

Summary
Synthetische biologie waardoor de engineering van eiwitten met ongekende eigenschappen met behulp van het co-translational inbrengen van niet-canonieke aminozuren. Hier, presenteerden we hoe een spectraal rood-verschoven variant van een GFP-type fluorophore met roman fluorescentie spectroscopische eigenschappen, genoemd "goud" fluorescent proteïne (GdFP), wordt geproduceerd in E. coli via selectieve druk opneming (SPI).
Abstract
Fluorescerende eiwitten zijn fundamentele hulpmiddelen voor de biowetenschappen, met name voor fluorescentie microscopie van levende cellen. Terwijl wild-type en gemanipuleerde varianten van de groen fluorescente proteïne uit Aequorea victoria (avGFP), alsmede van andere diersoorten homologen cover al grote delen van het optische spectrum, blijft een spectrale kloof in de regio nabij-infrarood, voor welke avGFP gebaseerde fluorophores zijn niet beschikbaar. Red-verschoven fluorescente proteïne (FP) varianten zou aanzienlijk uitbreiden van de toolkit voor de spectrale randverwijdering van meerdere moleculaire soorten, maar de natuurlijk voorkomende rood-verschoven FPs afgeleid van koralen of zeeanemonen hebben lagere fluorescentie quantumrendement en inferieur foto-stabiliteit t.o.v. de avGFP varianten. Verdere manipulatie en de mogelijke uitbreiding van de chromofore van geconjugeerd systeem ten aanzien van de far-red spectrale regio wordt ook beperkt door het repertoire van 20 canonieke aminozuren voorgeschreven door de genetische code. Deze beperkingen wilt opheffen, kunnen synthetische biologie verder spectrale rood-shifting via inbrengen van niet-canonieke aminozuren in de chromofore triade. We beschrijven de toepassingvan SPI de varianten van de avGFP van de ingenieur met nieuwe spectrale eigenschappen. Eiwit expressie wordt uitgevoerd in een tryptofaan-auxotrofe E. coli stam en door groei media aan te vullen met geschikte indool precursoren. In de cellen, zijn deze precursoren geconverteerd naar de corresponderende tryptofaan-analogen en eiwitten ingelijfd door de ribosomal machine in reactie op UGG codonen. De vervanging van Trp-66 in de verbeterde "cyaan" variant van avGFP (ECFP) door een elektron-doneren 4-aminotryptophan resulteert in GdFP met een 108 nm Stokes shift en een maximale sterk rood-verschoven emissie (574 nm), terwijl het thermodynamisch stabieler dan zijn voorganger ECFP. Residu-specifieke opneming van de niet-canonieke aminozuur wordt geanalyseerd door de Spectrometrie van de massa. De spectroscopische eigenschappen van GdFP worden gekenmerkt door time-resolved Fluorescentiespectroscopie als één van de waardevolle toepassingen van genetisch gecodeerde bps in biowetenschappen.
Introduction
Sinds de ontdekking van de groen fluorescente proteïne in de kwal Aequorea victoria (avGFP) in 19621 en de eerste heterologe expressie in 19942 in andere eukaryotische cellen, zijn fluorescente proteïnen van het GFP-familie geworden zeer waardevolle hulpmiddelen en doelen in de biowetenschappen. Uitgebreide genetische en moleculaire techniek opgenomen de aanpassing van de soortspecifieke codon gebruik, versnelling van vouwen, verbeterde rijping, verhoogde helderheid, preventie van oligomerisatie en afstemming van de spectrale en fotochemische eigenschappen inclusief de mogelijkheid om omkeerbaar photoswitch3,4,5,6. GFP dankt haar fluorescentie van haar 4-(p- hydroxybenzylidene) imidazolidin-5-one (HBDI) chromofoor. De laatste autocatalytically ontstaat uit de zogenaamde chromofore triade van aminozuren (Ser-65/Tyr-66/Glycine-67 in avGFP) na de vorming van een extra covalente binding binnen de ruggengraat van de peptide onder invloed van moleculaire zuurstof7. Het resoneert gestabiliseerde geconjugeerd systeem communiceert dynamisch met de moleculaire omgeving beschikbaar, zodat voor de absorptie in het zichtbare bereik en karakteristieke groene fluorescentie van deze proteïnen.
Binnen de chromofore godentrias is de aanwezigheid van een aromatisch aminozuur verplicht. Echter, de standaard aminozuur repertoire bestaat uit slechts vier aromatische residuen (zijn, Phe, Trp en Tyr). Dit houdt in dat conventionele mutagenese benaderingen om aanzienlijk meer varianten van het avGFP rood-verschoven ten opzichte van de meest rood-verschoven natuurlijke FPs zoals DsRed8 van Discosoma striata -coralimorphs of mKate/mNeptune9 van de zeeanemonen Entacmaea quadricolor. Dus, het far-red en nabij-infrarood gedeelte van het optisch spectrum boven 600 nm is dun vallende GFP varianten. Dit is natuurlijk een ernstige beperking voor fluorescentie microscopische benaderingen die vereisen spectrale demultiplexing van verschillende soorten van de fluorophore op hetzelfde moment. Bijvoorbeeld lange golflengte markeringen zijn ook nodig om het maken van het regime van de laag absorptie van huidweefsel tussen 700 en 1000 nm in instellingen voor diepe weefsel imaging10gebruiken.
Fluorescerende van avGFP afgeleide eiwitten zijn onderverdeeld in verschillende klassen op basis van de spectroscopische eigenschappen en chemische aard van hun chromophores11. Met haar triade Ser-65/Tyr-66/Glycine-67, de wild-type chromofore bestaat als een equilibrated mengsel tussen de neutrale, fenolische vorm (λmax = 395 nm, ε = 21.000 M-1cm-1) en de anionogene phenolate vorm (λmax = 475 nm, ε = 7100 M -1cm-1), en het emissiespectrum vertoont een één piek op 508 nm. De hydroxylgroep van Ser-65 is van cruciaal belang, aangezien het schenkt een H-obligatie te Glu-222 in de omgeving van chromofoor (afstand: 3,7 Å), dat bevordert de ionisatie van deze carboxylaat. Klasse ik wordt gekenmerkt door een chromofore anionogene phenolate, zoals in EGFP (Phe-64-Leu/Ser-65-Thr;max λ = 488 nm, ε = 35.600 M-1cm-1, λem = 509 nm). Als gevolg van de vervanging van de Ser-65-Thr(Ala,Gly), de 395 nm excitatie piek van het neutrale fenol-formulier wordt onderdrukt en de 470-475 nm piek van de anionactieve phenolate is vijf - tot zes keer uitgebreid en verschoven naar 490 nm. Klasse II bestaat uit eiwitten met een neutrale fenolische chromofore, zoals saffier-GFP. Hier, de vervanging van Thr-203-Ile bijna volledig onderdrukt de 475 nm excitatie, waardoor alleen de piek op 399 nm. Aangezien de anionogene chromofore niet naar behoren solvated zijn, is zijn neutrale vorm favoriet. Klasse III bestaat uit de "gele" fluorescerend varianten (EYFP; Ser-65-Gly/val-68-Leu/ser-72-Ala/thr-203-Tyr; Λmax ε = 514 nm, ε = 84, 600 M-1cm-1, λem = 527 nm) met π-stapelen interactie van een aromatische zijketen en de phenolate, zoals veroorzaakt door de Thr-203-His(Trp,Phe,Tyr) vervangingen, die leiden tot maximaal 20 nm rood-verschoven emissie-maxima (Thr-203-Tyr). Verdere substitutie (Gln-69-Lys) resulteert in een ander 1-2 nm red shift tot 529 nm, de meest rood-verschoven avGFP variant11bekend. De uitwisseling van de fenol voor een indool (Tyr-66-Trp) maakt klasse IV, zoals in de cyaan-belichting fluorescerende ECFP (Ser-65-Thr/Tyr-66-Trp; λmax1 = 434 nm, ε = 24,800 M-1cm-1; λmax2 = 452 nm, ε = 23,600 M-1cm -1 ; Λem1 = 477 nm, λem2 = 504 nm). De accommodatie van de omvangrijke indool is waarschijnlijk ingeschakeld door andere, compenserende mutaties. De ECFP excitatie en emissie-maxima vallen inbetween die van eiwitten met neutrale of anionogene chromophores. Klasse V eiwitten haven een imidazool in plaats van de fenol (Tyr-66-Hsi), bijv., blauw-belichting fluorescerende eiwitten zoals EBFP. Klasse VI wordt geproduceerd door een fenol-naar-fenyl uitwisseling ten gunste van de neutrale chromofore vorm uitsluitend, die bijgevolg tot de meest blauw-verschoven excitatie en uitstoot piek posities leidt (360 nm en 442 nm, respectievelijk).
Klassieke plaats-geleide mutagenese is vooral geschikt voor de productie van roman avGFP chromofore varianten, door de permutatie van de 65-67 tripeptide en interactie residuen in het kader van de canonieke 20 aminozuren. Deze mogelijkheden kunnen verder worden uitgebreid wanneer niet-canonieke varianten van aromatische aminozuren worden ingevoerd tijdens ribosomal eiwit synthese12. In principe zijn er twee manieren om dit te bereiken. De eerste strategie is afhankelijk van de tolerantie van de substraat van het eiwit vertaling machines, met name van het aminoacyl-tRNA synthetases (aaRSs) naar verwante aminozuur-analogen. Om dit te bereiken met een hoog rendement, auxotrofe E. coli expressie zijn stammen die niet in staat zijn om de bijbehorende natuurlijke aminozuur te synthetiseren. Hierdoor is de vervanging van het laatste door het toevoegen van geschikte niet-canonieke aminozuren (ncAAs) of precursoren daarvan aan het kweekmedium. Deze strategie, ook bekend als selectieve druk opneming (SPI)13,14, kunt residu-specifieke vervangingen, die leiden wereldwijde opneming van de ncAA tot. De tweede strategie gebruikt stop codon suppressor tRNAs die belast zijn met de ncAA door ontworpen jaarlijkse activiteitenverslagen van enzymen. Dit resulteert in de readthrough van in-frame stop codonen en laat site-specific ncAA opneming. Bijgevolg leidt deze methode van stop codon onderdrukking (SCS) tot de uitbreiding van de genetische code15. Via mutagenese, een stop codon in het target-gen op de gewenste site geplaatst. In principe, SPI ook inzetbaar maken van recombinante peptides en proteïnen, rekening houdend met een unieke installatie van de ncAA, gezien het feit dat zeldzame canonieke aminozuren zoals BMO of Trp worden gekozen voor vervanging. Met Trp, SPI benaderingen is aangetoond dat ze werken met een grote verscheidenheid aan analogen inclusief 4 - F-, 5 - F - en 6-F-Trp, 7-aza-Trp, 4-OH - en 5-OH-Trp, alsmede 4- en 5-NH2- Trp of zelfs β (thienopyrrolyl) alanine derivaten16 ,17,18,19,20. SPI zou dus zeer gunstig voor het vervangen van aromatische aminozuren van GFP chromophores door niet-canonieke varianten de mogelijkheid om verdere spectra en Stokes verschuiving van deze FPs te verkennen. Wat betreft alle wijzigingen van de volgorde van de eiwitten, moet de verenigbaarheid met FP vouwen en chromofore rijping experimenteel worden getest.
