

Summary
ניצול של Cas9 preassembled ribonucleoprotein מורכבים (RNP) היא שיטה חזקה לעריכת הגנום מדויק, יעיל. כאן, אנחנו להדגיש השירות שלה על פני מגוון רחב של תאים, אורגניזמים, כולל תאים אנושיים ראשי שני קלאסי והעתידיים דגם אורגניזמים.
Abstract
בייעודי לאתר הגנום האיקריוטים עריכה עם CRISPR (באשכולות interspaced בקביעות קצר palindromic חזרה)-Cas (CRISPR-הקשורים) מערכות הפך במהירות שגרתי בקרב החוקרים רודף מגוון רחב של שאלות ביולוגיות. משתמשים לרוב מעסיקים החלבון Cas9 נגזר הפישחה ינפל תומימת סטרפטוקוקוס בתצמיד עם מדריך בקלות reprogrammed RNA (gRNA). רכיבים אלה הם הציגו לתוך תאים, ודבק דרך בסיס זיווג עם אזור משלימים של הגנום כפול גדילי הדנ א (dsDNA), האנזים בשני הגדילים לייצר הפסקה כפולה-סטרנד (DSB). תיקון עוקבות מוביל ההכנסה אקראית או אירועי מחיקה (indels) או שילוב של DNA שסופק על-ידי הנסיין באתר של ההפסקה.
השימוש של יחיד-מדריך RNA ו- Cas9 חלבון מטוהרים, preassembled כדי ליצור RNP ומסר ישירות לתאים, היא גישה חזק להשגת עריכה ג'ין יעילים ביותר. במיוחד RNP עריכה מגבירה את הקצב של ג'ין ההכנסה, תוצאה כי היא לעיתים קרובות מאתגרת כדי להשיג. בהשוואה המשלוח באמצעות פלסמיד התמדתו קצר יותר של RNP Cas9 בתוך התא מוביל לאירועים מחוץ-יעד פחות.
למרות יתרונותיה, רבים משתמשים אקראיים של עריכת ג'ין CRISPR הם פחות מוכרים בטכניקה זו. כדי להוריד את חסם כניסה, אנו מכינים פרוטוקולים מפורט ליישום האסטרטגיה RNP במגוון של הקשרים, המדגיש את היתרונות ברורים שלה ואת יישומים שונים ומגוונים. אנחנו מכסים עריכה של שני סוגי ראשי תאים אנושיים, תאי T ותאי גזע hematopoietic/קדמון (HSPCs). אנו גם מראים איך RNP Cas9 עריכה מאפשר מניפולציה גנטית נתיישב של אורגניזמים כולו, כולל את מודל קלאסי העגולה Caenorhabditis elegans וככל לאחרונה הציג מודל סרטנאים, Parhyale hawaiensis.
Introduction
ויתרונם CRISPR-Cas9 מערכת מאפשרת למדענים לשנות אזורים יישוב של כל הגנום1. טכנולוגיה זו מהירה וזולה יש מהפכה מחקר בסיסי, ומבטיח לעשות השפעה עמוקה על ההתפתחות של מחלת אישית טיפולים, חקלאות מדייקת, ומעבר2. עריכה CRISPR הוא כלי דמוקרטי ודורש יישום המערכת במעבדה החדשה אין מומחיות מיוחדת בכישורי ביולוגיה מולקולרית הנדסית, בסיסית גנום. חוקרים יכולים ללמוד עכשיו בעבר סורר אורגניזמים עם כמה אמצעים חלופיים עבור הנדסה גנטית3,4. בחמש השנים האחרונות לבד, עריכה הגנום CRISPR שימש להנדס מעל 200 החולייתנים שונים, חסרי חוליות, צמח, ועוד מיני חיידקים.
הותאם מהשביל ההגנה prokaryotic CRISPR, האלמנטים הליבה הנדרשים לעריכת הגנום בייעודי לאתר החלבון Cas9, בדרך כלל מתוך ס' הפישחה ינפל תומימת , הממוטבים codon עם אות לוקליזציה גרעיני נוסף (שקל), ממוקצעות שלה מדריך RNA5,6. אך לא נדון כאן, Cas9 orthologues או CRISPR endonucleases אחרים עשוי לשמש גם. GRNA טבעי מורכב שתי חתיכות משועתקים בנפרד, את ה CRISPR (crRNA) והפעלת את הטרנס-crRNA (tracrRNA)7. אלה RNAs יכול להיות מותך לתוך תעתיק יחיד, המכונה RNA (sgRNA) יחיד-מדריך8. מרוב העורכים הגנום לבחור sgRNA מפושטת9, למרות כפול-המדריך הוא גם משמש באופן קבוע10,11. ניסויים לבחור 20-נוקלאוטיד (nt) גנומית DNA למטרה להבטיח כי הוא שוכן בסמוך חתימה רישוי קצר נדרש Cas9 הכרה, שנקרא מוטיב סמוך protospacer (פאם), ולעצב gRNA המכיל את רצף משלים12 .
פעם אחת בתוך התא, מאתרת RNP מורכבים מטרתה גנומית, זוגות בסיס gRNA עם ה-DNA משלימים לנטישה, ודבק ואז האנזים שני גדילי הדנ א כדי ליצור זוגיות-בשרך לשבור את2. תא תיקון מכונות פותר את DSB על ידי אחד לפחות שני מסלולים: דרך השביל (NHEJ) מועדת לטעויות הלא-הומולוגי סוף-הצטרפות או התיקון מכוון הומולוגיה (HDR), אשר משלבת בצורה חלקה DNA המכיל 'סמל' הומולוגיה אל צדי השבר. מסלול לשעבר תיקון בדרך כלל מוביל ויאסין היווצרות ובהפסקות ג'ין הסוגר, בעוד שהשני מאפשר ניסויים להוסיף או לשנות DNA רצפים1.
היעילות והדיוק העריכה תלויים האמצעי שבאמצעותו Cas9 ו- gRNA להיכנס לתוך התא. רכיבים אלה עשויים להינתן תאים בתרבית, העוברים או אורגניזמים בדמות של חומצות גרעין, או כמו preassembled RNP מורכבים13,14,15. שיטות משלוח מבוססי חומצת גרעין הנפוצות כוללות את התמרה חושית ויראלי, תרביות תאים או אלקטרופורציה של mRNA או פלסמיד ה-DNA. חלבון Cas9 מדריך RNA ואז מיוצרים בתוך התא ולשייך להם להקים מתחם.
מסירה ישירה של RNP דורש הטיהור נפרדים של חלבון Cas9, מדריך ה-RNA. ניתן לבצע זאת בתוך הבית, או חלבונים ו- sgRNA ניתן לרכוש מאחת מספר ספקי מסחרי. מרגע שהוא נרכש, Cas9 ואת gRNA מעורב להקים במתחם RNP enzymatically המוסמכת, הציג לתאים על-ידי הזרקה ישירה לתוך הביצים מופרית/עוברי, תרביות תאים מבוססת השומנים16או אלקטרופורציה. הדוח הראשון של עריכת RNP מעורב הזרקה לתוך C. elegans גונדות17. Microinjection הוא עדיין האמצעי המועדף של ידביקו RNP עוברי, אורגניזם שלם, אבל יעיל אלקטרופורציה הוכח ב העכבר18,19 ו עכברים20 עוברי. לתאר את הפרוטוקולים עבור ישירות הזרקת RNP לתוך גונדות C. elegans , פ hawaiensis עוברי ואנו ממליצים על סוג מיוחד של אלקטרופורציה לספק RNP בעת עריכת ראשי תאים אנושיים. שיטה זו, nucleofection, כרוכה אלקטרופורציה ממוטבת תוכניות ופתרונות ייחודיים לסוג התא ומאפשר את RNP להזין הציטופלסמה והן את גרעין21.
הגנום עריכה עם RNP מציע מספר יתרונות ברורים. כי ה-חלבון ורכיבי ה-RNA הם מראש, והן יכול להיות איכות לפני הלידה, RNP עריכה מונע החסרונות רבים הקשורים עם המשלוח מבוססי חומצת גרעין. כלומר, אין סיכון של שילוב קידוד Cas9 DNA הגנום מארח mRNA מעולם לא נחשף על השפלה, זה עוקף בעיות ויוו gRNA או חלבון, קיפול, ואת זכות ההתאגדות22,23. יתר על כן, באמצעות RNP מוביל להפחית רעילות ואירועים את המטרה הרבה פחות מאשר הביטוי מבוסס פלסמיד, תוצאה של מחצית חיים קצר של RNP פנימה התא24,25,26,27.
לבסוף, RNP עריכה כאפשרית מוביל תעריפי עריכה גבוהה במגוון של שורות תאים אנושיים, ראשי תאים כגון fibroblasts, תאי גזע עובריים (ESCs), המושרה בתאי גזע pluripotent (iSPCs), HSPCs, T תאים16,24, 25,26,27,28,29; בחסרי חוליות כולל C. elegans, פ hawaiensisשל זבובי פירות3,17,30; במינים חוליות כמו דג זברה, עכברים, חולדות31,32; ב לשתול מיני כולל תודרניתטבק, חסה, אורז, הגפן, אפל, תירס, חיטה33,34,35,36; וב Chlamydomonas, Penicilliumו קנדידה מינים37,38,39. התדירות של היווצרות ויאסין יכול להיות גבוה יותר בעת שימוש RNP בהשוואה המשלוח פלסמיד, ההכנסה בתיווך HDR דנ א יכול להיות קל יותר להשיג27,25,29.
הפרוטוקול המתואר כאן משתמש RNP Cas9 והיא טכניקה יעילה וישימה בקלות, זה פשוט להחיל על מגוון רחב של מערכות ביולוגיות40,41, בייחוד בתאים אחרת קשה לעבוד עם היצורים ללא מערכות ומבוססת על מניפולציה גנטית מדויקת. נתחיל על-ידי המתארות כיצד לעצב, להשיג, ולהרכיב את RNP Cas9 לפני מכסה את השימוש בו על פני סוגי תאים שונים דגם ואורגניזמים. גזע hematopoietic/קדמון תאים (HSPCs), תאי T נערכים באמצעות אותה שיטה, nucleofection, כך שהם ביחד מכוסים שלבים 2 ו- 3 של פרוטוקול זה. עריכת נהלים עבור C. elegans מתוארים שלבים 4 ו- 5, ו- P. hawaiensis עריכת מכוסה שלבים 6 ו- 7. לבסוף, מאז ההצלחה של ניסוי גנטי לעריכת ב מכל יצור עשוי להיות מוערך על ידי גנוטיפ רצף, substeps המתארת שיטות ניתוח אפשרי עבור כל התאים ואת אורגניזמים תיאר בפרוטוקול מתוארים בשלב 8.
