

Summary
Forurening under den genomisk sekventering af mikroskopiske organismer er fortsat et stort problem. Her viser vi en metode til sequence genomet af et tardigrade fra et enkelt eksemplar med så lidt som 50 pg genomisk DNA uden hele genom forstærkning til at minimere risikoen for kontaminering.
Abstract
De er mikroskopiske dyr, som angiver en ametabolic stat kaldet anhydrobiosis når man står overfor udtørring og kan vende tilbage til deres oprindelige tilstand, når vand er leveret. Genomisk Sekventeringen af mikroskopiske dyr som Biogeografi risici bakteriel forurening, der nogle gange fører til fejlagtige fortolkninger, for eksempel med hensyn til omfanget af horisontal genoverførsel i disse dyr. Her, leverer vi en ultralow input metode for at sekvens genomet af tardigrade, Hypsibius dujardini, fra et enkelt eksemplar. Ved at ansætte grundig vask og forurenende udstødelse sammen med en effektiv udvinding af 50 ~ 200 pg genomisk DNA fra et enkelt individ, vi bygget et bibliotek sekventeret med en DNA-sekventering instrument. Disse biblioteker var meget reproducerbare og upartiske, og informatik analyse af den omkostningsstyring læser med andre H. dujardini genomer viste en minimal mængde af forurening. Denne metode kan anvendes til unculturable biogeografi, der ikke kunne blive sekventeret tidligere metoder.
Introduction
De er mikroskopiske dyr, der kan angive en ametabolic stat kaldet anhydrobiosis, når man står overfor udtørring. De inddrive ved absorption af vand1,2. I ametabolic staten er de i stand til at tolerere forskellige ekstreme miljøer, der omfatter ekstreme temperaturer3 og pres4,5, en høj dosis af ultraviolet lys6, røntgen og gammastråler 7 , 8, og kosmisk plads9. Genomisk data er et uundværligt grundlag for studiet af molekylære mekanismer af anhydrobiosis.
Tidligere forsøg på at sekvens Biogeografi genom har vist tegn på bakteriel forurening10,11,12,13,14. Genomisk sekventering af sådanne små organismer kræver et stort antal dyr og er liggende hen til bakteriel forurening; Derfor har vi tidligere fastsat en sekventering protokollen ved hjælp af en ultralow inddata metode med udgangspunkt i et enkelt eksemplar af tardigrade, for at minimere risikoen for forureninger15. Ved hjælp af disse data, har vi yderligere gennemført en høj kvalitet resequencing og gensamling af genomet af H. dujardini16,17. Her vi beskriver i detaljer denne metode for genomisk sekventering fra en enkelt tardigrade individuelle ()figur 1). Valideringen af denne sekventering metode er ud over fokus i dette papir og allerede er blevet grundigt drøftet i vores tidligere rapport16.
Denne metode består af to dele: isolering af en enkelt tardigrade med den laveste forurening mulige og høj kvalitet udvinding af piktogram niveauer af DNA. Tardigrade er udsultet og skylles grundigt med vand, som antibiotika og observeret under et mikroskop med 500 X forstørrelse at sikre fjernelse af enhver bakteriel forurening. Tidligere skøn og målinger viser, at et enkelt individ af tardigrade indeholder cirka 50-200 pg genomisk DNA16, som er udvundet af sprængning chitin exoskeleton af fryse-tø cykler eller ved manuel homogenisering. Denne genomisk DNA er indsendt til bibliotek opbygning og sekventeret på en DNA-sekventering instrument. En yderligere Informatik analyse viser høj kvalitet sekventering, samt lave niveauer for forurening i forhold til tidligere tardigrade sekventering projekter.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. forberedelse
- Forberede 2%-agarosegel med destilleret vand (DW) som opløsningsmiddel i en 90 mm plast kultur parabol, og 10 mL af en 1% penicillin/streptomycin cocktail med DW. Gelen kan opbevares i 2-3 uger i en inkubator, indstillet på 18 ° C.
Bemærk: Undgå enhver oplagring af gel under 10 ° C, til den lave temperatur vil skrumpe agarosegel, hvilket resulterer i en ganske lille kløft mellem gel og kultur parabol væggen hvor Biogeografi kan blive fanget.