Hierbij maken we gebruik van klasse IV ECFP21, die in plaats van de wild-type avGFP Tyr, een residu Trp binnen haar chromofore triade draagt. Met SPI, wordt dit Trp-66 (en Trp-57, het alleen andere Trp residu in ECFP) vervangen door de 4-amino-Trp. De aanwezigheid van de elektron-doneren aminogroep van 4-amino-Trp binnen de chromofore gunsten de stabilisatie van de resonantie van een geëxciteerde toestand veel rood-verschoven proton overdracht (ESPT) begiftigd met een 108 nm Stokes verschuiving. Deze "gouden" fluorescent proteïne (GdFP) vormt de variant met de grootste rode de verschuiving van de fluorescentie maximale (574 nm) onder alle avGFP afkomstige eiwitten. We beschrijven de methode van GdFP productie van de eiwitten door SPI en de protocollen voorzien in de verplichte analyse van de resulterende gemodificeerde eiwitten door massaspectrometrie. Verder laten we zien hoe de GdFP kan worden gebruikt en geanalyseerd in time-resolved fluorescentie spectroscopie benaderingen.
Protocol
1. de transformatie van Trp-auxotrofe E. coli
- Chemisch transformeren of electrocompetent cellen (50 µL) van een stam van de Trp-auxotrofe E. coli , bv. ATTC 49980 (WP2, mutant afgeleid van E. coli stam B/R22), met 1 µL van een 1 ng/µL waterige oplossing van het pQE - 80 L His6-ECFP-plasmide met warmte schok of electroporation, respectievelijk. Raadpleeg de JoVE Science Education Database23,24 voor meer informatie.
Opmerking: De expressie vector pQE - 80L His6-ECFP codeert een N-terminaal 6 x zijn-gelabeld ECFP21 gedreven door een bacteriële T5 promotor met lac operator. Het draagt verder een AmpR selectie marker en een colE1 oorsprong van replicatie (de pQE - 80 L vector ruggengraat reeks vindt u op: https://www.qiagen.com/mx/resources/resourcedetail?id=c3b71572-4d82-4671-a79b-96357fe926d1&lang=en & autoSuggest = true). De theoretische molecuulgewicht van het eiwit van de wild-type His6-ECFG (na chromofore rijping25) is 28303.92 Da. De vertaalde doel eiwit volgorde is als volgt (zijne-tag onderstreept, vector-afgeleide reeksen in vet): MRGSHHHHHHGSMVSKGEELFTGVVPILVELDGDVNGHKFSVSGEGEGDATYGKLTLKFICTTGKLPVPWPTLVTTLTWGVQCFSRYPDHMK
QHDFFKSAMPEGYVQERTIFFKDDGNYKTRAEVKFEGDTLVNRIELKGIDFKEDGNILGHKLEYNYISHNVYITADKQKNGIKANFKIRHNIEDGS
VQLADHYQQNTPIGDGPVLLPDNHYLSTQSALSKDPNEKRDHMVLLEFVTAAGITLGMDELYK. - Plaat getransformeerde cellen op LB-agar platen (tabel 1) aangevuld met 10 g/L glucose, 100 µg/mL ampicilline en Incubeer de platen bij 37 ° C's nachts.
2. recombinant eiwit expressie
-
Overnachting cultuur van E. coli ATCC 49980 pQE - 80 L His6-ECFP
- Bereiden van 5 mL van LB medium (tabel 1; aangevuld met 10 g/L glucose, 100 µg/mL ampicilline) in een steriele 14 mL polystyreen cultuur buis voor aërobe groei en inoculeren met een afzonderlijk kolonie van een met behulp van een steriele Pipetteer tip of inoculatie lus agarplaat.
Opmerking: Gebruik van kolonies van vers getransformeerde cellen wordt aanbevolen. De platen met bacteriële kolonies (uit stap 1.2.) kunnen worden achtergelaten bij 4 ° C voor meerdere dagen. - Incubeer de cellen bij 37 ° C in een roteerschudapparaat op 200-250 rpm's nachts.
- Bereiden van 5 mL van LB medium (tabel 1; aangevuld met 10 g/L glucose, 100 µg/mL ampicilline) in een steriele 14 mL polystyreen cultuur buis voor aërobe groei en inoculeren met een afzonderlijk kolonie van een met behulp van een steriele Pipetteer tip of inoculatie lus agarplaat.
-
Expressie van wild-type ECFP
- Inoculeer 10 mL verse LB medium (tabel 1; aangevuld met 10 g/L glucose, 100 µg/mL ampicilline) met 100 µL van de overnachting cultuur in een 100 mL conische kolf. Incubeer de kolf bij 37 ° C in een roteerschudapparaat bij 200 omwentelingen per minuut.
Opmerking: Optioneel, deze stap kan worden uitgevoerd in 10 mL NMM19 medium (tabel 1) aangevuld met 100 µg/L ampicilline en 0,5 mM L-tryptofaan (anderzijds indool kan worden gebruikt). - Meten van de extinctie op 600 nm (OD600) elke 20 min. bij voorkeur gemeten celdichtheid door bepaling van de extinctie bij 600 nm (OD600) in een spectrofotometer een cuvet met een weglengte van 1 cm. altijd uitvoeren een verwijzing meting met behulp van het bijbehorende kweekmedium. Verdun de monsters en meng de monsters goed te verkrijgen van een meting waarde van 0.1-0.8 en vervolgens wordt berekend OD600 met behulp van de verdunningsfactor. Voor meer informatie, gelieve te verwijzen naar eerdere publicatie 26.
- Bij het bereiken van een600 OD-waarde van 0.5-0.8 (ongeveer 2-3 uur na de inenting), neemt u monster "before inductie" voor SDS-PAGE (polyacrylamide-gel-elektroforese van natrium dodecyl sulfaat, stap 4).
- Doel eiwituitdrukking veroorzaken door de vloeibare cultuur tot 0.5 mM IPTG (isopropyl β-D-1-thiogalactopyranoside, vanaf 1 M stockoplossing) aan te passen en Incubeer het bij 30 ° C in een roteerschudapparaat bij 200 omwentelingen per minuut gedurende 4-8 uur.
Opmerking: Cyaan fluorescente proteïnen zijn gewoonlijk uitgedrukt bij temperaturen onder de 37 ° C27. - Monster genomen "na expressie" voor SDS-pagina (stap 4.).
- Oogst de bacteriecellen door centrifugeren voor 10 min op 5.000 x g- en 4 ° C.
- Verwijder het supernatant door decanteren en bevriezen van de cel pellets bij-20 ° C of -80 ° C tot doel eiwitreiniging.
- Inoculeer 10 mL verse LB medium (tabel 1; aangevuld met 10 g/L glucose, 100 µg/mL ampicilline) met 100 µL van de overnachting cultuur in een 100 mL conische kolf. Incubeer de kolf bij 37 ° C in een roteerschudapparaat bij 200 omwentelingen per minuut.
-
SPI voor de productie van GdFP
- Inoculeer 10 mL van NMM19 medium (tabel 1) aangevuld met 100 µg/mL ampicilline, 15 µM tryptofaan en 10 µL van overnachting cultuur in een 100 mL conische kolf en de cultuur kolf 's nachts bij 30 ° C in een roteerschudapparaat op 200 rpm uit te broeden.
Opmerking: Een verscheidenheid van chemisch gedefinieerde media voor teelt van E. coli en SPI is beschikbaar. Naast NMM gebruikt hierin, MOPS middellange28, glucose-minerale zouten middellange29, Davis minimaal middellange30, M9 minimaal middellange31, of GMML32 kan worden gebruikt. - De volgende dag, meten OD600 elke 30 minuten, totdat de waarde alleen door minder dan 0,05 meer dan 30 min verandert. De waarde van het plateau moet ongeveer 1.
Opmerking: Afwijkingen door ± 0,3 eenheden zijn aanvaardbaar. Afhankelijk van de bacteriële stam en medium gebruikt, kan de concentratie van de oorspronkelijke tryptofaan (stap. 2.3.1) aanpassing nodig. - Neemt monster "before inductie" voor SDS-pagina (stap 4.).
- Oogst de bacteriecellen door centrifugeren voor 10 min op 5.000 x g- en 4 ° C. Verwijder het supernatant door decanteren.
- Resuspendeer de cellen in 10 mL NMM19 medium met 100 µg/mL ampicilline in een 100 mL conische kolf en 4-amino-indole toevoegen om een eindconcentratie van 1 mM met behulp van de stockoplossing van 50 mM. Blijven de incubatie gedurende 30 minuten bij 30 ° C in een roteerschudapparaat bij 200 omwentelingen per minuut.
Opmerking: Deze stap wordt aanbevolen vanwege de lage chemische stabiliteit van ampicilline en zorgt voor een cellulaire opname van 4-amino-indool. - Doel eiwituitdrukking veroorzaken door toevoeging van IPTG aan een eindconcentratie van 0.5 mM 1 M papier en Incubeer het monster 's nachts bij 30 ° C in een roteerschudapparaat bij 200 omwentelingen per minuut.
Opmerking: Cyaan fluorescente proteïnen zijn gewoonlijk uitgedrukt bij temperaturen onder de 37 ° C27. - De volgende dag, meten OD600.
- Monster genomen "na expressie" voor SDS-pagina (stap 4.).
- Oogst de bacteriecellen door centrifugeren voor 10 min op 5.000 x g- en 4 ° C en weggeworpen door decanteren.
- In het geval dat een dergelijk vaartuig niet gebruikt werd voor centrifugeren, breng de pellet cel in een 50 mL conische polystyreen buis met behulp van een spatel. De cel pellet bij-20 ° C of -80 ° C tot doel eiwitreiniging bevriezen.
- Inoculeer 10 mL van NMM19 medium (tabel 1) aangevuld met 100 µg/mL ampicilline, 15 µM tryptofaan en 10 µL van overnachting cultuur in een 100 mL conische kolf en de cultuur kolf 's nachts bij 30 ° C in een roteerschudapparaat op 200 rpm uit te broeden.
3. target eiwitreiniging via geïmmobiliseerdet metaalion affiniteitchromatografie (IMAC)
-
Lysis van de bacteriële cel
- Ontdooi de cel pellet op ijs voor 10-20 min.
- Resuspendeer de pellet cel in een 50 mL conische polystyreen buis met behulp van 5 mL ijskoud bindende buffer (tabel 1) op het ijs.
