

Summary
Синтетическая биология позволяет инженерия протеинов с беспрецедентной свойств, с помощью co-translational вставки неканонических аминокислот. Здесь мы представили, как спектрально смещается красный вариант GFP-типа Флюорофор с Роман флуоресценции спектральными свойствами, называют «золото» флуоресцентный белок (GdFP), производится в E. coli через селективного давления включения (SPI).
Abstract
Флуоресцентные белки являются основными инструментами для наук о жизни, в частности для микроскопии флуоресцирования живых клеток. В то время как одичал тип и инженерии варианты зеленого флуоресцентного белка Aequorea victoria (avGFP), а также от других видов гомолог уже охватывают большие части оптического спектра, спектральные разрыв остается в регионе ближней ИК-области спектра, для которых на основе avGFP флуорофоров не доступны. Красный смещается флуоресцентный белок (FP) варианты бы существенно расширить инструментарий для спектральных расслоение нескольких молекулярных видов, но естественным красно смещается FPs, полученных из кораллов или актинии ниже квантовый выход флуоресценции и Ниже Фото стабильность по сравнению с avGFP варианты. Дальнейшие манипуляции и возможного расширения хромофора конъюгированных системы к far-red спектральном диапазоне также ограничивается репертуар канонические 20 аминокислот, установленном генетического кода. Чтобы преодолеть эти ограничения, синтетической биологии можно добиться дальнейшего спектральных красный сдвиг через вставку неканонических аминокислот в триады хромофора. Мы описываем применение SPI инженер avGFP варианты с Роман спектральных свойств. Выражение протеина производится в триптофан ауксотрофных E. coli деформации и путем дополнения питательных сред с подходящим индола прекурсоров. Внутри клетки эти прекурсоры преобразованы в соответствующие аналоги триптофана и включены в белки рибосомных механизмом в ответ UGG кодонов. Замена ГТО-66 в варианте расширенной «голубой» avGFP (ECFP), жертвуя электрон 4-aminotryptophan приводит к GdFP, показывая 108 Нм Стоксов сдвиг и сильно смещается красный выбросов максимум (574 Нм), будучи термодинамически более стабилен, чем его предшественник ECFP. Остаток конкретного включения неканонических аминокислоты анализируется по масс-спектрометрии. Спектральные свойства GdFP были характерны отдела Флуоресценция время решен как один из ценных приложений генетически закодированный FPs в науках о жизни.
Introduction
С момента открытия Зеленый флуоресцирующий белок в медузы Aequorea victoria (avGFP) в 1962 году1 и первого гетерологичных выражения в 1994 году2 в других эукариотических клеток стали флуоресцентных белков семейства GFP очень ценные инструменты и цели в области биологических наук. Обширные генетические и молекулярные техники включены корректировки использования вегетационных кодон, ускорение складывания, улучшения созревания, повышенной яркости, предотвращения олигомеризации и пошив спектральными и фотохимическими свойствами включая возможность обратимого фотореле3,4,5,6. GFP обязан флуоресцентным из его 4-(p- hydroxybenzylidene) имидазолидин-5-1 (HBDI) хромофора. Последний autocatalytically формируется из триады так называемой хромофора аминокислот (Ser-65/Tyr-66/Gly-67 в avGFP) после формирования дополнительного ковалентной связи в пределах пептид позвоночника под влиянием молекулярного кислорода7. Резонансно стабилизированный конъюгированных система динамически взаимодействует с молекулярной среды, позволяя для поглощения в видимом диапазоне и характерные зеленой флуоресценцией этих белков.
В рамках триады хромофора присутствие ароматических аминокислот является обязательным. Однако стандартный аминокислоты репертуар включает в себя только четыре ароматических остатков (его, Phe, ГТО и Tyr). Это ограничивает обычных мутагенеза подходов к достижению существенн больше смещается красный avGFP варианты по отношению к наиболее красный смещается природных FPs например8 DsRed из Discosoma горлица coralimorphs или mKate/mNeptune по9 Актинии Entacmaea quadricolor. Таким образом, far-red и ближней инфракрасной части оптического спектра выше 600 Нм редко покрыта варианты гена GFP. Это, конечно, жесткое ограничение для флуоресценции микроскопических подходов, которые требуют спектральных демультиплексирование нескольких видов Флюорофор, в то же время. Например, маркеры длинноволновых необходимы также для использования режима низкой абсорбцией кожной ткани между 700-1000 Нм в настройках для глубоких тканей изображений10.
Флуоресцентные белки, полученные от avGFP делятся на несколько классов, основанных на спектральные свойства и химической природы их хромофоры11. С его триады Ser-65/Tyr-66/Gly-67, одичал тип хромофора существует как уравновешенной смесь между нейтральными, фенольные формы (λМакс = 395 Нм, ε = 21000 M-1см-1) и анионными фенолятные формы (λМакс = 475 Нм, ε = 7100 М -1см-1), и спектр излучения экспонаты одного пика 508 Нм. Группа гидроксила Ser-65 имеет решающее значение, поскольку оно donates H-Бонд в глу-222 в близости хромофора (расстояние: 3.7 Å), которая способствует ионизации этой карбоксилат. Класс, я характеризуется хромофора анионные фенолятные, как и EGFP (Phe-64-лей/Ser-65-Чет; λМакс = 488 нм, ε = 35 600 М-1см-1,Эм λ = 509 Нм). В связи с заменой Ser-65-Thr(Ala,Gly), 395 Нм возбуждения пик формы нейтральных фенола подавляется и 470-475 Нм пик анионные фенолятные пять - шесть раз расширение и перенесен на 490 Нм. Класс II состоит из белков с нейтральным фенольные хромофора, как сапфир GFP. Здесь, Чет-203-Иль замещения почти полностью подавляет 475 Нм возбуждения, оставляя только вершина в 399 Нм. Так как анионные хромофора не может быть должным образом сольватированного, благоприятствует его нейтральной форме. Класс III включает в себя «желтый» флуоресцентный варианты (EYFP; SER-65-Gly/Val-68-Leu/SER-72-Ala/THR-203-Tyr; ΛМакс ε = 514 Нм, ε = 84, 600 М-1см-1,Эм λ = 527 Нм) с укладки π взаимодействие ароматических боковой цепи и фенолятные, как в результате замены Thr-203-His(Trp,Phe,Tyr), которые приводят к до 20 Нм красный смещается выбросов Максима (чет-203-Tyr). Дальнейшие замены (Gln-69-Lys) приводит к другой 1-2 нм красный сдвиг 529 Нм, известный11вариант avGFP наиболее красный смещается. Обмен фенола для индола (Tyr-66-ГТО) создает класса IV, как циан люминесцентных ECFP (Ser-65-Чет/Tyr-66-Trp; λmax1 = 434 Нм, ε = 24,800 М-1см-1; λmax2 = 452 Нм, ε = 23600 М-1см-1 ; Em1 λ = 477 Нм, λЭМ2 = 504 нм). Размещения громоздких индола вероятно включена по другим, компенсационные мутации. ECFP возбуждения и выбросов Максима осенью inbetween тех белков с нейтральной или анионные хромофоры. Класс V белки гавани имидазола вместо фенола (Tyr-66-его), например., голубой флуоресцентные белки, как EBFP. Класс VI производится биржей фенол фенил пользу форме нейтральных хромофора исключительно, что соответственно приводит к позиции пик возбуждения и выбросов наиболее сместился синий (360 Нм и 442 нанометр, соответственно).
Классический сайт Направленный мутагенез особенно подходит для производства вариантов хромофора Роман avGFP, перестановка трипептид 65-67 и взаимодействующих остатков в рамке 20 канонические аминокислот. Эти возможности можно расширить когда нестандартные варианты ароматических аминокислот вводятся во время синтеза рибосомных белка12. В принципе существует два способа достижения этого результата. Первая стратегия опирается на субстрат терпимости белка перевод механизма, особенно аминоацил тРНК-синтетаз (aaRSs) к аналогов соответствующих аминокислот. Для достижения этой цели с высокой эффективностью, ауксотрофных штаммов E. coli выражения работают которые не способны синтезировать соответствующий природных аминокислот. Это позволяет замену последнего путем добавления подходящего неканонических аминокислоты (ncAAs) или прекурсоров его питательной среды. Эта стратегия, также известный как селективного давления включения (SPI)13,14, позволяет остатков конкретных замен, которые приводят к глобальным включение ncAA. Вторая стратегия использует стоп-кодон tRNAs супрессор, которые взимаются с ncAA, инженерии ферментов Орсе. Это приводит к readthrough стоп-кадр кодонов и участкам ncAA включение. Следовательно этот метод стоп-кодон подавления (СКС) приводит к расширению генетического кода15. Помощью мутагенеза стоп-кодон помещается в целевых генов на нужный сайт. В принципе SPI может также использоваться для создания рекомбинантных пептидов и белков, учитывая уникальные установки ncAA, учитывая, что редкие канонические аминокислот например Met или ГТО выбираются для замены. С ТРП, SPI подходы показали работать с большим разнообразием аналогов, включая 4 - Ф - 5 - F - и 6-F-ГТО, 7-аза ГТО, 4-OH - и 5-OH-ГТО, а также 4 - и 5-NH2- Trp или даже β (thienopyrrolyl) аланина производные16 ,17,18,19,20. Таким образом SPI может быть весьма выгодным для замены ароматических аминокислот GFP хромофоры нестандартные варианты для изучения возможности для дальнейшего уточнения спектры и Стоксов сдвиг эти FPs. Что касается всех модификаций последовательности белка совместимость с FP складывания и хромофора созревания должны испытываться экспериментально.
