

Summary
هذا البروتوكول بالتفصيل الخطوات، والتكاليف، والمعدات اللازمة لتوليد كولاي-على أساس مقتطفات الخلية وتنفذ في المختبر البروتين التوليف التفاعلات في غضون 4 أيام أو أقل. للاستفادة من الطابع المرن لهذا النظام الأساسي للتطبيقات الواسعة، ونحن نناقش رد فعل الظروف التي يمكن تكييفها والأمثل.
Abstract
على مدى السنوات الخمسين الماضية، برز توليف بروتين الخلية الحرة (الحراجية) كتقنية قوية لتسخير قدرات الخلايا داخل أنبوب اختبار النسخي ومتعدية الجنسيات. تنتفي الحاجة إلى الحفاظ على بقاء الخلية، وإزالة الجدار الخلوي، الحراجية كانت التأسيسية للتطبيقات المستجدة في التحويلية بروتينات الصعبة تقليديا، فضلا عن التطبيقات في النماذج الأولية السريعة الهندسة الأيضية، وعلم الجينوم الوظيفي. لدينا الوسائل لتنفيذ كولاي-أساس الحراجية منصة تسمح للمستخدمين الجدد الوصول إلى العديد من هذه التطبيقات. هنا، نحن تصف أساليب إعداد مقتطف من خلال استخدام وسائل الإعلام المخصب وقوارير حيرة واستنساخه بطريقة تحلل الخلية على أساس sonication الانضباطي. يمكن ثم استخدام هذا المقتطف للتعبير البروتين قادرة على إنتاج 900 ميكروغرام/مل أو أكثر من مجلد سوبر أخضر نيون البروتين (سفجفب) في ح 5 فقط من الإعداد التجريبية لتحليل البيانات، ونظرا لأن مخزون الكاشف المناسب قد أعدت مسبقاً. بدء التشغيل المقدرة تكلفة الحصول على الكواشف هو $4,500 الذي سيتم الحفاظ على الآلاف من ردود أفعال بتكلفة تقدر دولار 0.021 الواحدة ميكروغرام بروتين التي تنتجها أو $0.019 كل ميليلتر من رد الفعل. بالإضافة إلى ذلك، تعكس أساليب التعبير البروتين سهولة الإعداد رد فعل ينظر في النظم المتاحة تجارياً بسبب الاستغلال الأمثل يمزج قبل الكاشف، في جزء صغير من التكلفة. لتمكين المستخدم من الاستفادة من الطابع المرن لمنهاج تطبيقات واسعة الحراجية، حددنا مجموعة متنوعة من جوانب المنهاج التي يمكن ضبطها والأمثل اعتماداً على الموارد المتاحة ونتائج تعبير البروتين المرجوة.
Introduction
وبرز توليف بروتين الخلية الحرة (الحراجية) تكنولوجيا التي قد مقفلة عددا من الفرص الجديدة لإنتاج البروتين والجينوم الوظيفي والهندسة الأيضية وأكثر في غضون 50 عاماً الماضية1،2. بالمقارنة مع القياسية في فيفو منابر التعبير البروتين، الحراجية توفر ثلاث مزايا رئيسية: 1) طبيعة الخلية خالية من المنصة تتيح إنتاج البروتينات التي يحتمل أن تكون السمية أو الخارجية ل الخلية3،4 ،،من56؛ 2) المنظمة للحمض النووي والأخذ بقالب الحمض النووي ترميز gene(s) لمصلحة قناة كل الطاقة الشاملة في رد فعل لإنتاج protein(s) الفائدة؛ و 3) الطبيعة المفتوحة للمنصة تمكن المستخدم من تعديل ورصد رد فعل الظروف والتكوين في الوقت الحقيقي7،8. يدعم هذا الوصول المباشر إلى رد فعل زيادة النظم البيولوجية مع كيمياء الموسعة وظروف الأكسدة والاختزال لإنتاج البروتينات رواية وضبط العمليات الأيضية2،9، 10-توجيه الوصول كما يسمح للمستخدم بالجمع بين فعل الحراجية مع فحوصات النشاط في نظام واحد-وعاء لزيادة سرعة التصميم-البناء-اختبار دورات11. القدرة على القيام برد فعل الحراجية في قطيرات صغيرة الحجم أو على الأجهزة المستندة إلى الورق كذلك يدعم جهود الاكتشاف الفائق والنماذج الأولية السريعة12،،من1314،15 ،16. ونتيجة لهذه المزايا وطبيعة النظام التوصيل والتشغيل، مكن الحراجية فريد مجموعة متنوعة من تطبيقات التكنولوجيا الحيوية مثل إنتاج البروتينات التي يصعب على التعبير عن سلوبلي في فيفو17، 19،،من 1820، والكشف عن المرض21،،من2223، على الطلب التحويلية18،24 ،25،،من2627، والتعليم28،29، كل منها التحلي بالمرونة وفائدة من منصة خالية من الخلية.
