

Summary
Bakteri hücreleri son derece dağınık şekilde düzenlenir. Yavaş büyüyen Myxococcus xanthus hücreler zamanla bu organizasyonda takip etmek için bir tuzak birçok nesiller boyunca yüksek kronolojik zamanmekansal çözünürlük ile floresan canlı hücre görüntüleme için geliştirilmiştir. Bu yöntemi kullanarak, kromozom segregasyon ve hücre bölünmesi için önemli proteinlerin kronolojik zamanmekansal dynamics tespit edilemedi.
Abstract
Floresans canlı hücre görüntüleme bakteriyel hücre proteinleri ve merkezi hücre döngüsü olayları temel kromozom kayma ve zamansal dinamiği Analizi anahtar bir yöntemdir. Ancak, yavaş büyüyen bakteriler temsil bu moleküllerin resim alma sırasında bir sorun nedeniyle photobleaching fluorophores ve fototoksisite görüntüleme. Burada, biz (4-6 h oluşturma süresi olan) Myxococcus xanthus durumunda bu sınırlamaları aşmak için basit bir protokol tanımlamak. Bu amaçla, M. xanthus hücreleri üzerine kalın bir besin içeren agar yastık ısı kontrollü bir nemli ortamda yetiştirilen. Bu koşullar altında biz tek hücrelerinin büyümesini takip ederek tek tek hücreler katlama zamanını belirlemek. Ayrıca, kromozom segregasyon ve hücre bölünmesi yanında floresans canlı hücre düşsel çoklu ParB-YFP, FtsZ-GFP ve mCherry-PomX gibi ilgili fluorescently etiketli işaret protein içeren hücre yansıması gibi anahtar cep işler hücre döngüsü. Daha sonra elde edilen görüntüler montages ve/veya filmler üretmek için işlenir.
Introduction
Bakteri hücreleri son derece dağınık şekilde asimetrik olarak hücresel kompartmanlarda1,2,3,4içinde yerelleştirme birçok protein ile düzenlenir. Bu yerelleştirme genellikle son derece dinamik ve hücre döngüsü ipuçları veya dış sinyalleri karşılık olarak zaman içinde değişir. Aynı şekilde, bakteriyel kromozom dağınık şekilde son derece bireysel loci belirli hücre altı konumlara öncesi ve sırasında segregasyon işlemi5yerleştirilmiş ile düzenlenmektedir. Bu dinamik uzamsal organizasyon büyüme, bölünme, hücre döngüsü Yönetmeliği, farklılaşma, hareketliliği, sinyal iletimi gibi kromozom organizasyon ve segregasyon için önemlidir; Böylece, aslında tüm yönleriyle bakteriyel işlevi etkiler.
Bu hücresel süreçler kronolojik zamanmekansal dinamikleri Escherichia coli, Bacillus subtilis, Vibrio choleraeve Caulobacter crescentus kadar önemli hizmet ile farklı bakteri türleri çeşitli analiz ediliyor Model organizmalar. Ancak, bu dört tür büyük bakteriyel çeşitliliğin sadece küçük bir spektrum kapak ve belki de şaşırtıcı olmayan bir şekilde bu türler arasında büyük bir filogenetik mesafe göz önüne alındığında, hücresel organizasyon ve polarizasyon bunlar içinde farklı mekanizmalar bakteri. Bu sonunda genel ilkeleri bakteri hücreleri kronolojik zamanmekansal dinamikleri temel ayıklamak edebilmek için ek bakteriyel türler eğitim gereksinimini kaldırmak.
Ayagin Gram-negatif delta proteobacterium M. xanthus bir model organizma insan sosyal davranışları ve işbirliği bakteri6. M. xanthus sıkı bir aerobe olduğunu ve besin varlığında, hangi hücreleri yayılmış son derece koordineli olarak dışa, swarming moda ve diğer mikroorganizmalar7dualarının kolonileri oluşturur. Besin açlık yanıt olarak, hücre hücre ve hangi içinde binlerce oluşur olgun organları oluşumunda sonuçları bir gelişimsel programını başlatmak çubuk şekilli hareketli hücreler için küresel ayırt diploid Sporlar8. Her iki tür davranışları, yani, kaynıyor ve meyve vücut oluşumu, sadece düz bir yüzey üzerine yerleştirilir hücreleri tarafından yürütülür. Ayrıca, her iki besin koşullar altında hücre, hareketliliğini teşvik veya alıcı9,10 toksin olarak işlev dış membran lipoproteinler değişimi de dahil olmak üzere doğrudan hücre-hücre kişileri içeren süreçlerini meşgul , LPS11, motilite komşu tarihinde exopolysaccharides tarafından uyarılması alışverişini12hücreleri ve hücreler arası tarafından bir hücre sinyallemesi yüzeyi protein13,14sinyal bağlantılı.
Son zamanlarda, M. xanthus ayrıca hareketliliği ve onun Yönetmeliği15, hücre bölünmesi16,17,18ve kromozom organizasyon19 yatan mekanizmaları eğitim için bir model organizma haline gelmiştir ,20,21. Kritik adımlar M. xanthus hücre döngüsü analiz ayrıntılı olarak floresans mikroskobu görüntüsü görüntüleri veya kısa zaman hata kayıtları ilgili fluorescently etiketli proteinler16, taşıyan suşları kullanarak tarafından 17,18,19,20. İdeal olarak, birçok hücreye tek hücreli çözünürlük ile floresan canlı hücre hücre döngüsü parametrelerdeki sağlam nicel verileri elde etmek en az bir tam hücre döngüsü için Imaging tarafından takip edilmelidir. Ancak, bu durumda bir mücadeledir M. xanthus ve photobleaching fluorophores ve resim alma sırasında fototoksisite nedeniyle, nispeten uzun oluşturma süresi 4-6 h Standart laboratuvar koşullarında nedeniyle.
Burada, biz takip için bir protokol tarif M. xanthus hücreleri tek hücre kararı ile floresan yaşamak için en az 24 saat görüntüleme ve birkaç hücre döngüsü kapsayan hücre ile. Önemlisi, tüm protokol sırasında hücreleri bir agar altlığında korunur ve Kapat'kişiyi bağımlı faaliyetler için sosyal yaşam tarzı için temel sağlayan iletişim M. xanthus. İletişim kuralı da bırakmak kullanıcı-monitör şekil, boyut, bölüm ve floresan problar yüksek zamansal çözünürlükte ve tek hücre çözünürlük ile ve böylece, hücre hücre değişkenlik miktar ve hücre döngüsü olaylar korelasyon etkinleştirir.
Protocol
1. hazırlık ve büyüme M. xanthus suşlarının
Not: Tablo 1 ve Tablo 2bakın.
- %1 casitone suyu (CTT) büyüme orta %1 (w/v) pankreas özünü kazein (Örneğin, Bacto casitone), 10 mM Tris-HCl pH 8.0, 1 mM KH2PO4 pH 7,6, 8 mM MgSO422, sefaloridin (50 µg/mL) ile desteklenmiş hazırlamak veya oksitetrasikline (10 µg/mL). Viomisin (10 µg/mL) beri diğer bakteri ile bulaşma riskini azaltmak için tüm ortamına Ekle M. xanthus için doğal dirençli hücrelerdir.