- Voeg 20 µL van 50 mg/mL lysozym, 20 µL van 1 mg/mL DNase ik 20 µL van 1 mg/mL RNase A. Sluit de buis, meng zachtjes door 5 keer omkeren, en houden op ijs gedurende 30 minuten.
Opmerking: Gedeeltelijke cel onderbreking optreedt als gekatalyseerd door lysozym. - Lyse de cellen door sonification met behulp van een echografie homogenizer tip met behulp van drie cycli van 3 min in een polystyreen tube van 15 mL afgekoeld door slush ijs met 2 s voor pulse, 4 s van pauze en 45% amplitude.
Opmerking: U kunt ook hogedruk homogenisering kan worden gebruikt, bijv., 20 cycli bij 14.000 psi. Eventueel verdunnen met behulp van bindende buffer te bereiken van het volume van de minimale instrument. Eiwit extractievloeistoffen kunnen bovendien worden gebruikt voor de verstoring van de cel. Zie materialen tabel voor voorbeelden. - Centrifugeer het monster gedurende 30 minuten bij 15.000-18.000 x g, 4 ° C.
- Breng de bovendrijvende substantie in een verse buis en noteer het vloeibare volume.
- Filtreer de oplossing door een 0,45 µm spuit-filter door middel van een 5 mL kunststof Luer lock spuit en een Polyvinylideenfluoride (PVDF) syringe filter.
- Neemt monster "lysate" voor SDS-pagina (stap 4.).
- Resuspendeer cel puin pellet in ddH2O (gelijk volume als voormalige lysate).
- Nemen monster "pellet" voor SDS-pagina (stap 4.).
-
IMAC zuivering
- Gebruik een 1 mL voorverpakt of zelf Lunchpakket IMAC FPLC (snelle proteïne-vloeistofchromatografie) kolom volgens de instructies van de fabrikant. Bindende buffer (tabel 1) voor kolom evenwichtsinstelling alsook wat betreft de wassen stap die volgt nadat de cel lysate is toegepast op de kolom gebruiken.
- Verzamelen en bundelen eluaat breuken met GdFP die kunnen worden geïdentificeerd door zichtbaar licht gouden kleur.
Opmerking: Optioneel, de doel-eiwit kan worden geëlueerd met behulp van een lineaire imidazool verloop (0-250 mM) met behulp van een geautomatiseerd FPLC systeem. - Bepaal de concentratie van de eiwitten met behulp van de literatuur waarde uitsterven-coëfficiënt op 466 nm (ɛ466 nm = 23.700 M-1 cm-1)33 met elutie buffer als referentie. Raadpleeg voor meer informatie over de procedure, eerdere publicatie26.
- Neem monster "eluaat" voor SDS-pagina, en gebruik van 1-10 µg eiwit per rijstrook in geval van Coomassie-kleuring.
Opmerking: SDS steekproef bedragen kunnen variëren naargelang de kleuringstechniek en kleurstof gevoeligheid. - Dialyze een aliquoot gedeelte van het eluaat breuken tegen dialyse buffer of MS buffer met behulp van een membraan met een molecuulgewicht cutoff (MWCO) van 5.000-10.000. Bereiden dialyse membraan volgens de instructies van de fabrikant. Dialyze een 1 mL monster driemaal tegen 100 mL buffer voor ten minste 2 uur. Raadpleeg voor meer informatie over deze procedure, eerdere publicatie34.
- Voor opslag, bevriezen eiwitSteekproef in dialyse buffer bij-80 ° C.
Opmerking: Aliquots moet stabiel gedurende ten minste 6 maanden.
4. SDS-pagina de bereiding van de monsters van E. coli hele cel Extract
- Overdracht van een gelijkwaardig aan 1 OD600 mL celsuspensie = 1 afhanging (bv. 500 µL van OD600= 2) in een tube van 1,5 mL microcentrifuge.
- Het oogsten van de cellen door gedurende 10 minuten bij kamertemperatuur, 5000 x g centrifugeren. Verwijder het supernatant door pipetteren.
- Voeg 80 µL van ddH2O en 20 µL van 5 x SDS laden kleurstof buffer (tabel 1) naar de cel pellet en meng door pipetteren.
- Het denatureren van de cellen door verhitting tot 95 ° C gedurende 5 minuten in een water-bad of warmte-blok. Vervolgens koelen de monsters tot op kamertemperatuur.
- Gebruik 10 µL voor SDS-pagina Coomassie gebeitste volgens eerdere publicatie35.
Opmerking: SDS steekproef bedragen kunnen variëren naargelang de kleuringstechniek en kleurstof gevoeligheid.
5. intact eiwit massa analyse door krachtige vloeibare chromatografie (HPLC) gekoppeld aan Electrospray ionisatie Time-of-flight massaspectrometrie (LC-ESI-TOF-MS)
Opmerking: HPLC verloop, instellingen en buffers kunnen variëren afhankelijk van de scheiding tussen kolom en instrument gebruikt. Zie materialen tabel voor voorbeeldige apparatuur.
- Bepalen de eiwitconcentratie uit een monster dialyzed tegen MS buffer zoals eerder beschreven (stap 3.2.3.) met MS buffer (Zie materialen tabel) als referentie.
- Verdun de eiwitSteekproef tot 0,1 mg/mL met behulp van MS buffer voor een eindvolume van 80 µL, Meng door zorgvuldige pipetteren, breng de oplossing kwantitatief over in een MS autosampler flacon met inzetglas en sluit deze met een cap. Het verwijderen van luchtbellen door flicking de flacon.
- Vul een tweede autosampler flesje zonder Glasfront (buffer leeg) met 1 mL van MS buffer.
- Het toestaan van het instrument om op te warmen. Kalibreren van het instrument uit te voeren. Ervoor te zorgen dat voldoende hoeveelheid vloeibare chromatografie-grade oplosmiddelen beschikbaar zijn (> 100 mL).
- Een lineaire 20 min HPLC verloop van 5% tot 80% buffer een (0,1% mierenzuur in ddH2O), in combinatie met buffer B Program (0,1% mierenzuur in acetonitril).
- HPLC beginnen bij een stroom van 0.3 mL/min en wacht totdat de kolom druk stabiel is.
- Instellen van een autosampler geïnjecteerde volume van 5 µL voor de LC-ESI-TOF-MS methode, een composities maken voor een leeg draaien gevolgd door een monster run en toewijzen van de overeenkomstige autosampler flacon posities. De composities worden uitgevoerd.
- Na voltooiing van de composities, open het gegenereerde gegevens voorbeeldbestand. Een celbereik selecteren in het huidige perceel (TIC) van de totale ion voor deconvolutie en deconvolute van het spectrum van de MS met behulp van de maximale entropie deconvolutie algoritme.
Opmerking: Afhankelijk van de experimentele omstandigheden, andere soorten kan optreden van niet-gerijpt FP of buffer ion adducten.
6. fluorescentie levensduur metingen en verval-geassocieerde Spectra (DAS) van GdFP
Opmerking: Voor de instrumentatie van time-resolved Fluorescentiespectroscopie, Raadpleeg Tabel van materialen voor voorbeeldige apparatuur. Extinctie, alsmede fluorescentie excitatie en emissie spectra van fluorescente proteïnen kan ook worden opgenomen met behulp van laboratorium UV/Vis- en fluorescentie spectrofotometers.
-
Golflengte-resolved fluorescentie levensduur meting van GdFP
- Bereiden van 2 mL van een 1 µM oplossing van GdFP door verdunning in PBS buffer (tabel 1) met een pH van 7. Vul de oplossing in een 1 cm kwarts cuvette.
- Ps-gepulseerde 470 nm laser voor monster excitatie en de 488 nm emissie long pass filter installeren en aanpassen van 600 L/mm raspend van het tijd - en golflengte-gecorreleerde één foton tellen36 (TWCSPC) detector voor verwerving van het regime van de golflengte 500-700 nm.
- Verwerven fluorescentie emissie met een snelheid van de graaf van ongeveer 200 x 103 fotonen/s tot ongeveer 103 punten worden verzameld in het maximum van de overname van de fluorescentie decay bochten met één foton tellen software.
-
Meting van de instrumentale reactie functie37 (IRF)
- Vervang de monster Cuvet met een 1 cm kwarts cuvette gevuld met 1 g/L Coloïdale siliciumdioxide (/g ~ 220 m2) in PBS buffer met een pH van 7.
Opmerking: De silica schorsing is opgesteld op basis van een waterige suspensie van 400 g/L. - 488 nm emissie long pass filter en invoegen grijs filters aan te passen het tempo van de detector van de TWCSPC tot onder 100 x 103 graven/s met graaf verwijderen.
- Aangepast rooster voor de verwerving van 470 nm fotonen in kanaal 8 van de 16-kanaals TWCSPC detector.
- Verwerven de IRF tot ongeveer 10 x 10,3 punten worden verzameld in de maximale emissie.
- Convert fluorescentie decay krommen en IRF naar ASCII-gegevensbestanden met global passend38 programma.
- Gedrag Global passen volgens een model van een som van drie exponentiële componenten met levensduur als gekoppelde parameters.
- Perceel verval gekoppeld spectra (DAS) als amplitude distributies van verval van de afzonderlijke componenten in afhankelijkheid van de golflengte software van de analyse van de gegevens.
- Vervang de monster Cuvet met een 1 cm kwarts cuvette gevuld met 1 g/L Coloïdale siliciumdioxide (/g ~ 220 m2) in PBS buffer met een pH van 7.
Representative Results
Met behulp van de techniek van de opneming van de selectieve druk, kan Trp-66 in de trits chromofore van ECFP (en Trp-57, het alleen andere Trp residu in ECFP) worden vervangen door 4-amino-Trp, waardoor het genereren van de rood-verschoven GdFP met verschillende spectrale eigenschappen. Massaspectrometrie moet worden gebruikt om aan te tonen van de gewenste stoichiometrische integratie van de niet-canonieke aminozuur in het eiwit, met resultaten die worden weergegeven in Figuur 1. Daarna bieden we gegevens uit microscopie, UV-Vis Absorptie spectroscopie evenals steady-state en tijd - en golflengte-resolved Fluorescentiespectroscopie te karakteriseren van de eigenschappen van de GdFP fluorophore met een focus op de afhankelijkheid van de pH van de Spectra.
Om te bevestigen de uitwisseling van de twee Trp residuen in ECFP door 4-amino-Trp, wordt massaspectrometrische analyse uitgevoerd. Figuur 1 toont een representatieve deconvoluted ESI-MS spectrum van GdFP. Hoewel wild-type ECFP een massa berekende eiwit voor 28,283.9 Da na de rijping chromofore heeft, is de overeenkomstige massa van GdFP 28,313.9 Da. Het deconvoluted ESI-MS spectrum van GdFP vertoont een grote massa piek bij 28,314.1 ± 0,1 Da, die van de theoretische waarde door minder dan 10 ppm afwijkt. Wordt binnen het bereik van de gemiddelde nauwkeurigheid voor dit soort analyse25, dit bevestigt de opneming van de ncAA via SPI (experimentele waarde voor wild-type ECFP: 28,283.7 Da).