2. prøve forberedelse og forurenende udelukkelse
- Indsamle en enkelt tardigrade, placere den på den forberedte agar plade og vask det 2 x - 3 x med DW at fjerne enhver resterende partikler.
- Inkubér tardigrade på 18-22 ° C i 24 timer til at fjerne eventuelle overskydende mad fra tarmene.
- Placer den sultet tardigrade i penicillin/streptomycin antibiotika for 2-6 h at fjerne enhver bakteriel forurening og placere dekontamineret dyret på en ren dias glas ved hjælp af en P10 pipette.
- Observere tardigrade under et mikroskop på 500 X Forstørrelse og bekræfte, at der er ingen resterende bakterier.
Bemærk: Den høj forstørrelse med et stereomikroskop er optimal, men et optisk mikroskop kan anvendes alternativt uden anvendelse af en coverslip. - Indsamle den enkelte bruger P10 pipette med et maksimum på 5 µL af væske, læg det i en lav-bindende polymerase kædereaktion (PCR) rør og fjerne som meget overskydende væske som muligt.
Bemærk: Microtubes, PCR rør og pipette tips bør alle være lav bindende at minimere tabet af DNA.
Bemærk: Gennemføre en homogenisering så hurtigt som muligt for hurtig udtørring eller død af dyret kan væsentligt skade genomisk DNA.
3. homogenisering og DNA-ekstraktion
- Homogeniseres dyret for at få sin genomisk DNA (gDNA) med en af følgende metoder.
Bemærk: Det er afgørende for at bruge den angivne kit vist i Tabel af materialer i de følgende DNA udvinding skridt, for andre kits (enten kolonne og perle baseret) ikke er effektive på dette ekstremt lave input. Genomisk lysisbuffer har været leveret med 0,5% beta-mercaptoethanol før brug.- Homogeniseres dyr med fryse-tø cykler18.
- Umiddelbart efter trin 2.5, tilføje 100 µL af lysisbuffer til PCR rør indeholdende tardigrade.
- PCR-røret anbringes i flydende kvælstof i 10 min og i 10 min, flytte den til en varme blok varmet til 37 ° C. Gentag dette trin 2 x.
Bemærk: Dette trin kan gentages, når homogenisering er ikke tilstrækkeligt eller kan udføres efter manuel knusning.
- Manuelt knuse dyret.
- Umiddelbart efter trin 2.5, under et stereomikroskop, knuse enkelt med P10 spidsen af pipetten ved at trykke på dyret mod PCR rør væg, og straks tilføje 100 µL af lysisbuffer.
Bemærk: Det er afgørende at overholde denne procedure under et stereomikroskop, fordi tardigrade kan nemt glide væk, og ineffektive knusning vil resultere i en fejl til at udtrække gDNA. Sørg for, at tardigrade kutikula er brudt, så lysisbuffer kan infiltrere organismen.
- Umiddelbart efter trin 2.5, under et stereomikroskop, knuse enkelt med P10 spidsen af pipetten ved at trykke på dyret mod PCR rør væg, og straks tilføje 100 µL af lysisbuffer.
- Homogeniseres dyr med fryse-tø cykler18.
- Inkuber røret i 30 min. ved stuetemperatur for lysis at forekomme.
Bemærk: Mindst 30 min er nødvendig for en effektiv lysis, men inkubation kan være længere. - Overføre den komplette volumen (100 µL) af lysis blandingen til en ren 1,5 mL lav-bindende microtube.
- Tilføje 100 µL af lysisbuffer til lav-bindende PCR rør, der blev brugt til homogenisering og er nu tomme, og efter pipettering blandingen, overføre den til 1,5 mL lav-bindende microtube bruges i trin 3.3. Gentag dette trin 2 x.
- I trin 3.4, tilføje 300 µL af lysisbuffer til lav-bindende PCR rør og efter pipettering, flytte blandingen til 1,5 mL lav-bindende microtube. 1,5 mL lav-bindende microtube bør indeholde 600 µL af blandingen efter dette trin.
Bemærk: Disse trin er at minimere tabet af gDNA bundet til rørvæggen PCR ved at vaske prøven flere gange. - Tilføje i alt 600 µL af lysis blanding til kolonnen spin placeret i en collection tube og centrifugeres det på 10.000 x g i 1 minut.
- Igen gælder flow gennem kolonnen og centrifugeres det på 10.000 x g i 1 minut.