- 5 mL % 1'de aşılamak ilgili içeren CTT antibiotic(s) vahşetin taze yetişkin bir koloni ile (WT) DK1622 23, SA4420 (ΔmglA)24, SA4797 yazın (ΔmglA, ΔpomX/PpomZ mCherry-pomX )16, SA8241 (ΔmglA, ftsZ+/PnatftsZ-gfp), veya SA4749 (ΔmglA, parB+/PnatparB-yfp) içinde 1. gün sabah.
- Tek bir resuspend M. xanthus 1 CTT antibiyotik steril bir tüp ile desteklenmiş ve bir 50 mL Erlenmeyer şişesi 5 mL % 1'i de içeren tüm süspansiyon transfer % 500 µL kolonisinde CTT.
Not: bir Erlenmeyer şişesi ile kültür hacmi 10 kez yeterli aeriation ve optimal büyüme garanti altına almak için kullanın.
- Tek bir resuspend M. xanthus 1 CTT antibiyotik steril bir tüp ile desteklenmiş ve bir 50 mL Erlenmeyer şişesi 5 mL % 1'i de içeren tüm süspansiyon transfer % 500 µL kolonisinde CTT.
- Karanlıkta 220 rpm'de sallayarak 32 ° C'de sekiz nesillerdir (yaklaşık 4-6 h oluşturma süresi ile 40-48 h) hücreleri büyümek. Üstel büyüme aşamasında hücreleri korumak (OD550 < 1.2) ve durağan faz ulaşmasını engellemek. Gerekirse, içine taze % 1 CTT orta 0.1 OD550 ilgili antibiotic(s) içeren hücreleri seyreltik - 0.2.
Not: Bir tek hücre Mikroskopi için en uygun bir OD550 0,5 - olarak 0,7. Bu OD550, yeterli sayıda hücre başına miktar hem de hücresel parametrelerin istatistiksel analiz izin vermek için görüntü mevcuttur.
2. mikroskobu örnekleri hazırlanması
Not: mikroskobu tarafından görülebilmesini hücreleri mikroskop coverslip üzerinde yerleştirilir ve sonra besin içeren bir özel yastık tarafından kaplı. Coverslip bir plastik için yapıştırılmış veya mekanik sağlamak için metal çerçeve desteklemiyor. Mikroskopi için hazırlık olarak, % 1 ' agarose/TPM/0.2% büyük bir yastık CTT 2.1 2.3 adımlarda açıklandığı gibi önceden hazırlanmalıdır. Ayrıca burada kullanılan belirli ürünler için Malzemeler tablo bakın.
- TPM arabelleği (10 mM Tris-HCl pH 7,6, 1 mM KH2PO4 pH 7,6, 8 mM MgSO4) 500 mL hazırlama ve otoklav veya filtre sterilize bir şişe en iyi filtre kullanarak.
Not: Steril tampon oda sıcaklığında birkaç ay depolanabilir. - %0,2 içeren %1 özel mikroskobu çözüm hazırlamak CTT (mix özel 80 mL TPM arabellek ve % 1 CTT orta 20 mL ile 1 g). Bir mikrodalga fırında ısı özel erimiş kadar.
Not: % 0.2 CTT hücreleri büyümek ve starvation önlemek için izin vermek yeterli olur. CTT daha yüksek konsantrasyonlarda mikroskobu orta yüksek arka plan floresans neden olur. - (Bir 11,5 cm x 11,5 cm kare Petri dish, erimiş özel yaklaşık 60 mL gereklidir için) bir petri 0,5 cm kalınlığında için erimiş özel ile doldurun ve aşağı oda sıcaklığına kadar soğumaya bırakın.
Not: Özel yastık nemli bir ortamda 4 ° C'de 2 güne kadar saklanır.- En az 15 dakika kullanmak için önceden 32 ° C'de % 1 agarose/TPM/0.2% CTT pad önceden ısıtmak.
Not: hücreleri Mikroskopi için hazırlamak için 2,4-2.8 yapınız.
- En az 15 dakika kullanmak için önceden 32 ° C'de % 1 agarose/TPM/0.2% CTT pad önceden ısıtmak.
- Steril cam coverslip yer (60 mm x 22 mm, Kalınlık: 0.7 mm), Orta (resim 1-A); bir delik olan plastik veya metal çerçeve Bu çerçevede ince coverslip için mekanik bir destek olarak hizmet vermektedir ve mikroskopi sırasında drift azaltmaya yardımcı olur. Coverslip çerçeve için bant ile düzeltmek.
- Çerçeveyi hazırlamak için 1 mm kalınlığında metal plaka bir 75 mm × 25 mm çerçeveden kesip, sonra orta uygun büyüklükte bir delik (20 mm × 30 mm bu deneyde) kesip.
- 10-20 µL katlanarak büyüdü, eklemek M. xanthus hücre coverslip.
- Floresan 0.5 µm mikroküreler hücrelere hücre veya proteinler hızlandırılmış kayıtları izleme kolaylaştırmak indirgeme işaretleyicileri ekleyin.
- TPM arabelleği mikroküreler 1: 100 sulandırmak ve 4 ° C'de için birkaç ay kadar saklayın. İyice kullanmadan önce çalkalayınız ve 5-10 µL seyreltilmiş mikroküreler, hücrelere ekleme.
Not: Burada mikroküreler bu tüm ortak mavi, yeşil, sarı ve kırmızı floresan kanalları kullanıldı içinde floresan.
- TPM arabelleği mikroküreler 1: 100 sulandırmak ve 4 ° C'de için birkaç ay kadar saklayın. İyice kullanmadan önce çalkalayınız ve 5-10 µL seyreltilmiş mikroküreler, hücrelere ekleme.
- Küçük bir yastık yaklaşık büyük önceden ısıtılmış % 1 agarose/TPM/0.2% CTT pad coverslip boyutunu keser ve hücreleri (Şekil 1B) üstüne yerleştirir. Bir coverslip buharlaşma önlemek ve nemli bir ortamda hücreleri korumak için % 1 agarose/TPM/0.2% CTT özel yastık üstüne yerleştirin.
Not: Yalnız coverslip en az 2 h için önemli buharlaşma engeller. Artık bu hızlandırılmış kayıtları için % 1 agarose/TPM/0.2% CTT pad ve coverslip sandviç ile parafin film buharlaşma önlemek için kapalı olmalıdır. - 15-20 dk hücreleri özel yastık altına eklemek izin vermek için 32 ° C'de mikroskobu örnek kuluçkaya. Hızlandırılmış mikroskobu kayıtları başla o zaman.
3. mikroskop kurulum ve hızlandırılmış edinme
Not: burada açıklanan protokol bir ters widefield mikroskobu ile autofocus, bir 100 X için geliştirilmiş / 1,30 NA petrol PH3 amaç, bir X, Y motorlu sahne, sCMOS kamera, bir ışık kaynağı, yeşil-floresan için filtreler, kırmızı-floresan veya sarı-floresan proteinler ve sıcaklık kontrollü kuluçka odası. Bu odanın ışık ve sabit sıcaklık korumalı hücreler tutar.