يمكن إنشاء النظم الحراجية من مجموعة متنوعة من ليساتيس الخام من كلا خطوط الخلية بدائية النواة وحقيقية النواة. يسمح هذا للحصول على خيارات متنوعة في النظام للاختيار، وكل منها لديها مزايا وعيوب اعتماداً على التطبيق للفائدة. كما تتفاوت النظم الحراجية في إعداد الوقت والتكلفة والإنتاجية. الأكثر شيوعاً المستخدمة خلية تنتج مقتطفات من جرثومة القمح وارنب خلية شبكية وخلايا الحشرات وخلايا الإشريكيّة القولونية ، مع كونه الأخير الأكثر فعالية من حيث التكلفة حتى الآن بينما تنتج غلات الحجمي أعلى من البروتين30 . بينما النظم الحراجية الأخرى يمكن أن يكون مفيداً لجهازهم التعديل بوستترانسلاشونال الفطرية، التطبيقات استخدام كولايالناشئة-استناداً إلى الآلية قادرة على سد الفجوة عن طريق توليد سيتيسبيسيفيكالي فوسفوريلاتيد و الغليكوزيلاتي البروتينات على الطلب31،،من3233،،من3435.
يمكن تشغيل الحراجية ردود الفعل أما دفعة واحدة، التبادل المستمر خلية خالية (سيكف) أو تنسيقات (CFCF) الخلية الحرة تدفق مستمر. تنسيق دفعة هو نظام مغلق عمر رد الفعل الذي محدود بسبب تناقص كميات كواشف مختبر وتراكم تركات المثبطة لرد الفعل. أساليب سيكف و CFCF زيادة العمر من رد فعل، ومما يسفر عن البروتين الحجمي زيادة غلة مقارنة برد فعل المجموعة. يتم ذلك عن طريق السماح للمنتجات الثانوية تخليق البروتين المراد إزالتها من وعاء التفاعل في حين يتم توفيره كواشف مختبر جديد طوال فترة رد الفعل2. في حالة CFCF، يمكن أيضا إزالة بروتين الفائدة من دائرة رد الفعل، بينما في سيكف، البروتينات تظل الفائدة في دائرة رد الفعل تتألف من36،شبه نفاذية الأغشية37. هذه الأساليب ذات قيمة خاصة في التغلب على ضعف غلة الحجمي البروتينات للتعبير عن اهتمام38،39،،من4041،42، 43. التحديات التي يواجهها تنفيذ النهج سيكف و CFCF هي 1) في حين أنها تسفر عن استخدام أكثر كفاءة للآلية البيولوجية المسؤولة عن النسخ والترجمة، أنها تتطلب كميات أكبر لا سيما من الكواشف التي تزيد من التكلفة الإجمالية و 2) وهي تتطلب أكثر تعقيداً رد فعل الأجهزة والمعدات المتخصصة مقارنة ب تنسيق دفعة44. من أجل ضمان إمكانية الوصول للمستخدمين الجدد، توضحها البروتوكولات التركيز على شكل دفعة في مجلدات رد فعل من 15 ميليلتر مع توصيات محددة من أجل زيادة حجم رد فعل لمقياس المليلتر.