Figuur 2 toont confocal fluorescentie imaging microscopie (CFIM) beelden van bacteriële cellen uiten van EGFP, EYFP, ECFP en GdFP op resuspensie van bacteriën in PBS buffer. Alle beelden werden verworven op een microscoop voorzien van een UV objectieve en laser excitatie bij over dezelfde energie voor elk monster.
Figuur 3A geeft een overlay van CFIM beelden van E. coli bacteriën uiting van verschillende FPs met inbegrip van GdFP, altijd gecontroleerd met zeer vergelijkbaar excitatie energie (golflengten zoals in Figuur 2). Figuur 3B toont de chromofore structuren van de varianten van de FP komt te staan. Met betrekking tot de helderheid van de GdFP in vergelijking met ECFP (fluorescentie quantum opbrengst φ = 0,4), EGFP (φ = 0,6) en EYFP (φ = 0.6) het is belangrijk op te merken dat voor GdFP, een breder scala van de overname van de fluorescentie-licht (30 nm) werd gebruikt in tegenstelling tot 20 nm gebruikt voor alle andere spe cies, om aan te passen van de intensiteit van de afbeeldingen om gelijkaardige waarden. Met een iets lagere uitsterven coëfficiënt en een verminderde quantumrendement als gevolg van de unieke fotofysische eigenschappen, is de helderheid van de GdFP lager in vergelijking met de andere FPs komt te staan.
Het absorptiespectrum van ECFP (Figuur 3 c) heeft twee karakteristieke maxima op 434 nm en 452 nm. In tegenstelling, GdFP wordt gekenmerkt door een brede absorptie van rood-verschoven band met het maximum op 466 nm. De absorptie van EGFP is verder rood-verschoven naar 488 nm. Echter, als gevolg van de veel grotere Stokes-verschuiving van de GdFP (108 nm) in vergelijking met ECFP (41 nm) en EGFP (20 nm), het emissiespectrum van GdFP is het meest rode-verschoven van alle drie GFP derivaten hier onderzocht (figuur 3D). Terwijl de uitstoot van de fluorescentie van ECFP twee karakteristieke maxima op 475 toont nm en 505 nm, EGFP heeft een brede belangrijkste emissie band piek op 508 nm (λmax) met een lichte schouder bij 540 nm. De fluorescentie van GdFP blijkt bij ongeveer 565 nm (λmax.) (Afbeelding 3D). Het emissiespectrum bevat een kleine bijdrage te leveren van wild-type ECFP dat ook zichtbaar is als een kleine schouder naar 475 nm. Deze kleine ECFP-fractie wordt gesynthetiseerd voordat inductie tijdens de SPI-procedure, als beschreven33.
De 3E figuur toont de pH-afhankelijke veranderingen in het spectrum van de absorptie van GdFP. Voor een pH verandering van 8 tot en met 5, de maximale emissie verschuift iets naar de rode en een lichte verruiming van de absorptie-band wordt waargenomen. De vermindering van de amplitude van de absorptie is echter slechts ongeveer 10% tussen pH 8 en pH 5, waaruit blijkt dat de eigenschappen van de grondtoestand van het GdFP chromofore zeer zwak gewijzigd door pH.
De tijd opgelost fluorescentie emissie gecontroleerd door één foton tellen wordt weergegeven in Figuur 4. De curven van de verval gecontroleerd in de spectrale kanalen gecentreerd op 550 nm en 600 nm (figuur 4A) vertonen een iets snellere verval van de fluorescentie op 600 nm ten opzichte van het verval op 550 nm. De resultaten van een wereldwijde pasvorm van de fluorescentie decay krommen met twee exponentiële onderdelen resultaten in twee spectraal te onderscheiden fluorescentie decay componenten met tijd constanten van 1.0 ns en 3.3 ns (figuur 4C en D).
De uitstoot van de fluorescentie van GdFP afhankelijk sterk van pH, zoals het is typisch voor veel fluorescerende eiwit varianten van het GFP-familie. Figuur 4B vergelijkt de uitstoot van de fluorescentie van GdFP tussen pH 5 en pH 8, waaruit duidelijk een daling van de intensiteit van de fluorescentie bij een lagere pH, blijkt terwijl de spectrumkenmerken constant blijven.
De verval-geassocieerde spectra (DAS)39 van GdFP (figuur 4C en D) worden gekenmerkt door twee verschillende emissie bands. De bijdrage van de langzame 3.3 ns component is meer uitgesproken in het korte golflengtegebied rond 550 nm (60%) met kleine bijdrage van de snellere component (40%). Op 600 nm, beide componenten hebben over de dezelfde amplitude. Op een verschuiving van de pH 7 (figuur 4C) op een pH van 6 (Figuur 4 d), de spectrale kenmerken van de DAS nauwelijks veranderen en de tijd-constanten van de wereldwijde montage routine zijn ook hetzelfde (de nauwkeurigheid van de DAS tijd constanten is ongeveer ± 0,15 ns). Echter, het verschil in de absolute amplitudes van de twee componenten van de DAS is duidelijk herkenbaar, die volledig verantwoordelijk voor de amplitude van de uitstoot verlaagd fluorescentie op de dezelfde verschuiving van de pH in figuur 4B.
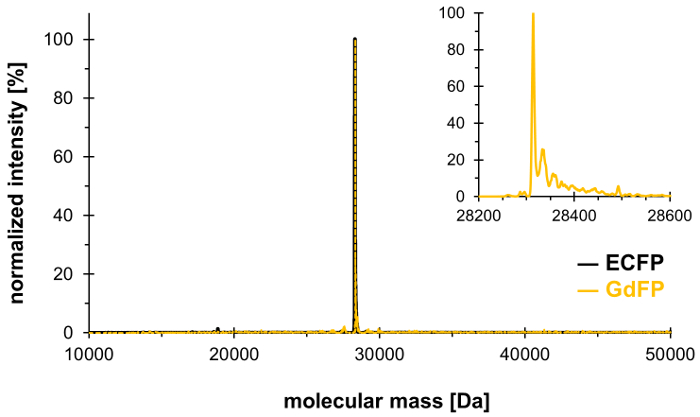
Figuur 1: vertegenwoordiger deconvoluted ESI-MS spectrum van GdFP. Het spectrum van de ESI-MS van GdFP (gouden kleur, vergrote perceel getoond als inzet) toont een belangrijkste piek bij 28314.1 Da (berekend van de waarde 28313.9 Da). Het spectrum voor wild-type ECFP wordt weergegeven in het zwart. Klik hier voor een grotere versie van dit cijfer.

Figuur 2: confocale fluorescentie microscopie beelden van bacteriële bevolking uiting van verschillende FOD. De volgende instellingen: golflengte werden gebruikt voor Beeldacquisitie: ECFP (λex = 457 nm, detectie: 461-480 nm), EGFP (λex = 488 nm, detectie: 495-515 nm), GDFP (λex = 476 nm, detectie: 560-590 nm), EYFP (λex = 514 nm, detectie: 520-530 nm). Klik hier voor een grotere versie van dit cijfer.

Figuur 3: spectrale eigenschappen van GdFP. (A) CFIM afbeelding van een mengsel van bacteriële cellen uiten van EGFP, ECFP en GdFP na de resuspensie van bacteriën in PBS buffer. (B) chromofoor structuren van GdFP (met 4-amino-Trp in plaats van residu 66), de ouderlijke ECFP (met Trp op positie 66) en EFGP (met Tyr op positie 66). (C) vergelijking voor de genormaliseerde absorptiespectra van GdFP, ECFP en EGFP, overwegende dat in (D), de genormaliseerde fluorescentie emissiespectrum van ECFP (excitatie op 430 nm) is in vergelijking met de fluorescentie emissie spectra van EGFP en GdFP (zowel enthousiast op 450 nm). (E) pH-afhankelijkheid van de Absorptiespectra (genormaliseerd absorptie bij 280 nm). Klik hier voor een grotere versie van dit cijfer.

Figuur 4: Time-resolved fluorescentie van GdFP. (A) fluorescentie verval van GdFP gevolgd door tijd - en golflengte-opgelost één foton tellen in de spectrale kanalen gecentreerd op 550 nm en 600 nm (± 12,5 nm) na excitatie met 470 nm laserpulsen. De instrumentale reactie-functie (IRF) bevat informatie over de resolutie van de tijd van de gebruikte setup. (B) variant van het emissiespectrum van GdFP afhankelijk van de pH (excitatie op 460 nm). (C, D) Verval-geassocieerde spectra (DAS) van GdFP bij pH 7 (C) en pH 6 (D) bepaald nadat deconvolutie van tijd - en golflengte-resolved fluorescentie vervalt en globale montage van de vervalt in alle kanalen door een algemene set van twee exponentiële functies met gekoppelde tijd-constanten zijn. Klik hier voor een grotere versie van dit cijfer.

Figuur 5: structuren van de intramoleculaire gratis overdracht van ECFP (zwart) en GdFP (goud)-chromophores. De toename van de omvang van het systeem van de chromofore door de goede elektronendonor van een amino groep als onderdeel van de ncAA kan de vorming van meer mesomeric structuren tot stabilisatie van de resonantie van de geëxciteerde toestand. De verbindingspunten aan de steiger van de FP worden weergegeven als semicircles. Klik hier voor een grotere versie van dit cijfer.