- Kuluçka odası ve 32 ° c ~ 1-2 h için mikroskop ön ısı mikroskobu başlamadan önce.
Not: mikroskop kurulumu bağlı olarak, Isıtma daha uzun sürebilir. Ön ısıtma drift azaltmak için önemlidir ve otofokus kontrol sistemi stabilize. - Mikroskobunun geçin ve mikroskop kontrol yazılımını başlatın. Doğru hedefi seçin ve doğru aynalar ve elde etmek için filtre kontrast görüntüler aynı zamanda yeşil-floresan, kırmızı-floresan veya sarı-floresan proteinlerin görüntüleri aşama.
Not: Mikroskop genellikle mikroskop kontrol ve görüntü edinme için tercih edilen bir yazılım ile sağlanır. Burada ticari bir yazılım ( Tablo reçetesigörmek) mikroskop ve görüntü alma kontrol etmek için kullanıldı. - Yüksek kaliteli Daldırma yağ amacın ve önceden 32 ° C'de inkübe örnek altına objektif üzerine bir damla ekleyin Amaç en düşük fiyat olası Z-örnek mikroskop sahne alanı'nda yerleştirildiğinde objektif lens zarar görmesini önlemek için konuma yerleştirin. Metal çerçeve "delik-tarafı" yönünde amacı ile mikroskop sahne alanı'na örnek ile yerleştirin. Örnek içinde sahne alanı sahibinin güvenli bir şekilde bağlayın.
- Hücreleri üzerinde sahne Z-yönünde daha yakın hedefiyle hareket ettirerek odaklan. Örnek alt yüzündeki yağ damlaları ve objektif lens yapmak zaman sahne daha yavaş hareket kişi. Sahne hareket X / Y yönü birden çok tek hücre kadar görünür görünümü, bölgede ne zaman odak düzlemi hücrelerdir. Daha sonra alınan görüntüleri hizalamak için görünümü bölgede en az bir o floresan microsphere olduğundan emin olun.
Not: optimum koşullar altında görünümü (2.048 x 2.048 piksel veya 133.1 x 133.1 µm) bölgenin başına 15-30 hücre hücre yoğunluğu erişilmesi gereken. - Gerekirse birden çok dalga boylarında ve sahne konumlarını görüntüleri elde etmek mikroskop sağlar bir zaman atlamalı deneme oluşturmak için mikroskop kontrol yazılımı Çok boyutlu alma sihirbazını açın.
- Ana sekmede Timelapse ve Birden çok dalga boylarındaetkinleştirin. Ek sekmeler penceresinin sol tarafında görüntülenir.
- Kaydetme sekmesinde ve alınan görüntüleri kaydetmek için bilgisayar sabit disk üzerindeki boş bir klasör seçmek için Dizin Seç'i tıklatın. Ardışık veri kümeleri öncekileri üzerine yazılmasını emin olmak için dosya varsa artışı temel adı etkinleştirin. Sonra deneme tarih ve soy adı veya başlık deneme ile bir ad verin.
- Hızlandırılmış parametrelerini ayarlamak için Timelapse sekmesini tıklatın. Süresi için 24 saat ve 20 dk Zaman aralığını ayarlayın. Zaman puan numarasını otomatik olarak değişecektir.
Not: Deneme ve hücresel işlev çözümlenmesi için en uygun zaman aralığı bağlıdır. Sık sık resim satın almalar photobleaching neden olabilir. Böylece, ampirik olarak zamansal çözünürlük ve photobleaching arasında bir denge kurulmalı. 4-6 h katlama zamanda görüntüleri kolayca 5 dk (ya da isterseniz bile daha küçük aralıklar) bir aralıkta faz kontrast mikroskobu için elde edilebilir. Floresans mikroskobu zamanlı kurs 24 h üzerinde görüntüleri yaklaşık 15-30 dakika bir aralıkta kaydedilmiş isteniyorsa. - Dalga boylarında sekmesini tıklayın, her zaman bir noktada her resim için sayıyı değiştirerek elde etmek için dalga boylarında sayısını seçin.
Not: her dalga boyu için sol tarafında yeni bir sekme görünür çok boyutlu edinme " Sihirbazı ve dalga boylarında elde edilebilir üst sırada yukarıdan aşağıya doğru. Her dalga boyu için satın alma ayarlarını ayrı ayrı değiştirilebilir. - Üstten ilk dalga boyu sekmesini tıklatın. Faz kontrast aydınlatma aþaðý açýlan listeden seçin. 100 ms pozlama için seçin ve Her zaman nokta edinme aþaðý açýlan listeden seçin. Otomatik ortaya çıkarmak hiçbir zaman aşağı açılan listeden seçerek devre dışı bırakın.
- Her zaman noktası elde edilebilir gerekiyor her dalga boyu için 3.5.5 arasındaki adımları yineleyin. Deneysel kurulum ve fluorescently etiketli proteinler açıklanan burada, kullanmak için aşağıdaki parametreleri için pozlama: mCherry füzyon proteinler için 250 ms, YFP füzyon proteinler için 200 ms ve GFP füzyon proteinler için 1000 ms.
Not: Her zorlanma ve floresan protein optimum aydınlatma ayarlarını önceden lamba yoğunluk ve her dalga boyu görüntü edinme süre değiştirerek tespit edilmelidir. Çok uzun görüntü edinme kez Fototoksik etkisini artırmak ve sonuçta büyüme tutuklama ve hücre ölümüne yol açar. Bu nedenle, görüntü kalitesi ve hücre canlılığı arasında bir denge elde edilmelidir. - Aynı denemede kaydedilen hücre sayısını artırmak için birden çok sahne pozisyonlar görüntülerden elde etmek.
- Birden çok sahne pozisyonlar görüntüleri elde etmek için Birden çok sahne pozisyonlar ana sekmesini etkinleştirin. Sonra sahne sekmesini ve görüş alanı bakmak için Live düğmesini tıklatın.
- Görüş alanı içinde faiz (ROI) bölgedir kadar sahne X/Y-yönde hareket ettirin. Kaydet X - ve Y-tıklayarak koordinatları "+" sahne sekmesini içinde sahne tekrar X/Y-yönünde yeni bir yatırım getirisi bulana ve koordinatları tekrar tıklatarak "+" hareket. Bölgeler için istediğiniz sayıyı kaydedilinceye kadar devam et.
Not: floresan resim alma durumunda faiz (ROIs) bölgelerinde hepsiyle alâkan olmadığından emin olun birbirlerine fototoksisite en aza indirmek için.
- Bir kez daha hücreler farklı kaydedilen X - ve Y-konumları üzerinde tıklatarak odakta olan ve donanım otofokus kaydedilmiş Z-pozisyon deneme boyunca sabit tutmak için AFC tutun tıklayarak başlayın kontrol edin.
- Hızlandırılmış kayıtları mikroskop kontrol yazılımı içinde Çok boyutlu alma sihirbazını Al tıklayarak başlayın.