Protocol
1. RNP הרכבה
-
תכנון הניסוי מראש, רכישת כל רכיבי ה-RNA DNA, חלבונים מבעוד מועד. כמו לעבור הראשון, נסה אחד מהפקדים חיובי המפורטים בטבלה 1 והשתמש של ריאגנטים מסחרי המתוארות בטבלה של החומרים כדי להבטיח עיצוב ניסיוני אמין והיושרה של החומרים. לקבלת עצות נוספות תכנון ניסוי חדש לעריכת הגנום, לראות את הניירות על זה42,12,נושא43.
הערה: לאחר התכנסה כפי שמתואר והשלבים הבאים, RNPs שהוכנה מראש ניתן לאחסן ב- 80 ° c- לאחר בחירת אילו גנים למטרה, להשתמש באחד הכלים מקוון חינם כדי לעצב של האופטימלית gRNA44,45,46,47,48. ודא כי המטרה של אקסון אם מקווה לייצר פצצה.
הערה: כלים אלה תסייע לזהות אתר יעד עם סמוכים הפישחה ינפל תומימת ס פאם רצף באיכות גבוהה ציון, ציון נמוך את המטרה. - לטהר את החלבון Cas9 הפישחה ינפל תומימת ס באמצעות שיטות שפורסמו8, או לרכוש אותו ספק מסחרי.
- להכין מאגר Cas9 אופייני RNA דילול הכנה RNP, אחסון חלבון, אשר מכיל 20 מ מ של HEPES pH 7.5, 150 מ מ של אשלגן כלורי, גליצרול 10% 1 מ מ TCEP. תמיד להשתמש במים נטולי נוקלאז במאגרי שישמשו resuspend או לדלל RNA כדי למנוע השפלה.
- לייצר מדריך RNA (tracrRNA, crRNA או sgRNA) דרך שעתוק במבחנה באמצעות שיטות שפורסם, או לרכוש אותו חומצת גרעין סינתזה החברה17,21,49, 50 , 51.
- אם הוספת גן, לסנתז או לרכוש תורם תבנית ה-DNA.
- לאחסן חלבונים ו- RNA aliquots-80 ° C, הפשרת הקרח מיד לפני השימוש.
הערה: כל ההקפאה-להפשיר מעט מוריד את היעילות. פרוטוקולים מפורט, גישה פתוחה עבור טיהור Cas952 ו- in vitro לקשרי שעתוק של sgRNAs53 זמינים במקום אחר.
- לאחר בחירת אילו גנים למטרה, להשתמש באחד הכלים מקוון חינם כדי לעצב של האופטימלית gRNA44,45,46,47,48. ודא כי המטרה של אקסון אם מקווה לייצר פצצה.
- אם עובדים עם C. elegans, דלג לשלב 1.5. עבור פרוטוקול hawaiensis פ , דלג לשלב 1.6. אם משתמש sgRNA, דלג לשלב מס ' 1.4. המשך לצעוד 1.3 להרכיב gRNA לעריכה התא הראשי.
-
להרכיב את gRNA על ידי ערבוב כמויות equimolar של tracrRNA ו- crRNA. להפוך µL 100 80 מיקרומטר gRNA במניה, כ-50 הגנום עריכת ניסויים.
- דגירה של gRNA ב 37 מעלות צלזיוס למשך 30 דקות, ואז לאפשר לו לאט להתקרר לטמפרטורת החדר.
-
RNP הכנה עבור HSPC ו- T תא עריכה: להרכיב RNP מורכבים על ידי ערבוב 1-2 x שן טוחנת כמות של gRNA כדי pmol 200 של חלבון Cas9 הנפח הכולל של µL 10. לאט מאוד, להוסיף Cas9 מרוכז gRNA (מדולל מראש במאגר Cas9) עבור 30 s , במעגלים מהירה עם הפיפטה מביא את הריכוז Cas9 הסופי 20 מיקרומטר.
- הכינו את אלקטרופורציה וואקום.
הערה: פרוטוקול זה ספציפיים למערכת מסחרי הנזכרים בטבלה של חומרים, אבל RNP עריכה גם ניתן להשיג עם התקנים אחרים אלקטרופורציה. - להוסיף 5 µL (100 pmols, תאי T) או 10 µL (200 pmol, HSPCs) של RNP כל cuvette.
- אם הוספת DNA חדש במקום לבצע נוק אאוט, להוסיף 1 µL של 100 מיקרומטר (100 pmol) חד-גדילי oligonucleotide התורם דנ א (ssODN)25,54,55 וואקום או בארות של הצלחת.
- דלג על שלב 2 עבור ההוראות הבאה עריכת התא הראשי פרוטוקול.
- הכינו את אלקטרופורציה וואקום.
-
פרטי RNP לעריכה C. elegans : להרכיב את RNP מורכבים על-ידי הוספה של ריאגנטים הבאים כדי ליצור נפח סופי של 20 µL (ריכוז סופי אתה מצוין בסוגריים): Cas9 (2 מיקרומטר), HEPES pH 7.5 (10 מיקרומטר), אשלגן כלורי (115 מיקרומטר), crRNA (12 מיקרומטר) , tracrRNA (40 µM), את תיקון תבניות במידת הצורך (0.5 מיקרומטר ssDNA או עד 350 dsDNA ng/µL).
הערה: היעילות של תיקון בתיווך Cas9 DSB תבניות הוא פרופורציונלי לריכוז הבונה תיקון dsDNA; לפיכך, גבוה יותר את הריכוז של תבנית תיקון, לייעל התיקון בתבניות. עם זאת, זריקה של תערובות המכילות גדול מ- 350 ng/µL של dsDNA הוכח כדי להקטין את הכדאיות של תולעים מוזרק. לכן, עדיף להשתמש, אך לא יותר מ 350 ng/µL של dsDNA בתערובת כדי למקסם את היעילות תיקון תוך מזעור הקטלניות שלה.- הוסף crRNAs מרובים כדי מטרה לוקוסים מרובים בו זמנית, כנדרש עבור הגישה ההקרנה co-CRISPR/co-conversion המתוארת בשלב 5.4. בעת הוספת crRNA אחד או יותר, הוסף אחד ברצף לתערובת הראשית.
הערה: הכמות של כל crRNA לא צריך להיות זהה, אפילו הכפלת ריכוז מוחלט של crRNAs בתערובת מאסטר מבלי לשנות את הריכוז של Cas9 לא מופיע להתערב עם תדירות מוטגנזה מכוונת-לוקוס ספציפי. דוגמאות מתוארים בפירוט Paix. et al. 56. - לערבב על ידי pipetting, לסובב את הפתרון RNP ב g 16,000 x 5 s כדי להבטיח שהפתרון בתחתית הצינורית.
- דגירה הפתרון ב 37 ° C 15 מ'.
- Centrifuge המדגם-16,000 g x עבור 1 דקות הצניפה חלקיקים כלשהם כי יכול לסתום את המחט microinjection דק-משועמם. להשתמש את תגובת שיקוע והשלבים הבאים.
- דלג לשלב 4 עד לתום של פרוטוקול C. elegans .
- הוסף crRNAs מרובים כדי מטרה לוקוסים מרובים בו זמנית, כנדרש עבור הגישה ההקרנה co-CRISPR/co-conversion המתוארת בשלב 5.4. בעת הוספת crRNA אחד או יותר, הוסף אחד ברצף לתערובת הראשית.
-
פרטי RNP לעריכה hawaiensis פ : להכין לשימוש יחיד Cas9 aliquots על ידי דילול אותם עם מים ללא נוקלאז פנול אדום (להמחשת זריקות) כדי ריכוז סופי של 6.25 מיקרומטר של Cas9 ו- 0.15% פנול אדום.
- להרכיב את RNP מורכבים על ידי ערבוב 2-5 x שן טוחנת עודף של gRNA את החלבון Cas9 של הנפח הכולל של 6 µl. להוסיף 12 pmol של Cas9 כדי gRNA, להביא את הריכוז הסופי של Cas9 2 מיקרומטר, gRNA 4-8 מיקרומטר לריכוז פנול אדום בריכוז 0.05%.
- התערובת בטמפרטורת החדר במשך 10 דקות למתחם תדגור על RNP
- דלג לשלב 6 על ההוראות הבאה בעריכת hawaiensis פ פרוטוקול.
2. התא תרבות והכנות
הערה: לבצע צעדים 2.1.1 כדי 3.3.3 בטיחות ביולוגית ארון.
-
האדם cryopreserved הרכישה גייסה דם היקפיים CD34+ HSPCs מספק.
- הפשרת ~ 1 x106 HSPCs במים 37 ° C אמבטיה למשך 3 דקות ומעבירים אותם אל צינור חרוטי 15 מ"ל. להוסיף 10 מ של מדיום הרחבה ללא סרום ממקור מסחרי ולסובב את התערובת ב g 100 x במשך 10 דקות להסיר את תגובת שיקוע resuspend התאים 2 מ של SFEM שהושלם. צלחת התאים לוחות 6-ובכן, תרבות אותם בתוך חממה 37 מעלות צלזיוס במשך 24-48 שעות לפני אלקטרופורציה RNP.
- לספור את התאים עם hemocytometer ולהעביר את המספר הכולל של HSPCs צריך (150,000-200,000 HSPCs לכל cuvette להיות electroporated) שפופרת צנטרפוגה.
- לסובב את הצינור ב- 100 גרם x 10 דקות הצניפה התאים.
-
לרכוש CD4 העיקרי האנושי+ T תאים מיצרן או לבודד אותם מן דם כל אדם על ידי צנטריפוגה הדרגתיות צפיפות29.
- לפני ההפעלה תא T, מראש מעיל 48-ובכן תרבות צלחות עם αCD3 (UCHT1), αCD28 (CD28.2). מעיל הצלחות עם 500 µL של αCD3 µg/mL 10, 10 αCD28 µg/mL. ב- PBS כבר לפחות שעתיים ב 37 º C.
הערה: עבור כמה לוקוסים, NHEJ יכולה להיות מושגת ללא גירוי מקדים, אך שלב זה כולל הגדלת היעילות שלה. - תרבות ה-T תאים עבור h 48-37 מעלות צלזיוס על αCD3/αCD28 צלחות נוגדן מכורך במדיום מלאה RPMI [RPMI-1640 בתוספת 5 מ"מ של HEPES, 2 מ מ של חלופה מסחרי-גלוטמין, 50 µg/mL של פניצילין/סטרפטומיצין, 50 מיקרומטר של מרקפטואתנול, 5 מ מ שאינם חיוניים חומצות אמינו, 5 מ מ של נתרן פירובט, ו-10% (vol/כרך) FBS]. תרבות ה-T תאים על צפיפות של 2,000,000 בתאי T ב- 500 µL של המדיה לכל טוב של צלחת 48-. טוב.
- ספירת ה-T תאים באמצעות hemocytometer העברת המספר הכולל של תאי T הדרושות אלקטרופורציה הניסוי (100,000-1,000,000 תאי T cuvette להיות electroporated) כדי שפופרת צנטרפוגה.