В этой работе мы используем класс IV ECFP21, которая осуществляет вместо одичал тип avGFP Tyr, ГТО остатков в пределах своей хромофора триады. С помощью SPI, этот ГТО-66 (и ГТО-57, только другие ГТО остатков в ECFP) заменяется на 4-амино ТРП. Присутствие электрон donating амино-группы 4-амино-ТРП внутри хромофора способствует стабилизации резонанс перенос протона далеко смещен красный возбужденное состояние (ESPT), наделенных 108 Нм Стоксов сдвиг. Этот «золото» флуоресцентный белок (GdFP) представляет собой вариант с крупнейшим красное смещение максимум флюоресценции (574 nm) среди всех белков, avGFP производные. Мы опишем способ производства белка GdFP, SPI и предоставляют протоколы для обязательного анализа результирующее изменение белков, масс-спектроскопии. Кроме того мы покажем, как GdFP могут быть использованы и во время решена флуоресценции спектроскопии подходов.
Protocol
1. Преобразование ГТО ауксотрофных E. coli
- Химически преобразования или electrocompetent клеток (50 мкл) штамма ГТО ауксотрофных E. coli , например. МЕСТА 49980 (WP2, мутант, производный от штамм E. coli B/R22), с 1 мкл 1 нг/мкл водный раствор плазмида His6-ECFP pQE - 80 Л, с использованием теплового шока или электропорация, соответственно. Пожалуйста, обратитесь к JoVE науки образования базы данных23,24 для деталей.
Примечание: Выражение вектор N-неизлечимо 6 x его меткой ECFP21 обусловлен бактериальной промоутер T5 с оператором лак кодирует pQE - 80L His6-ECFP. Она далее несет маркер выделения AmpR и colE1 происхождения репликации (последовательность позвоночника вектор pQE - 80 L можно найти на: https://www.qiagen.com/mx/resources/resourcedetail?id=c3b71572-4d82-4671-a79b-96357fe926d1&lang=en & с автозаполнением = true). Теоретические молекулярная масса белка одичал тип His6-ECFG (после созревания хромофора25) — 28303.92 да. Переведенные последовательности белка является следующим (его тег подчеркнул, вектор производные последовательности выделены жирным шрифтом): MRGSHHHHHHООMVSKGEELFTGVVPILVELDGDVNGHKFSVSGEGEGDATYGKLTLKFICTTGKLPVPWPTLVTTLTWGVQCFSRYPDHMK
QHDFFKSAMPEGYVQERTIFFKDDGNYKTRAEVKFEGDTLVNRIELKGIDFKEDGNILGHKLEYNYISHNVYITADKQKNGIKANFKIRHNIEDGS
VQLADHYQQNTPIGDGPVLLPDNHYLSTQSALSKDPNEKRDHMVLLEFVTAAGITLGMDELYK. - Пластина трансформированных клеток на плиты LB-агара (Таблица 1) дополнена глюкозы 10 г/Л, 100 мкг/мл Ампициллин и инкубировать пластины при 37 ° C на ночь.
2. рекомбинантных белков
-
На ночь культуры е. coli ATCC 49980 pQE - 80 L His6-ECFP
- Подготовить 5 мл среды фунтов (Таблица 1; дополнена глюкозы 10 г/Л, 100 мкг/мл ампициллин) в стерильных 14 мл полистирола культура трубки для аэробных роста и прививать с поодиночке колонии от агар пластины с помощью стерильной пипеткой отзыв или инокуляции цикла.
Примечание: Рекомендуется использовать колоний из свеже трансформированных клеток. Пластины с бактериальных колоний (от шага 1.2.) могут храниться при температуре 4 ° C на несколько дней. - На ночь инкубации клеток при 37 ° C в орбитальный шейкер на 200-250 об/мин.
- Подготовить 5 мл среды фунтов (Таблица 1; дополнена глюкозы 10 г/Л, 100 мкг/мл ампициллин) в стерильных 14 мл полистирола культура трубки для аэробных роста и прививать с поодиночке колонии от агар пластины с помощью стерильной пипеткой отзыв или инокуляции цикла.
-
Выражение одичал тип ECFP
- Прививать 10 мл свежего LB среднего (Таблица 1; дополнена глюкозы 10 г/Л, 100 мкг/мл ампициллин) с 100 мкл ночь культуры в 100 мл колбу Эрленмейера. Инкубируйте настой при 37 ° C в орбитальный шейкер на 200 об/мин.
Примечание: При необходимости, этот шаг может быть проведена в 10 мл NMM19 среднего (Таблица 1) дополняется 100 мкг/Л Ампициллин и 0,5 мм L-триптофан (в качестве альтернативы может использоваться индола). - Измеряют оптическую плотность на 600 Нм (600OD) каждые 20 минут преференциально измерить ячейку плотность путем определения вымирания на 600 Нм (600ОД) в спектрофотометр с помощью кювет с длиной пути от 1 см. всегда выполняют ссылку измерение с помощью соответствующей питательной среды. Развести образцы и смеси образцы хорошо для получения значения измерения 0,1-0,8, а затем рассчитать ОД600 с помощью коэффициента разрежения. Для подробной информации пожалуйста, обратитесь к предыдущей публикации 26.
- По достижении ОД600 значение 0,5-0,8 (примерно 2-3 ч после прививки), принимать образца «перед прививка» для SDS-PAGE (электрофорез геля полиакриламида додецилового сульфата натрия, шаг 4).
- Побудить целевых белков, регулируя культур в жидкой среде до 0,5 мм ИПТГ (изопропиловый β-D-1-тиогалактопиранозид, от 1 М раствор) и Инкубируйте на 30 ° C в орбитальный шейкер на 200 об/мин для 4-8 ч.
Примечание: Cyan флуоресцентные белки обычно выражена при температурах ниже 37 ° C27. - Взять образец «после выражения» для SDS-PAGE (шаг 4)..
- Урожай бактериальные клетки центрифугированием 10 мин на 5000 x g и 4 ° C.
- Отказаться от супернатанта, переливание и заморозить гранулы клеток при-20 ° C или ° C-80 до целевой очищение протеина.
- Прививать 10 мл свежего LB среднего (Таблица 1; дополнена глюкозы 10 г/Л, 100 мкг/мл ампициллин) с 100 мкл ночь культуры в 100 мл колбу Эрленмейера. Инкубируйте настой при 37 ° C в орбитальный шейкер на 200 об/мин.
-
SPI для производства GdFP
- Прививок 10 мл NMM19 среды (Таблица 1) дополняется 100 мкг/мл ампициллин, 15 мкм триптофана и 10 мкл в 100 мл колбу Эрленмейера ночь культуры и инкубировать культуры колбу на ночь в 30 ° C в орбитальный шейкер на 200 об/мин.
Примечание: Доступен целый ряд химически определенные СМИ для выращивания E. coli и SPI. В дополнение к НММ, используемый здесь MOPS средних28, глюкоза минеральных солей средних29, минимальный средний Дэвис30, минимальный средний M931, или может использоваться GMML32 . - Следующий день, измерьте ОД600 каждые 30 мин до тех пор, пока значение изменяется только менее чем 0,05 более 30 мин. Плато значение должно быть около 1.
Примечание: Отклонения от ± 0,3 единицы являются приемлемыми. В зависимости от бактериального штамма и используемой среды концентрация первоначальный триптофан (шаг. 2.3.1) может потребоваться корректировка. - Взять образец «перед прививка» для SDS-PAGE (шаг 4)..
- Урожай бактериальные клетки центрифугированием 10 мин на 5000 x g и 4 ° C. Отказаться от супернатанта, переливание.
- Ресуспензируйте клетки в 10 мл NMM19 среды с 100 мкг/мл ампициллин в 100 мл колбу Эрленмейера и добавить 4-амино индола в конечной концентрации 1 мм с использованием Стоковый раствор 50 мм. Продолжать инкубацию 30 мин при 30 ° C в орбитальный шейкер на 200 об/мин.
Примечание: Этот шаг рекомендуется из-за низкой химической стабильности Ампициллин и обеспечивает сотовой поглощение 4-амино индол. - Побудить целевых белков путем добавления IPTG конечная концентрация 0,5 мм, с использованием 1 М складе и Проинкубируйте образцы на ночь на 30 ° C в орбитальный шейкер на 200 об/мин.
Примечание: Cyan флуоресцентные белки обычно выражена при температурах ниже 37 ° C27. - Следующий день, измерьте ОД600.
- Взять образец «после выражения» для SDS-PAGE (шаг 4)..
- Урожай бактериальные клетки центрифугированием 10 мин на 5000 x g и 4 ° C и удалить супернатант на переливание.
- В случае, если такое судно не использовался для центрифугирования, передача Пелле клеток в 50 мл Конические трубки из полистирола с помощью шпателя. Заморозить Пелле клеток при-20 ° C или ° C-80 до целевой очищение протеина.
- Прививок 10 мл NMM19 среды (Таблица 1) дополняется 100 мкг/мл ампициллин, 15 мкм триптофана и 10 мкл в 100 мл колбу Эрленмейера ночь культуры и инкубировать культуры колбу на ночь в 30 ° C в орбитальный шейкер на 200 об/мин.
3. Цель очищение протеина через иммобилизованных ион металла аффинной хроматографии (ИАЦ)
-
Лизиса бактериальных клеток
- Оттепель Пелле клеток на льду на 10-20 мин.
- Ресуспензируйте Пелле клеток в 50 мл Конические трубки из полистирола с использованием 5 мл ледяной привязки буфера (Таблица 1) на льду.
- Добавьте 20 мкл лизоцима 50 мг/мл, 20 мкл 1 мг/мл DNase я, 20 мкл 1 мг/мл РНКазы A. закрыть трубку, осторожно перемешать, инвертирование 5 раз и держать его на льду за 30 мин.
Примечание: Разрушения частичной клеток происходит как катализируемой лизоцима. - Лизировать клетки путем озвучивания, с помощью УЗИ гомогенизатор отпечатка с помощью трех циклов 3 мин в 15 мл полистирола трубки, охлаждаемый слякоть льда с 2 s импульса, 4 s паузы и 45% амплитуды.