Not: Alınan her dalga boyu için bir pencere görünür ve alınan zaman puan ve sonraki resim satın kadar zaman sayısını gösteren ek bir pencere görünür. - Hücreleri ilk birkaç saat-puan görüntü kalitesini en üst düzeye çıkarmak ve gerekirse yönlendirmesi için hızlandırılmış kayıtları sonra hala odakta olduğundan emin olun.
4. nesil hızlandırılmış film ve resim hizalaması
Not: Birçok ticari ve özgür yazılım paketleri resim alma ve görüntü analizi için kullanılabilir. Biz ticari bir yazılım kullanın (bkz. Tablo reçetesi) birden çok önceden yüklenmiş eklentiler ve ek araçları ile.
- Görüntü analizi/işleme yazılımı yüklü olduğu bir bilgisayarda hızlandırılmış kayıtlarından bireysel görüntüleri kaydetmek.
- Belgili tanımlık bilgisayar yazılımı başlamak ve açık görüntüleri bir yığın bir daha gözden geçirme çok boyutlu veri tıklatarak | Temel dosya seçin | Dizini seçin. Çok boyutlu verilerle klasörü açın. Veri kümesi denetleyin ve görüntüle' yi tıklatın; Tek görüntüleri zaman sonuna kadar bir işaret olarak veri kümesi gösterilir. (İçin bir yığın oluşturma) dalga boyu etkinleştirmek, yığın içinde olmalı ve Yük resimleritıklatın tüm görüntüleri seçin. Tüm dalga boyları için bu adımı yineleyin ve tamamlanan yığınları kaydedin.
- (İsteğe bağlı) Dosyasını kullanarak film için gerekli tüm görüntüleri açın | Açık.
Not: Bu görüntüleri açmak için bir elde edilen dalga boyu tarafından teker teker işlem gücü sınırlı ise bilgisayarı değil yavaşlatmak için tavsiye edilir. Hızlandırılmış kayıtları, Örneğin, başlangıç, bitiş veya birkaç zaman puan bazı kısımlarını atlanması Eğer, o zaman bu tamamlanan filmde ayarlanabilir. - Drift için düzeltilmesi gereken görüntülerin yığınını etkinleştirin. Apps tarafından Hizalama aracını açın | Hizala otomatik..... onay yığın görüntüleri ve başvuru düzlemi olarak ilk uçak/saat noktası için kaynak olarak. Kaynak yığın düğmesini yığınıyla seçin ve Uygula' yı tıklatın.
Not: Otomatik hizalama biraz zaman ve bir işlem gücü alacak ama drift mikroskop tuzak için büyük yığınlar düzeltmek için iyi bir yoldur. De mikroküreler eklenir, ancak aynı zamanda onlar olmadan çalışabilir bu otomatik hizalama çalışır. - Hizalanmış yığın kaydedin.
- ROIs kullanın.
Not: Floresan hızlandırılmış mikroskobu kolayca almak yukarıya biraz çok-in bir işlem gücü ve bu filmler aşağı akım işleme kapalı yavaş veri dosyalarının büyük kümeleri oluşturur. Bu nedenle ROIs tanımlanmasını ve yalıtılmasını hücreleri daha küçük dosyalarla çalışmak için önerilir.- Dikdörtgen aracını seçin. Bir yatırım getirisi faiz hücrelerin etrafındaki el ile faz kontrast görüntü üzerinde bir yatırım getirisi çizerek oluşturma. Hücreleri ilgi ve odak bütün hızlandırılmış film boyunca görünür olduğundan emin olun.
- İkinci dalga boyu aynı veri kümesinin hızlandırılmış film açın. ROI faz kontrast resimlerden floresan için aktarmak için ikinci dalga boyu resimleri bölgelere Transfer bölgeleri aracıyla kullanmak | Bölgeler transfer. Kaynak görüntüyü ve ikinci dalga boyu veri kümesi olarak Hedef görüntüolarak faz kontrast veri kümesini seçin. Tüm bölgeleri seçin ve Tamam'ıtıklatın.
- Aynı veri kümesi için alınan her dalga boyu için 4.6.2 arasındaki adımları yineleyin.
- ROI seçin ve düzenleme ile bir yığın olarak yinelenen | Yinelenen | Yığın... veya Shift + Ctrl + D tuşlarına basın. Dosya yinelenen yığınıyla kaydetmek | Kaydet özgün veri olarak aynı klasöre.
- 4.6.4 için aynı veri kümesi için alınan her dalga boyu her yatırım getirisi yineleyin
- Bir film MOV veya AVI formatlarında üretmek için yığını üzerinden Yapmak film işlevini açın | Film yapmak. Kaynak yığını düğmesi ile hızlandırılmış kayıtları seçin. Çıkış biçimi, kare hızı, kare sayısını seçin ve Kaydet' i tıklatın.
Representative Results
M. xanthus katı yüzeyler üzerinde hareket eden bir yavaş büyüyen bir bakteridir. Deneysel yapımız test etmek için hareketli DK1622 WT hücreleri ile hızlandırılmış bir deneme yapılır. Faz kontrast görüntüleri 24 h (Şekil 2A, B) için 5 dk aralıklarla elde. Hücreleri çoğunluğu gruplar halinde düzene sokmak. Beklendiği gibi hücre hareketliliği görüntülenen ve ağırlıklı olarak gruplar halinde taşındı. Biz daha fazla hücreler zaman zaman hareket yönü tersine gözlenen. Bu bulgular WT hücreleri test edilmiş koşullar altında normal açısından hücre hareketliliği davranırlar öneririz. Hücreleri her 5 dk bile kaydedilir, ancak, tek tek hücreleri tanımlaması zordur. Ayrıca, hücreleri hareketli olduğundan, birçok hücreye kaçmak veya hücreleri uzun süre takip zorlaştırır görüş alanı girin.
Aynı izleme amacıyla M. xanthus hücreleri hücre döngüsü yanında canlı hücre düşsel birkaç tur için-ebilmek var olmak silmek bireysel suşları hareketliliği25için gerekli olan mglA gen için. Bu hücreleri görüntüleme iletişim kuralı sırasında görüş alanı dışına taşımalarını engeller. Çerçeve silme Shi ve arktarafından açıklandığı gibi oluşturulur. 26
Hareketli olmayan ΔmglA hücreleri (Şekil 3), ile faz kontrast canlı hücre görüntülemede beklendiği gibi hücreleri aktif hareket göstermedi. Biz büyüme ve tek tek hücreler bölünme microcolony oluşumu sırasında takip edebildik. İçinde 24 h için 5 dk aralıklarla görüntüleri elde zaman hata kayıtları bağlı olarak, tek hücre çözünürlük interdivision zamanla (iki hücre bölünmesi olaylar arasındaki süreyi) ölçmek mümkündü. ΔmglA mutant hücrelerinin vardı bir arası bölümü zaman 235 ± 50 dk (n = 97 hücre). Yaklaşık 4 h ile interdivision zaman süspansiyon kültürlerde WT hücreler için ölçülen katlama zaman benzer. Bu, bu kanıt sağlar M. xanthus hücrelerin büyümesine en iyi şekilde bu deneysel koşullar altında.