Bemærk: Dette trin er afgørende for at sikre, at de fleste af gDNA er bundet til kolonnen. - Tilføje 500 µL af wash buffer til kolonnen spin og centrifugeres det på 10.000 x g i 1 min. overførsel kolonnen spin til en ren 1,5 mL microtube.
- Anvende 20 µL af 10 mM Tris-HCl, pH 8,5, at kolonnen spin, vente i 5 min ved stuetemperatur, og centrifugeres det på 10.000 x g i 1 minut.
Bemærk: Eluering buffer må ikke indeholder EDTA, for det griber ind i biblioteket forberedelse enzymer. - Re-andrage flow gennem kolonnen spin, og efter 5 min inkubation ved stuetemperatur, centrifugeres i 1 minut ved 10.000 x g.
Bemærk: Dette trin er afgørende for at sikre maksimal eluering af gDNA bundet til kolonnen.
4. biblioteket konstruktion sekvens
-
DNA opsplitning
- Overføre 15 µL af gDNA eluant til en 15-µL microtube for DNA fragmentering og centrifugeres røret i 1 minut ved hjælp af en bordplade microcentrifuge.
Bemærk: Den angivne microtube vist i Tabel af materialer er optimal for en lav input. Fragmentering med lav-volumen sonikering er kritisk, og dette kan ikke erstattes med enzymatisk fragmentering på grund af den ekstremt lav koncentration af DNA. - Fragment gDNA til 550 bp.
Bemærk: Indstillingerne 550 bp brugte vi er som følger: hændelse spidseffekt = 30 W, duty faktor = 20%, cyklusser pr. burst = 50, indvirkningstid = 23 s. - Efter en grundig pipettering, overføre 10 µL af fragmenterede DNA blandingen til en ren lav-bindende PCR rør.
Bemærk: Eksperimentet kan standses her. Bevare DNA ved 4 ° C eller -20 ° C.
- Overføre 15 µL af gDNA eluant til en 15-µL microtube for DNA fragmentering og centrifugeres røret i 1 minut ved hjælp af en bordplade microcentrifuge.
-
Biblioteket konstruktion sekvens
Bemærk: Det er helt afgørende at bruge den angivne kit i Tabel af materialer i de følgende procedurer, på grund af det lave antal input DNA.- Tilsæt 2 µL af skabelon forberedelse buffer og 1 µL af skabelon forberedelse enzym og bland dem grundigt med en pipette.
- Udføre skabelon forberedelse reaktion på en termisk cycler med følgende betingelser: 22 ° C i 25 min, 55 ° C i 20 min., 4 ° C hold og opvarmet låg på 101-105 ° C. Når reaktionen er afsluttet, gå videre til næste trin.
- Tilføje 1 µL af bibliotek syntese buffer og 1 µL af bibliotek syntese enzym til reaktionsprodukt skabelon forberedelse og inkuberes blanding ved 22 ° C i 40 min (med et hold for 4 ° C). Når reaktionen er afsluttet, gå videre til næste trin.
- Tilføj 30 µL af bibliotek forstærkning master mix (25 µL af bibliotek forstærkning buffer, 1 µL af bibliotek forstærkning enzym og 4 µL nukleasen-gratis vand) og 5 µL af den indekserer reagens.
- Bland det hele grundigt med en pipette og centrifugeres blandingen kort med en bordplade centrifuge.
- Udføre en PCR med de betingelser, der er præsenteret i tabel 1.
| Temperatur | Tid | Cykler |
| 72 ˚C | 3 minutter | |
| 85 ˚C | 2 minutter | |
| 98 ˚C | 2 minutter | |
| 98 ˚C | 20 sekunder | 4 cyklusser |
| 67 ˚C | 20 sekunder | |
| 72 ˚C | 40 sekunder | |
| 98 ˚C | 20 sekunder | 16 cykler |
| 72 ˚C | 50 sekunder | |
| 4 ˚C | Hold |
Tabel 1: PCR betingelser.
-
Oprensning af PCR produkter
Bemærk: Dette trin kan erstattes med andre metoder, rensning. Hvis der benyttes en alternativ metode, Sørg for at tjekke for den resulterende DNA renhed ved hjælp af et spektrofotometer. Absorbansen nøgletal 260/280 og 260/230 skal både være over 1,8.- Tilsæt 50 µL af magnetiske perler, afpipetteres løsning 10 x, og centrifugeres det kort med en bordplade microcentrifuge.
