼에서 가장 레드-이동의 모든 3 개의 GFP 파생 상품 여기 조사 (그림 3D) 이다. ECFP의 형광 방출 475에 두 개의 특성 맥시 마를 보여 및 505 nm, EGFP는 하나의 광범위 한 주요 방출 밴드 최고치 508 nm 540에 약간의 어깨 (λ최대) nm. GdFP 형광 나타납니다 약 565 nm (λ최대) (그림 3d 이미지)입니다. 475에 작은 어깨로 볼 수도 있는 야생-타입 ECFP의 작은 기여를 포함 하는 그것의 방출 스펙트럼 nm. 이 작은 ECFP 분수 설명33SPI 절차 동안 유도 전에 합성 됩니다.
그림 3E GdFP의 흡수 스펙트럼의 pH 의존 변화를 보여줍니다. 5 8의 pH 변화, 방출 최대 약간 빨간색 그리고 흡수 밴드의 약간의 확대 관찰. 그러나, 흡수 진폭의 감소 pH 8 사이 pH 5, GdFP 발 색 단의 바닥 상태 속성은 매우 약한 산도 의해 수정 나타내는 약 10%만 이다.
시간 단일 광자를 계산 하 여 모니터링 하는 형광 방출은 그림 4에 나와 해결. 가운데 550에 스펙트럼 채널 모니터링 감퇴 곡선 nm와 600 nm (그림 4A) 전시 600에서 약간 빠른 형광 붕괴 550에 감퇴에 비해 nm nm. 형광의 글로벌 적합 결과 부패 1.0의 시간 상수와 2 개의 괴기 하 게 구별할 수 형광 붕괴 구성 요소에 두 가지 지 수 구성 요소 결과 곡선 ns 및 3.3 ns (그림 4C 및 D).
그것이 GFP 가족의 많은 형광 단백질 변형에 대 한 일반적인 GdFP의 형광 방출 강하게 pH에 따라 다릅니다. 그림 4B pH 5와 pH 8, 스펙트럼 특성 일정 동안 명확 하 게 낮은 pH에서 형광 강렬에 있는 감소를 보여주는 사이 GdFP의 형광 방출을 비교 합니다.
GdFP의 부패 관련 스펙트럼 (DAS)39 (그림 4C 및 D) 두 가지 방출 악대에 의해 특징입니다. 느린 3.3 기여 ns 구성 요소는 더 발음 짧은 파장 범위에 약 550 nm (60%) 빠른 구성 요소 (40%)의 사소한 기여. 600 nm, 두 구성 요소는 동일한 진폭에 대 한. PH 변화에 따라 7 (그림 4C) DAS의 pH 6 (그림 4D), 스펙트럼 특성을 거의 변화와 글로벌 피팅 루틴에서 시간 상수는 또한 동일 (DAS 시간 상수의 정확도 ± 0.15 ns에 대 한). 그러나, 두 신발 부품의 절대 진폭 차이가 명확 하 게 명백한는 완전히 차지 그림 4B의 동일한 pH 변화에 따라 감소 형광 방출 진폭.
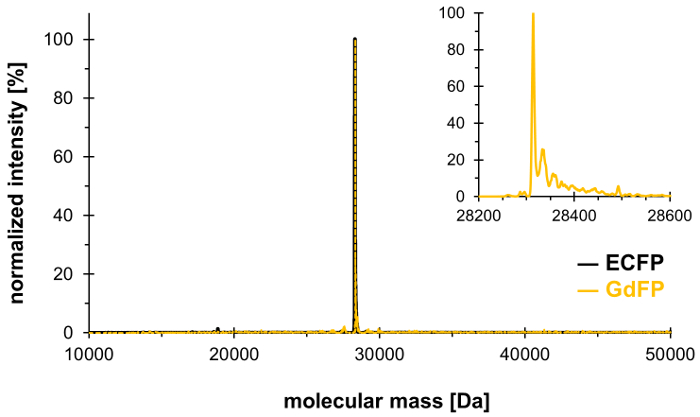
그림 1: GdFP의 대표 deconvoluted ESI MS 스펙트럼. ESI MS 스펙트럼 GdFP (골드 색상, 음각으로 표시 하는 확대 작)의 28314.1 다에서 주요 피크를 보여준다 (계산 값 28313.9 다). 야생-타입 ECFP에 대 한 스펙트럼 검정에 표시 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 2: 다양 한 프레임을 표현 하는 세균성 인구에서 Confocal 형광 현미경 이미지. 다음 파장 설정 이미지 수집을 위해 사용 되었다: ECFP (λ전 457 = nm, 탐지: 461-480 nm), EGFP (λ전 = 488 nm, 탐지: 495-515 nm), GDFP (λex 476 = nm, 탐지: 560-590 nm), EYFP (λ전 = 514 nm, 검색: 520-530 nm). 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 3: GdFP의 스펙트럼 속성. (A) CFIM 이미지 ECFP, EGFP GdFP PBS 버퍼에 박테리아의 물의 resuspension 후에 표현 하는 세균성 세포의 혼합물의. (B) (4-아미노-Trp 잔류물 66 대신)와 GdFP, (위치 66에서 Trp)와 부모의 ECFP 및 EFGP (위치 66에 티 르)와 발 색 단 구조. (C) 비교 GdFP, ECFP 및 EGFP, 정규화 된 흡수 스펙트럼의 반면 (D), ECFP의 정규화 된 형광 방출 스펙트럼 (430에서 여기 nm) EGFP GdFP (둘 다의 형광 방출 스펙트럼에 비해 450에 흥분 nm). 흡수 스펙트럼의 (E) pH 의존 (280에서 흡수 정규화 nm). 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 4: GdFP의 형광 시간 해결. (A) 형광 GdFP의 모니터링에 의해 시간-및 파장-해결 550 중심 스펙트럼 채널에서 단일 광자 nm와 600 nm (± 12.5 nm) 여기 470 nm 레이저 펄스와 후. 경 음악 응답 함수 (IRF) 사용된 설치의 시간 해상도 대 한 정보를 제공합니다. (B) 변형 GdFP pH에 의존의 방출 스펙트럼의 (여기에서 460 nm). (C, D) 부패 관련 (DAS)의 스펙트럼 GdFP pH 7 (C)와 pH 6 (D) 결정 후 시간 및 파장 해결 형광의 deconvolution 부패와 함께 두 지 수 함수의 글로벌 설정 하 여 모든 채널에서 자연 붕괴의 글로벌 피팅 연결 된 시간 상수입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 5: ECFP (검정) 및 GdFP (골드) chromophores intramolecular 충전 전송의 구조. 미식 축구의 일환으로 아미노 그룹의 좋은 전자 기증자에 의해 발 색 단 시스템의 크기에서 증가 흥분된 상태로의 공명 안정화를 달성 하기 위해 더 mesomeric 구조의 형성 수 있습니다. 연결 지점 FP 발판으로 반원을 표시 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
| 재고 솔루션 | 농도, 용 매 | 참고 | |
| 20 %D-포도 당 | 200 g/L D-포도 당 ddH2O | 0.45 μ m 기 공 크기 주사기 필터를 통해 여과 의해 소독 | |
| indole | 소 프로 파 놀에 50 m m | ||
| 4-아미노-indole | 20%의 에탄올 (ddH2O 채워 100 mL의 최종 볼륨에 20 mL 에탄올)에 50 m m | ||
| IPTG | DdH2O 1 M | ||
| L-트립토판 | 15mm ddH2O 1 M HCl을 사용 하 여에 녹 (분말은 dissoved 때까지 저 어 아래 HCl에 dropwise 추가) | ||
| lysozyme | ddH2O에서에서 50 mg/mL | ||
| DNase I | ddH2O 1 mg/mL | ||