Biz floresan ile görüntüleme hızlandırılmış gerçekleştirilen yapımız hücreleri normalde uzun sürelerle YFP etiketli proteinler izleme süre büyümeye izin verip vermediğini öğrenmek için M. xanthus YFP öğesini protein Hızlı hücre. Bu amaçla, çoğaltma (ori) kökeni için bir işaretleyici olarak ParB-YFP izledik. ParB olduğunu ParABS sistem bileşeni olarak M. xanthus ve oriiçin; proksimal parS sitelere bağlar Bu nedenle, kaynak çoğaltma ve kromozom segregasyon takip19,20,21olabilir. İle her 20 dk edinme (faz kontrast ve floresan, 200 ms edinme zamanında YFP kanal) görüntü, hücreleri büyüdü, bölünmüş ve büyüme bile 24 h (Şekil 4A) olarak görüntülenir. Kayıtları başlangıcında, ParB-YFP (Şekil 4A) hücrelerin çoğunda subpolar bölgelerde iki küme kurdu. Kısa bir süre önce veya sonra hücre bölünmesi, çoğaltılamaz eski hücre Kutbu'nda küme subpolar ParB YFP. İki kümeden eski hücre Kutbu yaklaşık 40-60 dakika sonra (Şekil 4A, B) son subpolar konumuna ulaşan yeni hücre kutup translocated ikinci kopya süre kaldı. Bu gözlemler kısa zaman hata kayıtları ince agar yastıkları19kullanarak oluşturulan önceki veri ile anlaşma vardır. Biz bu deneysel kurulum floresans hızlandırılmış mikroskobu kromozom segregasyon birkaç hücre yavaş büyüyen döngü izlemek sağlar sonucuna M. xanthus perturbing hücre büyümesini veya kromozom segregasyon makine olmayan hücreler.
Benzer bir deneyde, hızlandırılmış floresans mikroskobu ile hücre bölünmesi için işaretleri izleyin istedi. Neredeyse tüm diğer bakteri, benzer M. xanthus FtsZ, bakteriyel tübülin benzeri GTPazlar, hücre bölünmesi16,17,18için gerektirir. FtsZ, midcell, sözde Z-hücre bölünmesi27,28için gerekli diğer proteinler askere yardımcı olan yüzük, yüzük benzeri yapısı oluşturur. İçinde M. xanthus, Z-ring ve onun midcell konumlandırma oluşumu üç PomXYZ proteinler16,17tarafından uyarılır. Bu üç proteinler hemen "Anne" hücredeki hücre bölünmesi sitesinden çevrili iki kızı hücrelerdeki ortasına aktarır kromozom ilişkili bir kompleks oluştururlar. Hemen ortasında daha önce kromozom segregasyon ve burada midcell ile PomXYZ karmaşık acemi FtsZ çakışacak ve Z-halka oluşumu uyarır.
Buraya, ilk hareketli olmayan hücreleri ftsZ-gfpifade kadar takip etmiş. Çünkü FtsZ-GFP genel gösterir daha zayıf bir floresan sinyal ParB-YFP biz arttı çekim hızı 5-fold 1 s GFP kanaldaki. Beklendiği gibi güçlü FtsZ-GFP birikimi sadece midcell gözlendi ve hücre bölünmesi daralma (Şekil 5A) konumunu bu yerelleştirme dikte. FtsZ-GFP bir küme midcell daha uzun hücre içinde ağırlıklı olarak kurdu. Bu küme yoğunluğu zaman içinde artan açıktı. Sonra hücre bölünmesi, FtsZ-GFP yeniden midcell iki kız, birikmiş yaklaşık 2 saat sonra (Şekil 5B) hücreleri görülmektedir. Bu hücreler bir nüfus yaklaşık % 50'si görüntüsü analiz16,17tarihinde dayalı midcell adlı FtsZ yerelleştirme görüntülemek bulma ile tutarlıdır.
İkinci bir deneyde, mCherry-pomXhızlı hareketli olmayan ΔmglA hücreler için 24 saat takip ettik. PomXYZ sisteminin bir parçası olarak, PomX Kılavuzu Z-halka oluşumu ve konumlandırma, böylece hücre bölünmesi midcell16uyarıcı yardımcı olur. MCherry-PomX floresans sinyalinin ve güçlü bir çekim hızı 250 Bayan floresans kanalda da önemlisi sağlar, tüm hücreleri boyutunda büyüdü ve 24 h ( sonra microcolonies oluşturan deney boyunca bir hücre bölünmesi olayı görüntülenir Şekil 6A). Daha önce raporlanmış16olarak, hemen hemen tüm hücrelerin bir mCherry-PomX küme bulunan. Bunlar midcell ve deneme süresince midcell için translocated midcell uzak kümeleri yerelleştirilmiş çoğunluğu. Hücre bölünmeler sırasında mCherry-PomX kümeleri, her kızı bir küme alma hücre ile ayrılmış. FtsZ-GFP, aksine mCherry-PomX hücre döngüsünün midcell 80-90 oranında lokalize ve bu pozisyonda yakında sonra hücre bölünmesi (Şekil 6B) ulaştı.
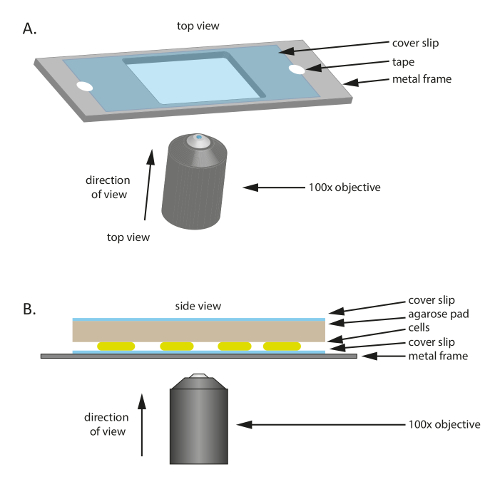
Resim 1 : Bu çalışma kullanılan deneysel tuzak şematik. (A) A metal veya plastik çerçeve hizmet örnek için bir destek olarak. Bir coverslip metal çerçeve hareket örnek azaltmak için bant ile sabittir. (B) yan deneysel örnek up görüntüleyin. Hücreleri (A) gösterilen coverslip üzerine monte edilir. Besin ve nem hücrelere sağlayan özel panel hücreleri üzerine yerleştirilir. Özel yastık buharlaşma azaltmak için ek bir coverslip tarafından kaplıdır. Yüksek kaliteli görüntüler için bir 100 X yağı daldırma faz kontrast objektif kullanılır. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Resim 2 : Faz kontrast WT hızlandırılmış mikroskopi M. xanthus hücreleri. Hücreler için 24 saat takip ve görüntüleri her 5 dk.(a)elde temsilci aynı görüş alanı her 5 dk görüntülerini gösterilir. Renkli oklar, bireysel hücrelerin hareketinin yönünü gösterir. Aynı renk, zaman içinde aynı hücreyi işaretler. Sayılar süreyi dakika cinsinden belirtir. Ölçek çubuğu: 5 görüntüleri µm. (B) her saat sonra aynı görüş alanı gösterilir. Not aynı görüş alanı gösterilir ancak hücreleri hareket ediyor çünkü, hücreler sürekli girme ve görüş alanı bırakarak. Sayılar süreyi saat gösterir. Ölçek çubuğu: 5 µm. PH: faz kontrast. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Şekil 3 : Faz kontrast, hareketli hızlandırılmış mikroskopi M. xanthus hücreleri. ΔmglA hücreleri için her 5 dk 24 h görüntüleri elde ve her saat sonra temsilcisi görüntüleri gösterilir izledi. Seçili hücre bölünmesi boğumların turuncu oklarla işaretlenmiş. Sayılar süreyi saat gösterir. PH: faz kontrast. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