الأساليب المقدمة هنا تمكين غير الخبراء ذوي المهارات المختبرية الأساسية (مثل طلاب المرحلة الجامعية) تنفيذ نمو الخلايا واستخراج إعداد الدفعة تنسيق إعداد رد فعل كولاي-استناداً إلى نظام الحراجية. وهذا النهج فعالة من حيث التكلفة مقارنة بمجموعات المتاحة تجارياً دون التضحية بالسهولة لإعداد رد فعل يستند إلى مجموعة. وعلاوة على ذلك، يمكن هذا النهج التطبيقات في المختبر وفي الميدان. المستخدمين الجدد تماما عند اتخاذ قرار لتنفيذ الحراجية، ينبغي تقييم فعالية أنظمة تعبير البروتين التقليدية للاستثمار بدء التشغيل، الحراجية قد لا تكون متفوقة في كل حالة. الطرق الحراجية الموضحة هنا تمكين المستخدم لمباشرة تنفيذ مجموعة متنوعة من التطبيقات، بما في ذلك الجينوم الوظيفي، الاختبار، وإنتاج البروتينات التي مستعصية على الحل للتعبير في فيفو ، فضلا عن الحقل الفائق التطبيقات بما في ذلك أجهزة استشعار العوامل البيولوجية ومواد تعليمية للبيولوجيا التركيبية. تطبيقات إضافية مثل الهندسة الأيضية وضبط شروط التعبير البروتين والكشف عن المرض، والارتقاء باستخدام أساليب سيكف أو CFCF لا تزال ممكنة ولكن قد تتطلب خبرة مع منصة الحراجية لمزيد من التعديل من رد فعل شروط. لدينا طرق الجمع بين النمو في وسائل الإعلام المخصب وقوارير حيرة، مع طرق سريعة نسبيا واستنساخه من تحلل الخلية عن طريق سونيكيشن، متبوعاً بإعداد رد فعل الحراجية مبسطة التي تستخدم قبل المزيج الأمثل45. بينما قد تصبح أساليب النمو الخلوي موحدة إلى حد ما داخل هذا الحقل، طرق لتحلل الخلية تختلف على نطاق واسع. بالإضافة إلى سونيكيشن، وتشمل المشتركة تحلل أساليب الاستفادة من صحافة فرنسية والخالطون، وحبه المضارب، أو lysozyme وأخرى تعطل البيوكيميائية والفيزيائية أساليب46،47،48، 49-استخدام أساليب عملنا، ويتم الحصول على حوالي 2 مل استخراج النفط الخام الخلية الواحدة 1 لتر خلايا. ويمكن دعم هذه الكمية من استخراج خلية أربعمائة 15 ميليلتر الحراجية من ردود الفعل، وكل إنتاج ~ 900 ميكروغرام/مل من البروتين سفجفب مراسل من قالب بلازميد pJL1-سفجفب. هذا الأسلوب يكلف $0.021/ميكروغرام سفجفب المنتجة ($.019/ميليلتر من رد الفعل)، باستثناء تكلفة العمالة والمعدات (تكميلية الشكل 1). ابتداء من الصفر، هذا الأسلوب يمكن تنفيذها في 4 أيام في شخص واحد، وكرر الحراجية ردود فعل يمكن أن تنجز في غضون ساعات (الشكل 1). بالإضافة إلى ذلك، يمكن الارتقاء بالبروتوكول في تخزين لدفعات أكبر من إعداد كاشف لتناسب احتياجات المستخدم. الأهم من ذلك، يمكن تنفيذ البروتوكول المعروضة هنا غير مختبر تدريب الخبراء مثل طلاب المرحلة الجامعية، كما أنه يتطلب فقط المهارات المختبرية الأساسية. قد وضعت خصيصا لتحسين إمكانية الحصول على منصة الحراجية كولاي للاستخدام الواسع الإجراءات الواردة أدناه، والفيديو المصاحبة.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1-وسائل الإعلام إعداد ونمو الخلايا
-
1 يوم
- خلايا BL21*(DE3) كولاي متواصلة من والغليسيرول الأسهم على صفيحة أجار رطل واحتضانها لمالا يقل عن 18 ح عند 37 درجة مئوية.
- إعداد 50 مل الإعلام رطل والاوتوكلاف الحل على أساس دورة سائل لمدة 30 دقيقة في 121 درجة مئوية. تخزين في درجة حرارة الغرفة.