| RNase A | ddH2O 1 mg/mL | ||
| Amp100 | 100 mg/mL 암 피 실린 ddH2O | ||
| 나트륨-dodecylsulfate (SDS) | ddH2O 200 g/L | ||
| 황산 암모늄 ((NH4)2등4) | DdH2O 1 M | 압력가 마로 소독 하 여 소독 | |
| 칼륨 디 인산 (KH2포4) | DdH2O 1 M | 압력가 마로 소독 하 여 소독 | |
| 디-칼륨 수소 인산 염 (K2HPO4) | DdH2O 1 M | 압력가 마로 소독 하 여 소독 | |
| 황산 마그네슘 (MgSO4) | DdH2O 1 M | 압력가 마로 소독 하 여 소독 | |
| D-포도 당 | DdH2O 1 M | 0.45 μ m 기 공 크기 주사기 필터를 통해 여과 의해 소독 | |
| 염화 나트륨 (NaCl) | DdH2O 5 M | 압력가 마로 소독 하 여 소독 | |
| 염화 칼슘 (CaCl2) | 1 g/L | 0.45 μ m 기 공 크기 주사기 필터를 통해 여과 의해 소독 | |
| iron(II) 염화 (FeCl2) | 1 g/L | 0.45 μ m 기 공 크기 주사기 필터를 통해 여과 의해 소독 | |
| 티 아민 | 10 g/L | 0.45 μ m 기 공 크기 주사기 필터를 통해 여과 의해 소독 | |
| 비타민 b 복합체 | 10 g/L | 0.45 μ m 기 공 크기 주사기 필터를 통해 여과 의해 소독 | |
| 추적 요소 혼합 | 황산 구리 (CuSO4), 아연 염화 물 (ZnCl2), 염화 망간 (MnCl2), 황화 몰 리브 덴 ((NH4)2무4); ddH2O 각 1 mg/L | 0.45 μ m 기 공 크기 주사기 필터를 통해 여과 의해 소독 | |
| 19 아미노산 믹스 | 1.) 0.5 g L-페닐알라닌 및 분말 녹아 때까지 저 어 아래 1 M HCl의 100 ml ddH2O dropwise 추가 0.5 g L-티로신 해산. | ||
| (L-트립토판)를 제외 하 고 나머지 L 아미노 산의 각 0.5 g으로 무게 2.). 22 mL fo 믹스 1 M KH2포4 및 48 mL 1 M K2HPO4의. 약 800 ml ddH2O를 추가 합니다. 솔루션 취소 될 때까지 저 어. | |||
| 3.) 추가 녹아 L-페닐알라닌 그리고 L-티로신 1 단계에서.) ddH2오 1 l 볼륨 조정 | |||
| 4) Sterilize 병 최고 필터 장치 진공 여과 의해 아미노산 혼합물. | |||
| 버퍼 및 미디어 | 구성/준비 | ||
| SDS 염료 버퍼 로드, 5 배 집중 | 0.25 M Tris pH 6.8, 50 %v / v 글리세롤, 0.25 %w / v bromphenol 블루, 0.5 M didhiothreitol (DTT, 또는 5% β-mercaptoethanol), 10 %w / v 나트륨-dodecylsulfate (SDS) | ||
| 바인딩 버퍼 | 50mm 나트륨 dihydrogenphosphate (NaH2포4), 500 m m NaCl, 이미 10 m m, pH 8 | ||
| 차입 버퍼 | 50mm 나트륨 dihydrogenphosphate (NaH2포4), 500 m m NaCl, 이미 250 m m, pH 8 | ||
| 투 석 버퍼 | 50mm 나트륨 dihydrogenphosphate (NaH2포4), 150 m m NaCl, 100 mL/L 글리세롤, pH 8 | ||
| MS 버퍼 | 10 mM Tris HCl, pH 8 | ||
| 19 L-아미노산 L-트립토판 (NMM19)를 제외 하 고 포함 하는 새로운 최소 매체 | 다음 최종 농도 얻기 위해 모든 재고 솔루션을 혼합: 7.5 m m (NH4)2등4, 1.7 m m NaCl, 22 mM KH2포4, 50 m K m2HPO4, 1mm MgSO4, 20 m m D-포도 당, 50 mg/L의 19 아미노산 믹스, 1 µ g/L CaCl2, 1 µ g/L FeCl2, 10 µ g/L 티 아민, 비오 틴 10 mg/L, 0.01 mg/L 성분 혼합 | ||
| LB 매체 | 구성: 10 g/L tryptone 5 g/L 효 모 추출 물, 10 g/L NaCl, pH 7.0 ddH2O | ||
| 준비: | |||
| 1.) 무게 50 g tryptone 밖으로, 25 g 효 모 추출 물, 5 g NaCl 1 L 유리 병에. | |||
| 2.) 추가 ddH2O ~ 800 mL까지와 감동에서 디졸브 구성 요소. | |||
| 3.) pH를 측정 하 고 pH 7에 dropwise 추가 1 M HCl 또는 1 M NaOH에 의해 필요한 경우 조정 합니다. 1 L. ddH2O 추가 | |||
| 압력가 마로 소독, 여 4) Sterilize 이후에 볼륨 손실에 대 한 확인 하 고 살 균 ddH2O 필요한 경우 보상을 추가. 사용 될 때까지 4 ° C에서 저장 합니다. | |||
| 파운드 한 천 배지 | 구성: 10 g/L tryptone 5 g/L 효 모 추출 물, 10 g/L NaCl, 우 무 15 g/L, pH 7.0 ddH2O | ||
| 준비: | |||
| 1.) 무게 50 g tryptone 밖으로, 25 g 효 모 추출 물, 5 g NaCl, 1 L 유리 병에 7.5 g 우 무. | |||
| 2.) 추가 ddH2O 교 아래 500 mL 및 디졸브 구성 요소까지. | |||
| 3.) pH를 측정 하 고 pH 7에 dropwise 추가 1 M HCl 또는 1 M NaOH에 의해 필요한 경우 조정 합니다. 1 L. ddH2O 추가 | |||
| 압력가 마로 소독, 여 4) Sterilize 이후에 볼륨 손실에 대 한 확인 하 고 필요한 경우 살 균 ddH2O 보상, 추가. (참고: 파운드 천 파운드 천 접시의 준비를 위해 사용 될 때까지 4 ° C에서 저장 될 수 있다. 조심 스럽게 전자 레인지를 사용 하 여 응고 한 천 녹) | |||
| 5.) 때 솔루션은 아직 따뜻한 (30-40 ° C), 암 피 실린 100 µ g/mL의 최종 농도에 추가 | |||
| 6.) 부 어 5 단계에서 액체의 약 15 mL.) 살 균 10 cm 배양으로 무 균 조건 하에서 요리. 천 경화 때 플레이트 사용까지 4 ° C에서 1 주 동안 저장할 수 있습니다. | |||
| 인산 염 버퍼 식 염 수 (PBS) | 구성: 137 m m NaCl, 2.7 m m KCl, 10mm 나2HPO4, 1.8 m m KH2포4, 1mm CaCl2, 0.5 m m MgCl2, pH 7. 압력가 마로 소독 또는 여과 하 여 소독. | ||
표 1: 주식 해결책 및 버퍼.