- לסובב את הצינור ב g x 90 במשך 8 דקות הצניפה התאים. אם התאים יש כבר צפיפות מופרד הדרגתי תוך יומיים, לסובב אותם ב g x 200 במשך 8 דקות.
- לפני ההפעלה תא T, מראש מעיל 48-ובכן תרבות צלחות עם αCD3 (UCHT1), αCD28 (CD28.2). מעיל הצלחות עם 500 µL של αCD3 µg/mL 10, 10 αCD28 µg/mL. ב- PBS כבר לפחות שעתיים ב 37 º C.
-
עבור שני סוגי תאים, וארוקן את תגובת שיקוע עם פיפטה/ואקום, הסרת בועות.
- Resuspend בעדינות את התאים עם µL 20 אלקטרופורציה מאגר לכל cuvette.
- הוסף 20 µL של התאים (150,000-200,000 HSPCs או תאי T 100,000-1,000,000) כל cuvette, וכבר מכילה 10 µL RNP, לערבב היטב על ידי pipetting למעלה ולמטה ללא יצירת בועות.
3. RNP אלקטרופורציה
- Electroporate כימיקלים לאחר הצבתם nucleofector. עבור HSPCs, להשתמש בקוד הדופק ER100. עבור תאי T, להשתמש בקוד הדופק EH-115.
-
HSPCs בלבד: להוסיף 100 µL שהושלם SFEM מדיום (התחמם עד 37 ° C) על כל cuvette מיד לאחר אלקטרופורציה ולתת שהתאים להתאושש במשך 10-15 דקות
- העבר את התאים תרבות אותם ב 96-ובכן עגול-תחתון צלחת ולהוסיף של µL 100 נוספים של המדיום SFEM שהושלם עבור 24 שעות.
- לשנות אותם למדיום SFEM שהושלם טריים, תקופת דגירה אותם של 24-72 h נוספים.
- להסיר את התאים עבור genotyping אותם 48-96 h פוסט-אלקטרופורציה. לסובב את התאים ב g x 300 במשך 5 דקות ולהסיר את תגובת שיקוע לפני תחילת הפקת דנ א (שלב 8.2).
-
T תאים בלבד: להוסיף 80 µL של RPMI להשלים את תרבות המדיה מראש התחמם עד 37 ° C מן המאגר לכל cuvette או, ובכן, בעזרת פיפטה של רב ערוצית (במקרה הצורך).
- דגירה אותם ב 37 מעלות צלזיוס למשך 15 דקות.
- הוסף את המדיה המתאימה, נוגדנים, ציטוקינים, וכו ' כדי plate(s) היעד, לחמם אותם בתוך חממה 37 ° C.
- העברה µL 107 של התאים electroporated מן הבארות צלחת 96-ובכן סיבוב המדרגה באמצעות פיפטה של רב ערוצית (במקרה הצורך).
- לקבלת מידע אודות הערכת התוצאות העריכה, דלג לשלב 8.
4. C. elegans הכנה
-
יום אחד לפני microinjection: להכין את רפידות agarose microinjection.
- להפוך את פתרון agarose 3% (w/v) במים על ידי הוספת agarose מים, מביא את הפתרון לרתיחה על פלטה חמה או במיקרוגל.
- לארגן 24 מ"מ x 50 מ"מ x 1.5 מ"מ חיפוי זכוכית שקופיות על שולחן ולהשתמש כוס פסטר פיפטה למקום טיפה קטן (µL ~ 15) של הפתרון agarose אותן לשקופית. במהירות לשטח הירידה agarose על ידי הצבת coverslip עוד למעלה. אפשר את agarose לחזק ולאחר מכן להסיר את אחד coverslips.
- להשאיר את הפנים coverslip מצופים agarose פלטת שולחן לילה יבש. לאחר 24 שעות, לאחסן את רפידות agarose במיכל נקי, יבש.
הערה: אלה יכולים לשמש ללא הגבלת זמן.
- משוך את המחטים microinjection: באמצעות נימי זכוכית בורוסיליקט זיריהם (הקוטר החיצוני 1.0 מ מ והפנימית קוטר מ"מ 0.58), למשוך את המחטים. המבוססת על מלו ואש57 ו משאבים אחרים58. המחטים יכול לשמש באופן מיידי או ניתן לאחסן במיכל נקי, יבש, חיבק מאת קליי תומך.
- לשם קיום התולעים, להכין אגר מדיה צמיחה תולעים נימיות (NGM) נשפך לתוך צלחות פטרי, מרובבים חיידקים OP50 (עבור פרוטוקולים על תקן C. elegans תחזוקה, מתכונים צמיחה מדיה, ראה Stiernagle59).
- שלב את התולעים עבור microinjection: 12-24 שעןת לפני microinjection, לאסוף אנדרוגינוסים מבוים-L4 ל צלחת NG-אגר חדשה עם חיידקים OP50 ואת דגירה אותם בן לילה ב 20 º C. עבור כל תערובת הזרקה היעד Cas9, לבחור ~ 30 תולעים לצלחת.
-
ימי microinjection: לטעון את המחט microinjection משך עם RNP הפתרון supernatant מוכן בשלב 1.5.
- Pipette את תגובת שיקוע מהשלב 1.5.4 לתוך פיפטה נימי משך מילוי הפתרון מ פיפטה נימי לתוך המחט microinjection מוכן (טעינת בדרך כלל פחות מ 0.1 µL).
- הר המחט טעון על המנגנון microinjection מצורף של micromanipulator. הגדר את הלחץ מכשיר הזרקת 250 kPa ואת הלחץ איזון 25 kPa.
-
לשבור את מחט טעון כדי ליצור קצה מחט חדה. המקום 15 מ"מ x 15 מ"מ x 1.5 מ מ מרובע coverslip בחלק העליון 24 מ"מ x 50 מ"מ x 1.5 מ מ coverslip.
- כיסוי לאחד מקצוות coverslip מרובע עם שמן halocarbon 700.
- מיקום המחט בשמן, בקצה coverslip מרובע 15 מ מ.
- תוך שימוש ביד להנחות את המיקרוסקופ הבמה ואת coverslip, מברשת השקופית, לאורך קצה המחט תוך מדכא דוושת/לחצן הזרקה. לשבור את מחט, להגדיל את זרימת הנוזל החוצה המחט. להשיג שיעור זרימה אופטימלית על ידי ביצוע הזריקה לערבב זרימה לאורך קצה המחט, ויצרו בועה ~ 1/s.
- לאשר כי התולעים L4 בחרה 12-24 שעןת לפני microinjection הן מבחינת בשלבים צעירים ביום של הזרקת. לבחור צעיר תולעים בוגרות כדי צלחת NG-אגר חסר חיידקים OP50 ולאפשר להם לזחול סביב במשך 5 דקות. זה מפחית את כמות החיידקים להעביר משטח הזרקה, מזעור המחט כפכפים.
- במקום משטח/coverslip של הזרקת agarose על גבי טווח לנתיחה. באמצעות איסוף תולעת, שכב על מסלול קטן של שמן halocarbon לאורך קצה אחד של משטח.
-
באמצעות האיסוף תולעת מצופה בשמן, הרם מספר תולעים רלוונטי NG-אגר, לתוך המסלול של שמן. עם שיער בסדר המצורפת פיפטה, כגון משערה עפעף או חתול, מקם את התולעים במקביל, בעדינות לדחוף את התולעים אל תוך כרית agarose. עד נוחים עם ההליך microinjection, רק הר ולהזריק תולעת אחת בכל פעם.
הערה: agarose יבש לפתיל הלחות מן התולעים, גורם להם לדבוק בלוח. כתוצאה מכך, אחד חייב לעבוד במהירות כמו התולעים יכולים וכשתתחילי.- שינוי מיקום, מצורף משטח, כיסוי התולעים עם עוד כמה טיפות שמן halocarbon (µL ~ 20) מהקצה של התולעת לבחור.
5. C. elegans בלוטת Microinjection עם RNPs ולדאוג שלאחר הזרקה
הערה: פרוטוקול microinjection מותאמת של מלו ואש57ותיאר בפירוט במקום60,61.
-
מניחים את coverslip עם התולעים רכוב על גבי מיקרוסקופ הזרקה. תחת הגדלה נמוכה (5 X אובייקטיבי, 10 X עינית), מקם את התולעים בניצב המחט הזרקה.
- המתג בהגדלה גדולה (40 X אובייקטיבי, 10 X עינית), למקם מחדש את המחט סמוכים את הזרוע בלוטת המין המתאים לאזור בקרבת הגרעינים ב אמצע - עד מאוחר-pachytene.
- שימוש של micromanipulator, להזיז את המחט נגד התולעת, מדכא את הקוטיקולה מעט. לאחר מכן, עם יד אחת, הקש על הצד של הבמה מיקרוסקופ כדי לדפוק לך את המחט הקוטיקולה. נלחצים דוושת/לחצן הזרקת לאט למלא את הזרוע בלוטת המין עם המיקס הזרקה, הסר את המחט.
- חזור על שלב זה עם היד השניה בלוטת המין.
-
ברגע שהם מזריקים את התולעים, להסיר את משטח coverslip/agarose והנח אותו תחת מיקרוסקופ ויבתר.
- באמצעות פיפטה נימי משך, תחסיר השמן מן התולעים מאת pipetting מאגר M9 מעליהם. לבצע טיפול זה לשחרר את התולעים מ אגר.
- לאחר 10 דקות, כאשר התולעים הן מכות סביב במאגר, להעביר אותם אל צלחת NG-אגר עם חיידקים OP50 באמצעות פיפטה נימי משך. מקם את הצלחת ב 20 מעלות צלזיוס במשך 2-3 h עד התולעים התאוששו הם לנוע.
- ברגע התאושש, בנפרד להעביר את התולעים NG-אגר צלחות עם OP50 ו להעביר את הצלחות חממה 25 ° C.
-
לאפשר את P0-מוזרק תולעים לגדול ולהניח רומא למשך 3 ימים. מסך אופספרינג1 נ.
- אם משתמש co-CRISPR או המרה שיתוף62,63,64,65, ולאחר מכן בחר את התולעים המועמד להקרנה בהתבסס על אם יש להם את ותפקיד מוטציה של הגן הפניה. בנפרד להעביר תולעים מסומן אלה לוחיות הרישוי NG-אגר עם OP50 ולאפשר להם להניח את רומא2 F ב- 20 ° C.
הערה: פנוטיפ המשמש עבור co-CRISPR הקרנה או בחירה צריך לספק הערכה מוקדמת להצלחת Cas9 עריכה. - אם פנוטיפ co-CRISPR אינו נוכח, microinject של פלסמיד בקרה חיובית כדי לסייע בשיפור היעילות microinjection.