Примечание: в качестве альтернативы, высокого давления гомогенизации может использоваться, например., 20 циклов на 14000 psi. При необходимости разбавьте с помощью привязки буфера для достижения минимальной инструмента объем. Кроме того реагенты экстракции белка может использоваться для разрушения клеток. Просмотреть материалы таблицы для примеров. - Центрифугуйте образцы на 30 мин в 15 000-18 000 x g, 4 ° C.
- Перенесите супернатант в свежий трубки и запишите объем жидкости.
- Фильтр решение через фильтр шприц 0,45 мкм, с помощью шприца блокировки пластиковых Luer 5 мл и поливинилиденфторид (ПВДФ) шприц фильтра.
- Взять образец «лизатных» для SDS-PAGE (шаг 4)..
- Ресуспензируйте клеток Пелле мусора в ddH2O (равный объем как бывший lysate).
- Взять образец «лепешки» для SDS-PAGE (шаг 4)..
-
IMAC очистки
- Используйте 1 мл расфасовываться или самостоятельно Упакованные IMAC ПСОК (быстрый протеин жидкостной хроматографии) столбца согласно инструкциям производителя. Использование привязки буфера (Таблица 1) для столбца уравновешивания, а также для мытья шаг, который следует после lysate клетки был применен к столбцу.
- Собрать и объединить элюата фракций с GdFP, который может быть идентифицирован видимый свет золотой цвет.
Примечание: При необходимости целевого белка может быть этого eluted используя имидазол линейный градиент (0-250 мм) с помощью автоматизированной системы ПСОК. - Определить концентрацию протеина, с использованием значения литературы для Коэффициент вымирания на 466 Нм (ɛ466 Нм = 23 700 M-1 см-1)33 с Элюирующий буфер как ссылка. Подробную информацию о процедуре пожалуйста, обратитесь к предыдущей публикации26.
- Взять образец «элюата» для SDS-PAGE и использовать 1-10 мкг белка на Лейн в случае Окрашивание Кумасси.
Примечание: SDS образца суммы могут варьироваться в зависимости от окрашивания методом и чувствительность окрашивания. - Dialyze Алиготе элюата фракций против диализа буфер или МС буфер с помощью мембраны с молекулярной массой отсечки (MWCO) 5000-10 000. Подготовьте диализа мембрана согласно инструкциям производителя. Dialyze образец 1 мл три раза против 100 мл буфера для минимум 2 ч. Подробную информацию по этой процедуре пожалуйста, обратитесь к предыдущей публикации34.
- Для хранения заморозить образец протеина диализа буфера-80 ° c.
Примечание: Аликвоты должна быть стабильной по крайней мере 6 месяцев.
4. SDS-PAGE пробоподготовки E. coli клеточных экстракт
- Передача суспензию клеток эквивалент 1 мл ОД600 = 1 подвеска (например. 500 мкл ОД600= 2) в трубку microcentrifuge 1,5 мл.
- Урожай клетки центрифугированием за 10 мин при комнатной температуре 5000 x г. Отказаться от супернатанта, закупорить.
- Добавить ddH2O 80 мкл и 20 мкл 5 x SDS загрузки краситель буфера (Таблица 1) клетки гранул и микс, закупорить.
- Денатурируйте клетки путем нагрева до 95 ° C за 5 мин в блоке водой ванну или тепла. Впоследствии охлаждения образцов до комнатной температуры.
- Для окрашенных Кумасси SDS-PAGE согласно предыдущей публикации35используйте 10 мкл.
Примечание: SDS образца суммы могут варьироваться в зависимости от окрашивания методом и чувствительность окрашивания.
5. интактных белков массового анализа высокой производительности жидкостной хроматографии (ВЭЖХ) в сочетании с электроспрей ионизации время полета масс-спектрометрия (LC-ЭСИ TOF-МС)
Примечание: ВЭЖХ градиент, настройки и буферы могут варьироваться в зависимости от разделения столбцов и инструментом. Просмотреть материалы таблицы для подражания оборудования.
- Определить концентрацию белка из образца dialyzed против МС буфер, как описано выше (шаг 3.2.3.) с использованием MS буфера (см. таблицу материалов) как ссылка.
- Разбавить образец протеина до 0,1 мг/мл, используя MS буфер для окончательный объем 80 мкл, смешивать, тщательно закупорить, передача решения в MS Автоматический пробоотборник флакон со стеклянной вставкой и закройте его крышкой. Удалите пузырьки воздуха, стряхивая флакона.
- Заполните второй автоматический пробоотборник флакон без стеклянной вставкой (пустой буфер) 1 мл MS буфера.
- Разрешить инструмент для разминки. Калибровки инструмента. Убедитесь, что достаточное количество растворителей жидких хроматографии класса доступны (> 100 мл).
- Программа линейной 20 мин ВЭЖХ градиент от 5% до 80% буфер (0,1% муравьиной кислоты в ddH2O), в сочетании с буфером B (0,1% муравьиной кислоты в ацетонитриле).
- Начало ВЭЖХ в поток 0,3 мл/мин и подождать до тех пор, пока давление столбца является стабильной.
- Установите автоматический пробоотборник объем впрыска 5 мкл для метода LC-ЭСИ-TOF-MS, создание рабочего списка для запуска пустым последовал запуск образца и присвойте соответствующие Автоматический пробоотборник флакон позиции. Запуск рабочего списка.
- После завершения рабочего списка откройте созданный образец файла данных. Выберите диапазон в общей Ион текущей (TIC) участок для деконволюция и deconvolute спектр MS, с использованием алгоритма деконволюция максимальной энтропии.
Примечание: В зависимости от экспериментальных условий, дополнительные виды могут произойти от не созрел FP или буфер Ион аддукты.
6. флуоресценции жизни измерений и распада связанных спектры (ДАС) GdFP
Примечание: Для инструментирования отдела Флуоресценция время решена, обратитесь к Таблице материалов для подражания оборудования. Поглощения, а также возбуждения флуоресценции и выбросов спектры флуоресцентных белков также могут быть записаны с помощью лабораторных флуоресценции и УФ-Вид спектрофотометры.
-
Длина волны решена флуоресценции жизни измерение GdFP
- Готовить 2 мл раствора 1 мкм GdFP путем разбавления в буфер (Таблица 1) PBS на рН 7. Заполните решение в кювет кварц 1 см.
- Установите л.с импульсного 470 Нм лазер для возбуждения образца и 488 нм фильтра выбросов Лонг перевал и отрегулировать 600 Л/мм, решеткой из одиночных фотонов коррелированных по времени и волны, подсчета36 (TWCSPC) детектор для приобретения режима волны 500-700 Нм.
- Приобрести флуоресценции выбросов в размере всего около 200 x 103 фотонов/с до тех пор, пока около 10 пунктам3 накапливаются в приобретение максимум флюоресценции распада кривых с одиночных фотонов, подсчет программного обеспечения.
-
Измерение инструментальные реакции функция37 (МАФ)
- В PBS буфера при pH 7 замените образец кювета с 1 см кварцевые кюветы, заполнены с 1 г/Л коллоидный кремнезем (~ 220 м2/г).
Примечание: Суспензии кремнезема готовится с использованием водной суспензией 400 г/Л. - Удаление 488 нм фильтра выбросов Лонг перевал и вставка серые фильтры для регулировки скорости счета на TWCSPC детектор для ниже 100 x 103 счетчики/s.
- Настройка решетки для приобретения 470 Нм фотонов в канале 8 16-канальный детектор TWCSPC.
- Приобрести МАФ, до тех пор, пока около 10 x 103 счетчики накапливаются в максимальное излучение.
- Преобразовать флуоресценции распада кривых и МАФ ASCII файлы данных с глобальной установки программы38 .
- Проведение глобального подходят согласно модели суммы трех экспоненциальных компонентов с продолжительностей жизни как связанные параметры.
- Участок распада связанных спектры (ДАС) как распределение амплитуды отдельных распада компонентов в зависимости от длины волны с программное обеспечение для анализа данных.
- В PBS буфера при pH 7 замените образец кювета с 1 см кварцевые кюветы, заполнены с 1 г/Л коллоидный кремнезем (~ 220 м2/г).
Representative Results
Используя технику селективного давления включения, ГТО-66 в триаде хромофора ECFP (и ГТО-57, только другие ГТО остатков в ECFP) могут быть заменены 4-амино ГТО, создавая тем самым GdFP красно перенесло с собственный спектральных свойств. Масс-спектрометрия должен использоваться для демонстрации желаемого стехиометрическим интеграции неканонических аминокислоты белка, с результаты, показанные на рисунке 1. После этого, мы предоставляем данные из микроскопии, UV-Vis абсорбционной спектроскопии, а также устойчивого состояния и время и волны решены отдела Флуоресценция характеризовать свойства GdFP Флюорофор с акцентом на зависимость рН спектры.
Чтобы подтвердить обмен двух ГТО остатков в ECFP 4-амино-ТРП, масс-спектрометрических анализ проводится. Рисунок 1 показывает представитель deconvoluted еси-МС спектр GdFP. В то время как одичал тип ECFP имеет массу вычисляемый белка 28,283.9 Да после созревания хромофора, соответствующей массы GdFP – 28,313.9 Да. Deconvoluted еси-МС спектр GdFP экспонатов основной массы пик 28,314.1 ± 0,1 Да, который отличается от теоретического значения, менее 10 ppm. Находясь в пределах диапазона типичный точности для этого типа анализа25, это подтверждает включение ncAA через SPI (экспериментальное значение одичал тип ECFP: 28,283.7 да).
Рисунок 2 показывает конфокальный флуоресцентной микроскопии изображений изображения (CFIM) бактериальной клетки, выражая ECFP, EGFP, EYFP и GdFP после взмучивания бактерий в PBS буфера. Все изображения были приобретены на микроскопе оснащены УФ цель и лазерного возбуждения в о той же энергии для каждого образца.