Discussion
매우 높은 ncAA 법인 효율성을 달성 하기 위해, auxotrophy 기반 SPI 메서드는 ncAA의 해당 자연 대응 합성 수 없습니다 metabolically 설계 호스트 세포의 사용에 의존 합니다. 대장균을 그러한 변종 쉽게 사용할 수 있습니다. 같은 단백질으로 여러 ncAAs의도 동시 설립은 multiauxotrophic 긴장을 사용 하 여 가능 합니다. 교체와 비슷한 화학 아날로그에 제한 되 고 화학 레 퍼 토리의 잔류물 관련 모드는 단점으로 볼 수 있습니다. 그럼에도 불구 하 고, 자연 세균성 번역 기기 허용 수많은 아미노산 아날로그로 단백질 이체의 많은 수를 생산 수 있습니다. 예를 들어 50 ncAAs 단백질 생체 외에서 번역, 다시40에 대 한 사용할 수 유전 코드의 모든 codons의 약 73%를 사용 하 여에 포함 될 수 있습니다. 또한, SPI는 또한 대상 단백질41의 효율적인 multisite 라벨 수 있습니다. 원칙적으로, SPI 방법론 대장균을 제한 되지 않습니다 하지만 사용할 수 있습니다 다른 호스트에서 정식 20 아미노산의 각 제공 하는 auxotrophic 긴장 및 정의 된 재배 미디어 사용할 수 있다. 예를 들어 두 개의 메티오닌 아날로그, azidohomoalanine (Aha) 및 homopropargylglycine (Hpg), 상업적으로 사용할 수 있는 사용 됩니다 단백질과 다양 한 유기 체에 있는 proteomes를 라벨에 대 한. 또한, Aha 침 생산 하 고 그 후 단백질42에 통합 될 수 있습니다. 이 번의 전미이 대학 bioorthogonal 활용형 클릭 화학 Tirrel와 동료에 의해 개발 등을 위해 특히 적당 하다: 예를 들어 누에나방 애벌레43, 초파리 에서 애기 thaliana의 조직에 공장 셀44, 애벌레 zebrafish45 로 뉴런46을 포함 한 포유류 세포, 단백질 Aha47,48이라는 수 있습니다. 마찬가지로, Trp 아날로그 Trp auxotrophic Lactococcus lactis 긴장49에서 항균 성 펩 티 드에 성공적으로 통합 했다. SPI는 Xenobiology50,51, 탐구 생활의 기본적인 화학 메이크업에 대 한 대안의 분야에도 유용 합니다. 예를 들어 대장균52 에 B. subtilis53이전 작품을 바탕으로, E. 콜라이 긴장 개발한 최근 대신 thienopyrrole를 사용 하는 선택적 압력 진화 전략 indole, 유전자 코드54thienopyrrole-알라닌, 트립토판의 프로테옴 전체 대체 결과 일반적으로, 단일 triplet (UGG)으로 인코딩된 Trp 정식 아미노산 단백질 공학 indole 화학, 수많은 화학 유사 콘텐츠를 제공의 풍부한 측면 때문에 유망한 대상을 선물 한다. 최근, SPI 기반 병합 하는 대신 SCS 플랫폼 세균 및 진 핵 호스트에서 site-specifically Trp 아날로그 통합 수 있다 소설 보고55. 이 더 스펙트럼 속성의 변경을 포함 하 여 ncAA-기반 단백질 공학, vivo에서 도구 상자를 높 혔 습니다.
Auxotrophic 식 호스트를 사용 하 여, 외 SPI 프로토콜 필요 엄격한 발효 조건, 대상 식 타이밍와 ncAA 법인 고효율 및 대상 단백질 수율에 도달 하기 위해서는 중간의 구성 56. 재배 본질적으로 주요 소금 외에 질소 (염화 소금)와 탄소 (D-포도 당), 비타민, 추적 요소에 대 한 소스를 포함 하는 화학적으로 정의 된 최소한의 미디어를 사용 하 여 수행 됩니다. 비록 엄격 하 게 필요한 추가 auxotrophies, 나머지 아미노산의 부재에 (20-n, n 아미노산 대체 하는 경우) 일반적으로57세균성 성장을 촉진에 추가 됩니다. 대상 단백질 식의 유도 하기 전에 초기 성장 단계 동안 교체 n 정식 아미노산 농도 제한에 추가 됩니다. 세포 성장 대상된 필수 아미노산 고갈, 실험적으로 고정 세600으로 표시 될 때까지 진행 한다. 그 후, 문화 매체 하 고갈된 아미노산 부족 풍부한 농도에서 미식 축구를 포함 하는 신선한 매체에 의해 대체 됩니다. 이 프로토콜에서와 같이 트립토판 아날로그의 ribosomal 설립은 침 트립토판 synthase58해당 트립토판 파생으로 변환 되는 indole 아날로그 먹이입니다. 다음, 대상 단백질 식이 유도 된다. 이 단계에서 세포는 전체 휴대폰 번호와 피트 니스 사이 균형으로 로그 성장의 끝. 존재와는 정식의 아미노 야생 형 단백질 생산으로 이어질 것, 그것은 중요 한 필수 아미노산 유도 전에 완전히 고갈 되도록. 마찬가지로, 그것은 질량 분석에 의해 일반적으로 대상 단백질에 번의 전미 대학 설립의 효율성을 검토 하는 필수입니다. 경우에 정식 아미노산, 재배 조건 필요가 조정 될, 예를 들어, 초기 성장 단계에 대 한 필수 amino acid(s)의 농도 또는 나중의 기간을 변경 하 여 실질적인 존재. 미식 축구 쪽으로 낮은 aaRS 활동, 시 생 효소의 overexpression 또는 미식 축구 쪽으로 더 활성화 되어 다른 aaRS의 공동 식 실시59될 수 있습니다.