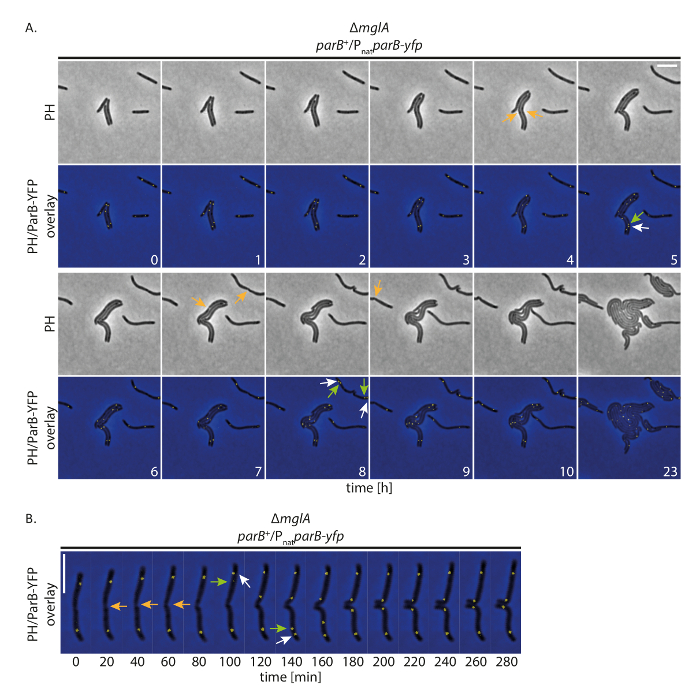
Şekil 4 : ParB-YFP floresan hızlandırılmış mikroskobu içinde hareketli olmayan M. xanthus hücreleri. Hücreleri bir ΔmglA mutantifade parB yfp huzurunda yerli parB (SA4749; ΔmglA; parB +/PnatparB-yfp) 24 saat faz kontrast ve floresan mikroskopi tarafından takip edildi. (A)görüntüleri her 20 dk elde ve temsilcisi görüntüler her saat 10 h gösterilir kadar aynı hücreler 24 h görüntüleri faz kontrast (PH) gösterilir sonra birlikte ve yerleşimi faz kontrast ve YFP sinyal gibi. Seçili hücre bölünmeler turuncu oklarla işaretlenmiş. Beyaz ve yeşil okları ParB-YFP küme çoğaltma olaylar, translocating küme işaretleme yeşil oklarla işaret ediyor. Sayılar süreyi saat gösterir. Ölçek çubuğu: 5 µm. (B) resim (Aolduğu gibi) satın alınan ama daha yüksek zamansal çözünürlükte görüntülenir. Sayılar süreyi dakika cinsinden belirtir. Okları (Aolduğu gibi) vardır. Ölçek çubuğu: 5 µm. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Şekil 5 : FtsZ-GFP floresans hızlandırılmış mikroskobu içinde hareketli olmayan M. xanthus hücreleri. FtsZ-gfp yerli ftsZ (SA8241; ΔmglA; huzurunda ifade ΔmglA mutant hücrelerinin ftsZ +/PnatftsZ-gfp) için 24 h faz kontrast ve floresan mikroskopi tarafından takip edildi. (A)görüntüleri her 20 dk elde ve temsilcisi görüntüleri 10 h kadar her saat gösterilir, 24 h sonra aynı hücreleri ile birlikte görüntüleri faz kontrast (PH) ve kaplama faz kontrast ve GFP sinyal olarak gösterilir. Seçili hücre bölünmeler turuncu oklarla işaretlenmiş. Beyaz ok FtsZ-GFP kümeleri midcell adlı göstermek. Sayılar süreyi saat gösterir. Ölçek çubuğu: 5 µm. (B) resim (Aolduğu gibi) satın alınan ama daha yüksek zamansal çözünürlükte görüntülenir. Sayılar süreyi dakika cinsinden belirtir. Yeşil ve beyaz okları FtsZ-GFP kümeleri sol ve sağ hücrelerde sırasıyla işaretleyin. Turuncu oklar hücre bölümler gösterir. Ölçek çubuğu: 5 µm. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Şekil 6 : MCherry-PomX içinde hareketli olmayan floresan hızlandırılmış mikroskopi M. xanthus hücreleri. MCherry-PomX (SA4797; ΔmglA; ΔpomX/PpomZ mCherry-pomX) biriken sigara hareketli ΔpomX hücreleri faz kontrast ve floresan mikroskopi tarafından her 20 dk.(a)24 saat takip temsilcisi görüntüleri 10 h kadar her saat gösterilir, 24 h sonra aynı hücreleri ile birlikte görüntüleri faz kontrast (PH) ve faz kontrast ve mCherry kaplama olarak gösterilir sinyal. Seçili hücre bölünmeler turuncu oklarla işaretlenmiş. Beyaz ve yeşil oklar, öncesi ve sonrası olaylar, sırasıyla yarma mCherry-PomX kümeleri gösterir. Sayılar süreyi saat gösterir. Ölçek çubuğu: 5 µm. (B) resim (Aolduğu gibi) satın alınan ve daha yüksek zamansal çözünürlük gösterilir. Okları (Aolduğu gibi) vardır. Ölçek çubuğu: 5 µm. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
| Bakteriyel zorlanma | İlgili genotip1 | Başvuru |
| DK1622 | Wildtype | 23 |
| SA4420 | ΔmglA | 24 |
| SA4749 | ΔmglA; parB+/attB:: PnatparB-yfp (pAH7) | Bu çalışmada |
| SA4797 | ΔmglA; ΔpomX / attB::PpomZ mCherry-pomX (pAH53) | 16 |
| SA8241 | ΔmglA; ftsZ+/ mxan18 / 19::PnatftsZ-gfp (pDS150) | Bu çalışmada |
| Plazmid parantez içinde belirtilen gen füzyon içeren ve genom üzerinde belirtilen sitelerde intergated. Plazmid attB sitesinde tümleşik veya mxan18-19 intergenic bölge üzerinden dile getirdi kendi yerel organizatörü (Pnat) veya pomZ (PpomZ) yerel organizatörü. |
||
Tablo 1: Bu çalışmada kullanılan bakteri suşları listesi.
| Plazmid | İlgili özellikleri | Başvuru |
| pAH7 | PnatparB-yfp; Mx8 attP; TetR | 19 |
| pAH53 | PpomZ mCherry-pomX; Mx8 attP ; KmR | 16 |
| pDS150 1 | PnatftsZ-gfp ; mxan18-19 ; TetR | Bu çalışmada |
| pMR3691 | Plazmid vanillate indüklenebilir gen ekspresyonu için | 18 |
| pKA51 | PnatftsZ-gfp ; Mx8 attP; TetR | 17 |
| 1 pDS150: pDS150 olduğunu hangi Mx8 attP sitesi ile mxan18-19 intergenic bölge yerine pKA51 bir türevi. Bunun için mxan18-19 intergenic bölge üzerinden pMR3691 Mxan18-19 fwd BsdRI astar ile güçlendirilmiş yapıldı. (GCGATCATTGCGCGCCAGACGATAACAGGC) ve Mxan18-19 rev BlpI (GCGGCTGAGCCCGCGCCGACAACCGCAACC) ve pKA51 içine klonlanmış. |
||
Tablo 2: Bu çalışmada kullanılan plazmid listesi.