-
2 يوم
- إعداد 750 مل 2 x YTP الوسائط و 250 مل من 0.4 م د-الجلوكوز الحل كما هو موضح في المعلومات التكميلية.
- صب وسائط الإعلام x YTP 2 في قارورة حيرة يعقم 2.5 لتر والحل د-الجلوكوز في يعقم 500 مل زجاجة. اﻷوتوكﻻف كل الحلول على أساس دورة سائل لمدة 30 دقيقة في 121 درجة مئوية.
- التأكد من أن كل الحلول العقيمة مخزنة في 37 درجة مئوية إذا كان يتم إجراء نمو الخلايا في اليوم التالي، زيادة معدلات النمو على التطعيم. لا تجمع الحلول حتى التطعيم.
ملاحظة: يمكن تخزين الحلول عند 4 درجة مئوية عن د 1-2 إذا لزم الأمر، على الرغم من أن وسائل الإعلام x YTP 2 العالية المعرضة للتلوث. - ابدأ بثقافة بين عشية وضحاها من BL21(DE3) بتطعيم 50 مل الإعلام رطل مع مستعمرة واحدة من BL21(DE3) باستخدام حلقة معقمة وتقنية معقمة لتجنب التلوث.
- ضع 50 مل الثقافة BL21*(DE3) رطل في 37 درجة مئوية 250 دورة في الدقيقة تهتز حاضنة وتنمو بين عشية وضحاها على ح 15-18.
- تحضير وتعقيم جميع المواد اللازمة لأيام 3 و 4، بما في ذلك: زجاجتين من أجهزة الطرد المركزي ل 1، 4 x الباردة 50 مل مخروطية أنابيب (وزن والجماهير سجل ثلاثة)، والعديد من 1.5 مل [ميكروفوج] أنابيب.
-
3 يوم
- إزالة ثقافة بين عشية وضحاها 50 مل BL21*(DE3) رطل من حاضنة الهز وقياس OD600 على جهاز المطياف الضوئي استخدام 01:10 إضعاف مع وسائل الإعلام رطل. حساب حجم بين عشية وضحاها الثقافة اللازمة لإضافة إلى 1 لتر من وسائط الإعلام لانطلاق OD600 من 0.1 (على سبيل المثال، إذا كان OD600 من 01:10 إضعاف للقراءة ك 0.4، تطعيم 25 مل OD غير مخفف600 = 4.0 الثقافة بين عشية وضحاها إلى 1 لتر 2 x YTP ز).
- إزالة حرارة 2 x YTP وسائل الإعلام والحلول د-الجلوكوز من حاضنة 37 درجة مئوية جنبا إلى جنب مع 50 مل الثقافة رطل. بعناية باستخدام تقنية تعقيم، صب الحل د-الجلوكوز في وسائل الإعلام x YTP 2 (تجنب جانبي قارورة حيرة).
ملاحظة: إضافة د-الجلوكوز يكمل وصفه للأم 1 من 2 x YTPG. - الحفاظ على أسلوب تعقيم، تلقيح ل 1 2 x YTPG حل مع المقدار المناسب من ثقافة 50 مل تبدأ ثقافة 1 لتر في التطوير التنظيمي 0.1600. فورا بوضع ثقافة 1 لتر الملقحين في 37 درجة مئوية تهز الحاضنة 200 لفة في الدقيقة.
- تأخذ OD الأولى600 القراءة بعد الساعة الأولى من النمو (مرحلة انتقالية نموذجية يأخذ ح 1). لا تضعف الثقافة. مواصلة أخذ القياسات600 OD تقريبا كل 20-30 دقيقة حتى OD600 يبلغ 0.6.
- عند بلوغ OD600 = 0.6، أضف 1 مل 1 م إيبتج (التركيز النهائي في الثقافة ل 1 = 1 مم) لثقافة x YTPG 2.
ملاحظة: التعريف المثالي OD600 هو 0.6؛ مجموعة من 0.6-0.8 غير مقبولة. تحريض من إيبتج للإنتاج الذاتية من T7 رنا بوليميراز (T7RNAP). - بعد التعريفي، وقياس OD600 تقريبا كل 20-30 دقيقة حتى تصل إلى 3.0.