- Inkuber løsning for 2 min ved stuetemperatur.
- Inkuber det på en magnetisk stå i 5 min eller indtil løsningen bliver helt klart, og Fjern supernatanten.
- Tilføje 200 µL af frisklavede 80% ethanol til lav-bindende PCR rør på en magnet stå, vente 30 s, og Fjern supernatanten. Gentag dette trin 2 x. Forstyr ikke perlerne.
- Kort centrifugeres lav-bindende PCR rør med en bordplade centrifuge og fjerne eventuelle overskydende ethanol på den magnetiske stand. Lufttørre perlerne men undgå overtørring.
- Resuspend perler med 15 µL af 10 mM Tris-HCl, pH 8,5, grundigt afpipetteres løsningen, således at de magnetiske perler er ensartet fordelt, inkuberes rør i 2 min. ved stuetemperatur, og efter en kort centrifugering, inkuberes det på en magnetisk står for 2 min indtil løsningen er klar.
Bemærk: Eluering buffer bør ikke indeholder EDTA, for det griber ind i sekventering kemi. - Overføre supernatanten, uden at forstyrre pelleten, til en ny lav-bindende PCR rør.
Bemærk: Eksperimentet kan standses her. Bevare DNA ved 4 ° C eller ved-20 ° C i lang tid opbevaring.
5. kvalitet Check, kvantificering og sekvens af DNA
Bemærk: En kvalitetskontrol er ikke gennemført forud for dette trin på grund af det lave antal DNA.
-
Validering af DNA bibliotek størrelse distribution
Bemærk: Andre høj-følsomme elektroforese systemer med en digital indberetning af størrelse distribution kan bruges.- Returnere elektroforese buffer reagens og gel patron (25-1.000 bp i range) til stuetemperatur.
- Tilføje 3 µL af elektroforese buffer reagens med 1 µL af sekventering bibliotek og bland dem grundigt i 1 min. med en vortex, og kortvarigt centrifugeres dem med en bordplade centrifuge.
- Gennemføre elektroforese og validere bibliotek størrelse distribution med det tilknyttede programmel. Vigtigste fragment peak bør bredt spænder fra omkring 300 til 1.000 bp.
-
DNA kvantificering
Bemærk: Andre fluorescens-baserede metoder eller kvantitativ PCR kan også bruges, men spektrofotometri bør undgås, da det ikke er præcis nok for kvantificering sekventering biblioteker.- Tilføj 796 µL af løsning buffer og 4 µL af fluorescerende reagens og bland dem grundigt. Undvære 190 µL af brugsopløsning til to assay rør og 197 µL til en assay tube.
- Tilsæt 10 µL standarder med kendte koncentrationer af DNA til hvert assay rør indeholdende 190 µL af brugsopløsning og 3 µL af biblioteket parat til assay rør indeholdende 197 µL af brugsopløsning.
- Vortex rør kort og centrifugeres dem på en bordplade centrifuge. Kvantificere DNA ved hjælp af en fluorometer med 3-µL indstillinger.
-
DNA bibliotek sekvens
Bemærk: Sekventering platform skal være kompatibel med den angivne bibliotek byggesæt.- Forberede sekventering bibliotek baseret på producentens protokol.
- Sæt reagens kassette og flow celle til sekvensering instrument og Indtast sekventering køre oplysninger efter producentens protokol.
- Køre Sekventeringen.
Bemærk: Vi har gennemført to sekventering kører: en prøve/køre, samt fire prøver multipleksede i en køre.
6. beregningsmæssige analyse
- Base-opkald og eventuelt afmultiplekse læser.
- Validere kvaliteten af sekvens data med FastQC19.
Bemærk: For en mere dybdegående validering af de fremkomne data, se vores forrige rapport16.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Forurenende udelukkelse:
Denne protokol omfatter en grundig vask af tardigrade og en sterilisation med antibiotika behandling at minimere forurening. Det indebærer også en visuel kontrol proces for at sikre fuldstændigheden af disse processer. Et mikroskop billede under validering (trin 2.4 i protokollen) er vist i figur 2. Når observeret ved 500 X forstørrelse, kan bakterieceller ses som små partikler, der bevæger sig omkring de enkelte tardigrade.