На рисунке 3A показывает наложение изображений CFIM бактерий E. coli , выражая различные FPs, включая GdFP, всегда наблюдает с очень похожие энергии возбуждения (волн как показано на рис. 2). Рисунок 3B показывает структуру хромофора FP вариантов показано. Что касается яркости GdFP, по сравнению с ECFP (флуоресценции квантовый выход φ = 0,4), EGFP (φ = 0,6) и EYFP (φ = 0,6) важно отметить, что для GdFP, приобретение более широкий спектр света флуоресцирования (30 Нм) был использован в отличие от 20 Нм, используется для всех других spe Сиес, для того чтобы отрегулировать интенсивность изображения или подобный. С немного меньший коэффициент вымирания и снижение квантовый выход вследствие изучены фотофизические уникальных свойств яркость GdFP ниже по сравнению с другие FPs показано.
Спектр поглощения ECFP (рис. 3 c) имеет две характерные Максима на 434 Нм и 452 Нм. В отличие от GdFP характеризуется одной широкой красно смещается поглощения группы с максимум на 466 Нм. Поглощение EGFP далее красно перенесен на 488 нм. Однако, из-за гораздо большей Стоксов сдвиг GdFP (108 Нм) по сравнению с ECFP (41 Нм) и EGFP (20 Нм), спектр излучения GdFP является наиболее красно смещается из всех трех GFP производных расследование здесь (рис. 3D). В то время как флуоресценции выбросов ECFP показывает две характерные Максима на 475 Нм и 505 нм, EGFP имеет один широкий основных выбросов группа достигнув 508 Нм (λМакс) с небольшим плеча на 540 Нм. Флуоресценции GdFP появляется около 565 Нм (λМакс.) (Рис. 3D). Его спектр излучения содержит небольшой вклад одичал тип ECFP, которая также видна как небольшое плечо на 475 Нм. Эта незначительная ECFP синтезируется перед индукции во время процедуры SPI, как описано33.
3E рисунок показывает рН зависимые изменения в спектре поглощения GdFP. Для изменения рН от 8 до 5 максимальный выброс слегка сдвигает красный и незначительное расширение полосу поглощения наблюдается. Однако уменьшение амплитуды поглощения составляет только около 10% между pH 8 и рН 5, указывая, что основного состояния свойства GdFP хромофора очень слабо изменения рН.
Время решить что флуоресценции выбросов контролируются одиночных фотонов подсчета показан на рисунке 4. Распад кривых, мониторинг в спектральных каналов, центрированного 550 Нм и 600 Нм (рис. 4A) экспонат немного быстрее распада флуоресценции в 600 Нм, по сравнению с распада 550 Нм. Результаты глобального fit флуоресценции распад кривых с двух экспоненциальных компонентов результаты в двух спектрально различимых флуоресценции распада компонентов с константы времени 1.0 ns и 3.3 ns (рис. 4 c и D).
Флуоресценции выбросов GdFP сильно зависит от pH, как это характерно для многих вариантов флуоресцентного белка GFP семьи. Рисунок 4B сравнивает флуоресценции выбросов GdFP между pH 5 и рН 8, который ясно показывает снижение интенсивности флуоресценции при более низких значениях рН, в то время как спектральные характеристики останется неизменной.
Распада связанных спектры (DAS)39 GdFP (рис. 4 c и D) характеризуются наличием двух различных выбросов полос. Вклад медленно 3.3 ns компонент является более выраженным в диапазоне коротких волн около 550 Нм (60%), с незначительными вклад компонента быстрее (40%). На 600 Нм, обе компоненты имеют о же амплитуды. После перехода от рН 7 (рис. 4 c) до pH 6 (рис. 4 d), спектральные характеристики DAS вряд ли изменится и константы времени от рутины глобальной установки также то же самое (точность константы времени DAS идет о ± 0,15 ns). Однако, которая очевидна разница в абсолютных амплитуд двух компонентов Дас, полностью приходится выбросов амплитуды сокращения флуоресценции после же сдвиг pH в рисунке 4В.
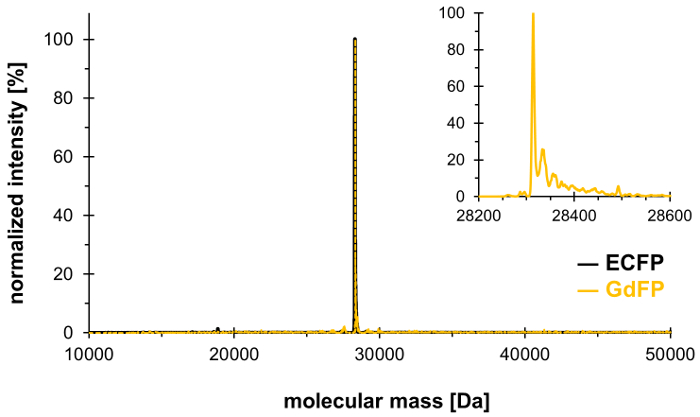
Рисунок 1: представитель deconvoluted еси-МС спектр GdFP. ЭСИ-МС спектр GdFP (Золотой цвет, увеличенное сюжет, показанный как врезные) показывает основной пик на 28314.1 Да (вычисляется значение 28313.9 Да). Спектра для одичал тип ECFP показан в черном. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 2: Confocal флуоресценции микроскопии изображений от бактериальных популяций, выражая различные FPs. Следующие параметры волны были использованы для захвата изображений: ECFP (λex = 457 Нм, обнаружение: 461-480 Нм), EGFP (λex = 488 нм, обнаружение: 495-515 нм), GDFP (λex = 476 Нм, обнаружение: 560-590 нм), EYFP (λex = 514 Нм, обнаружение: 520-530 Нм). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 3: спектральных свойств GdFP. (A) CFIM изображения смеси бактериальной клетки, выражая EGFP, ECFP и GdFP после взмучивания бактерий в PBS буфера. (B) хромофора структуры GdFP (с 4-амино Trp вместо остатков 66), родительских ECFP (с ГТО в позиции 66) и EFGP (с Tyr в позиции 66). (C) сравнение нормализованных спектров поглощения GdFP, ECFP и EGFP, тогда как в (D), нормализованных флуоресценции эмиссионный спектр ECFP (возбуждения в 430 Нм) по сравнению с выбросов спектры флуоресценции EGFP и GdFP (оба возбужденных в 450 Нм). (E) рН зависимость спектров поглощения (нормированный поглощения на 280 Нм). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 4: время решена флуоресценции GdFP. (A) флуоресценции распад GdFP контроль по времени - и волны решена одиночных фотонов, подсчет в спектральных каналов, центрированного 550 Нм и 600 Нм (± 12,5 Нм) после возбуждения с 470 Нм лазерных импульсов. Функцию инструментальные реакции (МАФ) содержит сведения о времени резолюции используется программа установки. (B) вариации выбросов спектра GdFP зависит от pH (возбуждения на 460 Нм). (C, D) Распада связанных спектры (ДАС) GdFP на pH 7 (C) и рН 6 (D) определяется после деконволюции решена раз и волны флуоресценции разлагается и глобальные установки разлагается во всех каналах глобальной набором двух экспоненциальных функций с связанные время константы. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 5: структуры внутримолекулярной заряда передачи ECFP (черный) и GdFP (золото) хромофоры. Увеличение размера хромофора системы хорошо электрона донором аминогруппы частью ncAA позволяет формирование более мезомерных структур для достижения стабилизации резонанс возбужденного состояния. Точки подключения к эшафоту FP отображаются в виде полукругов. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
| Стоковый раствор | концентрация, растворитель | Примечание | |
| 20% D-глюкозы | 200 г/Л Д-глюкозы в ddH2O | стерилизуют фильтрацией через фильтр шприц размер поры 0,45 мкм | |
| индола | 50 мм в изопропиловый спирт | ||
| 4-амино индол | 50 мм в 20% этанола (20 мл этанола в окончательный объем 100 мл, наполненный ddH2O) | ||
| IPTG | 1 M в ddH2O | ||
| L-триптофан | 15 мм растворяют в ddH2O с 1 M HCl (добавить HCl каплям при перемешивании до тех пор, пока порошок является dissoved) | ||
| лизоцим | 50 мг/мл в ddH2O | ||
| DNase I | 1 мг/мл в ddH2O | ||
| РНКазы A | 1 мг/мл в ddH2O | ||
| Amp100 | Ампициллин 100 мг/мл в ddH2O | ||
| Лаурилсульфат натрия (SDS) | 200 г/Л в ddH2O | ||
| Сульфат аммония ((NH4)2т-4) | 1 M в ddH2O | Стерилизация автоклавированием | |
| калия дигидрогенфосфат (KH2PO4) | 1 M в ddH2O | Стерилизация автоклавированием | |
| ди калия гидрогенфосфат (K2HPO4) | 1 M в ddH2O | Стерилизация автоклавированием | |
| Сульфат магния (4MgSO) | 1 M в ddH2O | Стерилизация автоклавированием | |
| D-глюкоза | 1 M в ddH2O | стерилизуют фильтрацией через фильтр шприц размер поры 0,45 мкм | |
| хлорид натрия (NaCl) | 5 М в ddH2O | Стерилизация автоклавированием | |
| Хлорид кальция (2CaCl) | 1 г/Л | стерилизуют фильтрацией через фильтр шприц размер поры 0,45 мкм | |
| железа хлорид (2FeCl) | 1 г/Л | стерилизуют фильтрацией через фильтр шприц размер поры 0,45 мкм | |
| тиамин | 10 г/Л | стерилизуют фильтрацией через фильтр шприц размер поры 0,45 мкм | |
| биотин | 10 г/Л | стерилизуют фильтрацией через фильтр шприц размер поры 0,45 мкм | |
| Прослеживающие элементы микс | Медный купорос CuSO4, цинка хлорида (ZnCl2), марганца хлорид (НКД2), Молибдат аммония ((NH4)2MoO4); каждый 1 мг/Л в ddH2O | стерилизуют фильтрацией через фильтр шприц размер поры 0,45 мкм | |
| 19 аминокислот микс | 1.) растворяют в 0,5 g L-фенилаланина и 0,5 g L-тирозин в 100 мл ddH2O с добавлением прикапывают HCl 1 M при помешивании до полного растворения порошка. | ||
| 2.) весят, 0,5 г каждого из оставшихся L-аминокислоты (за исключением L-триптофан). Смешать с 22 мл fo 1м х2PO4 и 48 мл 1 M K2HPO4. Добавьте ddH2O около 800 мл. Перемешайте до тех пор, пока решение становится ясно. | |||