| Stockoplossing | concentratie, oplosmiddel | Opmerking | |
| 20% D-glucose | 200 g/L D-glucose in ddH2O | steriliseren door filtratie door een 0,45 µm porie grootte spuit filter | |
| indool | 50 mM in isopropanol | ||
| 4-amino-indole | 50 mM in 20% ethanol (20 mL ethanol in een eindvolume van 100 mL gevuld met ddH2O) | ||
| IPTG | 1 M in ddH2O | ||
| L-tryptofaan | 15 mM opgelost in ddH2O met behulp van 1 M HCl (toevoegen van HCl ontkleuring onder roeren tot poeder dissoved is) | ||
| lysozym | 50 mg/mL in ddH2O | ||
| DNase ik | 1 mg/mL in ddH2O | ||
| RNase A | 1 mg/mL in ddH2O | ||
| Amp100 | 100 mg/mL ampicilline in ddH2O | ||
| natrium-dodecylsulfate (SDS) | 200 g/L in ddH2O | ||
| ammoniumsulfaat ((NH4)2SO4) | 1 M in ddH2O | steriliseren in autoclaaf | |
| kalium Natriumdiwaterstoffosfaat fosfaat (KH2PO4) | 1 M in ddH2O | steriliseren in autoclaaf | |
| di-kalium waterstof fosfaat (K2HPO4) | 1 M in ddH2O | steriliseren in autoclaaf | |
| magnesium-sulfaat (MgSO4) | 1 M in ddH2O | steriliseren in autoclaaf | |
| D-glucose | 1 M in ddH2O | steriliseren door filtratie door een 0,45 µm porie grootte spuit filter | |
| natriumchloride (NaCl) | 5 M in ddH2O | steriliseren in autoclaaf | |
| calciumchloride (CaCl2) | 1 g/L | steriliseren door filtratie door een 0,45 µm porie grootte spuit filter | |
| ijzer(II) chloride (FeCl2) | 1 g/L | steriliseren door filtratie door een 0,45 µm porie grootte spuit filter | |
| Thiamine | 10 g/L | steriliseren door filtratie door een 0,45 µm porie grootte spuit filter | |
| Biotine | 10 g/L | steriliseren door filtratie door een 0,45 µm porie grootte spuit filter | |
| mix van sporenelementen | kopersulfaat CuSO4, zink chloride (ZnCl2), mangaan chloride (MnCl2), ammoniummolybdaat ((NH4)2MoO4); elke 1 mg/L in ddH2O | steriliseren door filtratie door een 0,45 µm porie grootte spuit filter | |
| 19 aminozuren mix | 1.) Los 0,5 g L-fenylalanine en 0,5 g L-tyrosine in 100 ml ddH2O met dropwise toevoeging van 1 M HCl onder roeren tot poeder wordt ontbonden. | ||
| 2.) Weeg 0,5 g van elk van de resterende L-amino zuren (met uitzondering van L-tryptofaan). Meng met 22 mL fo 1 M KH2PO4 en 48 mL 1 M K2HPO4. DdH2O aan ongeveer 800 mL toevoegen. Roer de oplossing pas duidelijk. | |||
| 3.) toevoegen de opgeloste L-fenylalanine en L-tyrosine uit stap 1). en pas het volume aan 1 L met ddH2O. | |||
| 4.) Sterilize het aminozuur mengsel door vacuüm filtratie met een fles top filter eenheid. | |||
| Buffers en Media | Samenstelling/voorbereiding | ||
| SDS laden kleurstof buffer, 5 x geconcentreerd | 0,25 M Tris pH 6.8, 50% v/v glycerol, 0,25% w/v broomfenolblauw, 0.5 M didhiothreitol (DTT; u kunt ook β-mercaptoethanol 5%), 10% w/v natrium-dodecylsulfate (SDS) | ||
| bindende buffer | 50 mM natrium dihydrogenphosphate (NaH2PO4), 500 mM NaCl, 10 mM imidazool, pH 8 | ||
| elutie buffer | 50 mM natrium dihydrogenphosphate (NaH2PO4), 500 mM NaCl, 250 mM imidazool, pH 8 | ||
| dialyse buffer | 50 mM natrium dihydrogenphosphate (NaH2PO4), 150 mM NaCl, 100 mL/L glycerol, pH 8 | ||
| MS buffer | 10 mM Tris-HCl, pH 8 | ||
| nieuwe minimaal medium met 19 L-amino zuren met uitzondering van L-tryptofaan (NMM19) | Vermeng alle voorraad oplossingen om het verkrijgen van de volgende eindconcentraties: 7.5 mM (NH4)2SO4, 1.7 mM NaCl, 22 mM KH2PO4, 50 mM K2HPO4, 1 mM MgSO4, 20 mM D-glucose, 50 mg/L 19 aminozuren mix, 1 µg/L CaCl21 µg/L FeCl2, 10 µg/L thiamine, Biotine 10 mg/L, 0,01 mg/L sporenelementen mix | ||
| LB medium | Samenstelling: 10 g/L trypton, 5 g/L gistextract, 10 g/L NaCl, pH 7,0 in ddH2O | ||
| Bereiding: | |||
| 1.) Weeg uit trypton 50 g 25 g gistextract, 5 g NaCl in een 1 L-fles. | |||
| 2.) toevoegen ddH2O tot ~ 800 mL en los onderdelen onder roeren. | |||
| 3.) meten van pH en pas op een pH van 7 door dropwise toevoeging van 1 M HCl of 1 M NaOH, indien nodig. Toevoegen van ddH2O tot 1 L. | |||
| 4.) Sterilize in autoclaaf, Controleer volume verlies daarna en toevoegen van steriele ddH2O te compenseren indien nodig. Bewaren bij 4 ° C tot gebruik. | |||
| LB agar platen | Samenstelling: 10 g/L trypton, 5 g/L gistextract, 10 g/L NaCl, 15 g/L agar-agar, pH 7.0 in ddH2O | ||
| Bereiding: | |||
| 1.) Weeg uit trypton 50 g 25 g gistextract, 5 g NaCl, 7,5 g agar-agar in een 1 L-fles. | |||
| 2.) toevoegen ddH2O tot 500 mL en los componenten onder roeren. | |||
| 3.) meten van pH en pas op een pH van 7 door dropwise toevoeging van 1 M HCl of 1 M NaOH, indien nodig. Toevoegen van ddH2O tot 1 L. | |||
| 4.) Sterilize in autoclaaf, Controleer volume verlies daarna en toevoegen van steriele ddH2O te vergoeden, indien nodig. (Opmerking: LB agar kan worden achtergelaten bij 4 ° C tot gebruik voor bereiding van LB agar platen. Zorgvuldig smelten gestolde agar met behulp van een magnetron) | |||
| 5.) wanneer de oplossing is nog steeds warm (30-40 ° C), ampicilline toevoegen om een eindconcentratie van 100 µg/mL | |||
| 6.) pour ongeveer 15 mL van de vloeistof uit stap 5.) in een steriele 10 cm petrischaal onder steriele omstandigheden. Wanneer de agar is gestold, kunnen platen worden opgeslagen voor 1 week bij 4 ° C tot gebruik. | |||
| fosfaatgebufferde zoutoplossing (PBS) | Samenstelling: 137 mM NaCl, 2,7 mM KCl, 10 mM nb2HPO4met 1,8 mM KH2PO4, 1 mM CaCl2, 0.5 mM MgCl2, pH 7. Steriliseren in autoclaaf of filtratie. | ||
Tabel 1: Stock oplossing en buffer.
Discussion
Om te bereiken zeer hoge efficiëntie van de opneming van de ncAA, berust de SPI auxotrophy gebaseerde methode op het gebruik van metabolisch gemanipuleerde gastheer cellen, die zijn niet in staat voor het synthetiseren van de bijbehorende natuurlijke tegenhanger van de ncAA. Voor E. colizijn dergelijke stammen beschikbaar. Zelfs de gelijktijdige integratie van meerdere ncAAs in hetzelfde eiwit is haalbaar met behulp van de multiauxotrophic stammen. De residu-specifieke wijze van vervanging en de chemische repertoire wordt beperkt tot soortgelijke chemische analogen kan worden gezien als nadelen. Een groot aantal eiwit varianten kan echter worden geproduceerd zoals het natuurlijke bacteriële vertaling apparaat talrijke aminozuur-analogen tolereert. Bijvoorbeeld, kan meer dan 50 ncAAs in eiwitten met behulp van in vitro vertaling, ongeveer 73% van alle codonen van de genetische code beschikbaar te zijn voor hertoewijzing40worden opgenomen. SPI kunnen bovendien ook efficiënte multisite etikettering van de target eiwit41. In principe de SPI-methodologie is niet beperkt tot E. coli, maar kan werken in een willekeurige andere host en voor elk van de canonieke 20 aminozuren, mits auxotrofe stammen en gedefinieerde teelt media beschikbaar zijn. Bijvoorbeeld, zijn twee methionine analogen, azidohomoalanine (Aha) en homopropargylglycine (Hpg), commercieel beschikbaar en worden gebruikt voor het labelen van proteïnen en proteomes in verschillende organismen. Aha kan bovendien worden geproduceerd intracellulair en vervolgens opgenomen in eiwit42. Deze ncAA is vooral geschikt voor bioorthogonal vervoegingen zoals Klik chemie zoals ontwikkeld door Tirrel en collega's: plant bijvoorbeeld in weefsel van Arabidopsis thaliana, Bombyx mori larven43, Drosophila cellen44, larvale zebrafish45 evenals zoogdiercellen met inbegrip van neuronen46, eiwitten kunnen worden voorzien van Aha47,48. Trp-analogen zijn ook met succes opgenomen in antimicrobiële peptiden in Trp-auxotrofe Lactococcus lactis stammen49. SPI is ook handig voor het gebied van Xenobiology50,51, die onderzoekt de alternatieven voor de fundamentele chemische samenstelling van het leven. Bijvoorbeeld, werd gebaseerd op vorige kunstwerken op E. coli52 en B. subtilis53, een stam E. coli ontwikkeld onlangs door een evolutionaire strategie met selectieve druk om te gebruiken in plaats van thienopyrrole indool, resulterend in Proteoom bestrijkende vervanging van tryptofaan door thienopyrrole-alanine in de genetische code54. In het algemeen, het canonieke aminozuur Trp, die is gecodeerd door een enkele triplet (UGG), presenteert een veelbelovende doelwit voor Eiwittechnologie toe te schrijven aan de rijke facetten van indool chemie, die talrijke chemische variaties biedt. Onlangs, en als een alternatief voor SPI gebaseerde integratie, een roman die SCS platform staat te nemen Trp-analogen site-specifically in zowel de bacteriële en de eukaryote gastheren is gemeld55. Dit verbreedt verder de werkset van in vivo ncAA gebaseerde Eiwittechnologie, met inbegrip van de wijziging van de spectrale eigenschappen.
Naast het gebruik van auxotrofe expressie hosts vereist het SPI-protocol strikte gisting voorwaarden, zowel in termen van doel expressie timing en de samenstelling van het medium om ncAA opneming hoogrenderende en doel eiwit opbrengst te bereiken 56. teelt wordt uitgevoerd met behulp van chemisch gedefinieerde minimale media, die in wezen bevatten naast grote zouten van de bronnen van stikstof (ammoniumzout) en koolstof (D-glucose), vitaminen en sporenelementen. Hoewel niet strikt vereist in het ontbreken van verdere auxotrophies, de resterende aminozuren (20 -n, als n aminozuren worden vervangen) worden vaak toegevoegd ter bevordering van de bacteriegroei57. Tijdens een eerste groeifase voor inductie van doel eiwit expressie, worden de n -canonieke aminozuren worden vervangen toegevoegd in het beperken van de concentraties. Cellulaire groei gaat door tot de gerichte essentiële aminozuren zijn uitgeput, zo experimenteel aangegeven door een stationaire OD600. Vervolgens wordt het kweekmedium vervangen door verse medium dat ontbreekt het verarmd aminozuur en bevat de ncAA in overvloedige concentraties. Voor de ribosomal opname van tryptofaan-analogen zoals in dit protocol, wordt een analoge indool gevoed, die intracellulair wordt geconverteerd naar de corresponderende tryptofaan afgeleide door tryptofaan synthase58. Vervolgens wordt doel eiwit expressie veroorzaakt. In dit stadium zijn de cellen dicht bij het einde van logaritmische groei, zoals een evenwicht tussen totale celaantal en fitness. Zoals de aanwezigheid en de opneming van de canonieke amino tot wild-type eiwitproductie leiden zou, is het essentieel om ervoor te zorgen dat het essentiële aminozuur volledig vóór inductie uitgeput is. Ook is het verplicht om de onderzoeken van de efficiëntie van ncAA opneming in het doel-eiwit, vaak door massaspectrometrie. In geval van substantiële aanwezigheid van het canonieke aminozuur, de teelt voorwaarden moeten worden aangepast, bijvoorbeelddoor een wijziging van de concentratie van de essentiële amino acid(s) voor de initiële groeifase of de duur van de laatste. In het geval van de activiteit van de lage jaarlijkse activiteitenverslagen naar de ncAA kunnen de overexpressie van het endogene enzym of co uitdrukking van een verschillende jaarlijkse activiteitenverslagen, oftewel actiever naar de ncAA, transiënte59.