Validering af DNA bibliotek kvalitet:
Det samlede beløb for den beregnede DNA-Seq bibliotek er ca 109.5 ng (7,3 ng/µL x 15 µL)16. For at validere længde fordelingen af fragmenteringen, en elektroforese mønster bør svare til figur 3. Som vi sætter fragmentering størrelse til 550 bp med en DNA klipning system, biblioteket bør være 550-600 bp, herunder sekventering adaptere. Det kan bemærkes, at størstedelen af biblioteket sekvens er indeholdt mellem 200-1000 bp og er konsekvent mellem flergangsbestemmelser (N1 - N4).
Sekvens dataanalyse:
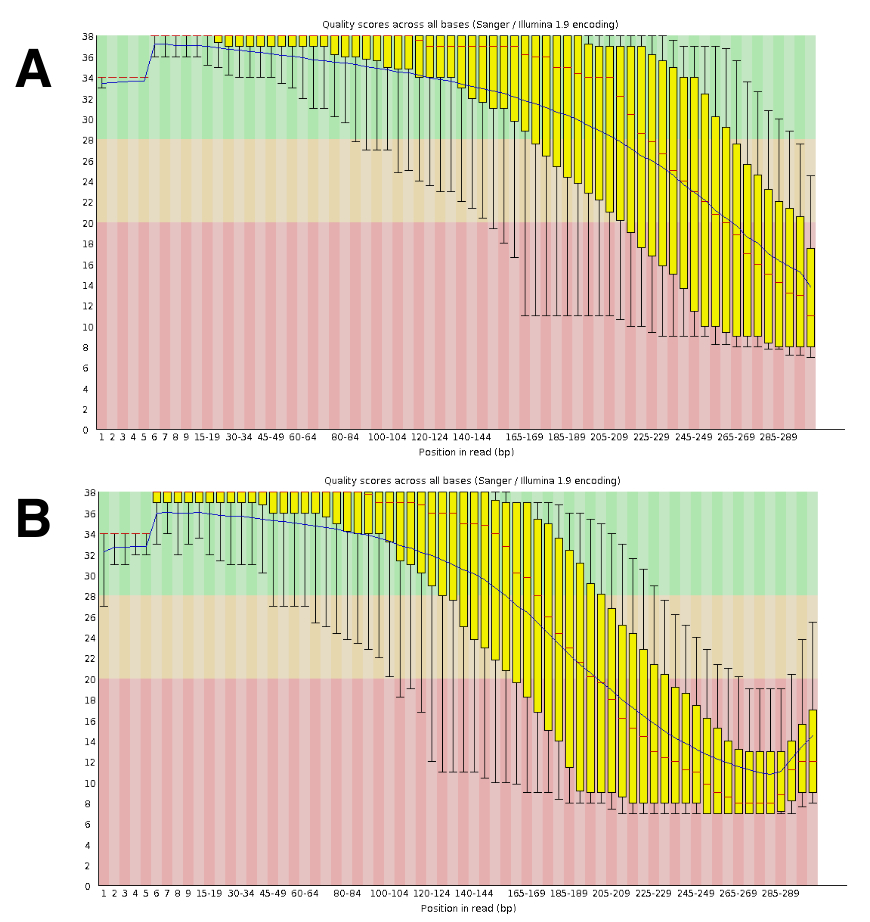
DNA-sekventering genereret omtrent 20 - til 25-M parrede læser per run. Validering af kvaliteten blev gennemført ved hjælp af FastQC (figur 4). Fordelingen af kvalitet langs sekventeret Læs er typisk for en 300-bp parrede køre.

Figur 1: arbejdsprocessen for denne protokol. Denne figur viser en oversigt over denne protokol. Venligst klik her for at se en større version af dette tal.

Figur 2 : Repræsentativt billede af en bakterier-fri tardigrade. Denne figur viser billeder af en forurenet (venstre) tardigrade og renset (højre) tardigrade (Hypsibius dujardini), sammen med yderligere forstørrede billeder (nederst). Stavformet celler omkring tardigrade er forurenende stoffer og er angivet med en pil. Skalalinjen angiver 100 µm. venligst klik her for at se en større version af dette tal.