| 3.) добавить растворенные L-фенилаланина и L-тирозин из шага 1.) и отрегулируйте громкость до 1 Л с ddH2O. | |||
| 4.) Стерилизируй-отпусти смеси аминокислот, вакуумной фильтрации с единицей фильтрации верхнего бутылку. | |||
| Буферы и средства массовой информации | Композиция/подготовка | ||
| SDS загрузки буфера краситель, 5 x сосредоточены | 0,25 М трис pH 6.8, 50% v/v глицерина, 0,25% w/v bromphenol синий, 0,5 М didhiothreitol (DTT; или же 5% β-меркаптоэтанол), 10% w/v натрия натрия (SDS) | ||
| Привязка буфер | dihydrogenphosphate натрия 50 мм (NaH2PO4), 500 мм NaCl, 10 мм имидазола, рН 8 | ||
| Элюирующий буфер | dihydrogenphosphate натрия 50 мм (NaH2PO4), 500 мм NaCl, 250 мм имидазола, рН 8 | ||
| диализ буфер | dihydrogenphosphate натрия 50 мм (NaH2PO4), 150 мм NaCl, глицерин 100 мл/Л, pH 8 | ||
| МС буфер | 10 мм трис-HCl, рН 8 | ||
| новый минимальный носитель, содержащий 19 L-аминокислоты Кроме L-триптофан (NMM19) | Смешайте все биржевые решения для получения следующих окончательный концентрации: 7,5 мм (NH4)2т4, 1,7 мм NaCl, 22 мм х2PO4, 50 мм K2HPO4, 1 мм MgSO4, 20 мм D-глюкоза, 50 мг/Л 19 аминокислот смесь, 1 мкг/Л CaCl2, 1 мкг/Л FeCl2, 10 мкг/Л тиамина, биотин 10 мг/Л, смесь микроэлементов 0,01 мг/Л | ||
| Средний LB | Состав: 10 г/Л Триптон, экстракт дрожжей 5 г/Л, 10 г/Л NaCl, pH 7.0 ddH2O | ||
| Подготовка: | |||
| 1.) весят вне Триптон 50 g, 25 г дрожжей экстракт, 5 г NaCl в стеклянной бутылки 1 Л. | |||
| 2.) добавить ddH2O до ~ 800 мл и растворения компонентов при помешивании. | |||
| 3.) измерения рН и приспособиться к рН 7 каплям добавлением 1 М HCl или 1 M NaOH, в случае необходимости. Добавить ddH2O до 1 л | |||
| 4.) Стерилизируй-отпусти, автоклавирования, потом проверить объем потерь и добавить стерильные ddH2O компенсировать в случае необходимости. Хранить при 4 ° C до использования. | |||
| Плиты агара LB | Состав: 10 г/Л Триптон, экстракт дрожжей 5 г/Л, 10 г/Л NaCl, агар-агар 15 г/Л, pH 7.0 в ddH2O | ||
| Подготовка: | |||
| 1.) весят вне Триптон 50 g, 25 г дрожжей экстракт, 5 г NaCl, 7.5 g агар-агар в стеклянной бутылки 1 Л. | |||
| 2.) добавить ddH2O до 500 мл и растворения компонентов при помешивании. | |||
| 3.) измерения рН и приспособиться к рН 7 каплям добавлением 1 М HCl или 1 M NaOH, в случае необходимости. Добавить ddH2O до 1 л | |||
| 4.) Стерилизируй-отпусти, автоклавирования, потом проверьте потеря объема и добавить стерильные ddH2O компенсации, в случае необходимости. (Примечание: LB агар могут храниться при температуре 4 ° C до использования для подготовки плиты агара фунтов. Тщательно расплава затвердевших агар с помощью микроволновой) | |||
| 5.) когда решение еще теплой (30-40 ° C), добавить ампициллин в конечной концентрации 100 мкг/мл | |||
| 6.) залить около 15 мл жидкости из шага 5.) в стерильные 10 см Петри в стерильных условиях. Когда агар затвердевает, пластины могут храниться для 1 недели при температуре 4 ° C до использования. | |||
| фосфат амортизированное saline (PBS) | Состав: 137 мм NaCl, 2,7 мм KCl, 10 мм Na2HPO4, 1.8 мм х2PO4, 1 мм CaCl2, MgCl 0,5 мм2, рН 7. Стерилизация автоклавированием или фильтрации. | ||
Таблица 1: Stock решения и буфера.
Discussion
Для достижения высокой эффективности включения ncAA, метод auxotrophy на основе SPI основывается на использовании метаболически инженерии хост клеток, которые не способны синтезировать соответствующий естественной коллегой ncAA. Для е. coliтакие штаммы являются доступными. Даже одновременное включение нескольких ncAAs в же белков возможно с помощью multiauxotrophic штаммов. Остаток конкретных режим замены и химических репертуар, будучи привязанными к аналогичные химических аналогов можно рассматривать как недостатки. Тем не менее большое количество вариантов белка могут быть изготовлены как естественный бактериальный перевод аппарат выдерживает многочисленные аналоги аминокислоты. Например более чем 50 ncAAs могут быть включены в белки, с использованием в vitro перевода, приходится около 73% всех кодонов генетического кода для назначений40. Кроме того SPI может также позволяют эффективно мультисайтовой маркировки целевой белок41. В принципе методология SPI не ограничивается кишечной палочки, но может работать в любой другой хост и для каждого из канонических 20 аминокислот, при условии наличия ауксотрофных штаммов и определенных культивирования СМИ. Например два аналогов метионина, azidohomoalanine (Aha) и homopropargylglycine (Hpg), коммерчески доступны и используются для маркировки белков и протеомов в различных организмах. Кроме того Aha могут быть произведены внутриклеточно и впоследствии включены в белок42. Этот ncAA особенно подходит для спряжения bioorthogonal например нажмите кнопку химии, разработанной Tirrel и коллегами: например, в завод ткани Arabidopsis thaliana, в Тутовый шелкопряд личинки43, дрозофила клетки44, личинок данио рерио45 , а также клетки млекопитающих, включая нейроны46, белки могут быть маркированы с Aha47,48. Аналогичным образом ГТО аналогов были успешно включены в антимикробных пептидов в ГТО ауксотрофных Lactococcus lactis штаммов49. SPI является также полезным для поля Xenobiology50,,51, который исследует альтернативы основного химического состава жизни. Например на основе предыдущих работ по E. coli52 и53 B. subtilis, штамм E. coli была разработана недавно эволюционной стратегии с селективного давления использовать thienopyrrole вместо индол, что приводит к протеома общесистемной замещения триптофана, thienopyrrole аланина в генетический код54. Как правило каноническое аминокислоты ГТО, который кодируется с одной триплет (UGG), представляет многообещающие мишенью для инженерных белка из-за богатых аспекты химии индола, который предлагает многочисленные химические изменения. Недавно и в качестве альтернативы на основе SPI инкорпорации роман, которую SCS платформы способны включить ГТО аналогов site-specifically бактерий и эукариот хостов был сообщили55. Это далее расширяет панель в естественных условиях на основе ncAA белка техники, включая изменение спектральных свойств.
Помимо использования ауксотрофных выражение хостов протокол SPI требует строгого ферментации условий, как с точки зрения целевого выражения сроки и состав среды для достижения высокой эффективности включения ncAA и целевого белка урожай 56. Культивирование осуществляется с использованием химически определенные минимальные СМИ, которые по существу содержат помимо основных солей источники для азота (соли аммония) и углерода (D-глюкозы), витамины и микроэлементы. Хотя строго говоря, не требуется в случае отсутствия дальнейших auxotrophies, остальные аминокислоты (20 -n, если n аминокислоты должны быть заменены) часто добавляются для содействия росту бактерий57. На этапе первоначального роста до индукции целевых белков n канонические аминокислоты замены добавляются ограничения концентрации. Клеточного роста продолжается до тех пор, пока целевой незаменимые аминокислоты истощаются, как экспериментально обозначается стационарных ОД600. Впоследствии питательной среды заменяется свежей средой, которая не хватает обедненного аминокислоты и содержит ncAA в изобилии концентрации. Для рибосомальной включения триптофан аналогов, как показано в настоящем Протоколе подается индола аналоговый, который становится внутриклеточно преобразованы в соответствующие производные триптофан триптофан синтазы58. Далее целевых белков индуцируется. На этой стадии клетки близки к концу логарифмического роста, как баланс между общей ячейки номер и фитнес. Как присутствие и включение канонической амино приведет к одичал тип протеина производства, важно обеспечить что незаменимые аминокислоты полностью разряжен до индукции. Аналогичным образом он является обязательным для изучения эффективности ncAA включение в целевого белка, обычно по масс-спектрометрии. В случае существенного присутствия канонической аминокислота, условий выращивания необходимо скорректировать, например, путем изменения концентрации основных amino acid(s) для этапа первоначального роста или продолжительность последнего. В случае низкой Орсе деятельности к ncAA гиперэкспрессия эндогенных ферментов или совместного выражения различных Орсе, который является более активным, к ncAA, может быть проведены59.