ملاحظة: يبرد الطرد المركزي إلى 4 درجات مئوية خلال هذا الوقت. إعداد المخزن المؤقت S30 الباردة كما هو مفصل في "المعلومات التكميلية". إذا كان المخزن المؤقت S30 مستعدة مسبقاً، ضمان أن لا يتم إضافة القيمة حتى يوم الاستخدام. - مرة واحدة OD600 يصل إلى 3.0 (الشكل 2A)، من أجل الثقافة في زجاجة 1 لتر للطرد مركزي باردة في حمام الماء المثلج. إعداد أجهزة الطرد مركزي 1 لتر مملوءة بالمياه زجاجة من وزن متساو لاستخدامها كتوازن في أجهزة الطرد المركزي.
ملاحظة: تختلف قيم امتصاص من الصك للصك. بينما OD600 من حصاد BL21(DE3) غير متغير حساسة، من المستحسن أن المستخدم تقييم وتحسين هذا المتغير كإجراء استكشاف الأخطاء وإصلاحها. قد يؤدي أقل نسبيا من القراءات600 OD الطيفية على أساس ومبومو أصغر بالمقارنة مع أكبر الطيفية. - الطرد المركزي الزجاجات 1 لتر لمدة 10 دقيقة مبلغ 000 5 x ز و 10 درجات مئوية بيليه الخلايا.
- صب ببطء بعيداً عن المادة طافية والتخلص منه وفقا لإجراءات النفايات البيولوجية للمؤسسة. مكان بيليه على الجليد.
- باستخدام ملعقة عقيمة، كشط بيليه الخلية من الزجاجة للطرد المركزي وتحويلها إلى أنبوب مخروطي باردة 50 مل.
- إضافة 30 مل من البرد S30 العازلة للأنبوبة المخروطية وريسوسبيند بيليه الخلية التي فورتيكسينج مع رشقات نارية قصيرة (20-30 ثانية) وفترات الراحة (1 دقيقة) على الجليد حتى حراكه تماما مع لا قطع.
- بمجرد الكامل هو حراكه بيليه، استخدام أنبوب مخروطي 50 مل آخر مع المياه كالتوازن والطرد المركزي لمدة 10 دقائق في 5000 × ز و 10 درجات مئوية (قبل تبريده إلى 4 درجات مئوية).
ملاحظة: هذا يكمل 1st ليغسل 3 المطلوبة عند حصاد الخلايا. - من أجل المادة طافية والتخلص منه وفقا لإجراءات النفايات البيولوجية للمؤسسة. ريسوسبيند بيليه مع 20-25 مل من البرد S30 المخزن المؤقت والطرد المركزي لمدة 10 دقائق في 5000 س ز و 10 درجات مئوية (قبل تبريده إلى 4 درجات مئوية).
ملاحظة: هذا يكمل 2nd ليغسل 3. - ومرة أخرى، من أجل المادة طافية والتخلص منه وفقا لإجراءات النفايات البيولوجية للمؤسسة. إضافة بالضبط 30 مل من المخزن المؤقت S30 ودوامه مرة أخرى ريسوسبيند بيليه.
- استخدام الأنابيب المخروطية قبل موزون، الباردة 50 مل 3 وحشو ماصة مصلية ماصة معقمة، الكوة 10 مل مخلوط حراكه بيليه/S30 المخزن المؤقت في كل من أنابيب مخروطية الشكل 3.
ملاحظة: تقسيم الخلايا في أنابيب 3 غير مطلوب، ولكن نتائج هذه الخطوة في أصغر خلية الكريات (~ 1 ز) لزيادة الراحة في خطوات لاحقة. - الطرد المركزي جميع الأنابيب، باستخدام الأرصدة المناسبة حسب الحاجة، لمدة 10 دقيقة على 5000 × ز و 10 درجات مئوية (قبل تبريده إلى 4 درجات مئوية).
ملاحظة: هذا يكمل الخطوة يغسل النهائي. - من أجل المادة طافية والتخلص منه وفقا لإجراءات النفايات البيولوجية للمؤسسة. إزالة المخزن المؤقت S30 الزائدة عن طريق المسح بعناية داخل الأنبوبة المخروطية وكاب مع أنسجة نظيفة؛ تجنب لمس بيليه.