Discussion
Floresans canlı hücre görüntüleme bakteri hücreleri kronolojik zamanmekansal dinamiklerini incelemek için güçlü bir araç haline gelmiştir. Hızlandırılmış floresans mikroskobu gibi büyüyen bakterilerin hareketli ve yavaş M. xanthus, ancak, zor olmuştur ve sadece kısa bir süre süreleri için gerçekleştirildi. Burada, canlı hücre görüntüleme için kullanımı kolay ve güçlü bir yöntemi mevcut M. xanthus tarafından hızlandırılmış floresans mikroskobu. Bu yöntem hücreleri ve hücre döngüsü tek hücre çözünürlük ile birkaç tur için fluorescently etiketli proteinler izleyin düzenlemesine olanak sağlar.
Yavaş büyüyen canlı hücre görüntüleme başarısını etkileyen birkaç önkoşul M. xanthus hücreleri de dahil olmak üzere: 1) bir katı yüzey için hücre Eki; 2) besin ve oksijen durumu; 3) sürekli nem ve sıcaklık; ve 4) maruz kalma süresi ve görüntü edinme frekans gibi deneysel koşullar optimizasyonu.
Deneysel yapımız besin ile desteklenmiş kalın özel yastıkları kullanın. Tek hücre takip etmek mikrosıvısal cihazlar aksine kalın özel pedleri kullanarak bazı temel faydaları ama aynı zamanda bazı dezavantajları vardır. İlk olarak, özel yastık sağlar sadece bir yüzey için M. xanthus hücre eki ve hareket aynı zamanda büyüme için en az 24 saat için yeterli besin. İkinci olarak, atış analizleri fluorescently etiketli proteinlerin hücre içi yerelleştirme çalışma için yaygın olarak kullanılan özel yastıkları16,17,29aynı türde daha önce yapıldı kendine gel. Bu nedenle, ek çekim analizleri verilerden burada açıklanan yöntemi ile elde edilen verileri doğrudan karşılaştırılabilir. Üçüncü olarak, özel yastıkları kolayca değiştirilebilir ve antibiyotik veya CuSO4 gibi diğer takviyeleri ile takıma ve vanillate bu yaygın gen ifade indüksiyon18,30için kullanılır. Son olarak, hücre bir deney süresince formu microcolonies için izin çünkü bu kurulum da doğrudan hücre-hücre etkileşimleri analiz ediliyor belirli parametre üzerinde etkisini eğitim sağlar. Bu açıdan özellikle durumunda önemlidir M. xanthus çünkü bu bakteri birkaç kişiyi bağımlı etkileşimleri görüntüler. Bu yöntemin en büyük dezavantajı deneysel koşullar bir deneme süresi için önceden belirlenmiş olduğunu. Buna karşılık, mikrosıvısal aygıtları genellikle örneğin antibiyotikler31ekleyerek bir deney süresince deneysel koşullar değiştirilmesine izin ver.
Özgür yazılım paketleri (Örneğin, MicrobeJ, Oufti) otomatik olarak, tek hücre ve protein yerelleştirme tek tek hücreleri içinde analiz etmek kullanılabilir. Ancak, bu bilgisayar yazılımı are yalnızca tek hücre veya hücre küçük gruplar analizi için oldukça uygundur. Böylece, otomatik olarak burada açıklanan 24 h kayıtları için oluşturulan verileri çözümlemek için bir meydan okuma olarak kalır.
Özetle, yavaş büyüyen ile canlı hücre görüntüleme gerçekleştirmek için kullanımı kolay ve tekrarlanabilir bir protokol açıkladığımız M. xanthus bakteri. Basit özel besin takıma yastıkları büyümesi için en az 24 saat ayakta tutmak ve gözlemleyerek ve protein yerelleştirme ve birçok nesiller içinde tek hücre çözünürlük ile büyüme analiz için izin vermek yeterli olduğunu göstermektedir.
Disclosures
Yazarlar onlar rakip hiçbir mali çıkarları var bildirin.
Acknowledgments
Bu çalışma çerçevesinde Transregio 174 "kronolojik zamanmekansal dinamiklerini bakteri hücreleri, Alman Araştırma Konseyi (DFG) tarafından desteklenen" ve Max Planck toplum.
Materials
| Name | Company | Catalog Number | Comments |
| DMI6000B with AFC | Leica microsystems | 11888945 | Automated inverted widefield fluorescence microscope with adaptive focus control |
| Universal mounting frame | Leica microsystems | 11532338 | Stage holder for different sample sizes |
| HCX PL FLUOTAR 100x/1.30 oil PH3 | Leica microsystems | 11506197 | Phase contrast objective |
| Orca Flash 4.0 camera | Hamamatsu | 11532952 | 4.0 megapixel sCMOS camera for picture aquisition |
| Filter set TXR ET, k | Leica microsystems | 11504170 | Fluorescence filter set, Ex: 560/40 Em: 645/75 |
| Filter set L5 ET, k | Leica microsystems | 11504166 | Fluorescence filter set, Ex: 480/40 Em: 527/30 |
| Filter set YFP ET, k | Leica microsystems | 11504165 | Fluorescence filter set, Ex: 500/20 Em: 535/30 |
| ProScan III | Prior | H117N1, V31XYZEF, PS3J100 | Microscope automation controller with interactive control center |
| EL 6000 light source | Leica microsystems | 11504115 | External fluorescence light source |
| Incubator BLX Black | Pecon | 11532830 | Black incubation chamber surrounding the microscope |
| Tempcontrol 37-2 digital | Leica microsystems | 11521719 | Automated temperature control for incubation chamber |
| Gentmycin sulphate | Carl Roth | 0233.4 | Gentamycin |
| Oxytetracylin dihydrate | Sigma Aldrich | 201-212-8 | Oxytetracyclin |
| Kanamycin sulphate | Carl Roth | T832.3 | Kanamycin |
| Filtropur BT25 0.2 bottle top filter | Sarstedt | 831,822,101 | Bottle top filter for sterilization of buffers |
| Deckgläser | VWR | 630-1592 | Glass cover slip (60 x 22 mm, thickness: 0.7 mm) |
| Seakem LE agarose | Lonza | 50004 | Agarose for microscopy slides |
| Leica Metamorph AF | Leica microsystems | 11640901 | Microscope control software and software for picture analysis |
| Tetraspeck Microsperes, 0.5 µm | ThermoFisher | T7281 | Fluorescent microspheres |
| petri dish | Greiner Bio-one | 688102 | 120 mm x 120 mm x 17 mm squared petri dish for agarose pads |
| BD Bacto Casitone | Becton Dickinson | 225930 | Casitone |
| Parafilm M | VWR | 291-1213 | Parafilm |
| Tris(hydroxymethyl)-aminomethane | Carl Roth | AE15.2 | Tris |
| Magnesium sulphate heptahydrate | Carl Roth | P027.2 | Magnesium sulphate |
| Potassium dihydrogen phosphate p.a. | Carl Roth | 3904.1 | Potassium dihydrogen phosphate |
| 1% CTT medium: 1 % (w/v) BD Bacto™ casitone, 10 mM Tris-HCl ph 8.0, 1 mM potassium phosphate buffer pH 7.6, 8 mM MgSO4 | Cultivation medium for M.xanthus | ||
| TPM buffer: 10 mM Tris-HCl ph 8.0, 1 mM potassium phosphate buffer pH 7.6, 8 mM MgSO4 | Buffer for preparation of microscopy slides for M.xanthus |
References
- Shapiro, L., McAdams, H. H., Losick, R. Why and how bacteria localize proteins. Science. 326 (5957), 1225-1228 (2009).