Figur 3 : Validering af fragment længde distribution af biblioteket bygget DNA. Dette panel viser fordelingen af sekventering bibliotek størrelse. De lilla og grønne linjer angiver øvre og nedre markører på 1.500 og 25 bp, henholdsvis. L = stigen, S = 1 prøve/run, N1 - N4 = 4 flergangsbestemmelser/køre. Venligst klik her for at se en større version af dette tal.

Figur 4 : Eksempel på validering af DNA-Seq kvalitet med FastQC. DNA-Seq data blev indsendt til FastQC at validere sekvens ydeevne. Et repræsentativt resultat for DRR055040 pr. base sekvens kvalitet er vist (DDBJ sekvens Læs Arkiv DRA00445516). (A) dette panel viser de fremad læser (R1). (B) dette panel viser de omvendte læser (R2). Venligst klik her for at se en større version af dette tal.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Bakteriel forurening udgør en trussel mod den genomisk sekventering af mikroskopiske organismer. Mens tidligere undersøgelser på tardigrade genome sequencing har filtreret ud forurening ved hjælp af omfattende Informatik metoder12,20, sekventeret vi genom fra en enkelt person til at minimere risikoen for kontaminering. Da en individuel tardigrade indeholder cirka 50-200 pg genomisk DNA16 og er indkapslet i et tykt lag af chitin exoskeleton, udelukkelse af forurenende stoffer og høj kvalitet DNA-ekstraktion er de kritiske punkter i denne protokol. Eksisterende Biogeografi kulturer er ikke aseptisk, og dem, der indsamles fra de vilde bære en masse af forurenende stoffer på overfladen, samt rester af fødevarer i deres tarme. Tidligere genome sequencing projekter af Biogeografi har sekventeret 10.000-100.000 personer kollektivt som en prøve12,14, hvilket betyder, at resultaterne er meget tilbøjelige til at være påvirket af bakterielle kontaminanter. I deres betænkning, Boothby et al. indsamlet H. dujardini personer ved hjælp af deres negative phototaxis adfærd14, og gruppen ansætte ikke nogen anti-bakteriel metoder.
For at visuelt undersøge, om der er forurenende stoffer, vi inkuberes tardigrade i antibiotika (penicillin/streptomycin) og undersøgt enkelt under 500 X mikroskop. Ved at isolere et enkelt individ og omhyggeligt undersøge det for alle forurenende stoffer, minimeret vi risikoen for eventuelle forureninger. Lave niveauer af forurening blev bekræftet fra sequencing data som godt16. Med hensyn til DNA-ekstraktion ansat vi manuel homogenisering, samt termisk homogenisering18. Ved at sende den tardigrade enkelte til flydende kvælstof og 37 ° C, revner blev induceret i kitin exoskeleton og lysisbuffer var i stand til at trænge ind i kroppen og lyse celler. Når DNA udbytte er fortsat lavere end forventet, kan både termiske og manuel homogenisering gennemføres for at maksimere udbyttet.
Den metode, der er anført i denne artikel har flere begrænsninger. Først, homogenisering af fryse-tø cykler blev anvendt fra en undersøgelse af nematoder; metoden kan derfor kun være effektivt mod ecdysozoa. For det andet på grund af forstærkning af DNA-fragmenter i DNA sekvens bibliotek fase, kan ikke muligheden af PCR fejl ignoreres. Således sekvens data anbefales ikke til analyse, der kræver høj-nøjagtighed læser (dvs., SNP analyse). Desuden, som vi har anført i protokollen, brugen af de angivne SNA-Seq kit vist i Tabel af materialer er helt afgørende, på grund af det lave antal input DNA. Dette DNA bibliotek byggesæt ligates Illumina adapter sekvenser før forstærkning; Derfor, dette bibliotek kan ikke anvendes for længe-Læs sekventering ved hjælp af PacBio eller Nanopore teknologi. Endelig, en kvalitetskontrol af de beregnede DNA bibliotek under denne protokol forekommer kun én gang, efter sekvensering bibliotek konstruktion. Dette er på grund af den lave indgang på DNA siden de fleste DNA kvantificering og elektroforese metoder ikke kan registrere 50-200 pg af DNA. Derfor har vi gennemført kvalitetskontrol, såsom elektroforese (figur 1) og fluorescens-baserede kvantificeringer, kun efter PCR-amplifikation.