Канонический аминокислота ГТО наделен три замечательных особенностей: (i) его естественной изобилия в белках низка; (ii) его биофизические и химических свойств являются уникальными (например., это обычно доминирующим происхождение внутренней флуоресценции белков и пептидов) и (iii) она способствует целый ряд биохимических взаимодействий и функций, включая Π-укладка, H-склеивание и катион π взаимодействие. Все эти особенности радикально изменили на ГТО → 4-амино Trp замещения в GdFP. Помимо сомнения, Дизайн «золото» класса avGFPs является замечательным примером для инженерных индивидуальные autofluorescent белков. С различных спектральных свойств FPs могут быть настроены на определенных спектральных windows через мутагенеза и ncAA включение. В случае GdFP это достигается путем простого обмена H → NH2 в рамках химического кольца индола, содержащихся в ECFP хромофора триады. Рисунок 5 показывает последствия включения ncAA в пределах хромофора. Введение электрон donating группы из 4-амино индол (внутриклеточно преобразованы в 4-амино ГТО) позволяет целый ряд мезомерных структур, которые могут объяснить стабилизированный возбужденное состояние. Спектроскопически его расширенном Стоксов сдвиг и перешли красное флуоресцирование выбросов в результате этих различных свойств расширенной конъюгированных системы. Как сообщалось ранее, перенос расширенной внутримолекулярной заряда в GdFP хромофора изначально чувствителен к рН (рис. 4B) и сопровождается более крупные изменения в момент диполя между землей0 S и S1 возбужденные состояния по отношению к ECFP33. Как альтернативных групп, жертвуя электрон триптофан аналогов, принимая кольцо индола, заменяется гидрокси группы могут использоваться, как сообщалось в сравнительное исследование с модель белка барстар41.
Спектры поглощения и флуоресценции GdFP расширен по сравнению с ECFP и EGFP (рис. 3 c и D). Однородного расширения полос поглощения и флуоресценции обычно вызвано колебательных режимов в хромофора и, Кроме того, сцепление хромофора далее колебательных режимах в белков60. Муфта для окружающей среды местными белка поддерживается обвинения, локализуются на хромофора. Поддерживаются как структурной неоднородности белка приводит к местные вариации вибронных спектра, такая связь между вибронных спектров хромофора и на остальной части белка делокализация заряда и мезомерных государствами, как указано в Рисунок 5. Это соединение также поддерживает большой Стоксов сдвиг и обязательно уменьшает квантовый выход флуоресценции. По сравнению с другой красный смещается FPs GdFP даже экспонаты улучшение белка стабильности и низкая склонность для агрегирования33,,6162. Это не только отличается по цвету от других вариантов FP, но также экспонаты существенно повышенной термостойкостью и расширение совместных складывания33. Его интенсивность флуоресценции составляет по меньшей мере 90% сохранились после нагрева до 60 ° C, в то время как ECFP флуоресценции уменьшается до около 30%. В белках ароматические аминокислоты часто способствуют сети взаимодействующих боковых цепей, которые часто оказывают стабилизирующее воздействие на третичную структуру протеина. avGFP гавани такой стороны цепи сети, которая состоит из хромофора, сам, а также как Пе-165, его-148 и Tyr-145. Эти боковые цепи не только в GdFP структура33довольно жесткой, но главное, они формируют гидрофобные контакты с хромофора. Наиболее характерной особенностью романа в GdFP является более проксимальнее Phe-165 Аминированных хромофора. Это взаимодействие — это функция, не наблюдается в других известных avGFPs. Как 3.2-4.5 Å друг от друга, два остатков аминокислот ароматические взаимодействия могут присутствовать также. Вместе с аминирования индуцированной резонанс стабилизации хромофора эти скорее стабилизировать этой сети гидрофобных аминокислот в духе сотрудничества. Более эффективной передачи внутримолекулярной заряда могут поддерживаться эти взаимодействия в возбуждённом состоянии по сравнению с государственной земли хромофора, а он по крайней мере частично приходится 108 Нм Стокса сдвиг33,62 .
В рациональный дизайн Флюорофор свойства приведет к красно смещается возбуждения волны прогнозируется увеличение размера делокализованных π-системы. Это правило является повиновано серии аминокислот в 66 ведущие позиции к нейтральной хромофоры: пластинчатые теплообменники (λМакс = 355 Нм) < его (λМа x= 386 Нм) < Tyr (λМакс = 395 Нм) < ГТО (λМакс = 436 Нм)63. В природе это расширение хромофора конъюгированных системы π-облигаций было достигнуто на различные стратегии. Для DsRed от Discosoma горлицаон продлевается на интеграции дополнительных аминокислота, таким образом ветра λМакс 573 Нм64. Хромофора asFP595 (λМакс = 595 Нм) от Anemonia бороздчатая был продлен имино группой, увеличивая его π-системы65. Так как хромофора GdFP и другие avFPs того же размера, другой принцип должны влечь за собой длина волны излучения в диапазоне расширенной DsRed и asFP595 хромофоры. Глубокие Стоксов сдвиг 108 Нм приписывается четкую структуру хромофора GdFP, который показывает новый принцип изучены фотофизические в дизайн autofluorescent белков. Предварительные расчеты (как сообщалось в 62) показали, что дипольный момент возбужденные состояния хромофора GdFP значительно больше, чем в основном состоянии, в отличие от соответствующих значений ECFP. Дипольный момент GdFP увеличивается от ~ 3 D (Дебая) в состояние0 S до ~ 15 D в S1, в то время как для ECFP хромофора изменение довольно умеренные (от D ~ 4 ~ 6 D). Таким образом уникальные золотые флуоресценции GdFP обусловлено существенной внутримолекулярной заряда передачи в пределах хромофора, который увеличивает разнообразие возможных мезомерных структур (см. Рисунок 5), которые позволяют для стабилизации резонанс. Это уменьшает уровень энергии, от которого происходит выбросов. В результате глубоких изменений в момент диполя после возбуждения внутримолекулярной заряда разделение является основной причиной изменений в электростатического потенциала окружающей среды хромофора. Окружающие матрица белка, в свою очередь, подстраивается под изменения в распределении заряда после возбуждения хромофора. Последующей структурной релаксации понижает уровень энергии возбужденных хромофора, который сдвигает спектра флуоресценции Красного из-за его характера переноса заряда. По той же причине, вследствие большой Стоксов сдвиг и повышение ставок radiationless процессов, флуоресценции квантовый выход GdFP уменьшается по сравнению с ECFP33.
Высокий квантовый выход и небольших Стоксов сдвиг ECFP и EGFP обычно приписывается среде жесткой белка хромофора, который уменьшает степень свободы и, следовательно, внутреннее преобразование в пользу радиационного релаксации возбужденного состояния 66. Следовательно, молекулярный дизайн более жестко встроенный хромофоры с сокращением муфта к матрице оставшиеся белок может служить в качестве руководства производить дальше красно смещается GFP производных с высоким флуоресценции квантовый выход. Таким образом для дальнейшего инженерные подходы к производят красный смещается autofluorescent белки, расширение системы π-электронов и жесткой хромофора структуры с слабого сцепления белка окружающей среды весьма желательно. Такие изменения также могут быть введены непосредственно в на основе GFP хромофоры либо путем размещения желаемого ncAAs в близости хромофора.
Disclosures
Авторы заявляют, что они не имеют никаких финансовых интересов.
Acknowledgments
Эта работа была поддержана в Германии исследовательский фонд (кластер передового опыта «унификации понятий в катализ) т.ф. и Н.Б. и федерального министерства образования и науки (BMBF программа «HSP-2020», ту-WIMIplus проекта SynTUBio) для ф-и.с.