정식 아미노산 Trp 세 놀라운 기능 부여: (i) 단백질에 그것의 자연적인 풍부는 낮다; (ii)는 생물과 화학 속성은 고유 (예., 그것은 일반적으로 단백질 및 펩 티 드의 본질적인 형광의 지배적인 근원), 및 (iii) 그것은 다양 한 생 화 학적 상호 작용에 기여 하 고 기능을 포함 하 여 Π 스태킹, H 결합 및 양이온-π 상호 작용. 이러한 모든 기능은 근본적으로 Trp → 4-아미노-Trp 대체 GdFP 넘어 의심에서 시 변경, avGFPs의 "금" 클래스의 디자인은 맞춤형 autofluorescent 단백질 엔지니어링에 대 한 놀라운 예입니다. 고유한 스펙트럼 속성, FPs mutagenesis와 번의 전미 대학 설립을 통해 특정 스펙트럼 창 쪽으로 조정 될 수 있습니다. GdFP, 경우이 간단한 화학의 교환에 의해 H → NH2 프레임에 ECFP 발 색 단 깡패에 포함 된 indole 링 수행 됩니다. 그림 5 는 발 색 단 내에서 번의 전미 대학 설립의 효과 표시합니다. 4-아미노-indole (침 4-아미노-Trp로 변환)에서 발생 하는 전자 기증 그룹의 소개의 mesomeric 구조는 안정된 상태를 설명할 수 있는 다양 한 수 있습니다. 시선, 그 확대 Stokes shift 키와 레드 이동 형광 방출 확장된 활용된 시스템의 이러한 고유 속성에서 유래한 다. 로 이전, GdFP 발 색 단 내에서 향상 된 intramolecular 충전 전송은 본질적으로 pH (그림 4B) 보고 하 고 S1 와 S0 지상 쌍 극 자 모멘트에 큰 변화에 의해 동반 흥분 상태 ECFP33상대적입니다. 대체 전자 기증 그룹으로 트립토판 아날로그 hydroxy 그룹 대체 indole 링 베어링 사용 될 수, 모델 단백질 barstar41와 비교 연구에서 보고.
GdFP의 흡수 및 형광 스펙트럼 ECFP EGFP 비해 확대는 (그림 3C 및 D). 균질 흡수와 형광 밴드의 확대는 일반적으로 발생 진동 모드는 발 색 단에 의해 하 고, 또한, 추가 진동 모드 단백질60에 발 색 단의 커플링에 의해 합니다. 지방 단백질 환경 결합 요금 지역화는 발 색 단에 의해 지원 됩니다. 단백질의 구조적 이질성 vibronic 스펙트럼의 현지 변이에 리드는 발 색 단의 vibronic 스펙트럼와 나머지 단백질의 결합을 같은 충전 delocalization 및 mesomeric 상태에 표시 된에 의해 지원 됩니다. 그림 5. 이 커플링 또한 큰 Stokes shift를 지원 하 고 반드시 형광 양자 수율을 감소 시킨다. 레드 이동 하는 다른 FPs에 비해 GdFP도 향상 된 단백질 안정성 및 집계33,,6162낮은 추세를 전시 한다. 그것은 다른 FP의 변종에서 컬러로 다릅니다 뿐만 아니라 또한 실질적으로 증가 열적 안정성 및 향상 된 협동33접는 전시. 그것의 형광 강도 적어도 90 %ECFP 형광은 약 30% 감소 하는 동안 60 ° C에 난방 시 유지. 단백질, 향기로운 아미노산은 종종 일반적으로 단백질의 3 차 구조에 안정화 효과가지고 하는 상호 작용 측면 체인의 네트워크에 기여할. avGFP 이러한 측면 체인 네트워크, 자체로 페-165, 그의-148, 잘하고 Tyr-145로 발 색 단으로 구성 된 항만. 이 쪽 체인은만 GdFP 구조33, 매우 딱딱한 하지만 중요 한 것은, 그들은 소수 성 접촉은 발 색 단을 형성. GdFP에서 확인 된 가장 눈에 띄는 새로운 기능 aminated 발 색 단 페-165 더 근입니다. 이 상호 작용은 다른 알려진된 avGFPs에서 관찰 되지 않는 기능입니다. 두 잔류물은 3.2-4.5 Å 떨어져, 향기로운 아미노산 상호 작용 존재 또한 수 있습니다. Amination 유도 공명 안정화는 발 색 단, 함께이 가장 가능성이 안정 협력 방식에서 아미노산의이 소수 성 네트워크. 더 효과적인 intramolecular 충전 전송 발 색 단의 바닥 상태에 비해 흥분된 상태에서 이러한 상호 작용에 의해 지원 될 수 있습니다 그리고 그것은 적어도 부분적으로는 108 nm Stokes shift33,62 차지 .