הערה: למשל, כולל על פלסמיד בתערובת הזרקת מקודד מתויג mCherry MYO-2 תסייע להעריך את יעילות הזרקת. תולעים בהצלחה הזריק pCFJ90 יהיו כמה צאצאים עם pharynxes פלורסנט.
- אם משתמש co-CRISPR או המרה שיתוף62,63,64,65, ולאחר מכן בחר את התולעים המועמד להקרנה בהתבסס על אם יש להם את ותפקיד מוטציה של הגן הפניה. בנפרד להעביר תולעים מסומן אלה לוחיות הרישוי NG-אגר עם OP50 ולאפשר להם להניח את רומא2 F ב- 20 ° C.
- לבחון את התולעים1 F לנוכחות של שינויי העריכה הרצוי. לבחור האם1 F טוב בודדים של צלחת 96-ובכן, lyse אותה, ולבחון את הדנ א שלה על-ידי הוספה ספציפיים PCR הגברה, ניתוח רצף הדנ א או מודד נוקלאז assay (CEL-1)66.
הערה: מבחני אלה ניתן לבצע כאשר באמצעות הקרנה co-CRISPR/co-conversion או אחרים או בחירה משטרים65,66,67,68. - לקבלת מידע אודות הערכת התוצאות העריכה, דלג לשלב 8.
6. הכנה hawaiensis פ
- יום אחד לפני microinjection, להעשיר עבור העוברים מוקדם על-ידי הגדרת 'טנק זוג' בלילה שלפני; נקבות מופרדים החדש יכיל העוברים טריים מופרית. ראה Rehm. et al. 69 לפרטים.
- ביום microinjection, לאסוף את העוברים Parhyale תא בודד (0-4 h שלאחר ההפריה) מאת מאלחש נשים gravid עם שמן ציפורן 0.02% במי ים ולשרוט בעדינות את העוברים מחוץ לה מחדירות הגחון באמצעות להבה-משך ו פיפטה מזכוכית מעוגלות, זוג מלקחיים #3 משעמם.
7. hawaiensis פ Microinjection העובר עם RNPs ולדאוג שלאחר הזרקה
- מילוי צינור קפילרי משך עם 1 µL של המיקס הזרקת RNP שתוארו לעיל.
-
השתמש חנקן דחוס כדי microinject כל העובר כפי שמתואר Rehm. et al. 69.
- להזריק את העוברים Parhyale תחת מיקרוסקופ ויבתר באמצעות microinjector של micromanipulator. לטעון µL 1.5 של המיקס הזרקת לחלק האחורי של צינור קפילרי משך (4 אינץ ' - 1.0 מ"מ זיריהם, משך שימוש micropipette מושך המנגנון) באמצעות טיפ פיפטה microloader.
- להגדיר את המחט על המנגנון הזרקת ולשבור את קצה המחט (כמות קטנה מאוד) באמצעות זוג מלקחיים מתחת למיקרוסקופ ויבתר. כיילו את עוצמת הקול על-ידי הזרקת לתוך שמן halocarbon 700 ומדידת קוטר הבועה.
- חותכים "שוקת". הסוכן ריפוי בעזרת סכין גילוח. למלא אותו באמצע הים מחוטא-מסנן מים, בשורה העוברים Parhyale בשוקת לייצב.
- להזריק את העוברים באמצעות הגדרת microinjection, ייצוב כל העובר עם זוג מלקחיים במהלך ההזרקה. לאחר ההזרקה, השתמש פיפטה העברה של זכוכית להעברת העוברים מעל לצלחת תרבות טריים 60 מ מ מלא באמצע הים מחוטא-מסנן מים.
-
אם לליגה הראשונה כבר התרחשה לטופס העובר 2-תא (4-6 שעות לאחר ההפריה), ליצור חיות באופן מלא-מוטציה על ידי הזרקת blastomeres שני. כדי להבטיח המחשוף הכוללת של השלב 2 תאים, שיתוף להזריק את blastomeres עם FITC או TRITC לתוספי ולצפות כי האות הוא מוגבל blastomere יחיד תחת פלורסנט לנתח היקף לאחר הזריקה.
- לחלופין, צור 'חצי-מוטציה' חיות באמצעות הזרקת רק לאחד שני blastomeres בשלב 2-תא (מחולק בערך משמאל בהתאם רקמות תפקיד לאורך הציר A-P).
- להזריק תא אחד העובר שמונה תאים (7.5-9 h שלאחר ההפריה) כדי להגביל את עריכת לשכבה נבט יחיד. ראה Gerberding. et al. 70 עבור מפה של שושלות blastomere המוקדמות.
-
דגירה העוברים במנות 60 מ מ תרבות (לא יותר מ 25 לכל מנה), מלא באמצע הדרך עם ים מחוטא-מסנן 'מראש חמצן' באמצעות bubbler אקווריום של או ע י ניעור נמרצות.
- מקם את המנות של עוברי plasticware אטום באופן רופף עם מגבות נייר רטוב כדי לשמור על לחות ומניחים חממה 26 ° C עם מחזור בהירה-כהה 12 שעות.
- העברת העוברים התקינים לניקוי מי ים מנות כל כמה ימים.
הערה: עוברי עשוי להיות מחונן בטמפרטורת החדר, למרות שהם יפתחו הרבה יותר לאט
-
לפרק ולתקן את העוברים בשלבים שונים עבור ניתוח ביטוי מאת בחיי עיר הכלאה או נוגדן מכתימים (ראה בראון. et al. 71 מדריך הזמני, הפניות נוספות עבור ניתוח של קיבוע72, בחיי עיר הכלאה73נוגדן מכתים74).
- לעשות ניתוח מחטים על ידי שרשור חתיכת מושחת של טונגסטן תילים כ 0.5 ב אורך לתוך סוף מחט של אינסולין. לחדד את המחט בתוך נתרן הידרוקסידי תחת זרם. השתמש מזרק 1 מ"ל כמו הידית של המחט לנתיחה.
- מילוי טוב אחד של צלחת זכוכית 3-טוב באמצע הדרך עם פתרון טרייה של 9 חלקים PEM מאגר (0.1 M של צינורות pH 6.95, 2 מ מ EGTA, 1 מ מ MgSO4), 1-חלק 10 x PBS ו- % 32 1 חלק מחברים. מקום 3-5 עוברי לתוך המנה ולעקור חור קטן לתוך כל העובר, באמצעות מחט חדה טונגסטן כדי לתקוע. ואחד dulled מעט לייצב, המאפשר את החלמון לזרום החוצה את מקבע לרוץ.
- באמצעות זוג מחטים טונגסטן מחודדים, בעדינות להקניט משם את הקרומים שני החיצוני המקיף את העובר Parhyale . נתח אותם לשבועיים כדי להפוך את העוברים עמידים יותר אבל עבודה במהירות כדי למנוע הקרום להיות מקובע העובר, מה שהופך את הסרת קרום קשה יותר. לאפשר את העוברים לתקן עבור סכום כולל של 15-20 דקות עבור צביעת נוגדן או 40-50 דקות של הכלאה בחיי עיר .
- תמונה הוולדות בשידור חי, לנתח אותם עבור פנוטיפים התנהגותיים מורפולוגיים או לתקן ולא תכתים אותם עבור ניתוח מפורט יותר. הרימו את הוולדות לבגרות מינית תוך 2-3 חודשים כדי ליצור הסתרה וקווים הטרנסגניים (ראה Kontarakis ו- Pavlopoulos75 עבור טיפול אבקוע ופרטים שימושיים אחרים).
8. הערכת עריכה תוצאות
- אם רלוונטי, חפשו הפנוטיפ חזותי או פונקציונלי הערוך תאים או אורגניזמים.
הערה: תהליך זה צפויים להשתנות על-ידי יישום, כמה דוגמאות מתוארים בסוף הפרוטוקול הרלוונטיות שלהם השלבים שלעיל. לאחר תיקון המוטציה חרמשית ב HSPCs, לנתח את ייצור ההמוגלובין ידי erythroblasts הבדיל באמצעות HPLC (איור 1א'). פצצה של הגן לקולטן il-2 בתאי T יכול להיות מאושרות על ידי צביעת משטח, לזרום cytometry (איור 1B). כדי להעריך פנוטיפים C. elegans , פ hawaiensis , להתבונן על התנהגות תחת מיקרוסקופ אור או פלורסנט (דמויות 1C וד' 1) ומורפולוגיה בעלי חיים. - כדי לקבוע את היעילות ואת סוג העריכות גנומית שנוצר, lyse הבריכות של תאים הערוך ולחלץ את הדנ א הגנומי שלהם באמצעות ערכת חילוץ מסחרי21.
-
עבור הערכה מהירה היווצרות ויאסין, PCR-להגביר לפחות 200 זוגות בסיס סביב החתך באתר ולבצע את endonuclease1 T7 assay (T7E1)76 או מודד (נוקלאז צ'ל-1)77.
- אם מערך ויאסין-Cas9-דרך האתר או HDR מוצלח ליצור או להסיר אתר ההגבלה ידוע, שקול להשתמש עיכול אנזים הגבלה כדי להעריך את יעילות העריכה6. הבדיקה פולימורפיזם (RFLP) הגבלת אורך קטע יכול להיות דרך נוחה כדי לבדוק את היעילות אם זה יקרה יהיה זמין.
- עבור כימות מדויק של יעילות העריכה ונחישות של תוצאות העריכה הדומיננטית, שלח את אמפליקון PCR סנגר סטנדרטי רצף עם תחל ויוצאים.
הערה: אם ניתוח של שיבוט יחיד או אורגניזם, הניתוח של תוצאות סאנגר היא פשוטה, כפי שמתואר באיור 2א. אם ניתוח בריכה של תאים, ואז לנתח את chromatograms עם כלי מקוון78, כפי שמוצג באיור 2B. - כימות מלא, רצפים של עריכת תוצאות, לבצע רצף עמוק27,54, כפי שהיא מתוארת באיור 2C.
- כדי להעריך מסוימת של שינויים-יעד, PCR-להגביר האתרים את המטרה החזוי ולשלוח אותם על המיתרים. כדי לאפשר הגילוי של translocations כרומוזומלית, לבצע מדריך-seq79 או תפוקה גבוהה, רוברטסונית ברמת הגנום רצפי (HTGTS)80. לקבלת תמונה מלאה של שינויי העריכה את המטרה בקרב אוכלוסיה המשובטים, לבצע רצף הגנום כולו (WGS)81,82,83.
הערה: ישנם מגוון שיטות, עבור לכימות--את-ויעד הגנום עריכות, הסביר בהמשך בסקירה שונים מאמרים84,85,86.