- Treuner-Lange, A., Søgaard-Andersen, L. Regulation of cell polarity in bacteria. J Cell Biol. 206 (1), 7-17 (2014).
- Laloux, G., Jacobs-Wagner, C. Spatiotemporal control of PopZ localization through cell cycle-coupled multimerization. J Cell Biol. 201, 827-841 (2013).
- Rudner, D. Z., Losick, R. Protein subcellular localization in bacteria. Cold Spring Harb. Perspect. Biol. 2 (4), 000307 (2010).
- Badrinarayanan, A., Le, T. B. K., Laub, M. T. Bacterial chromosome organization and segregation. Annu Rev Cell Dev Biol. 31 (1), 171-199 (2015).
- Munoz-Dorado, J., Marcos-Torres, F. J., Garcia-Bravo, E., Moraleda-Munoz, A., Perez, J. Myxobacteria: Moving, Killing, Feeding, and Surviving Together. Front Microbiol. 7, 781 (2016).
- Berleman, J. E., Kirby, J. R. Deciphering the hunting strategy of a bacterial wolfpack. FEMS Microbiol Rev. 33 (5), 942-957 (2009).
- Konovalova, A., Petters, T., Søgaard-Andersen, L. Extracellular biology of Myxococcus xanthus. FEMS Microbiol. Rev. 34, 89-106 (2010).
- Nudleman, E., Wall, D., Kaiser, D. Cell-to-cell transfer of bacterial outer membrane lipoproteins. Science. 309, 125-127 (2005).
- Vassallo, C. N., et al. Infectious polymorphic toxins delivered by outer membrane exchange discriminate kin in myxobacteria. eLife. 6, 29397 (2017).
- Vassallo, C., et al. Cell rejuvenation and social behaviors promoted by LPS exchange in myxobacteria. Proc Natl Acad Sci USA. 112 (22), 2939-2946 (2015).
- Li, Y., et al. Extracellular polysaccharides mediate pilus retraction during social motility of Myxococcus xanthus. Proc. Natl. Acad. Sci. USA. 100, 5443-5448 (2003).
- Kim, S. K., Kaiser, D. Cell alignment required in differentiation of Myxococcus xanthus. Science. 249, 926-928 (1990).
- Lobedanz, S., Søgaard-Andersen, L. Identification of the C-signal, a contact dependent morphogen coordinating multiple developmental responses in Myxococcus xanthus. Genes Dev. 17, 2151-2161 (2003).
- Schumacher, D., Søgaard-Andersen, L. Regulation of cell polarity in motility and cell division in Myxococcus xanthus. Annu Rev Microbiol. 71 (1), 61-78 (2017).
- Schumacher, D., et al. The PomXYZ proteins self-organize on the bacterial nucleoid to stimulate cell division. Dev Cell. 41 (3), 299-314 (2017).
- Treuner-Lange, A., et al. PomZ, a ParA-like protein, regulates Z-ring formation and cell division in Myxococcus xanthus. Mol Microbiol. 87 (2), 235-253 (2013).
- Iniesta, A. A., Garcia-Heras, F., Abellon-Ruiz, J., Gallego-Garcia, A., Elias-Arnanz, M. Two systems for conditional gene expression in Myxococcus xanthus inducible by isopropyl-beta-D-thiogalactopyranoside or vanillate. J Bacteriol. 194 (21), 5875-5885 (2012).
- Harms, A., Treuner-Lange, A., Schumacher, D., Søgaard-Andersen, L. Tracking of chromosome and replisome dynamics in Myxococcus xanthus. reveals a novel chromosome arrangement. PLoS Genet. 9 (9), 1003802 (2013).
- Iniesta, A. A. ParABS system in chromosome partitioning in the bacterium Myxococcus xanthus. PLoS One. 9 (1), 86897 (2014).
- Lin, L., Osorio Valeriano, M., Harms, A., Søgaard-Andersen, L., Thanbichler, M. Bactofilin-mediated organization of the ParABS chromosome segregation system in Myxococcus xanthus. Nat Commun. 8 (1), 1817 (2017).
- Hodgkin, J., Kaiser, D. Cell-to-cell stimulation of movement in nonmotile mutants of Myxococcus. Proc Natl Acad Sci U S A. 74 (7), 2938-2942 (1977).
- Kaiser, D. Social gliding is correlated with the presence of pili in Myxococcus xanthus. Proc Natl Acad Sci USA. 76 (11), 5952-5956 (1979).
- Miertzschke, M., et al. Structural analysis of the Ras-like G protein MglA and its cognate GAP MglB and implications for bacterial polarity. EMBO J. 30 (20), 4185-4197 (2011).
- Hodgkin, J., Kaiser, D. Genetics of gliding motility in Myxococcus xanthus. (Myxobacterales): Two gene systems control movement. Mol Gen Genet. 171, 177-191 (1979).
- Shi, X., et al. Bioinformatics and experimental analysis of proteins of two-component systems in Myxococcus xanthus. J Bacteriol. 190 (2), 613-624 (2008).
- Bi, E. F., Lutkenhaus, J. FtsZ ring structure associated with division in Escherichia coli. Nature. 354 (6349), 161-164 (1991).
- Lutkenhaus, J., Pichoff, S., Du, S. Bacterial cytokinesis: From Z ring to divisome. Cytoskeleton. 69 (10), 778-790 (2012).
- McLoon, A. L., et al. MglC, a Paralog of Myxococcus xanthus GTPase-Activating Protein MglB, Plays a Divergent Role in Motility Regulation. J Bacteriol. 198 (3), 510-520 (2015).
- Gomez-Santos, N., et al. Comprehensive set of integrative plasmid vectors for copper-inducible gene expression in Myxococcus xanthus. Appl Environ Microbiol. 78 (8), 2515-2521 (2012).
- Treuner-Lange, A., et al. The small G-protein MglA connects to the MreB actin cytoskeleton at bacterial focal adhesions. J Cell Biol. 210 (2), 243-256 (2015).