En fuld diskussion af Bioinformatik analyser af disse data ligger uden for rammerne af denne artikel; dog har vi kort sagt flere analyser har vi gennemført. En kvalitetskontrol af sequencing data med FastQC19 beregner per-base kvaliteter, sekvens dobbeltarbejde, etc. sekvens data, der er blevet valideret kan indgives til genom-assembly. Vi har samlet en 132 Mb genom med MaSuRCA v3.1.321 og har sammenlignet kortlægning statistikken beregnes med BWA22 og i denne DNA-sekventering bibliotek med andre H. dujardini genom forsamlinger16QualiMap,23 . Desuden, vi også har brugt denne DNA-sekventering data for udelukkelse af forurenende stoffer i vores undersøgelse17, og har observeret at de sekventeret læsninger er fordelt jævnt over hele genomet.
De fleste projekter på ikke-modelorganismer starter fra dyrkning nok prøvemateriale, som det var tilfældet med de24. Tekniske fremskridt i kultur teknikker har aktiveret høje mængder af tardigrade kultur, men aktuelle kultur metoder er endnu ikke aseptisk, og da de fleste Biogeografi er stadig unculturable i labs, det har været næsten umuligt at gennemføre genom eller transkriptom sekvensering. Denne DNA-sekventering metode fra et enkelt individ gør det muligt at analysere sjældne tardigrade arter, herunder havarter, der er blevet undersøgt mindre. Ved at gennemføre Komparativ genomforskning på et bredere phyletic område, kan en yderligere forståelse af anhydrobiosis mekanismer i Biogeografi være opnået.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ikke noget at oplyse.
Acknowledgments
Forfatterne takke Nozomi Abe, Yuki Takai og Nahoko Ishii for deres tekniske support i genomisk sekvensering. Dette arbejde blev støttet af licensbetaling for Japan-samfund til fremme af videnskab (JSP'ER) Research Fellow, KAKENHI licensbetaling for unge forskere (No.22681029) og KAKENHI licensbetaling for videnskabelig forskning (B), No. 17 H 03620 fra JSP'ER, ved en Tilskud til grundlæggende videnskab forskningsprojekter fra The Sumitomo Foundation (No.140340), og dels af forskningsmidler fra Yamagata Prefectural regering og Tsuruoka City, Japan. Chlorella vulgaris bruges til at fodre Biogeografi var som Chlorella Industry Co Ltd
Materials
| Name | Company | Catalog Number | Comments |
| SZ61 microscope | OLYMPUS | ||
| BactoAgar | Difco Laboratories | 214010 | |
| Penicillin Streptomycin (10,000 U/mL) | Gibco by life technologies | 15140-148 | |
| VHX-5000 System | Keyence | ||
| 0.2mL Silicone coating tube | Bio Medical Science | BC-bmb20200 | |
| Quick-DNA Microprep Kit | ZYMO Research | D3021 | Use of this kit is absolutey critical; see step 3.1 |
| 1.5 mL microtube | greiner bio-one | 616-201 | See 4.1.1 |
| HIgh speed refrigerated micro centrifuge | TOMY | MX-307 | |
| Covaris M220 | Covaris Inc. | 4482277 | |
| ThruPLEX DNA-Seq kit | Rubicon Genomics | CAT. NO. R400406 | Use of this kit is absolutey critical; see step 4.2 |
| Thermal Cycler | Bioer Technology | TC-96GHbC | |
| AMPure XP reagent | BECKMAN COULTER Life Science | A63881 | |
| Ethanol | Wako | 054-027335 | |
| EB buffer | QIAGEN | 19086 | |
| 2200 TapeStation | Agilent | G2965AA | |
| D1000 Reagents | Agilent | 5067-5583 | |
| D1000 ScreenTape | Agilent | 5067-5582 | |
| Qubit dsDNA BR Buffer/Reagent | ThermoFisher Scientific | Q32850 | |
| Cubee Mini-Centrifuge | RecenttecGenereach | R5-AQBD01aqbd | |
| MiSeq 600 cycle v3 | Illumina Inc. | MS-102-3003 | |
| MiSeq Sequencer | Illumina Inc. | SY-410-1003 |
References
- Crowe, J. H., Hoekstra, F. A., Crowe, L. M. Anhydrobiosis. Annual Review of Physiology. 54 (1), 579-599 (1992).