Materials
| Name | Company | Catalog Number | Comments |
| Chemicals | |||
| 4-aminoindole | Sigma-Aldrich | 525022 | |
| acetonitrile | VWR | HiPerSolv CHROMANORM ULTRA for LC-MS, 83642 | LC-MS grade required |
| agar-agar | Carl Roth | 5210 | |
| ammonium molybdate ((NH4)2MoO4) | Sigma-Aldrich | 277908 | |
| ammonium sulfate ((NH4)2SO4) | Sigma-Aldrich | A4418 | |
| ampicillin sodium salt | Carl Roth | K029 | |
| biotin | Sigma-Aldrich | B4501 | |
| bromophenol blue | Sigma-Aldrich | B0126 | |
| calcium chloride (CaCl2) | Sigma-Aldrich | C5670 | |
| colloidal silica | Sigma-Aldrich | Ludox HS-40, 420816 | |
| Coomassie Brillant Blue R 250 | Carl Roth | 3862 | |
| copper sulfate (CuSO4) | Carl Roth | CP86.1 | |
| D-glucose | Carl Roth | 6780 | |
| di-sodium hydrogen phosphate (Na2HPO4) | Carl Roth | X987 | |
| di-potassium hydrogen phosphate (K2HPO4) | Carl Roth | P749.1 | |
| 1,4-dithiothreitol (DTT) | Carl Roth | 6908 | |
| DNase I | Sigma-Aldrich | D5025 | |
| ethanol | Carl Roth | 9065.1 | |
| formic acid | VWR | HiPerSolv CHROMANORM for LC-MS, 84865 | LC-MS grade required |
| glycerol | Carl Roth | 3783 | |
| imidazole | Carl Roth | X998 | |
| indole | Sigma-Aldrich | I3408 | |
| iron(II) chloride (FeCl2) | Sigma-Aldrich | 380024 | |
| isopropanol | Carl Roth | AE73.1 | |
| isopropyl β-D-1-thiogalactopyranoside (IPTG) | Sigma-Aldrich | I6758 | |
| lysozyme | Sigma-Aldrich | L6876 | |
| magnesium chloride (MgCl2) | Carl Roth | KK36.1 | |
| magnesium sulfate (MgSO4) | Carl Roth | 8283.2 | |
| manganese chloride (MnCl2) | Sigma-Aldrich | 63535 | |
| β-mercaptoethanol | Carl Roth | 4227.3 | |
| potassium chloride (KCl) | Carl Roth | 6781.3 | |
| potassium dihydrogen phosphate (KH2PO4) | Sigma-Aldrich | P5655 | |
| RNase A | Carl Roth | 7156 | |
| sodium chloride (NaCl) | Carl Roth | P029 | |
| sodium dihydrogen phosphate (NaH2PO4) | Carl Roth | T879 | |
| sodium dodecyl sulphate (NaC12H25SO4) | Carl Roth | 0183 | |
| thiamine | Sigma-Aldrich | T4625 | |
| Tris(hydroxymethyl)-aminomethane (Tris) | Carl Roth | 5429 | |
| Tris hydrochloride (Tris-HCl) | Sigma-Aldrich | 857645 | |
| tryptone | Carl Roth | 8952 | |
| yeast extract | Carl Roth | 2363 | |
| zinc chloride (ZnCl2) | Sigma-Aldrich | 229997 | |
| Name | Company | Catalog Number | Comments |
| amino acids | |||
| L-alanine | Sigma-Aldrich | A7627 | |
| L-arginine | Sigma-Aldrich | A5006 | |
| L-asparagine | Sigma-Aldrich | A8381 | |
| L-aspartic acid | Sigma-Aldrich | A0884 | |
| L-cysteine | Sigma-Aldrich | C7352 | |
| L-glutamic acid | Sigma-Aldrich | G2128 | |
| L-glutamine | Sigma-Aldrich | G3126 | |
| L-glycine | Sigma-Aldrich | G7126 | |
| L-histidine | Sigma-Aldrich | H8000 | |
| L-isoleucine | Sigma-Aldrich | I2752 | |
| L-leucine | Sigma-Aldrich | L8000 | |
| L-lysine | Sigma-Aldrich | L5501 | |
| L-methionine | Sigma-Aldrich | M9625 | |
| L-proline | Sigma-Aldrich | P0380 | |
| L-phenylalanine | Sigma-Aldrich | P2126 | |
| L-serine | Sigma-Aldrich | S4500 | |
| L-threonine | Sigma-Aldrich | T8625 | |
| L-tryptophan | Sigma-Aldrich | T0254 | |
| L-tyrosine | Sigma-Aldrich | T3754 | |
| L-valine | Sigma-Aldrich | V0500 | |
| Name | Company | Catalog Number | Comments |
| Lab materials | |||
| 0.45 µm syringe filter with PVDF membrane | Carl Roth | CCY1.1 | |
| 1.5 mL microcentrifuge tubes | Eppendorf | 30120086 | |
| conical polystyrene (Falcon) tubes, 50 mL | Fisher Scientific | 14-432-22 | |
| Luer-Lock syringe 5 mL | Carl Roth | EP96.1 | |
| dialysis membrane, Molecular Weight Cut-Off (MWCO) 5,000 | Spectrum Medical Industries | Spectra/Por MWCO 5000 dialysis membrane, 133198 | |
| Immobilized Metal ion Affinity Chromatography (IMAC) column 1 mL, Ni-NTA | Macherey Nagel | Protino series, 745410.5 | |
| petri dishes (polystyrene, sterile) | Carl Roth | TA19 | |
| pQE-80L plasmid vector | Qiagen | no longer available | replaced by N-terminus pQE Vector set Cat No./ID: 32915 |
| protein extraction reagent BugBuster | EMB Millipore | 70921-4 | |
| round-bottom polystyrene tubes, 14 mL | Fisher Scientific | Corning Falcon, 14-959-1B | |
| Trp-auxotrophic E. coli strain | ATCC | ATCC 49980 | Bridges BA et al., Chem Biol Interact., 1972, 5(2):77-84; see main text for alternatives |
| Name | Company | Catalog Number | Comments |
| Mass Spectrometry equipment | |||
| mass spectrometer for LC-ESI-TOF-MS | Agilent | Agilent 6530 Accurate-Mass QTOF | coupled with Infinity LC system |
| mass spectrometry data analysis software | Agilent | MassHunter Qualitative Analysis software v. B.06.00 | |
| High-Performance Liquid Chromatography (HPLC) column for LC-ESI-TOF-MS | Sigma-Aldrich | Supelco Discovery BIO Wide Pore C5 HPLC column, 3 µm particle size, 10 cm x 2.1 mm | |
| HPLC autosampler vials 1.5 mL | Sigma-Aldrich | Supelco 854165 | with conical 0.1 mL glass inserts, screw caps and septa |
| Name | Company | Catalog Number | Comments |
| General equipment | |||
| benchtop centrifuge for 1.5 mL Eppendorf tubes | Eppendorf | 5427 R | |
| cooling centrifuge for 50 mL Falcon tubes | Eppendorf | 5810 R | |
| high pressure microfluidizer for bacterial cell disruption | Microfluidics | LM series with “Z” type chamber | |
| peristaltic pump for LC | GE Healthcare | P-1 | |
| Fast Protein Liquid Chromatography (FPLC) system | GE Healthcare | ÄKTA pure 25 L | |
| orbital shaker for bacterial cultivation | Infors HT | Minitron | |
| UV/Vis spectrophotometer | Biochrom | ULTROSPEC 2100 | |
| ultrasonic homogenizer for bacterial cell disruption | Omnilab | Bandelin SONOPULS HD 3200, 5650182 | with MS72 sonifier tip |
| Name | Company | Catalog Number | Comments |
| Fluorescence spectroscopy equipment | |||
| ps-pulsed laser 470 nm | Picoquant GmbH | PDL-470 | |
| time- and wavelength-correlated single photon counting (TWSPC) acquisition software | Picoquant GmbH | SymPhoTime 64 | |
| time- and wavelength-correlated single photon counting (TWSPC) detector | Picoquant GmbH | PML-16C | 16 spectral channels, to be selected by grating settings |
| single photon counting software | Picoquant GmbH | SPCM 9.75 | |
| global fitting software | Picoquant GmbH | SPC2Glo(R) | |
| fluorescence decay data analysis software | Picoquant GmbH | FluoFit program | |
| data analysis software | OriginLab Inc. | Origin 9.2 | |
| neutral density filter set | Schott | NG1 to NG11 | (400 - 650 nm, transmission 50 %, 20%, 10 %, 5 %) |
| 488 nm long-pass emission filter | AHF Analysentechnik | AHF-488 | |
| quartz cuvette | Thorlabs GmbH | CV10Q1400 | 1 cm pathlength |
References
- Shimomura, O., Johnson, F. H., Saiga, Y. Extraction, Purification and Properties of Aequorin, a Bioluminescent Protein from the Luminous Hydromedusan, Aequorea. J Cell Compar Physl. 59 (3), 223-239 (1962).
- Chalfie, M., Tu, Y., Euskirchen, G., Ward, W. W., Prasher, D. C. Green fluorescent protein as a marker for gene expression. Science. 263 (5148), 802-805 (1994).
- Andresen, M., et al. Structure and mechanism of the reversible photoswitch of a fluorescent protein. P Natl Acad Sci USA. 102 (37), 13070-13074 (2005).
- Andresen, M., et al. Structural basis for reversible photoswitching in Dronpa. P Natl Acad Sci USA. 104 (32), 13005-13009 (2007).
- Brakemann, T., et al. A reversibly photoswitchable GFP-like protein with fluorescence excitation decoupled from switching. Nat Biotechnol. 29 (10), 942-947 (2011).
- Kremers, G. -J., Gilbert, S. G., Cranfill, P. J., Davidson, M. W., Piston, D. W. Fluorescent proteins at a glance. J Cell Sci. 124 (Pt 2), 157-160 (2011).
- Shimomura, O. Structure of the chromophore of aequorea 0. shimomura green fluorescent protein. FEBS Lett. 104 (2), 220-222 (1979).
- Shaner, N. C., Campbell, R. E., Steinbach, P. A., Giepmans, B. N. G., Palmer, A. E., Tsien, R. Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat Biotechnol. 22 (12), 1567-1572 (2004).
- Shcherbo, D., et al. Bright far-red fluorescent protein for whole-body imaging. Nat Methods. 4 (9), 741-746 (2007).
- Shcherbakova, D. M., Subach, O. M., Verkhusha, V. V. Red fluorescent proteins: advanced imaging applications and future design. Angew Chem Int Edit. 51 (43), 10724-10738 (2012).
- Stepanenko, O. V., Verkhusha, V. V., Kuznetsova, I. M., Uversky, V. N., Turoverov, K. K. Fluorescent proteins as biomarkers and biosensors: throwing color lights on molecular and cellular processes. Curr Protein Pept Sc. 9 (4), 338-369 (2008).
- Wang, L., Xie, J., Deniz, A. A., Schultz, P. G. Unnatural amino acid mutagenesis of green fluorescent protein. J Org Chem. 68 (1), 174-176 (2003).
- Budisa, N., Steipe, B., Demange, P., Eckerskorn, C., Kellermann, J., Huber, R. High-level biosynthetic substitution of methionine in proteins by its analogs 2-aminohexanoic acid, selenomethionine, telluromethionine and ethionine in Escherichia coli. Eur J Biochem. 230 (2), 788-796 (1995).
- Sharma, N., Furter, R., Kast, P., Tirrell, D. A. Efficient introduction of aryl bromide functionality into proteins in vivo. FEBS Lett. 467 (1), 37-40 (2000).
- Liu, C. C., Schultz, P. G. Adding new chemistries to the genetic code. Annu Rev Biochem. 79, 413-444 (2010).
- Twine, S. M., Murphy, L., Phillips, R. S., Callis, P., Cash, M. T., Szabo, A. G. The Photophysical Properties of 6-Azaindole. J Phys Chem B. 107 (2), 637-645 (2003).
- Lepthien, S., Hoesl, M. G., Merkel, L., Budisa, N. Azatryptophans endow proteins with intrinsic blue fluorescence. P Natl Acad Sci USA. 105 (42), 16095-16100 (2008).
- Budisa, N., et al. Probing the role of tryptophans in Aequorea victoria green fluorescent proteins with an expanded genetic code. Biol Chem. 385 (2), 191-202 (2004).
- Ross, J. B., et al. Spectral enhancement of proteins: biological incorporation and fluorescence characterization of 5-hydroxytryptophan in bacteriophage lambda cI repressor. P Natl Acad Sci USA. 89 (24), 12023-12027 (1992).