Representative Results
ניסויים אלה RNP Cas9 איך מראש התאספו הצג יכול לשמש כדי לתפעל את הגנום של תאים ראשוני ואורגניזמים שלם. חוקרים לטהר או לרכוש חלבון Cas9, sgRNA, לשלב שני מרכיביה להקים מראש את המתחם, להציג את RNP לתוך תאים או אורגניזם עניין שלהם. לאחר מתן אפשרות מספיק זמן עבור עריכת להתרחש, כלפי הצאצאים של הדור הבא להיוולד (אם רלוונטי), לבדוק פנוטיפים ו/או לאסוף תאים עבור genotyping. פנוטיפים ייבחנו באמצעות מבחני פונקציונלי, מבחני הבעה, ויזואליזציה (לפי העין או עם מיקרוסקופ) או בשיטות אחרות, בהתאם הניסוי.
לדוגמה, HSPCs שנערכו כדי לתקן את המוטציה β-גלובין שגורם מחלה חרמשית יכול להיות מובחן אריתרוציטים, לבדיקה של הייצור של בריאות או מגל המוגלובין27,87 (איור 1 A). תאי T נערך לדפוק את הגן לקולטן il-2 גבוה-זיקה, CD25 (IL2RA), יכול להיות מנותח על ידי צביעת משטח cytometry זרימה88, וניתח באופן פונקציונלי כדי לזהות תגובה איתות il-2 גירוי (איור 1B ). תאי T שניתן לתכנת גם בדרכים קליניות חשובות רבות שדורשות הערכה של פנוטיפים שונים, כולל את היעילות של זיהום ב- HIV89 , ויוו antitumor היעילות של המכונית-T תאים11.
שימוש בגישה ההקרנה co-CRISPR/co-conversion, בתולעים המוטנטיות נערכים בו זמנית על שני לוקוסים62. HDR-הגן הפניה dpy-10 באמצעות ssODN של תיקון תבנית התוצאה מוטציה רווח-של-הפונקציה הדומיננטית הבקיע בקלות dpy-10 . משפחתית ולא משפחתית הטרוזיגוטיים F1dpy-10(gof) חיות רולר (Rol), homozygous dpy-10(gof) בעלי החיים גוצה (Dpy). הנוכחות של פנוטיפ מציין עריכה Cas9 הזה התרחש החיות הללו ומשפר את הסיכויים לזיהוי אירוע עריכה-מיקומה השני את החיות1 Rol או Dpy F. עריכת ניסוי יירוט מוצלח צריך לגרום 33-50% של מוזרק P0 תולעים מניב הצאצאים1 F 20 או יותר כי הם Rol או Dpy90. אז אפשרי לבחור שלא שבעזרתם חיות כדי לחזור wildtype dpy 10 ולבחור עבור עריכה homozygous עניין. כלל אצבע, ריכוז crRNA מיקוד הגן הפניה co-CRISPR צריך להיות חצי של crRNA פילוח הגן עניין. אם עריכה בגן עניין לא החלים, ניתן להתאים את היחס של RNAs שני CRISPR כדי להגדיל את הסבירות של מחלים את השינוי הרצוי. למשל, להגדיל את כמות crRNA עבור הגן עניין ביחס crRNA גנטי הפניית יגדיל את אחוז תולעים הפו עריכות בגן עניין של תולעים שקיימים גם עריכות-מיקומה ג'ין הפניה בתוך האוכלוסייה. המרה שיתוף תדרים משתנים, אבל המחירים הם בדרך כלל 20-60%, לעיתים קרובות מניב עריכות homozygous דור1 נ (איור 1C).
P. hawaiensis הוולדות שנערכו כדי להמם את הגן Abdominal-B (עבד-B) להציג חריגות מורפולוגיים ברורים3 (איור 1D). גן זה נדרש עבור המתבנת בטן נכונה, תוצאותיו הפרעה מסוג בית החזה קפיצה, הליכה רגליים החלפת הרגליים שחייה ו עוגן כי הם בדרך כלל מציגים על הבטן.
קביעת הגנום עריכה תוצאות ברמת genotypic דורש רצף או assay במבחנה שמזהה רצף השינויים. כאן, אנו מראים נתונים רצף נציג שלנו סוגי תאים דגם, אורגניזמים, הדגשת גישות שונות לעריכת כימות. שימו לב כי התוויות איור הם כללית כי כל השיטות המוצגות כאן ניתן להחיל על כל מערכת ביולוגית.
גישות רצף להשתנות הטכניות המורכבות והעומק של תוצאות. עבור אוכלוסיות הערוך המשובטים או אורגניזמים נפרדים בקלות ספרבילי, יחידים הערוך יכול להיות רציף בעקבות מיצוי הדנ א הגנומי. תקן סנגר רצף תוצאות תחשוף את השינוי רצף באתר Cas9-חתך של אדם נתון, עם frameshifts היפותטי לשבש את תפקידה (איור 2א). כלי מקוון המשמש רצף הוא סנגר המבוססת על רצף גישה אחרת שניתן להחיל אוכלוסיות מעורבות מאשר מוטציות בודדות78. רצפים מנותחים עם כלי מקוון באפשרותך להעריך את יעילות העריכה הכללית, כמו גם תוצאות רצף הדומיננטית. נציג הנתונים מוצגים באיור 2B.
שיטה היסודיות ביותר המתוארות כאן הוא רצף עמוק (המכונה לפעמים רצף תפוקה גבוהה או הדור הבא). שיטה זו מספקת רצפי DNA מן הגנום בודדים בקרב אוכלוסיה מעורבת. נתונים כאלה ניתן לתיאור במגוון דרכים. כאן, אנו יש לסווג רצף בודדים קריאות מתאי הערוך בהתבסס על התוצאה העריכה (איור 2C). רוב התאים נערכים דרך השביל NHEJ, שתוצאתה בדרך כלל שיבוש גנטי. במקרים אחרים, כתקלת את הגן היעד עבור גרסה חלופית באמצעות HDR27.
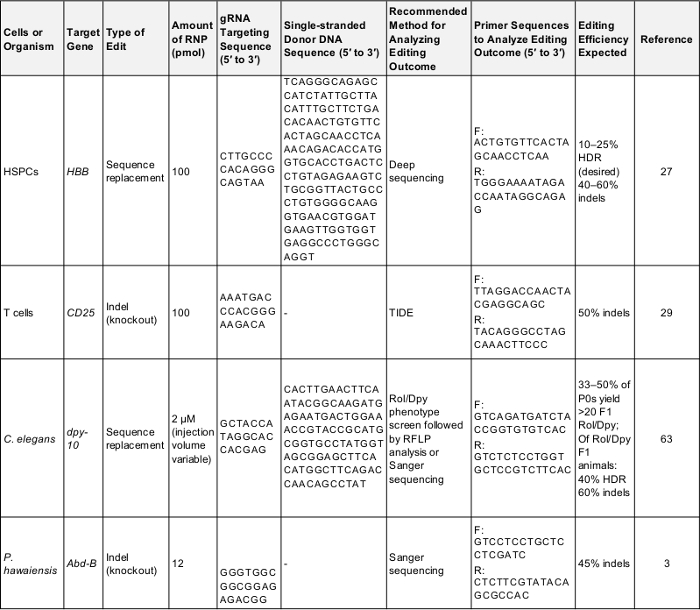
טבלה 1: חיובי שולט על הגנום ראשוני עריכת ניסויים. טבלה זו מציגה את המידע העיקרי הדרושים לביצוע גנום לראשונה עריכת הניסוי בכל אחד של תאים אורגניזמים שמתואר פרוטוקול זה. בעקבות פרמטרים אלה צפוי להניב תוצאה מוצלחת יכול לשמש כדי לבדוק את הפרוטוקול או כקו להשוואה פעם הנסיין מכוון הגן על האינטרס שלהם. F: קדימה, r: הפוך, HDR: תיקון מכוון הומולוגיה. אנא לחץ כאן כדי להוריד את השולחן הזה.

איור 1 : נציג פנוטיפי נובעת Cas9 RNP עריכה של תאים אנושיים ראשוני ואורגניזמים. (א) זה מעקב HPLC להראות את זה אחרי מוצלחת הגנום עריכה, HSPCs זה מובחנים לתוך erythroblasts בשלב מאוחר יהיה לייצר המוגלובין יותר פונקציונלי יותר אנמיה המוגלובין. אריתרוציטים מוטציה לייצר המוגלובין מגל (HbS), ואילו תאים נערך בהצלחה יהיה לייצר המוגלובין בריא (HbA ו HbA2) כמו גם המוגלובין עוברית (HbF). ספיגת בגרף ביחידות שרירותי (au). לוח זה פורסם לראשונה ב DeWitt e.t al... 27. מודפס באישור האגודה האמריקאית לקידום המדע. (B) משמאל, עבור כל תנאי, לוח זה מציג נתונים cytometry זרימה מראה כי שהוכתמו משטח T תאים שאינם מבטאים CD25 אחרי הגן CD25 נוצח עם RNP. שפע CD25 מותווה בציר ה-x עם גודל התא בציר ה-y. מימין, עבור כל תנאי, חלונית זו מציגה כימות פוספו-Stat5 (pStat5) לאחר אינדוקציה עם il-2. האיתות מופחתת כאשר הקולטן il-2 היא נעדרת (CD25 KO). שפע pStat5 מותווה בציר ה-x, הנתונים הנובעים שלוש רמות שונות של il-2 קלט מושווים אנכית. (ג) לוח זה מציג Caenorhabditis elegans co-CRISPR/co-conversion מסך מיקוד dpy-10 כמו דה מרקר ההמרה משותף. 2 מדריך RNAs יעד שני לוקוסים, dpy-10 , ג'ין האהוב שלך (yfg), אותו P0-חיה מוזרק. HDR- dpy-10 תוצאות הפנוטיפ Rol או Dpy. הבחירה של Rol - או Dpy-F1 חיות מגדיל את הסיכויים לזיהוי עריכות-מיקומה השני. (ד) לוח זה מראה שיש wildtype Parhyale hawaiensis הוולדות נורמלי הבטן עם רגליים שחייה ו עוגן. הצאצאים מעלף עבד-B (F0 יחידים) לפתח של הבטן הפכה לכיוון החזה. לפיכך, שחייה והרגליים עוגנים הם נעלם והוחלף הרגליים קופץ והליכה המשויך של החזה נורמלי. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.