Fluorophore 속성의 합리적인 디자인, delocalized π-시스템의 크기에서 증가 레드 이동 여기 파장에서 발생할 전망 이다. 이 규칙의 엄지손가락 위치 중립 chromophores 66 선도에서 아미노산의 시리즈에 의해 순종: 페 (λ최대 = 355 nm) < 그의 (λma x= 386 nm) < Tyr (λ최대 = 395 nm) < Trp (λ최대 = 436 nm)63. 자연 속에서 π 유대의 발 색 단의 활용된 시스템의이 확장 다른 전략에 의해 달성 되었습니다. Discosoma 풀에서 DsRed에 대 한 그것은 따라서 573 nm64λ최대 변화는 추가 아미노산의 통합에 의해 확장 됩니다. AsFP595의 발 색 단 (λ최대 = 595 nm) Anemonia sulcata 에서 그것의 π 시스템65확대 imino 그룹에 의해 확장 되었다. GdFP 및 다른 avFPs의 발 색 단 같은 크기의 이기 때문에, 다른 원리는 확장 된 DsRed와 asFP595 chromophores 범위에서 방출 파장을 수반 해야 합니다. 108의 심오한 Stokes shift nm GdFP 발 색 단, autofluorescent 단백질의 디자인에 새로운 photophysical 원리를 밝혀의 독특한 구조에 기인한 다. (로 62에 보고 된) 예비 계산 GdFP의 흥분 상태 발 색 단의 쌍 극 자 모멘트는 ECFP의 해당 값과는 달리 지상 상태에 보다 실질적으로 더 큰으로 나타났습니다. ECFP 발 색 단에 대 한 변화는 오히려 온건한 GdFP의 쌍 극 자 순간 S1~ 15 D ~ 3 D S0 상태에서 (Debye)에서 증가, 반면 (~ 4 D에서 ~ 6 D). 따라서, GdFP의 독특한 골든 형광 증가 가능한 mesomeric 구조 ( 그림 5참조) 공명 안정화에 대 한 허용 하는 다양 한 발 색 단 내 상당한 intramolecular 충전 전송에 의해 발생 합니다. 이는 방출 발생 하는 에너지 레벨을 줄일 수 있습니다. 여기에 쌍 극 자 모멘트에서 심오한 변화 결과로 intramolecular 충전 분리 발 색 단 환경의 정전기 잠재력에 있는 변화에 대 한 주요 이유입니다. 주변 단백질 매트릭스, 차례 차례로, 발 색 단 여기 후 충전 분포에 변화를 조정합니다. 이후 구조 휴식 형광 스펙트럼의 충전 전송 특성으로 인해 레드에 교대는 흥분된 발 색 단의 에너지 레벨을 낮춘 다. 같은 이유로, 큰 Stokes shift 및 radiationless 프로세스, GdFP의 형광 양자 수율의 향상 된 속도 결과로 감소에 대 한 ECFP33에 비해.
높은 양자 수율 및 ECFP의 EGFP 작은 스톡 변화는 보통의 자유도 감소 시키는 발 색 단의 딱딱한 단백질 환경 관찰 작용 하 고, 따라서, 흥분 상태의 복사 휴식을 선호 하는 내부 변환 66. 결과적으로, 더 엄격 하 게 임베디드 chromophores 나머지 단백질 매트릭스의 감소 된 결합으로의 분자 디자인 멀리 레드 이동 GFP 파생 상품과 높은 형광 양자 수율을 생산 하 가이드 역할 수 있습니다. 따라서 더 레드 이동 autofluorescent 단백질을 생산 하기 위해 접근 엔지니어링, π-전자 시스템 및 경직 된 발 색 단 구조 약한 단백질 환경에 커플링의 확대는 매우 바람직한. 이러한 수정 chromophores GFP 기반으로 직접 또는 발 색 단 부근에서 원하는 ncAAs의 배치에 의해 도입 될 또한.
Disclosures
저자 들은 아무 경쟁 금융 관심사 선언 합니다.
Acknowledgments
이 작품은 독일 연구 재단 (우수 "통합 개념에서 촉매의 클러스터) T.F. 주의 하 고 교육과 과학 (BMBF 프로그램" HSP 2020 ", TU WIMIplus 프로젝트 SynTUBio)의 연방 내각에 의해 F. 제를 지원 했다
Materials
| Name | Company | Catalog Number | Comments |
| Chemicals | |||
| 4-aminoindole | Sigma-Aldrich | 525022 | |
| acetonitrile | VWR | HiPerSolv CHROMANORM ULTRA for LC-MS, 83642 | LC-MS grade required |
| agar-agar | Carl Roth | 5210 | |
| ammonium molybdate ((NH4)2MoO4) | Sigma-Aldrich | 277908 | |
| ammonium sulfate ((NH4)2SO4) | Sigma-Aldrich | A4418 | |
| ampicillin sodium salt | Carl Roth | K029 | |
| biotin | Sigma-Aldrich | B4501 | |
| bromophenol blue | Sigma-Aldrich | B0126 | |
| calcium chloride (CaCl2) | Sigma-Aldrich | C5670 | |
| colloidal silica | Sigma-Aldrich | Ludox HS-40, 420816 | |
| Coomassie Brillant Blue R 250 | Carl Roth | 3862 | |
| copper sulfate (CuSO4) | Carl Roth | CP86.1 | |
| D-glucose | Carl Roth | 6780 | |
| di-sodium hydrogen phosphate (Na2HPO4) | Carl Roth | X987 | |
| di-potassium hydrogen phosphate (K2HPO4) | Carl Roth | P749.1 | |
| 1,4-dithiothreitol (DTT) | Carl Roth | 6908 | |
| DNase I | Sigma-Aldrich | D5025 | |
| ethanol | Carl Roth | 9065.1 | |
| formic acid | VWR | HiPerSolv CHROMANORM for LC-MS, 84865 | LC-MS grade required |
| glycerol | Carl Roth | 3783 | |
| imidazole | Carl Roth | X998 | |
| indole | Sigma-Aldrich | I3408 | |
| iron(II) chloride (FeCl2) | Sigma-Aldrich | 380024 | |
| isopropanol | Carl Roth | AE73.1 | |
| isopropyl β-D-1-thiogalactopyranoside (IPTG) | Sigma-Aldrich | I6758 | |
| lysozyme | Sigma-Aldrich | L6876 | |
| magnesium chloride (MgCl2) | Carl Roth | KK36.1 | |
| magnesium sulfate (MgSO4) | Carl Roth | 8283.2 | |
| manganese chloride (MnCl2) | Sigma-Aldrich | 63535 | |
| β-mercaptoethanol | Carl Roth | 4227.3 | |
| potassium chloride (KCl) | Carl Roth | 6781.3 | |
| potassium dihydrogen phosphate (KH2PO4) | Sigma-Aldrich | P5655 | |
| RNase A | Carl Roth | 7156 | |
| sodium chloride (NaCl) | Carl Roth | P029 | |
| sodium dihydrogen phosphate (NaH2PO4) | Carl Roth | T879 | |
| sodium dodecyl sulphate (NaC12H25SO4) | Carl Roth | 0183 | |
| thiamine | Sigma-Aldrich | T4625 | |
| Tris(hydroxymethyl)-aminomethane (Tris) | Carl Roth | 5429 | |
| Tris hydrochloride (Tris-HCl) | Sigma-Aldrich | 857645 | |
| tryptone | Carl Roth | 8952 | |
| yeast extract | Carl Roth | 2363 | |
| zinc chloride (ZnCl2) | Sigma-Aldrich | 229997 | |
| Name | Company | Catalog Number | Comments |
| amino acids | |||
| L-alanine | Sigma-Aldrich | A7627 | |
| L-arginine | Sigma-Aldrich | A5006 | |
| L-asparagine | Sigma-Aldrich | A8381 | |
| L-aspartic acid | Sigma-Aldrich | A0884 | |
| L-cysteine | Sigma-Aldrich | C7352 | |
| L-glutamic acid | Sigma-Aldrich | G2128 | |
| L-glutamine | Sigma-Aldrich | G3126 | |
| L-glycine | Sigma-Aldrich | G7126 | |