Het canonieke aminozuur Trp is begiftigd met drie opmerkelijke functies: (i) de natuurlijke overvloed in eiwitten is laag; (ii) de biofysische en chemische eigenschappen uniek zijn (bv., het is meestal de dominante oorsprong van de intrinsieke fluorescentie van eiwitten en peptiden), en (iii) het draagt bij aan een verscheidenheid van biochemische interacties en functies met inbegrip van Π-stapelen, H-binding en catie-π interacties. Al deze functies zijn radicaal veranderd op Trp → 4-amino-Trp vervanging GdFP. Beyond twijfel, het ontwerp van een "gouden" klasse van avGFPs is een opmerkelijk voorbeeld voor engineering op maat autofluorescent eiwitten. Met verschillende spectrale eigenschappen, kan de FPs naar bepaalde spectrale windows via mutagenese en ncAA opneming worden afgestemd. In het geval van GdFP, wordt dit bereikt door een eenvoudige chemische uitwisseling H → NH2 in het kader van de ring van de indool opgenomen in de ECFP chromofore triade treft. Figuur 5 toont de effecten van ncAA integratie binnen de chromofore. De invoering van de elektron-doneren groep afkomstig uit 4-amino-indool (intracellulair geconverteerd naar 4-amino-Trp) kan een verscheidenheid van mesomeric structuren die een gestabiliseerde geëxciteerde toestand verklaren kunnen. Spectroscopically, haar uitgebreide Stokes shift en rood-verschoven fluorescentie uitstoot het gevolg zijn van deze verschillende eigenschappen van het uitgebreide geconjugeerd systeem. Zoals gemeld eerder, de verbeterde intramoleculaire gratis overdracht binnen de GdFP chromofore is inherent gevoelig voor pH (figuur 4B) en vergezeld gaat van een grotere verandering in dipoolmoment tussen de S0 grond en S1 opgewonden staat ten opzichte van ECFP33. Als alternatieve elektron-doneren groepen, kunnen tryptofaan analogen rekening houdend met een indool-ring vervangen door hydroxy groepen worden gebruikt, zoals gerapporteerd in een vergelijkende studie met de model eiwit barstar41.
De absorptie en fluorescentie spectra van GdFP worden uitgebreid ten opzichte van ECFP en EGFP (Figuur 3 c en D). Homogene verbreding van de absorptie en fluorescentie bands is over het algemeen veroorzaakt door vibrationele modi in de chromofoor en, bovendien, koppeling van de chromofore aan verdere vibrationele modi aanwezig in de eiwit-60. De koppeling aan de lokale eiwit-omgeving wordt ondersteund door kosten gelokaliseerd op de chromofore. Omdat de structurele heterogeniteit van het eiwit leidt tot lokale variaties van het vibronic spectrum, dergelijke koppeling tussen de vibronic spectra van de chromofoor en de rest van het eiwit worden ondersteund door de verplaatsing van de lading en mesomeric staten zoals aangegeven in Figuur 5. Door deze koppeling ondersteunt ook de grote verschuiving van Stokes en noodzakelijkerwijs vermindert het quantumrendement van fluorescentie. In vergelijking met andere rood-verschoven FPs vertoont GdFP zelfs betere eiwitstabiliteit en een lage tendens voor aggregatie33,61,62. Het is niet alleen verschilt in kleur van andere FP-varianten, maar vertoont ook een aanzienlijk hogere thermostabiliteit en verbeterde coöperatie33vouwen. De intensiteit van de fluorescentie is ten minste 90% behouden bij verhitting tot 60 ° C, terwijl ECFP fluorescentie is teruggebracht tot ongeveer 30%. In eiwitten bijdragen aromatische aminozuren vaak aan netwerken van interagerende zijketens, die vaak een stabiliserende invloed op de tertiaire structuur van het eiwit hebben. avGFP herbergt zo'n kant keten netwerk, dat uit de chromofore zelf, als goed als Phe-165, zijne-148 en Tyr-145 bestaat. Deze zijketens zijn niet alleen heel stijf in de GdFP structuur33, maar bovenal vormen ze hydrofobe contacten met de chromofore. De meest opvallende nieuwe kenmerk geïdentificeerd in GdFP is dat de aminated chromofore meer proximale tot Phe-165. Deze interactie is een functie niet waargenomen in andere bekende avGFPs. Als de twee residuen 3.2-4.5 Å apart zijn, kunnen amino-aromatische interacties ook aanwezig zijn. Samen met de aminering-geïnduceerde resonantie te stabiliseren de chromofore, deze waarschijnlijk deze hydrofobe netwerk stabiliseren van aminozuren in een coöperatieve manier. Een meer doeltreffende intramoleculaire gratis overdracht mogelijk ondersteund door deze interacties in de geëxciteerde toestand ten opzichte van de grondtoestand van het chromofoor en het op zijn minst gedeeltelijk verantwoordelijk voor de 108 nm Stokes shift33,62 .
In rationele ontwerp van fluorophore eigenschappen, is een toename van de omvang van de gedelokaliseerd π-systeem voorspelde resulteren in een rood-verschoven excitatie golflengte. Deze vuistregel wordt nageleefd door de reeks van aminozuren in positie 66 leidt tot neutraal chromophores: Phe (λmax = 355 nm) < zijn (λma x= 386 nm) < Tyr (λmax = 395 nm) < Trp (λmax = 436 nm)63. In de natuur, is de uitbreiding van de chromofore van geconjugeerd systeem van pi-bindingen bereikt door verschillende strategieën. Voor DsRed vanaf Discosoma striata, is zij uitgebreid door de integratie van een extra amino zuur, dus verschuiven λmax naar 573 nm64. De chromofore van asFP595 (λmax = 595 nm) van Anemonia sulcata werd uitgebreid door een imino groep, uitbreiding van de π-systeem-65. Aangezien de chromofore van GdFP en andere avFPs van dezelfde grootte, moet een ander beginsel inhouden een emissie golflengte in het bereik van de uitgebreide DsRed en asFP595 chromophores. De diepe Stokes verschuiving van 108 nm wordt toegeschreven aan de aparte structuur van de GdFP chromofore, die een nieuwe fotofysische beginsel in het ontwerp van autofluorescent eiwitten onthult. Voorlopige berekeningen (zoals gerapporteerd in 62) is gebleken dat het dipoolmoment van het opgewonden statuswaarden chromofore van GdFP aanzienlijk groter dan in de grondtoestand, in tegenstelling tot de respectievelijke waarden van ECFP. Overwegende dat het dipoolmoment van GdFP van ~ 3 D (Debye) in de S0 Braziliaanse tot ~ 15 D in S1 stijgt, de verandering voor de ECFP chromofore was vrij gematigd (van ~ 4 D tot ~ 6 D). Dus, het unieke gouden fluorescentie van GdFP wordt veroorzaakt door aanzienlijke intramoleculaire gratis overdracht binnen de chromofore, waardoor het scala aan mogelijke mesomeric structuren (Zie Figuur 5) die het mogelijk voor de stabilisatie van de resonantie maken. Dit vermindert het energieniveau waaruit emissie plaatsvindt. Als gevolg van de ingrijpende veranderingen in het dipoolmoment op excitatie is de intramoleculaire gratis scheiding de belangrijkste reden voor de wijzigingen in het elektrostatische potentieel van het chromofore milieu. De omliggende eiwit matrix, beurtelings, aangepast aan de veranderingen in de ladingsverdeling na chromofore excitatie. De daaropvolgende structurele versoepeling verlaagt het energieniveau van de opgewonden chromofore, die de fluorescentie-spectrum naar de rode vanwege haar gratis overdracht karakter verschuift. Voor dezelfde reden, als gevolg van de grote Stokes shift en de verbeterde tarieven van radiationless processen, de fluorescentie quantumrendement van GdFP wordt verminderd in vergelijking met ECFP33.
De hoge quantumrendement en kleine Stokes verschuiving van ECFP en EGFP zijn meestal toegeschreven aan een rigide eiwit-omgeving van de chromofore, waardoor het aantal vrijheidsgraden en, bijgevolg, interne conversie naar de gunst van de radiatieve versoepeling van de geëxciteerde toestand 66. bijgevolg de moleculaire design van meer rigide ingesloten chromophores met verminderde koppeling naar het resterende eiwit matrix kan dienen als een gids voor het produceren van verder rood-verschoven GFP derivaten met hoge fluorescentie quantumrendement. Daarom, voor verdere engineering benaderingen voor de productie van rode-verschoven autofluorescent eiwitten, uitbreiding van het systeem van de π-elektronen en een stijve chromofore structuur met zwak koppelen aan het eiwit milieu is zeer wenselijk. Dergelijke wijzigingen kunnen ook worden ingevoerd rechtstreeks in GFP gebaseerde chromophores of door plaatsing van de gewenste ncAAs in de omgeving van chromofore.
Disclosures
De auteurs verklaren dat zij geen concurrerende financiële belangen hebben.
Acknowledgments
Dit werk werd ondersteund door de Duitse Research Foundation (Cluster van Excellence "verenigen concepten in katalyse) T.F. en N.B. en door het federale ministerie van onderwijs en Wetenschappen (goedgekeurd programma"HSP 2020", TU-WIMIplus Project SynTUBio) naar F.-J.S.