איור 2 : תוצאות טיפוסי עריכה שיטות ניתוח התוצאה. (א) לוח זה מציג דוגמאות של סנגר רצף תוצאות בודדות F1 hawaiensis פ אורגניזמים, כולל את רצף wildtype ו- indels שונים 3 לשבש את הפונקציה הגן על ידי הסטת מסגרת קריאה פתוחה. (B) תוצאות אלה גאות להציג את הטווח של הוספות ואירועים מחיקת שהתרחשו באתר Cas9-יעד בבריכה של תאי T ברצף. ציר ה-x מציין את אורך נתון ההוספה או המחיקה של נוקלאוטידים. (ג) תוצאות רצף עמוק אלו מראות ללא גנום עריכה ללא nucleofection או gRNA, מוצלחת עריכה עם שלם Cas9 RNP, המקובצות לפי תיקון ה-DNA התוצאה HSPCs. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
Discussion
הקמת גנום חזקים עריכת פרוטוקול בתא קו או אורגניזם עניין דורש אופטימיזציה, אמפירי בדיקה של פרמטרים מרכזיים, שהוזכר בסעיף זה. מנסה כמה וריאציות של הגישות הכללית המובאת כאן היא מעודדים. המגבלה מפתח של פרוטוקול זה זה יישום שיטות אלה לתאים אחרים או אורגניזמים עלול להוביל לתוצאה אחרת בהתאם למין, למד, עיצוב ניסיוני שמוביל נוקאאוט גנטי יעילות גבוהה לא עשוי לקדם את הדנ א ההכנסה. לפיכך, אנו ממליצים החל השיטות המובאות כאן ופתרון בעיות כמתואר להלן.
פתרון בעיות הגנום עריכה ריאגנט איכות:
יצירת או רכישת ריאגנטים באיכות גבוהה היא שלב קריטי בכל הגנום עריכת פרוטוקול. חלבון Cas9 ניתן לטהר במעבדה או לרכוש באופן מסחרי. פרוטוקולים רבים הערה ריכוז סופי עבור Cas9 מתכונים RNP, אך הגן האופטימלי לעריכת פעילות יהיה תלוי הפעילות הספציפי של כל הכנה חלבון Cas9 בודדים, אשר משתנה בהתאם למקור. ברגע פרוטוקול המוצג כאן הוא עובד, שקול מיטוב כמות RNP להשתמש בה על ידי רמות Cas9 titrating כדי לקבוע ריכוז של האופטימלית: אחד המספק יעד ספציפי מאוד DNA המחשוף בלי מחשוף את המטרה מיותרים הנגרמת על ידי Cas9 מוגזמת40.
מדריך RNA טוהר, הומוגניות ניתן גם גורמים של הגנום עריכה הצלחה22. SgRNAs שנרכשו או רכיבים נפרדים crRNA ו- tracrRNA הם בדרך כלל באיכות גבוהה ריאגנטים, מגוון של שינויים כימיים הינם זמינים כדי להתמודד עם בעיות עם ה-RNA השפלה או הקניית תכונות נוספות ה RNP91. בזמן שעבר שינוי כימי gRNAs לא יהיה צורך הגנום תקן עריכת ניסויים, קבוצות מסוימות מקיימות גבוה בהרבה עריכת יעילות עם ריאגנטים כזה, אז הם עשויים להיות שווה לנסות אחרי מאסטרינג התהליך ו/או כאשר gRNA השפלה נראה22,גיליון91. במבחנה שעתוק ו ג'ל עוקבות טיהור היא חלופה זולה, אשר עשוי להיות מספיק עבור גנום שגרתית עריכת ניסויים17,21,49,50. יתר על כן, מספר גישות בדרך כלל מוחלים כדי לייצר gRNA הומוגנית אוכלוסיות ויוו, לרבות ריבוזים tRNA מבוססות כריתה של קווי עזר, יורחב כדי במבחנה RNA הכנה ליצירת שואב האבק מוצרים92.
מדריך RNA ותורם DNA עיצוב טיפים:
מדריך RNA הבחירה היא גורם קריטי להשגת ביעד יעילים ביותר לעריכת תוך צמצום הסיכויים של חופש-יעד המחשוף. כדי לסייע בבחירה מדריך, מספר מחקרים השתמשו מסכי תפוקה גבוהה בשילוב עם הדור הבא רצפי כדי לקמפל את רצף תכונות של מדריכים מוצלחים47,79,93,94, 95,96. תכונות אלה שימשו לפיתוח אלגוריתמים חזוי של כלים מקוונים כדי לסייע מדריך בחירת44,45,46,47,48. אלגוריתמים כאלה הם מבוססים על המסכים באמצעות מערכות מבוססות DNA עבור מדריך ביטוי RNA. מדריכים מבוטאים באמצעות מקדם Pol III, הביטוי שלהם ולכן הוא נוטה המגבלות הקשורות שעתוק Pol III, כגון סיום מוקדם בעת היתקלות רצועות של אורציל97,98, 99. עם זאת, השימוש RNPs עשה עם במבחנה-מדריך מסונתז RNAs עוקף אותם חששות ומפשטת את האילוצים מדריך עיצוב. תכונה נפוצה הגיח באלגוריתמים אלה, אושרה במחקרים רבים עם הגנום יעילים עריכת, היא הנוכחות של פורין, במיוחד גואנין, בסוף 3′ במטרה רצף של המדריך. תכונה זו מדריך היה מוצלח מאוד בין אורגניזמים ועד יונקים C. elegans, זבובי פירות, דג זברה65,100,101. בנוסף, עבור C. elegans, עיצוב מדריכים dinucleotide GG בסוף 3′ אזור מיקוד של המדריך היא אסטרטגיית אפקטיבי לניבוי מדריך יעיל מאוד RNAs65. באופן אידיאלי, מבחן עזר מרובות במקביל כדי לקבוע אשר הוא המוצלח ביותר עבור יישום מסוים.
כאשר מנסים להציג רצף ה-DNA לתוך הגנום, העיצוב של התורם או תבנית ה-DNA הוא גם מכריע. Oligonucleotide חד-גדילי תורמים (ssODNs) יוכנסו בצורה מהימנה יותר מאשר אחרים תבניות תיקון טיפוסי, כפול גדילי לינארי, פלסמיד דנ א54,55,102. במלון יש לוקוסים, ניתן לשפר יעילות HDR עם ssODNs כי הם משלימים שאינם-היעד או שנעקרו נ להחזיק נשק הומולוגיה כי הם אסימטרי ב אורך27,55. מאז תבנית תיקון להיות מוכנס באתר לחתוך וכוללת את רצף יישוב, צעדים יש לנקוט כדי למנוע Cas9 של ביקוע התורם DNA לפני או אחרי ההוספה גנומית. זו מושגת על ידי הפיכת מוטציות שקטות פאם רצף או זרע באזור, הימנעות ההכרה על-ידי Cas9 תוך שמירה על הפונקציה של21,הגן שנוספו103. למרות שינויים נוקלאוטיד יחיד אפילו פאם נוטים לבטל את איגוד104, לנסות לשנות לפחות ארבעה נוקלאוטידים כדי להיות בטוחים.
משמעות ויישומים עתידיים:
הגנום עריכה עם CRISPR-Cas9 התפתחה שיטה רב עוצמה המאפשר נתיישב מניפולציה גנטית של כל אורגניזם. עריכה RNP Cas9 לוקח קצת יותר מאמץ בהתחלה אבל פשוטה לשימוש ברגע ריאגנטים ופרוטוקולים נקבעו במעבדה. עריכת תאים עם RNP מראש במקום דנ א פלסמיד מוביל גבוה יותר יעילות העריכה הכללי, כולל ההוספה ג'ין קשה להשיג באמצעות HDR, עם פחות תופעות מחוץ-יעד24,25,26 , 27 , 29. עוד יותר, ניסויים למנוע בעיות עם ביטוי גנים, RNA השפלה, קיפול חלבונים ו הקשר בין gRNA לבין מולקולות Cas9 מסונתז בנפרד בתוך תא22,23. עריכת RNP עוקף גם חששות בטיחות מוטגנזה מכוונת insertional ומשמשת ביטוי מתמשכת שעלולות להתעורר כאשר שיטות משלוח ויראלי קלינית14. בגלל יתרונות אלה, מדענים רבים עורכים ניסויים פרה-קליניים, ניסויים הוכחה הרעיון טובה RNP עריכה ליישומים טיפוליים אנושי. הן ויוו לשעבר vivo הגנום מבוסס-RNP העריכה גישות נמצאים בשלבי פיתוח כדי לטפל או לרפא אפילו מגוון תנאים, ממחלות גנטיות כמו דושן ניוון שרירים105 ואנמיה חרמשית27 HIV29 ו סרטן11. מעניין, Cas9 RNP הוא מועסק יותר ויותר כמו שיטת משלוח עבור הנדסה חקלאית, מכיוון שהיא מאפשרת 'ללא ה-DNA' עריכה של צמחים33,34,36.
Disclosures
המחברים אלכסנדר Marson ותירס אי יעקב הם השותפים המייסדים של זרקור הרפוי. יעקב אי תירס הוא יועץ המשימה Therapeutics ו המעבדה שלו קיבלה את תמיכת מחקר ממומן של חברת AstraZeneca, פייזר. אלכסנדר Marson הוא יועץ ג'ונו Therapeutics ו ברית הרפוי, המעבדה שלו קיבלה תמיכת מחקר ממומן של ג'ונו הרפוי Epinomics, סאנופי. המעבדה שלו הוחל גם עבור פטנטים הקשורים בטכנולוגיה Cas9 RNP.
Acknowledgments
אנו מודים רבים מבני הקודם שלנו מעבדות, אזור מפרץ הגנום קהילת על תרומתם לפיתוח שיטות אלה. אנו מודים וילסון רוס אנושות קריאת כתב היד הזה.
המחקר של אלכסנדר Marson נתמך על-ידי מתנה ג'ייק ארונוב ולהעניק חברה טרשת הלאומי (CA 1074-A-21). אלכסנדר Marson מחזיק פרס הקריירה מדענים רפואיים מהקרן Wellcome בורוז, חוקר Biohub צ'אן צוקרברג. המחקר של יעקב אי התירס נתמך על ידי קרן Li קה שינג, מכון מורשת רפואי מחקר רפואי, מכון קליפורניה עבור רפואה רגנרטיבית. Behnom Farboud ומאייר ברברה ג' של המחקר זה מומן בחלקו על ידי המענק NIGMS R01 GM030702 כדי ברברה ג' מאייר, מי הוא חוקר במכון רפואי הווארד יוז. ארין ג'רביס ופאטל Nipam ה של המחקר זה מומן בחלקו על ידי המענק אכ מ IOS-1257379 ומקבל ארין ג'רביס תמיכה של GRFP של ה-NSF מלגת בוגר Philomathia.