| L-histidine | Sigma-Aldrich | H8000 | |
| L-isoleucine | Sigma-Aldrich | I2752 | |
| L-leucine | Sigma-Aldrich | L8000 | |
| L-lysine | Sigma-Aldrich | L5501 | |
| L-methionine | Sigma-Aldrich | M9625 | |
| L-proline | Sigma-Aldrich | P0380 | |
| L-phenylalanine | Sigma-Aldrich | P2126 | |
| L-serine | Sigma-Aldrich | S4500 | |
| L-threonine | Sigma-Aldrich | T8625 | |
| L-tryptophan | Sigma-Aldrich | T0254 | |
| L-tyrosine | Sigma-Aldrich | T3754 | |
| L-valine | Sigma-Aldrich | V0500 | |
| Name | Company | Catalog Number | Comments |
| Lab materials | |||
| 0.45 µm syringe filter with PVDF membrane | Carl Roth | CCY1.1 | |
| 1.5 mL microcentrifuge tubes | Eppendorf | 30120086 | |
| conical polystyrene (Falcon) tubes, 50 mL | Fisher Scientific | 14-432-22 | |
| Luer-Lock syringe 5 mL | Carl Roth | EP96.1 | |
| dialysis membrane, Molecular Weight Cut-Off (MWCO) 5,000 | Spectrum Medical Industries | Spectra/Por MWCO 5000 dialysis membrane, 133198 | |
| Immobilized Metal ion Affinity Chromatography (IMAC) column 1 mL, Ni-NTA | Macherey Nagel | Protino series, 745410.5 | |
| petri dishes (polystyrene, sterile) | Carl Roth | TA19 | |
| pQE-80L plasmid vector | Qiagen | no longer available | replaced by N-terminus pQE Vector set Cat No./ID: 32915 |
| protein extraction reagent BugBuster | EMB Millipore | 70921-4 | |
| round-bottom polystyrene tubes, 14 mL | Fisher Scientific | Corning Falcon, 14-959-1B | |
| Trp-auxotrophic E. coli strain | ATCC | ATCC 49980 | Bridges BA et al., Chem Biol Interact., 1972, 5(2):77-84; see main text for alternatives |
| Name | Company | Catalog Number | Comments |
| Mass Spectrometry equipment | |||
| mass spectrometer for LC-ESI-TOF-MS | Agilent | Agilent 6530 Accurate-Mass QTOF | coupled with Infinity LC system |
| mass spectrometry data analysis software | Agilent | MassHunter Qualitative Analysis software v. B.06.00 | |
| High-Performance Liquid Chromatography (HPLC) column for LC-ESI-TOF-MS | Sigma-Aldrich | Supelco Discovery BIO Wide Pore C5 HPLC column, 3 µm particle size, 10 cm x 2.1 mm | |
| HPLC autosampler vials 1.5 mL | Sigma-Aldrich | Supelco 854165 | with conical 0.1 mL glass inserts, screw caps and septa |
| Name | Company | Catalog Number | Comments |
| General equipment | |||
| benchtop centrifuge for 1.5 mL Eppendorf tubes | Eppendorf | 5427 R | |
| cooling centrifuge for 50 mL Falcon tubes | Eppendorf | 5810 R | |
| high pressure microfluidizer for bacterial cell disruption | Microfluidics | LM series with “Z” type chamber | |
| peristaltic pump for LC | GE Healthcare | P-1 | |
| Fast Protein Liquid Chromatography (FPLC) system | GE Healthcare | ÄKTA pure 25 L | |
| orbital shaker for bacterial cultivation | Infors HT | Minitron | |
| UV/Vis spectrophotometer | Biochrom | ULTROSPEC 2100 | |
| ultrasonic homogenizer for bacterial cell disruption | Omnilab | Bandelin SONOPULS HD 3200, 5650182 | with MS72 sonifier tip |
| Name | Company | Catalog Number | Comments |
| Fluorescence spectroscopy equipment | |||
| ps-pulsed laser 470 nm | Picoquant GmbH | PDL-470 | |
| time- and wavelength-correlated single photon counting (TWSPC) acquisition software | Picoquant GmbH | SymPhoTime 64 | |
| time- and wavelength-correlated single photon counting (TWSPC) detector | Picoquant GmbH | PML-16C | 16 spectral channels, to be selected by grating settings |
| single photon counting software | Picoquant GmbH | SPCM 9.75 | |
| global fitting software | Picoquant GmbH | SPC2Glo(R) | |
| fluorescence decay data analysis software | Picoquant GmbH | FluoFit program | |
| data analysis software | OriginLab Inc. | Origin 9.2 | |
| neutral density filter set | Schott | NG1 to NG11 | (400 - 650 nm, transmission 50 %, 20%, 10 %, 5 %) |
| 488 nm long-pass emission filter | AHF Analysentechnik | AHF-488 | |
| quartz cuvette | Thorlabs GmbH | CV10Q1400 | 1 cm pathlength |
References
- Shimomura, O., Johnson, F. H., Saiga, Y. Extraction, Purification and Properties of Aequorin, a Bioluminescent Protein from the Luminous Hydromedusan, Aequorea. J Cell Compar Physl. 59 (3), 223-239 (1962).
- Chalfie, M., Tu, Y., Euskirchen, G., Ward, W. W., Prasher, D. C. Green fluorescent protein as a marker for gene expression. Science. 263 (5148), 802-805 (1994).
- Andresen, M., et al. Structure and mechanism of the reversible photoswitch of a fluorescent protein. P Natl Acad Sci USA. 102 (37), 13070-13074 (2005).
- Andresen, M., et al. Structural basis for reversible photoswitching in Dronpa. P Natl Acad Sci USA. 104 (32), 13005-13009 (2007).