- Mobjerg, N., et al. Survival in extreme environments - on the current knowledge of adaptations in tardigrades. Acta Physiologica. 202 (3), 409-420 (2011).
- Becquerel, P. La suspension de la vieau dessous de 1/20 K absolu par demagnetization adiabatique de L'alun de fer dans le vide les plus eléve. Comptes Rendus de l'Académie des Sciences. 231, 261-264 (1950).
- Ono, F., et al. Effect of ultra-high pressure on small animals, tardigrades and Artemia. Cogent Physics. 3 (1), 1167575 (2016).
- Horikawa, D. D., et al. Tolerance of anhydrobiotic eggs of the Tardigrade Ramazzottius varieornatus to extreme environments. Astrobiology. 12 (4), 283-289 (2012).
- Horikawa, D. D., et al. Analysis of DNA repair and protection in the Tardigrade Ramazzottius varieornatus and Hypsibius dujardini after exposure to UVC radiation. PLoS One. 8 (6), e64793 (2013).
- Horikawa, D. D., et al. Radiation tolerance in the tardigrade Milnesium tardigradum. International Journal of Radiation Biology. 82 (12), 843-848 (2006).
- May, R. M., Maria, M., Gumard, J. Action différentielle des rayons x et ultraviolets sur le tardigrade Macrobiotus areolatus, a L'état actif et desséché. Bulletin Biologique de la France et de la Belgique. 98, 349-367 (1964).
- Jonsson, K. I., Harms-Ringdahl, M., Torudd, J. Radiation tolerance in the eutardigrade Richtersius coronifer. International Journal of Radiation Biology. 81 (9), 649-656 (2005).
- Bemm, F., Weiss, C. L., Schultz, J., Forster, F. Genome of a tardigrade: Horizontal gene transfer or bacterial contamination? Proceedings of the National Academy of Sciences of the United States of America. 113 (22), E3054-E3056 (2016).
- Delmont, T. O., Eren, A. M. Identifying contamination with advanced visualization and analysis practices: metagenomic approaches for eukaryotic genome assemblies. PeerJ. 4, e1839 (2016).
- Koutsovoulos, G., et al. No evidence for extensive horizontal gene transfer in the genome of the tardigrade Hypsibius dujardini. Proceedings of the National Academy of Sciences of the United States of America. 113 (18), 5053-5058 (2016).
- Boothby, T. C., Goldstein, B., et al. Reply to Bemm et al. and Arakawa: Identifying foreign genes in independent Hypsibius dujardini genome assemblies. Proceedings of the National Academy of Sciences of the United States of America. 113 (22), E3058-E3061 (2016).
- Boothby, T. C., et al. Evidence for extensive horizontal gene transfer from the draft genome of a tardigrade. Proceedings of the National Academy of Sciences of the United States of America. 112 (52), 15976-15981 (2015).
- Arakawa, K. No evidence for extensive horizontal gene transfer from the draft genome of a tardigrade. Proceedings of the National Academy of Sciences of the United States of America. 113 (22), E3057 (2016).
- Arakawa, K., Yoshida, Y., Tomita, M. Genome sequencing of a single tardigrade Hypsibius dujardini individual. Scientific Data. 3, 160063 (2016).
- Yoshida, Y., et al. Comparative genomics of the tardigrades Hypsibius dujardini and Ramazzottius varieornatus. PLoS Biology. 15 (7), e2002266 (2017).
- He, F. Total RNA Extraction from C. elegans. Bio-protocol. Bio101, e47 (2011).
- Andrews, S. FastQC a quality-control tool for high-throughput sequence data. , http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (2015).
- Hashimoto, T., et al. Extremotolerant tardigrade genome and improved radiotolerance of human cultured cells by tardigrade-unique protein. Nature Communications. 7, 12808 (2016).
- Zimin, A. V., et al. The MaSuRCA genome assembler. Bioinformatics. 29 (21), 2669-2677 (2013).
- Li, H., Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics. 25 (14), 1754-1760 (2009).
- Okonechnikov, K., Conesa, A., Garcia-Alcalde, F. Qualimap 2: advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics. 32 (2), 292-294 (2016).
- Horikawa, D. D., et al. Establishment of a rearing system of the extremotolerant tardigrade Ramazzottius varieornatus: a new model animal for astrobiology. Astrobiology. 8 (3), 549-556 (2008).

{kind=link}