Materials
| Name | Company | Catalog Number | Comments |
| Reagents/Materials | |||
| DNA oligonucleotides | Integrated DNA Technologies | - | IDT will provide custom DNA sequences, including those in Table 1 |
| Guide RNAs | Synthego | - | Synthego will provide high-quality sgRNAs for S. pyogenes Cas9, including custom sgRNAs containing the targeting sequences included in Table 1 |
| Purified Cas9 protein (EnGen Cas9 NLS, S. pyogenes) | New England Biosciences | M0646T | If possible, purifying Cas9 in-house or purchasing from local core facilities is a less expensive option |
| Normal peripheral blood CD34+ stem/progenitor cells | AllCells | PB032-2 | |
| StemSpan SFEM | StemCell Technologies | 09650 | |
| StemSpan CC110 | StemCell Technologies | 02697 | |
| P3 Primary Cell 4D-Nucleofector X Kit | Lonza | V4XP-3032 | |
| RPMI-1640 Medium, With sodium bicarbonate, without L-glutamine, liquid | Sigma | R0883-6X500ML | |
| EasySep™ Human T Cell Isolation Kit | Stemcell | 17951 | |
| cell culture plate, 96 wells, round | Fisher Scientific | 3799 | |
| CTS (Cell Therapy Systems) Dynabeads CD3/CD28 | Life Tech | 40203D | |
| Reombinant Human IL-2 | UCSF Pharmacy | NA | |
| SepMate-50 500-pack IVD | Stemcell Technologies | 85460 | |
| OP50 Escherichia coli | Caenorhabditis Genetics Center | OP-50 | https://cgc.umn.edu/ |
| Nematode Growth Media agar in petri dishes | - | - | See Stiernagle, T (ref. 59) |
| Standard borosilicate glass capillaries with filament: 4 in (100 mm), 1/0.58 OD/ID | World Precision Instruments | 1B100F-4 | |
| Single-barrel standard borosilicate glass capillaries: 6 in (152 mm), 2/1.12 OD/ID | World Precision Instruments | 1B200-6 | |
| Cover glass; 24 × 50 mm | Thermo Fisher Scientific | 12-544E | |
| Cover glass; 22 × 22 mm | Thermo Fisher Scientific | 12-518-105K | |
| Apex LE agarose | Genesee Scientific | 20-102 | |
| Halocarbon oil 700 | Sigma-Aldrich | H8898-100ML | |
| pCFJ90 plasmid | Addgene | 19327 | |
| Compressed nitrogen | - | ||
| 60 mM culture dishes | BD | ||
| Capillary tubes with filament: 4 in (1.0 mm) | World Precision Instruments | T2100F-4 | |
| Sylgard 184 | Dow Corning | ||
| Petri dishes (100 × 15 mm) | - | ||
| Tungsten wire (0.005 in. diameter) | Ted Pella | ||
| Perfluoroalkoxy alkane (PFA) | - | ||
| Marine salt | - | ||
| 9" pasteur pipettes | - | ||
| Phenol red | - | ||
| Nuclease-free water | - | ||
| Equipment | |||
| 4D Nucleofector | Lonza | AAF-1002X | |
| MZ75 Stereomicroscope | Leica | Out-of-production. Current model is the M80 Stereomicroscope | |
| Axio Vert35 inverted phase contrast fluorescent microscope | Zeiss | Out-of-production. Current model is the Axio VertA.1 | |
| Laser-based micropipette puller (for C. elegans protocol) | Sutter Instrument | FG-P2000 | |
| Picoliter Microinjector (for C. elegans protocol) | Warner Instruments | PLI-100A | |
| Three-axis Joystick oil hydraulic micromanipulator | Narishige International | MO-202U | |
| Coarse manipulator | Narishige International | MMN-1 | |
| Micropipette puller (for P. hawaiensis protocol) | Sutter Instrument | P-80/PC | |
| Microinjector (for P. hawaiensis protocol) | Narishige | IM300 | |
| Microloader pipette tips | Eppendorf | 5242956003 | |
| NG-agar |
References
- Komor, A. C., Badran, A. H., Liu, D. R. CRISPR-based technologies for the manipulation of eukaryotic genomes. Cell. , 1-17 (2016).
- Barrangou, R., Horvath, P. A decade of discovery: CRISPR functions and applications. Nature Microbiology. 2, 1-9 (2017).
- Martin, A., Serano, J. M., et al. CRISPR/Cas9 mutagenesis reveals versatile roles of Hox genes in crustacean limb specification and evolution. Current Biology. 26 (1), 14-26 (2016).
- Goldstein, B., King, N. The future of cell biology: emerging model organisms. Trends in Cell Biology. 26 (11), 818-824 (2016).
- Mali, P., Yang, L., et al. RNA-guided human genome engineering via Cas9. Science. 339 (6121), 823-826 (2013).
- Cong, L., Ran, F. A., et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 339 (6121), 819-823 (2013).
- Deltcheva, E., Chylinski, K., et al. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature. 471 (7340), 602-607 (2011).
- Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 337 (6096), 816-821 (2012).
- Nowak, C. M., Lawson, S., Zerez, M., Bleris, L. Guide RNA engineering for versatile Cas9 functionality. Nucleic Acids Research. 44 (20), 9555-9564 (2016).
- Jiang, W., Cox, D., Zhang, F., Bikard, D., Marraffini, L. A. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nature Biotechnology. , 1-9 (2013).
- Rupp, L. J., Schumann, K., et al. CRISPR/Cas9-mediated PD-1 disruption enhances anti-tumor efficacy of human chimeric antigen receptor T cells. Scientific Reports. 7 (1), 737 (2017).
- Graham, D. B., Root, D. E. Resources for the design of CRISPR gene editing experiments. Genome Biology. 16, 260 (2015).
- Wang, H., La Russa, M., Qi, L. S. CRISPR/Cas9 in genome editing and beyond. Annual Review of Biochemistry. 85, 2270 (2016).
- Nelson, C. E., Gersbach, C. A. Engineering delivery vehicles for genome editing. Annual Review of Chemical and Biomolecular Engineering. 7, 637-662 (2016).
- Yin, H., Kauffman, K. J., Anderson, D. G. Delivery technologies for genome editing. Nature Reviews Drug Discovery. 16 (6), 387-399 (2017).
- Zuris, J. A., Thompson, D. B., et al. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nature Biotechnology. 33 (1), 73-80 (2015).
- Cho, S. W., Lee, J., Carroll, D., Kim, J. -S., Lee, J. Heritable gene knockout in Caenorhabditis elegans by direct injection of Cas9-sgRNA ribonucleoproteins. Genetics. 195 (3), 1177-1180 (2013).
- Wang, W., Kutny, P. M., et al. Delivery of Cas9 protein into mouse zygotes through a series of electroporation dramatically increases the efficiency of model creation. Journal of Genetics and Genomics. 43 (5), 319-327 (2016).
- Chen, S., Lee, B., Lee, A. Y. -F., Modzelewski, A. J., He, L. Highly efficient mouse genome editing by CRISPR ribonucleoprotein electroporation of zygotes. Journal of Biological Chemistry. 291 (28), 14457-14467 (2016).
- Remy, S., Chenouard, V., et al. Generation of gene-edited rats by delivery of CRISPR/Cas9 protein and donor DNA into intact zygotes using electroporation. Scientific Reports. 7 (1), 16554 (2017).
- DeWitt, M. A., Corn, J. E., Carroll, D. Genome editing via delivery of Cas9 ribonucleoprotein. Methods. , 1-7 (2017).
- Hendel, A., Bak, R. O., et al. Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells. Nature Biotechnology. 33 (9), 985-989 (2015).
- Thyme, S. B., Akhmetova, L., Montague, T. G., Valen, E., Schier, A. F. Internal guide RNA interactions interfere with Cas9-mediated cleavage. Nature Communications. 7, 11750 (2016).
- Kim, S., Kim, D., Cho, S. W., Kim, J., Kim, J. -S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Research. 24 (6), 1012-1019 (2014).
- Lin, S., Staahl, B. T., Alla, R. K., Doudna, J. A. Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery. eLife. 3, 04766 (2014).
- Liang, X., Potter, J., et al. Rapid and highly efficient mammalian cell engineering via Cas9 protein transfection. Journal of Biotechnology. 208, 44-53 (2015).
- DeWitt, M. A., Magis, W., Bray, N. L., Wang, T. Selection-free genome editing of the sickle mutation in human adult hematopoietic stem/progenitor cells. Science Translational Medicine. 8 (360), (2016).
- Ramakrishna, S., Kwaku Dad, A. -B., Beloor, J., Gopalappa, R., Lee, S. -K., Kim, H. Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA. Genome Research. 24 (6), 1020-1027 (2014).
- Schumann, K., Lin, S., et al. Generation of knock-in primary human T cells using Cas9 ribonucleoproteins. Proceedings of the National Academy of Sciences of the United States of America. 112 (33), 10437-10442 (2015).
- Lee, J. -S., Kwak, S. -J., et al. RNA-guided genome editing in Drosophila with the purified Cas9 protein. G3: Genes, Genomes, Genetics (Bethesda, MD). 4 (7), 1291-1295 (2014).
- Sung, Y. H., Kim, J. M., et al. Highly efficient gene knockout in mice and zebrafish with RNA-guided endonucleases. Genome Research. 24 (1), 125-131 (2014).
- Menoret, S., De Cian, A., et al. Homology-directed repair in rodent zygotes using Cas9 and TALEN engineered proteins. Scientific Reports. 5, 14410 (2015).
- Woo, J. W., Kim, J., et al. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nature Biotechnology. 33 (11), 1162-1164 (2015).
- Malnoy, M., Viola, R., et al. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Frontiers in Plant Science. 7, 1904 (2016).
- Svitashev, S., Schwartz, C., Lenderts, B., Young, J. K., Mark Cigan, A. Genome editing in maize directed by CRISPR-Cas9 ribonucleoprotein complexes. Nature Communications. 7, 13274 (2016).
- Liang, Z., Chen, K., et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nature Communications. 8, 14261 (2017).
- Shin, S. -E., Lim, J. -M., et al. CRISPR/Cas9-induced knockout and knock-in mutations in Chlamydomonas reinhardtii. Scientific Reports. 6, 27810 (2016).
- Pohl, C., Kiel, J. A. K. W., Driessen, A. J. M., Bovenberg, R. A. L., Nygård, Y. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synthetic Biology. 5 (7), 754-764 (2016).
- Grahl, N., Demers, E. G., Crocker, A. W., Hogan, D. A. Use of RNA-protein complexes for genome editing in non-albicans Candida species. mSphere. 2 (3), (2017).
- Rivera-Torres, N., Kmiec, E. B. A standard methodology to examine on-site mutagenicity as a function of point mutation repair catalyzed by CRISPR/Cas9 and ssODN in human cells. Journal of Visualized Experiments. (126), (2017).
- Nandal, A., Mallon, B., Telugu, B. P. Efficient generation and editing of feeder-free IPSCs from human pancreatic cells using the CRISPR-Cas9 system. Journal of Visualized Experiments. (129), (2017).
- Mohr, S. E., Hu, Y., Ewen-Campen, B., Housden, B. E., Viswanatha, R., Perrimon, N. CRISPR guide RNA design for research applications. The FEBS Journal. 283 (17), 3232-3238 (2016).
- Bauer, D. E., Canver, M. C., Orkin, S. H. Generation of genomic deletions in mammalian cell lines via CRISPR/Cas9. Journal of Visualized Experiments. (95), e52118 (2015).