- Brakemann, T., et al. A reversibly photoswitchable GFP-like protein with fluorescence excitation decoupled from switching. Nat Biotechnol. 29 (10), 942-947 (2011).
- Kremers, G. -J., Gilbert, S. G., Cranfill, P. J., Davidson, M. W., Piston, D. W. Fluorescent proteins at a glance. J Cell Sci. 124 (Pt 2), 157-160 (2011).
- Shimomura, O. Structure of the chromophore of aequorea 0. shimomura green fluorescent protein. FEBS Lett. 104 (2), 220-222 (1979).
- Shaner, N. C., Campbell, R. E., Steinbach, P. A., Giepmans, B. N. G., Palmer, A. E., Tsien, R. Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat Biotechnol. 22 (12), 1567-1572 (2004).
- Shcherbo, D., et al. Bright far-red fluorescent protein for whole-body imaging. Nat Methods. 4 (9), 741-746 (2007).
- Shcherbakova, D. M., Subach, O. M., Verkhusha, V. V. Red fluorescent proteins: advanced imaging applications and future design. Angew Chem Int Edit. 51 (43), 10724-10738 (2012).
- Stepanenko, O. V., Verkhusha, V. V., Kuznetsova, I. M., Uversky, V. N., Turoverov, K. K. Fluorescent proteins as biomarkers and biosensors: throwing color lights on molecular and cellular processes. Curr Protein Pept Sc. 9 (4), 338-369 (2008).
- Wang, L., Xie, J., Deniz, A. A., Schultz, P. G. Unnatural amino acid mutagenesis of green fluorescent protein. J Org Chem. 68 (1), 174-176 (2003).
- Budisa, N., Steipe, B., Demange, P., Eckerskorn, C., Kellermann, J., Huber, R. High-level biosynthetic substitution of methionine in proteins by its analogs 2-aminohexanoic acid, selenomethionine, telluromethionine and ethionine in Escherichia coli. Eur J Biochem. 230 (2), 788-796 (1995).
- Sharma, N., Furter, R., Kast, P., Tirrell, D. A. Efficient introduction of aryl bromide functionality into proteins in vivo. FEBS Lett. 467 (1), 37-40 (2000).
- Liu, C. C., Schultz, P. G. Adding new chemistries to the genetic code. Annu Rev Biochem. 79, 413-444 (2010).
- Twine, S. M., Murphy, L., Phillips, R. S., Callis, P., Cash, M. T., Szabo, A. G. The Photophysical Properties of 6-Azaindole. J Phys Chem B. 107 (2), 637-645 (2003).
- Lepthien, S., Hoesl, M. G., Merkel, L., Budisa, N. Azatryptophans endow proteins with intrinsic blue fluorescence. P Natl Acad Sci USA. 105 (42), 16095-16100 (2008).
- Budisa, N., et al. Probing the role of tryptophans in Aequorea victoria green fluorescent proteins with an expanded genetic code. Biol Chem. 385 (2), 191-202 (2004).
- Ross, J. B., et al. Spectral enhancement of proteins: biological incorporation and fluorescence characterization of 5-hydroxytryptophan in bacteriophage lambda cI repressor. P Natl Acad Sci USA. 89 (24), 12023-12027 (1992).
- Soumillion, P., Jespers, L., Vervoort, J., Fastrez, J. Biosynthetic incorporation of 7-azatryptophan into the phage lambda lysozyme: Estimation of tryptophan accessibility, effect on enzymatic activity and protein stability. Protein Eng Des Sel. 8 (5), 451-456 (1995).
- Heim, R., Tsien, R. Y. Engineering green fluorescent protein for improved brightness, longer wavelengths and fluorescence resonance energy transfer. Curr Biol. 6 (2), 178-182 (1996).
- Bridges, B. A., Mottershead, R. P., Rothwell, M. A., Green, M. H. L. Repair-deficient bacterial strains suitable for mutagenicity screening: tests with the fungicide captain. Chem Biol Interact. 5 (2), 77-84 (1972).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Bacterial Transformation: The Heat Shock Method. J Vis Exp. , (2017).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Bacterial Transformation: Electroporation. J Vis Exp. , (2017).
- Grigorenko, B. L., Krylov, A. I., Nemukhin, A. V. Molecular modeling clarifies the mechanism of chromophore maturation in the green fluorescent protein. J Am Chem Soc. , (2017).
- JoVE Science Education Database. General Laboratory Techniques. Introduction to the Spectrophotometer. J Vis Exp. , (2017).
- Goedhart, J., et al. Structure-guided evolution of cyan fluorescent proteins towards a quantum yield of 93%. Nat Commun. 3, 751 (2012).
- Neidhardt, F. C., Bloch, P. L., Smith, D. F. Culture medium for enterobacteria. J Bacteriol. 119 (3), 736-747 (1974).
- Hörnsten, E. G. On culturing Escherichia coli on a mineral salts medium during anaerobic conditions. Bioprocess Eng. 12 (3), 157-162 (1995).
- Davis, B. D. The Isolation of Biochemically Deficient Mutants of Bacteria by Means of Penicillin. P Natl Acad Sci USA. 35 (1), 1-10 (1949).
- Sambrook, J., Russell, D. W. Molecular Cloning: A Laboratory Manual. , Cold Spring Harbor Laboratory Press. Cold Spring Harbor, NY, USA. (2001).
- Wang, Y. -S., et al. The de novo engineering of pyrrolysyl-tRNA synthetase for genetic incorporation of L-phenylalanine and its derivatives. Mol Biosyst. 7 (3), 714-717 (2011).
- Bae, J. H., et al. Expansion of the genetic code enables design of a novel "gold" class of green fluorescent proteins. J Mol Biol. 328 (5), 1071-1081 (2003).
- JoVE Science Education Database. Dialysis: Diffusion Based Separation. J Vis Exp. , Cambridge, MA. (2017).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Separating Protein with SDS-PAGE. J Vis Exp. , (2017).
- Petrásek, Z., et al. Excitation energy transfer from phycobiliprotein to chlorophyll d in intact cells of Acaryochloris marina studied by time- and wavelength-resolved fluorescence spectroscopy. Photoch Photobio Sci. 4 (12), 1016-1022 (2005).
- Kolber, Z. S., Barkley, M. D. Comparison of approaches to the instrumental response function in fluorescence decay measurements. Anal Biochem. 152 (1), 6-21 (1986).
- Pelet, S., Previte, M. J. R., Laiho, L. H., So, P. T. C. A fast global fitting algorithm for fluorescence lifetime imaging microscopy based on image segmentation. Biophys J. 87 (4), 2807-2817 (2004).