- Soumillion, P., Jespers, L., Vervoort, J., Fastrez, J. Biosynthetic incorporation of 7-azatryptophan into the phage lambda lysozyme: Estimation of tryptophan accessibility, effect on enzymatic activity and protein stability. Protein Eng Des Sel. 8 (5), 451-456 (1995).
- Heim, R., Tsien, R. Y. Engineering green fluorescent protein for improved brightness, longer wavelengths and fluorescence resonance energy transfer. Curr Biol. 6 (2), 178-182 (1996).
- Bridges, B. A., Mottershead, R. P., Rothwell, M. A., Green, M. H. L. Repair-deficient bacterial strains suitable for mutagenicity screening: tests with the fungicide captain. Chem Biol Interact. 5 (2), 77-84 (1972).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Bacterial Transformation: The Heat Shock Method. J Vis Exp. , (2017).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Bacterial Transformation: Electroporation. J Vis Exp. , (2017).
- Grigorenko, B. L., Krylov, A. I., Nemukhin, A. V. Molecular modeling clarifies the mechanism of chromophore maturation in the green fluorescent protein. J Am Chem Soc. , (2017).
- JoVE Science Education Database. General Laboratory Techniques. Introduction to the Spectrophotometer. J Vis Exp. , (2017).
- Goedhart, J., et al. Structure-guided evolution of cyan fluorescent proteins towards a quantum yield of 93%. Nat Commun. 3, 751 (2012).
- Neidhardt, F. C., Bloch, P. L., Smith, D. F. Culture medium for enterobacteria. J Bacteriol. 119 (3), 736-747 (1974).
- Hörnsten, E. G. On culturing Escherichia coli on a mineral salts medium during anaerobic conditions. Bioprocess Eng. 12 (3), 157-162 (1995).
- Davis, B. D. The Isolation of Biochemically Deficient Mutants of Bacteria by Means of Penicillin. P Natl Acad Sci USA. 35 (1), 1-10 (1949).
- Sambrook, J., Russell, D. W. Molecular Cloning: A Laboratory Manual. , Cold Spring Harbor Laboratory Press. Cold Spring Harbor, NY, USA. (2001).
- Wang, Y. -S., et al. The de novo engineering of pyrrolysyl-tRNA synthetase for genetic incorporation of L-phenylalanine and its derivatives. Mol Biosyst. 7 (3), 714-717 (2011).
- Bae, J. H., et al. Expansion of the genetic code enables design of a novel "gold" class of green fluorescent proteins. J Mol Biol. 328 (5), 1071-1081 (2003).
- JoVE Science Education Database. Dialysis: Diffusion Based Separation. J Vis Exp. , Cambridge, MA. (2017).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Separating Protein with SDS-PAGE. J Vis Exp. , (2017).
- Petrásek, Z., et al. Excitation energy transfer from phycobiliprotein to chlorophyll d in intact cells of Acaryochloris marina studied by time- and wavelength-resolved fluorescence spectroscopy. Photoch Photobio Sci. 4 (12), 1016-1022 (2005).
- Kolber, Z. S., Barkley, M. D. Comparison of approaches to the instrumental response function in fluorescence decay measurements. Anal Biochem. 152 (1), 6-21 (1986).
- Pelet, S., Previte, M. J. R., Laiho, L. H., So, P. T. C. A fast global fitting algorithm for fluorescence lifetime imaging microscopy based on image segmentation. Biophys J. 87 (4), 2807-2817 (2004).
- Loefroth, J. E. Time-resolved emission spectra, decay-associated spectra, and species-associated spectra. J Phys Chem. 90 (6), 1160-1168 (1986).
- Hartman, M. C. T., Josephson, K., Lin, C. -W., Szostak, J. W. An expanded set of amino acid analogs for the ribosomal translation of unnatural peptides. PLoS One. 2 (10), e972 (2007).
- Budisa, N., et al. Global replacement of tryptophan with aminotryptophans generates non-invasive protein-based optical pH sensors. Angew Chem Int Edit. 41 (21), 4066-4069 (2002).
- Ma, Y., Biava, H., Contestabile, R., Budisa, N., di Salvo, M. L. Coupling bioorthogonal chemistries with artificial metabolism: intracellular biosynthesis of azidohomoalanine and its incorporation into recombinant proteins. Molecules. 19 (1), 1004-1022 (2014).
- Teramoto, H., Kojima, K. Incorporation of Methionine Analogues Into Bombyx mori Silk Fibroin for Click Modifications. Macromol Biosci. 15 (5), 719-727 (2015).
- Deal, R. B., Henikoff, J. G., Henikoff, S. Genome-wide kinetics of nucleosome turnover determined by metabolic labeling of histones. Science. 328 (5982), 1161-1164 (2010).
- Hinz, F. I., Dieterich, D. C., Tirrell, D. A., Schuman, E. M. Non-canonical amino acid labeling in vivo to visualize and affinity purify newly synthesized proteins in larval zebrafish. ACS Chem Neurosci. 3 (1), 40-49 (2012).
- Dieterich, D. C., et al. In situ visualization and dynamics of newly synthesized proteins in rat hippocampal neurons. Nat Neurosci. 13 (7), 897-905 (2010).
- Dieterich, D. C., Link, A. J., Graumann, J., Tirrell, D. A., Schuman, E. M. Selective identification of newly synthesized proteins in mammalian cells using bioorthogonal noncanonical amino acid tagging (BONCAT). P Natl Acad Sci USA. 103 (25), 9482-9487 (2006).
- Glenn, W. S., et al. Bioorthogonal Noncanonical Amino Acid Tagging (BONCAT) Enables Time-Resolved Analysis of Protein Synthesis in Native Plant Tissue. Plant Physiol. 173 (3), 1543-1553 (2017).
- Zhou, L., et al. Incorporation of tryptophan analogues into the lantibiotic nisin. Amino Acids. 48 (5), 1309-1318 (2016).
- Acevedo-Rocha, C. G., Budisa, N. Xenomicrobiology: a roadmap for genetic code engineering. Microb Biotechnol. 9 (5), 666-676 (2016).
- Agostini, F., Völler, J. -S., Koksch, B., Acevedo-Rocha, C. G., Kubyshkin, V., Budisa, N. Biocatalysis with Unnatural Amino Acids: Enzymology Meets Xenobiology. Angew Chem Int Edit. 56 (33), 9680-9703 (2017).
- Bacher, J. M., Ellington, A. D. Selection and characterization of Escherichia coli variants capable of growth on an otherwise toxic tryptophan analogue. J Bacteriol. 183 (18), 5414-5425 (2001).
- Wong, J. T. Membership mutation of the genetic code: loss of fitness by tryptophan. Pc Natl Acad Sci USA. 80 (20), 6303-6306 (1983).
- Hoesl, M. G., et al. Chemical Evolution of a Bacterial Proteome. Angew Chem Int Edit. 54 (34), 10030-10034 (2015).
- Italia, J. S., et al. An orthogonalized platform for genetic code expansion in both bacteria and eukaryotes. Nat Chem Biol. 13 (4), 446-450 (2017).
- Völler, J. -S., Thi To, T. M., Biava, H., Koksch, B., Budisa, N. Global substitution of hemeproteins with noncanonical amino acids in Escherichia coli with intact cofactor maturation machinery. Enzyme Microb Tech. 106, 55-59 (2017).
- Budisa, N., Steipe, B., Demange, P., Eckerskorn, C., Kellermann, J., Huber, R. High-level biosynthetic substitution of methionine in proteins by its analogs 2-aminohexanoic acid, selenomethionine, telluromethionine and ethionine in Escherichia coli. Eur J Biochem. 230 (2), 788-796 (1995).
- Völler, J. -S., Budisa, N. Coupling genetic code expansion and metabolic engineering for synthetic cells. Curr Opin Biotech. 48, 1-7 (2017).
- Johnson, J. A., Lu, Y. Y., Van Deventer, J. A., Tirrell, D. A. Residue-specific incorporation of non-canonical amino acids into proteins: recent developments and applications. Curr Opin Chem Biol. 14 (6), 774-780 (2010).
- Somsen, O. J., van Grondelle, R., van Amerongen, H. Spectral broadening of interacting pigments: polarized absorption by photosynthetic proteins. Biophys J. 71 (4), 1934-1951 (1996).
- Kurschus, F. C., Pal, P. P., Bäumler, P., Jenne, D. E., Wiltschi, B., Budisa, N. Gold fluorescent annexin A5 as a novel apoptosis detection tool. Cytom Part A. 75 (7), 626-633 (2009).
- Lepthien, S., Wiltschi, B., Bolic, B., Budisa, N. In vivo engineering of proteins with nitrogen-containing tryptophan analogs. Appl Microbiol Biot. 73 (4), 740-754 (2006).
- Wachter, R. M., Elsliger, M. -A., Kallio, K., Hanson, G. T., Remington, S. J. Structural basis of spectral shifts in the yellow-emission variants of green fluorescent protein. Structure. 6 (10), 1267-1277 (1998).
- Verkhusha, V. V., Lukyanov, K. A. The molecular properties and applications of Anthozoa fluorescent proteins and chromoproteins. Nat Biotechnol. 22 (3), 289-296 (2004).
- Martynov, V. I., Savitsky, A. P., Martynova, N. Y., Savitsky, P. A., Lukyanov, K. A., Lukyanov, S. A. Alternative cyclization in GFP-like proteins family. The formation and structure of the chromophore of a purple chromoprotein from Anemonia sulcata. J Biol Chem. 276 (24), 21012-21016 (2001).
- Piatkevich, K. D., Malashkevich, V. N., Morozova, K. S., Nemkovich, N. A., Almo, S. C., Verkhusha, V. V. Extended Stokes shift in fluorescent proteins: chromophore-protein interactions in a near-infrared TagRFP675 variant. Sci Rep. 3 (1), 1847 (2013).