- Hsu, P. D., Scott, D. A., et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nature Biotechnology. 31 (9), 827-832 (2013).
- Heigwer, F., Kerr, G., Boutros, M. E-CRISP: fast CRISPR target site identification. Nature Methods. 11 (2), 122-123 (2014).
- Moreno-Mateos, M. A., Vejnar, C. E., et al. CRISPRscan: designing highly efficient sgRNAs for CRISPR-Cas9 targeting in vivo. Nature Methods. 12 (10), 982-988 (2015).
- Labun, K., Montague, T. G., Gagnon, J. A., Thyme, S. B., Valen, E. CHOPCHOP v2: a web tool for the next generation of CRISPR genome engineering. Nucleic Acids Research. 44, 272-276 (2016).
- Haeussler, M., Schönig, K., et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biology. 17 (1), 148 (2016).
- Lo, T. -W., Pickle, C. S., et al. Precise and heritable genome editing in evolutionarily diverse nematodes using TALENs and CRISPR/Cas9 to engineer insertions and deletions. Genetics. 195 (2), 331-348 (2013).
- Bassett, A., Liu, J. -L. CRISPR/Cas9 mediated genome engineering in Drosophila. Methods. 69 (2), 128-136 (2014).
- Prior, H., Jawad, A. K., MacConnachie, L., Beg, A. A. Highly efficient, rapid and co-CRISPR independent genome editing in Caenorhabditis elegans. G3: Genes, Genomes, Genetics. , Bethesda, MD. (2017).
- Hirsh, A. Cas9 expression and purification protocol. protocols.io. , (2017).
- DeWitt, M. A., Wong, J. In vitro transcription of guide RNAs. protocols.io. , (2017).
- Yang, L., Guell, M., et al. Optimization of scarless human stem cell genome editing. Nucleic Acids Research. 41 (19), 9049-9061 (2013).
- Richardson, C. D., Ray, G. J., DeWitt, M. A., Curie, G. L., Corn, J. E. Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA. Nature Biotechnology. 34 (3), 339-344 (2016).
- Paix, A., Folkmann, A., Seydoux, G. Precision genome editing using CRISPR-Cas9 and linear repair templates in C. elegans. Methods. 121-122, 86-93 (2017).
- Mello, C., Fire, A. DNA transformation. Methods in Cell Biology. 48, 451-482 (1995).
- Sutter Pipette Cookbook. , Available from: https://www.sutter.com/PDFs/pipette_cookbook.pdf (2017).
- Stiernagle, T. Maintenance of C. elegans. WormBook: the online review of C. elegans biology. , (2006).
- Evans, T. C. Transformation and microinjection. WormBook: the online review of C. elegans biology. , (2006).
- Berkowitz, L. A., Knight, A. L., Caldwell, G. A., Caldwell, K. A. Generation of stable transgenic C. elegans using microinjection. Journal of Visualized Experiments. (18), (2008).
- Kim, H., Ishidate, T., et al. A co-CRISPR strategy for efficient genome editing in Caenorhabditis elegans. Genetics. 197 (4), 1069-1080 (2014).
- Arribere, J. A., Bell, R. T., Fu, B. X. H., Artiles, K. L., Hartman, P. S., Fire, A. Z. Efficient marker-free recovery of custom genetic modifications with CRISPR/Cas9 in Caenorhabditis elegans. Genetics. 198 (3), 837-846 (2014).
- Ward, J. D. Rapid and precise engineering of the Caenorhabditis elegans genome with lethal mutation co-conversion and inactivation of NHEJ repair. Genetics. 199 (2), 363-377 (2015).
- Farboud, B., Meyer, B. J. Dramatic enhancement of genome editing by CRISPR/Cas9 through improved guide RNA design. Genetics. 199 (4), 959-971 (2015).
- Wood, A. J., Lo, T. -W., et al. Targeted genome editing across species using ZFNs and TALENs. Science. 333 (6040), 307 (2011).
- Friedland, A. E., Tzur, Y. B., Esvelt, K. M., Colaiácovo, M. P., Church, G. M., Calarco, J. A. Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nature Methods. 10 (8), 741-743 (2013).
- Dickinson, D. J., Ward, J. D., Reiner, D. J., Goldstein, B. Engineering the Caenorhabditis elegans genome using Cas9-triggered homologous recombination. Nature Methods. 10 (10), 1028-1034 (2013).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Injection of Parhyale hawaiensis blastomeres with fluorescently labeled tracers. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Gerberding, M., Browne, W. E., Patel, N. H. Cell lineage analysis of the amphipod crustacean Parhyale hawaiensis reveals an early restriction of cell fates. Development (Cambridge, England). 129 (24), 5789-5801 (2002).
- Browne, W. E., Price, A. L., Gerberding, M., Patel, N. H. Stages of embryonic development in the amphipod crustacean, Parhyale hawaiensis. Genesis. 42 (3), New York, NY. 124-149 (2005).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Fixation and dissection of Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. In situ hybridization of labeled RNA probes to fixed Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Antibody staining of Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Kontarakis, Z., Pavlopoulos, A. Transgenesis in non-model organisms: the case of Parhyale. Methods in Molecular Biology. 1196, Clifton, NJ. 145-181 (2014).
- Kim, H. J., Lee, H. J., Kim, H., Cho, S. W., Kim, J. -S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Research. 19 (7), 1279-1288 (2009).
- Qiu, P., Shandilya, H., D'Alessio, J. M., O'Connor, K., Durocher, J., Gerard, G. F. Mutation detection using Surveyor nuclease. BioTechniques. 36 (4), 702-707 (2004).
- Brinkman, E. K., Chen, T., Amendola, M., van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Research. 42 (22), 168 (2014).
- Tsai, S. Q., Zheng, Z., et al. GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases. Nature Biotechnology. 33 (2), 187-197 (2015).
- Frock, R. L., Hu, J., Meyers, R. M., Ho, Y. -J., Kii, E., Alt, F. W. Genome-wide detection of DNA double-stranded breaks induced by engineered nucleases. Nature Biotechnology. 33 (2), 179-186 (2015).
- Smith, C., Gore, A., et al. Whole-genome sequencing analysis reveals high specificity of CRISPR/Cas9 and TALEN-based genome editing in human iPSCs. Cell Stem Cell. 15 (1), 12-13 (2014).
- Veres, A., Gosis, B. S., et al. Low incidence of off-target mutations in individual CRISPR-Cas9 and TALEN targeted human stem cell clones detected by whole-genome sequencing. Cell Stem Cell. 15 (1), 27-30 (2014).
- Kim, D., Bae, S., et al. Digenome-seq: genome-wide profiling of CRISPR-Cas9 off-target effects in human cells. Nature Methods. 12 (3), 237-243 (2015).
- Hendel, A., Fine, E. J., Bao, G., Porteus, M. H. Quantifying on- and off-target genome editing. Trends in Biotechnology. 33 (2), 132-140 (2015).
- O'Geen, H., Yu, A. S., Segal, D. J. How specific is CRISPR/Cas9 really. Current Opinion in Chemical Biology. 29, 72-78 (2015).
- Tsai, S. Q., Joung, J. K. Defining and improving the genome-wide specificities of CRISPR-Cas9 nucleases. Nature Reviews Genetics. 17 (5), 300-312 (2016).
- Hoban, M. D., Cost, G. J., et al. Correction of the sickle cell disease mutation in human hematopoietic stem/progenitor cells. Blood. 125 (17), 2597-2604 (2015).
- Simeonov, D. R., Gowen, B. G., et al. Discovery of stimulation-responsive immune enhancers with CRISPR activation. Nature. , (2017).
- Hultquist, J. F., Schumann, K., et al. A Cas9 ribonucleoprotein platform for functional genetic studies of HIV-host interactions in primary human T cells. Cell Reports. 17 (5), 1438-1452 (2016).
- Paix, A., Wang, Y., et al. Scalable and versatile genome editing using linear DNAs with microhomology to Cas9 sites in Caenorhabditis elegans. Genetics. 198 (4), 1347-1356 (2014).
- Lee, K., Mackley, V. A., et al. Synthetically modified guide RNA and donor DNA are a versatile platform for CRISPR-Cas9 engineering. eLife. 6, (2017).
- Minkenberg, B., Wheatley, M., Yang, Y. CRISPR/Cas9-enabled multiplex genome editing and its application. Progress in Molecular Biology and Translational Science. 149, 111-132 (2017).
- Doench, J. G., Fusi, N., et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nature Biotechnology. 34 (2), 184-191 (2016).
- Doench, J. G., Hartenian, E., et al. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nature Biotechnology. , 1-8 (2014).
- Liu, H., Wei, Z., Dominguez, A., Li, Y., Wang, X., Qi, L. S. CRISPR-ERA: a comprehensive design tool for CRISPR-mediated gene editing, repression and activation. Bioinformatics (Oxford, England). 31 (22), 3676-3678 (2015).
- Wu, X., Scott, D. A., et al. Genome-wide binding of the CRISPR endonuclease Cas9 in mammalian cells. Nature Biotechnology. 32 (7), 670-676 (2014).
- Bogenhagen, D. F., Brown, D. D. Nucleotide sequences in Xenopus 5S DNA required for transcription termination. Cell. 24 (1), 261-270 (1981).
- Cozzarelli, N. R., Gerrard, S. P., Schlissel, M., Brown, D. D., Bogenhagen, D. F. Purified RNA polymerase III accurately and efficiently terminates transcription of 5S RNA genes. Cell. 34 (3), 829-835 (1983).
- Chen, B., Gilbert, L. A., et al. Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system. Cell. 155 (7), 1479-1491 (2013).
- Gagnon, J. A., Valen, E., et al. Efficient mutagenesis by Cas9 protein-mediated oligonucleotide insertion and large-scale assessment of single-guide RNAs. PLoS ONE. 9 (5), 98186 (2014).
- Ren, X., Yang, Z., et al. Enhanced specificity and efficiency of the CRISPR/Cas9 system with optimized sgRNA parameters in Drosophila. Cell Reports. 9 (3), 1151-1162 (2014).
- Ran, F. A., Hsu, P. D., Wright, J., Agarwala, V., Scott, D. A., Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nature Protocols. 8 (11), 2281-2308 (2013).
- Serano, J. M., Martin, A., et al. Comprehensive analysis of Hox gene expression in the amphipod crustacean Parhyale hawaiensis. Developmental Biology. 409 (1), 297-309 (2016).
- Sternberg, S. H., Redding, S., Jinek, M., Greene, E. C., Doudna, J. A. DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Nature. , 1-17 (2014).
- Lee, K., Conboy, M., et al. Nanoparticle delivery of Cas9 ribonucleoprotein and donor DNA in vivo induces homology-directed DNA repair. Nature Biomedical Engineering. 1 (11), 889-901 (2017).