Materials
| Name | Company | Catalog Number | Comments |
| Chemicals | |||
| 4-aminoindole | Sigma-Aldrich | 525022 | |
| acetonitrile | VWR | HiPerSolv CHROMANORM ULTRA for LC-MS, 83642 | LC-MS grade required |
| agar-agar | Carl Roth | 5210 | |
| ammonium molybdate ((NH4)2MoO4) | Sigma-Aldrich | 277908 | |
| ammonium sulfate ((NH4)2SO4) | Sigma-Aldrich | A4418 | |
| ampicillin sodium salt | Carl Roth | K029 | |
| biotin | Sigma-Aldrich | B4501 | |
| bromophenol blue | Sigma-Aldrich | B0126 | |
| calcium chloride (CaCl2) | Sigma-Aldrich | C5670 | |
| colloidal silica | Sigma-Aldrich | Ludox HS-40, 420816 | |
| Coomassie Brillant Blue R 250 | Carl Roth | 3862 | |
| copper sulfate (CuSO4) | Carl Roth | CP86.1 | |
| D-glucose | Carl Roth | 6780 | |
| di-sodium hydrogen phosphate (Na2HPO4) | Carl Roth | X987 | |
| di-potassium hydrogen phosphate (K2HPO4) | Carl Roth | P749.1 | |
| 1,4-dithiothreitol (DTT) | Carl Roth | 6908 | |
| DNase I | Sigma-Aldrich | D5025 | |
| ethanol | Carl Roth | 9065.1 | |
| formic acid | VWR | HiPerSolv CHROMANORM for LC-MS, 84865 | LC-MS grade required |
| glycerol | Carl Roth | 3783 | |
| imidazole | Carl Roth | X998 | |
| indole | Sigma-Aldrich | I3408 | |
| iron(II) chloride (FeCl2) | Sigma-Aldrich | 380024 | |
| isopropanol | Carl Roth | AE73.1 | |
| isopropyl β-D-1-thiogalactopyranoside (IPTG) | Sigma-Aldrich | I6758 | |
| lysozyme | Sigma-Aldrich | L6876 | |
| magnesium chloride (MgCl2) | Carl Roth | KK36.1 | |
| magnesium sulfate (MgSO4) | Carl Roth | 8283.2 | |
| manganese chloride (MnCl2) | Sigma-Aldrich | 63535 | |
| β-mercaptoethanol | Carl Roth | 4227.3 | |
| potassium chloride (KCl) | Carl Roth | 6781.3 | |
| potassium dihydrogen phosphate (KH2PO4) | Sigma-Aldrich | P5655 | |
| RNase A | Carl Roth | 7156 | |
| sodium chloride (NaCl) | Carl Roth | P029 | |
| sodium dihydrogen phosphate (NaH2PO4) | Carl Roth | T879 | |
| sodium dodecyl sulphate (NaC12H25SO4) | Carl Roth | 0183 | |
| thiamine | Sigma-Aldrich | T4625 | |
| Tris(hydroxymethyl)-aminomethane (Tris) | Carl Roth | 5429 | |
| Tris hydrochloride (Tris-HCl) | Sigma-Aldrich | 857645 | |
| tryptone | Carl Roth | 8952 | |
| yeast extract | Carl Roth | 2363 | |
| zinc chloride (ZnCl2) | Sigma-Aldrich | 229997 | |
| Name | Company | Catalog Number | Comments |
| amino acids | |||
| L-alanine | Sigma-Aldrich | A7627 | |
| L-arginine | Sigma-Aldrich | A5006 | |
| L-asparagine | Sigma-Aldrich | A8381 | |
| L-aspartic acid | Sigma-Aldrich | A0884 | |
| L-cysteine | Sigma-Aldrich | C7352 | |
| L-glutamic acid | Sigma-Aldrich | G2128 | |
| L-glutamine | Sigma-Aldrich | G3126 | |
| L-glycine | Sigma-Aldrich | G7126 | |
| L-histidine | Sigma-Aldrich | H8000 | |
| L-isoleucine | Sigma-Aldrich | I2752 | |
| L-leucine | Sigma-Aldrich | L8000 | |
| L-lysine | Sigma-Aldrich | L5501 | |
| L-methionine | Sigma-Aldrich | M9625 | |
| L-proline | Sigma-Aldrich | P0380 | |
| L-phenylalanine | Sigma-Aldrich | P2126 | |
| L-serine | Sigma-Aldrich | S4500 | |
| L-threonine | Sigma-Aldrich | T8625 | |
| L-tryptophan | Sigma-Aldrich | T0254 | |
| L-tyrosine | Sigma-Aldrich | T3754 | |
| L-valine | Sigma-Aldrich | V0500 | |
| Name | Company | Catalog Number | Comments |
| Lab materials | |||
| 0.45 µm syringe filter with PVDF membrane | Carl Roth | CCY1.1 | |
| 1.5 mL microcentrifuge tubes | Eppendorf | 30120086 | |
| conical polystyrene (Falcon) tubes, 50 mL | Fisher Scientific | 14-432-22 | |
| Luer-Lock syringe 5 mL | Carl Roth | EP96.1 | |
| dialysis membrane, Molecular Weight Cut-Off (MWCO) 5,000 | Spectrum Medical Industries | Spectra/Por MWCO 5000 dialysis membrane, 133198 | |
| Immobilized Metal ion Affinity Chromatography (IMAC) column 1 mL, Ni-NTA | Macherey Nagel | Protino series, 745410.5 | |
| petri dishes (polystyrene, sterile) | Carl Roth | TA19 | |
| pQE-80L plasmid vector | Qiagen | no longer available | replaced by N-terminus pQE Vector set Cat No./ID: 32915 |
| protein extraction reagent BugBuster | EMB Millipore | 70921-4 | |
| round-bottom polystyrene tubes, 14 mL | Fisher Scientific | Corning Falcon, 14-959-1B | |
| Trp-auxotrophic E. coli strain | ATCC | ATCC 49980 | Bridges BA et al., Chem Biol Interact., 1972, 5(2):77-84; see main text for alternatives |
| Name | Company | Catalog Number | Comments |
| Mass Spectrometry equipment | |||
| mass spectrometer for LC-ESI-TOF-MS | Agilent | Agilent 6530 Accurate-Mass QTOF | coupled with Infinity LC system |
| mass spectrometry data analysis software | Agilent | MassHunter Qualitative Analysis software v. B.06.00 | |
| High-Performance Liquid Chromatography (HPLC) column for LC-ESI-TOF-MS | Sigma-Aldrich | Supelco Discovery BIO Wide Pore C5 HPLC column, 3 µm particle size, 10 cm x 2.1 mm | |
| HPLC autosampler vials 1.5 mL | Sigma-Aldrich | Supelco 854165 | with conical 0.1 mL glass inserts, screw caps and septa |
| Name | Company | Catalog Number | Comments |
| General equipment | |||
| benchtop centrifuge for 1.5 mL Eppendorf tubes | Eppendorf | 5427 R | |
| cooling centrifuge for 50 mL Falcon tubes | Eppendorf | 5810 R | |
| high pressure microfluidizer for bacterial cell disruption | Microfluidics | LM series with “Z” type chamber | |
| peristaltic pump for LC | GE Healthcare | P-1 | |
| Fast Protein Liquid Chromatography (FPLC) system | GE Healthcare | ÄKTA pure 25 L | |
| orbital shaker for bacterial cultivation | Infors HT | Minitron | |
| UV/Vis spectrophotometer | Biochrom | ULTROSPEC 2100 | |
| ultrasonic homogenizer for bacterial cell disruption | Omnilab | Bandelin SONOPULS HD 3200, 5650182 | with MS72 sonifier tip |
| Name | Company | Catalog Number | Comments |
| Fluorescence spectroscopy equipment | |||
| ps-pulsed laser 470 nm | Picoquant GmbH | PDL-470 | |
| time- and wavelength-correlated single photon counting (TWSPC) acquisition software | Picoquant GmbH | SymPhoTime 64 | |
| time- and wavelength-correlated single photon counting (TWSPC) detector | Picoquant GmbH | PML-16C | 16 spectral channels, to be selected by grating settings |
| single photon counting software | Picoquant GmbH | SPCM 9.75 | |
| global fitting software | Picoquant GmbH | SPC2Glo(R) | |
| fluorescence decay data analysis software | Picoquant GmbH | FluoFit program | |
| data analysis software | OriginLab Inc. | Origin 9.2 | |
| neutral density filter set | Schott | NG1 to NG11 | (400 - 650 nm, transmission 50 %, 20%, 10 %, 5 %) |
| 488 nm long-pass emission filter | AHF Analysentechnik | AHF-488 | |
| quartz cuvette | Thorlabs GmbH | CV10Q1400 | 1 cm pathlength |
References
- Shimomura, O., Johnson, F. H., Saiga, Y. Extraction, Purification and Properties of Aequorin, a Bioluminescent Protein from the Luminous Hydromedusan, Aequorea. J Cell Compar Physl. 59 (3), 223-239 (1962).
- Chalfie, M., Tu, Y., Euskirchen, G., Ward, W. W., Prasher, D. C. Green fluorescent protein as a marker for gene expression. Science. 263 (5148), 802-805 (1994).
- Andresen, M., et al. Structure and mechanism of the reversible photoswitch of a fluorescent protein. P Natl Acad Sci USA. 102 (37), 13070-13074 (2005).
- Andresen, M., et al. Structural basis for reversible photoswitching in Dronpa. P Natl Acad Sci USA. 104 (32), 13005-13009 (2007).
- Brakemann, T., et al. A reversibly photoswitchable GFP-like protein with fluorescence excitation decoupled from switching. Nat Biotechnol. 29 (10), 942-947 (2011).
- Kremers, G. -J., Gilbert, S. G., Cranfill, P. J., Davidson, M. W., Piston, D. W. Fluorescent proteins at a glance. J Cell Sci. 124 (Pt 2), 157-160 (2011).
- Shimomura, O. Structure of the chromophore of aequorea 0. shimomura green fluorescent protein. FEBS Lett. 104 (2), 220-222 (1979).
- Shaner, N. C., Campbell, R. E., Steinbach, P. A., Giepmans, B. N. G., Palmer, A. E., Tsien, R. Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat Biotechnol. 22 (12), 1567-1572 (2004).
- Shcherbo, D., et al. Bright far-red fluorescent protein for whole-body imaging. Nat Methods. 4 (9), 741-746 (2007).
- Shcherbakova, D. M., Subach, O. M., Verkhusha, V. V. Red fluorescent proteins: advanced imaging applications and future design. Angew Chem Int Edit. 51 (43), 10724-10738 (2012).
- Stepanenko, O. V., Verkhusha, V. V., Kuznetsova, I. M., Uversky, V. N., Turoverov, K. K. Fluorescent proteins as biomarkers and biosensors: throwing color lights on molecular and cellular processes. Curr Protein Pept Sc. 9 (4), 338-369 (2008).
- Wang, L., Xie, J., Deniz, A. A., Schultz, P. G. Unnatural amino acid mutagenesis of green fluorescent protein. J Org Chem. 68 (1), 174-176 (2003).
- Budisa, N., Steipe, B., Demange, P., Eckerskorn, C., Kellermann, J., Huber, R. High-level biosynthetic substitution of methionine in proteins by its analogs 2-aminohexanoic acid, selenomethionine, telluromethionine and ethionine in Escherichia coli. Eur J Biochem. 230 (2), 788-796 (1995).
- Sharma, N., Furter, R., Kast, P., Tirrell, D. A. Efficient introduction of aryl bromide functionality into proteins in vivo. FEBS Lett. 467 (1), 37-40 (2000).