- Loefroth, J. E. Time-resolved emission spectra, decay-associated spectra, and species-associated spectra. J Phys Chem. 90 (6), 1160-1168 (1986).
- Hartman, M. C. T., Josephson, K., Lin, C. -W., Szostak, J. W. An expanded set of amino acid analogs for the ribosomal translation of unnatural peptides. PLoS One. 2 (10), e972 (2007).
- Budisa, N., et al. Global replacement of tryptophan with aminotryptophans generates non-invasive protein-based optical pH sensors. Angew Chem Int Edit. 41 (21), 4066-4069 (2002).
- Ma, Y., Biava, H., Contestabile, R., Budisa, N., di Salvo, M. L. Coupling bioorthogonal chemistries with artificial metabolism: intracellular biosynthesis of azidohomoalanine and its incorporation into recombinant proteins. Molecules. 19 (1), 1004-1022 (2014).
- Teramoto, H., Kojima, K. Incorporation of Methionine Analogues Into Bombyx mori Silk Fibroin for Click Modifications. Macromol Biosci. 15 (5), 719-727 (2015).
- Deal, R. B., Henikoff, J. G., Henikoff, S. Genome-wide kinetics of nucleosome turnover determined by metabolic labeling of histones. Science. 328 (5982), 1161-1164 (2010).
- Hinz, F. I., Dieterich, D. C., Tirrell, D. A., Schuman, E. M. Non-canonical amino acid labeling in vivo to visualize and affinity purify newly synthesized proteins in larval zebrafish. ACS Chem Neurosci. 3 (1), 40-49 (2012).
- Dieterich, D. C., et al. In situ visualization and dynamics of newly synthesized proteins in rat hippocampal neurons. Nat Neurosci. 13 (7), 897-905 (2010).
- Dieterich, D. C., Link, A. J., Graumann, J., Tirrell, D. A., Schuman, E. M. Selective identification of newly synthesized proteins in mammalian cells using bioorthogonal noncanonical amino acid tagging (BONCAT). P Natl Acad Sci USA. 103 (25), 9482-9487 (2006).
- Glenn, W. S., et al. Bioorthogonal Noncanonical Amino Acid Tagging (BONCAT) Enables Time-Resolved Analysis of Protein Synthesis in Native Plant Tissue. Plant Physiol. 173 (3), 1543-1553 (2017).
- Zhou, L., et al. Incorporation of tryptophan analogues into the lantibiotic nisin. Amino Acids. 48 (5), 1309-1318 (2016).
- Acevedo-Rocha, C. G., Budisa, N. Xenomicrobiology: a roadmap for genetic code engineering. Microb Biotechnol. 9 (5), 666-676 (2016).
- Agostini, F., Völler, J. -S., Koksch, B., Acevedo-Rocha, C. G., Kubyshkin, V., Budisa, N. Biocatalysis with Unnatural Amino Acids: Enzymology Meets Xenobiology. Angew Chem Int Edit. 56 (33), 9680-9703 (2017).
- Bacher, J. M., Ellington, A. D. Selection and characterization of Escherichia coli variants capable of growth on an otherwise toxic tryptophan analogue. J Bacteriol. 183 (18), 5414-5425 (2001).
- Wong, J. T. Membership mutation of the genetic code: loss of fitness by tryptophan. Pc Natl Acad Sci USA. 80 (20), 6303-6306 (1983).
- Hoesl, M. G., et al. Chemical Evolution of a Bacterial Proteome. Angew Chem Int Edit. 54 (34), 10030-10034 (2015).
- Italia, J. S., et al. An orthogonalized platform for genetic code expansion in both bacteria and eukaryotes. Nat Chem Biol. 13 (4), 446-450 (2017).
- Völler, J. -S., Thi To, T. M., Biava, H., Koksch, B., Budisa, N. Global substitution of hemeproteins with noncanonical amino acids in Escherichia coli with intact cofactor maturation machinery. Enzyme Microb Tech. 106, 55-59 (2017).
- Budisa, N., Steipe, B., Demange, P., Eckerskorn, C., Kellermann, J., Huber, R. High-level biosynthetic substitution of methionine in proteins by its analogs 2-aminohexanoic acid, selenomethionine, telluromethionine and ethionine in Escherichia coli. Eur J Biochem. 230 (2), 788-796 (1995).
- Völler, J. -S., Budisa, N. Coupling genetic code expansion and metabolic engineering for synthetic cells. Curr Opin Biotech. 48, 1-7 (2017).
- Johnson, J. A., Lu, Y. Y., Van Deventer, J. A., Tirrell, D. A. Residue-specific incorporation of non-canonical amino acids into proteins: recent developments and applications. Curr Opin Chem Biol. 14 (6), 774-780 (2010).
- Somsen, O. J., van Grondelle, R., van Amerongen, H. Spectral broadening of interacting pigments: polarized absorption by photosynthetic proteins. Biophys J. 71 (4), 1934-1951 (1996).
- Kurschus, F. C., Pal, P. P., Bäumler, P., Jenne, D. E., Wiltschi, B., Budisa, N. Gold fluorescent annexin A5 as a novel apoptosis detection tool. Cytom Part A. 75 (7), 626-633 (2009).
- Lepthien, S., Wiltschi, B., Bolic, B., Budisa, N. In vivo engineering of proteins with nitrogen-containing tryptophan analogs. Appl Microbiol Biot. 73 (4), 740-754 (2006).
- Wachter, R. M., Elsliger, M. -A., Kallio, K., Hanson, G. T., Remington, S. J. Structural basis of spectral shifts in the yellow-emission variants of green fluorescent protein. Structure. 6 (10), 1267-1277 (1998).
- Verkhusha, V. V., Lukyanov, K. A. The molecular properties and applications of Anthozoa fluorescent proteins and chromoproteins. Nat Biotechnol. 22 (3), 289-296 (2004).
- Martynov, V. I., Savitsky, A. P., Martynova, N. Y., Savitsky, P. A., Lukyanov, K. A., Lukyanov, S. A. Alternative cyclization in GFP-like proteins family. The formation and structure of the chromophore of a purple chromoprotein from Anemonia sulcata. J Biol Chem. 276 (24), 21012-21016 (2001).
- Piatkevich, K. D., Malashkevich, V. N., Morozova, K. S., Nemkovich, N. A., Almo, S. C., Verkhusha, V. V. Extended Stokes shift in fluorescent proteins: chromophore-protein interactions in a near-infrared TagRFP675 variant. Sci Rep. 3 (1), 1847 (2013).