- Liu, C. C., Schultz, P. G. Adding new chemistries to the genetic code. Annu Rev Biochem. 79, 413-444 (2010).
- Twine, S. M., Murphy, L., Phillips, R. S., Callis, P., Cash, M. T., Szabo, A. G. The Photophysical Properties of 6-Azaindole. J Phys Chem B. 107 (2), 637-645 (2003).
- Lepthien, S., Hoesl, M. G., Merkel, L., Budisa, N. Azatryptophans endow proteins with intrinsic blue fluorescence. P Natl Acad Sci USA. 105 (42), 16095-16100 (2008).
- Budisa, N., et al. Probing the role of tryptophans in Aequorea victoria green fluorescent proteins with an expanded genetic code. Biol Chem. 385 (2), 191-202 (2004).
- Ross, J. B., et al. Spectral enhancement of proteins: biological incorporation and fluorescence characterization of 5-hydroxytryptophan in bacteriophage lambda cI repressor. P Natl Acad Sci USA. 89 (24), 12023-12027 (1992).
- Soumillion, P., Jespers, L., Vervoort, J., Fastrez, J. Biosynthetic incorporation of 7-azatryptophan into the phage lambda lysozyme: Estimation of tryptophan accessibility, effect on enzymatic activity and protein stability. Protein Eng Des Sel. 8 (5), 451-456 (1995).
- Heim, R., Tsien, R. Y. Engineering green fluorescent protein for improved brightness, longer wavelengths and fluorescence resonance energy transfer. Curr Biol. 6 (2), 178-182 (1996).
- Bridges, B. A., Mottershead, R. P., Rothwell, M. A., Green, M. H. L. Repair-deficient bacterial strains suitable for mutagenicity screening: tests with the fungicide captain. Chem Biol Interact. 5 (2), 77-84 (1972).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Bacterial Transformation: The Heat Shock Method. J Vis Exp. , (2017).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Bacterial Transformation: Electroporation. J Vis Exp. , (2017).
- Grigorenko, B. L., Krylov, A. I., Nemukhin, A. V. Molecular modeling clarifies the mechanism of chromophore maturation in the green fluorescent protein. J Am Chem Soc. , (2017).
- JoVE Science Education Database. General Laboratory Techniques. Introduction to the Spectrophotometer. J Vis Exp. , (2017).
- Goedhart, J., et al. Structure-guided evolution of cyan fluorescent proteins towards a quantum yield of 93%. Nat Commun. 3, 751 (2012).
- Neidhardt, F. C., Bloch, P. L., Smith, D. F. Culture medium for enterobacteria. J Bacteriol. 119 (3), 736-747 (1974).
- Hörnsten, E. G. On culturing Escherichia coli on a mineral salts medium during anaerobic conditions. Bioprocess Eng. 12 (3), 157-162 (1995).
- Davis, B. D. The Isolation of Biochemically Deficient Mutants of Bacteria by Means of Penicillin. P Natl Acad Sci USA. 35 (1), 1-10 (1949).
- Sambrook, J., Russell, D. W. Molecular Cloning: A Laboratory Manual. , Cold Spring Harbor Laboratory Press. Cold Spring Harbor, NY, USA. (2001).
- Wang, Y. -S., et al. The de novo engineering of pyrrolysyl-tRNA synthetase for genetic incorporation of L-phenylalanine and its derivatives. Mol Biosyst. 7 (3), 714-717 (2011).
- Bae, J. H., et al. Expansion of the genetic code enables design of a novel "gold" class of green fluorescent proteins. J Mol Biol. 328 (5), 1071-1081 (2003).
- JoVE Science Education Database. Dialysis: Diffusion Based Separation. J Vis Exp. , Cambridge, MA. (2017).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Separating Protein with SDS-PAGE. J Vis Exp. , (2017).
- Petrásek, Z., et al. Excitation energy transfer from phycobiliprotein to chlorophyll d in intact cells of Acaryochloris marina studied by time- and wavelength-resolved fluorescence spectroscopy. Photoch Photobio Sci. 4 (12), 1016-1022 (2005).
- Kolber, Z. S., Barkley, M. D. Comparison of approaches to the instrumental response function in fluorescence decay measurements. Anal Biochem. 152 (1), 6-21 (1986).
- Pelet, S., Previte, M. J. R., Laiho, L. H., So, P. T. C. A fast global fitting algorithm for fluorescence lifetime imaging microscopy based on image segmentation. Biophys J. 87 (4), 2807-2817 (2004).
- Loefroth, J. E. Time-resolved emission spectra, decay-associated spectra, and species-associated spectra. J Phys Chem. 90 (6), 1160-1168 (1986).
- Hartman, M. C. T., Josephson, K., Lin, C. -W., Szostak, J. W. An expanded set of amino acid analogs for the ribosomal translation of unnatural peptides. PLoS One. 2 (10), e972 (2007).
- Budisa, N., et al. Global replacement of tryptophan with aminotryptophans generates non-invasive protein-based optical pH sensors. Angew Chem Int Edit. 41 (21), 4066-4069 (2002).
- Ma, Y., Biava, H., Contestabile, R., Budisa, N., di Salvo, M. L. Coupling bioorthogonal chemistries with artificial metabolism: intracellular biosynthesis of azidohomoalanine and its incorporation into recombinant proteins. Molecules. 19 (1), 1004-1022 (2014).
- Teramoto, H., Kojima, K. Incorporation of Methionine Analogues Into Bombyx mori Silk Fibroin for Click Modifications. Macromol Biosci. 15 (5), 719-727 (2015).
- Deal, R. B., Henikoff, J. G., Henikoff, S. Genome-wide kinetics of nucleosome turnover determined by metabolic labeling of histones. Science. 328 (5982), 1161-1164 (2010).
- Hinz, F. I., Dieterich, D. C., Tirrell, D. A., Schuman, E. M. Non-canonical amino acid labeling in vivo to visualize and affinity purify newly synthesized proteins in larval zebrafish. ACS Chem Neurosci. 3 (1), 40-49 (2012).
- Dieterich, D. C., et al. In situ visualization and dynamics of newly synthesized proteins in rat hippocampal neurons. Nat Neurosci. 13 (7), 897-905 (2010).
- Dieterich, D. C., Link, A. J., Graumann, J., Tirrell, D. A., Schuman, E. M. Selective identification of newly synthesized proteins in mammalian cells using bioorthogonal noncanonical amino acid tagging (BONCAT). P Natl Acad Sci USA. 103 (25), 9482-9487 (2006).
- Glenn, W. S., et al. Bioorthogonal Noncanonical Amino Acid Tagging (BONCAT) Enables Time-Resolved Analysis of Protein Synthesis in Native Plant Tissue. Plant Physiol. 173 (3), 1543-1553 (2017).
- Zhou, L., et al. Incorporation of tryptophan analogues into the lantibiotic nisin. Amino Acids. 48 (5), 1309-1318 (2016).
- Acevedo-Rocha, C. G., Budisa, N. Xenomicrobiology: a roadmap for genetic code engineering. Microb Biotechnol. 9 (5), 666-676 (2016).
- Agostini, F., Völler, J. -S., Koksch, B., Acevedo-Rocha, C. G., Kubyshkin, V., Budisa, N. Biocatalysis with Unnatural Amino Acids: Enzymology Meets Xenobiology. Angew Chem Int Edit. 56 (33), 9680-9703 (2017).
- Bacher, J. M., Ellington, A. D. Selection and characterization of Escherichia coli variants capable of growth on an otherwise toxic tryptophan analogue. J Bacteriol. 183 (18), 5414-5425 (2001).
- Wong, J. T. Membership mutation of the genetic code: loss of fitness by tryptophan. Pc Natl Acad Sci USA. 80 (20), 6303-6306 (1983).
- Hoesl, M. G., et al. Chemical Evolution of a Bacterial Proteome. Angew Chem Int Edit. 54 (34), 10030-10034 (2015).
- Italia, J. S., et al. An orthogonalized platform for genetic code expansion in both bacteria and eukaryotes. Nat Chem Biol. 13 (4), 446-450 (2017).
- Völler, J. -S., Thi To, T. M., Biava, H., Koksch, B., Budisa, N. Global substitution of hemeproteins with noncanonical amino acids in Escherichia coli with intact cofactor maturation machinery. Enzyme Microb Tech. 106, 55-59 (2017).
- Budisa, N., Steipe, B., Demange, P., Eckerskorn, C., Kellermann, J., Huber, R. High-level biosynthetic substitution of methionine in proteins by its analogs 2-aminohexanoic acid, selenomethionine, telluromethionine and ethionine in Escherichia coli. Eur J Biochem. 230 (2), 788-796 (1995).
- Völler, J. -S., Budisa, N. Coupling genetic code expansion and metabolic engineering for synthetic cells. Curr Opin Biotech. 48, 1-7 (2017).
- Johnson, J. A., Lu, Y. Y., Van Deventer, J. A., Tirrell, D. A. Residue-specific incorporation of non-canonical amino acids into proteins: recent developments and applications. Curr Opin Chem Biol. 14 (6), 774-780 (2010).
- Somsen, O. J., van Grondelle, R., van Amerongen, H. Spectral broadening of interacting pigments: polarized absorption by photosynthetic proteins. Biophys J. 71 (4), 1934-1951 (1996).
- Kurschus, F. C., Pal, P. P., Bäumler, P., Jenne, D. E., Wiltschi, B., Budisa, N. Gold fluorescent annexin A5 as a novel apoptosis detection tool. Cytom Part A. 75 (7), 626-633 (2009).
- Lepthien, S., Wiltschi, B., Bolic, B., Budisa, N. In vivo engineering of proteins with nitrogen-containing tryptophan analogs. Appl Microbiol Biot. 73 (4), 740-754 (2006).
- Wachter, R. M., Elsliger, M. -A., Kallio, K., Hanson, G. T., Remington, S. J. Structural basis of spectral shifts in the yellow-emission variants of green fluorescent protein. Structure. 6 (10), 1267-1277 (1998).
- Verkhusha, V. V., Lukyanov, K. A. The molecular properties and applications of Anthozoa fluorescent proteins and chromoproteins. Nat Biotechnol. 22 (3), 289-296 (2004).
- Martynov, V. I., Savitsky, A. P., Martynova, N. Y., Savitsky, P. A., Lukyanov, K. A., Lukyanov, S. A. Alternative cyclization in GFP-like proteins family. The formation and structure of the chromophore of a purple chromoprotein from Anemonia sulcata. J Biol Chem. 276 (24), 21012-21016 (2001).
- Piatkevich, K. D., Malashkevich, V. N., Morozova, K. S., Nemkovich, N. A., Almo, S. C., Verkhusha, V. V. Extended Stokes shift in fluorescent proteins: chromophore-protein interactions in a near-infrared TagRFP675 variant. Sci Rep. 3 (1), 1847 (2013).