- روي الأنابيب على توازن التحليلي وتسجيل وزن بيليه النهائي على كل أنبوبة.
ملاحظة: يمكن أن يكون مؤقتاً في البروتوكول في هذه المرحلة. يمكن أن تكون الكريات فلاش المجمدة في النتروجين السائل والمخزنة في-80 درجة مئوية لتصل إلى سنة حتى اللازمة لإعداد استخراج.
2-النفط الخام الخلية استخراج إعداد-يوم 4
- لإعداد استخراج، إبقاء الخلايا الباردة على الجليد خلال كل خطوة. أضف 1 مل من المخزن المؤقت S30 الباردة كل 1 غرام كتلة الخلية لبيليه. التأكد من أن ديثيوثريتول (DTT) قد استكملت إلى المخزن المؤقت S30 لتركيز نهائي من 2 مم.
ملاحظة: يبرد ميكروسينتريفوجي إلى 4 درجات مئوية خلال هذا الوقت. - ريسوسبيند بيليه الخلية التي فورتيكسينج مع رشقات نارية قصيرة (20-30 ثانية) وفترات الراحة (1 دقيقة) على الجليد حتى حراكه تماما. إذا كان من الصعب استثارة، ترك الكريات على الجليد لمدة 30 دقيقة تذويب.
- نقل 1.4 مل خلايا حراكه في أنبوب 1.5 مل [ميكروفوج].
- ضع أنبوب 1.5 مل واحد يحتوي على 1.4 مل خلايا حراكه في حمام الماء المثلج في كوب. Sonicate ل 45 ق على تليها 59 مجموعة s إيقاف لدورات مجموع 3، مع السعة 50 في المائة. قم بإغلاق وعكس الأنابيب المزيج بلطف أثناء فترات قبالة. وفي المجموع، تسليم ي 800-900 من الطاقة لكل 1.5 مل [ميكروفوج] أنابيب تحتوي على 1.4 مل حراكه الخلايا (الشكل 3 ألف و 3 باء).
ملاحظة: هذه الخطوة حساسة لنوع سونيكاتور ونموذج المستخدمة، وينبغي أن يكون الأمثل إذا كانت معدات مختلفة من سردها لهذا الإجراء. يمكن استخدام نهجين متكاملين للارتقاء بمبلغ استخراج أعدت خلال هذه الخطوة: 1.5 مل 1) متعددة [ميكروفوج] أنابيب يمكن أن تكون سونيكاتيد بالتوازي، و/أو يمكن أن سونيكاتيد 2) أكبر من وحدات التخزين في الأنابيب المخروطية (تصل إلى 15 مل استثارة الخلية الواحدة أنبوب) ، زيادة مقدار الطاقة الموردة كما تم وصفه سابقا 29،45. - فورا بعد اكتمال sonication إضافة ميليلتر 4.5 متر واحد DTT (المكمل إضافي 2 مم DTT) إلى 1.4 مل من ليستي وعكس عدة مرات لخلط. ضع الأنبوب على الجليد. كرر الخطوات من 2.4 و 2.5 لأي أنابيب إضافية من الخلايا حراكه قبل الشروع في استخدام الطرد المركزي.
- عينات ميكروسينتريفوجي في 18,000 س ز و 4 درجة مئوية لمدة 10 دقائق (الشكل 3).
- "الماصة؛" المادة طافية في أنبوب [ميكروفوج] 1.5 مل جديدة. عدم الإزعاج بيليه؛ فمن الأفضل أن تترك بعض المادة طافية الحفاظ على النقاء من عرقلة بيليه في الجهود الرامية إلى تحقيق أقصى قدر من الإنتاجية.
- احتضان المادة طافية من الخطوة السابقة في 250 دورة في الدقيقة و 37 درجة مئوية لمدة 60 دقيقة بتسجيل هذه الأنابيب إلى منصة تهز الحاضنة (وهذا هو رد فعل الجريان السطحي).
- عينات ميكروسينتريفوجي 10 آلاف س ز و 4 درجة مئوية لمدة 10 دقائق.
- إزالة المادة طافية دون إزعاج بيليه وتحويلها إلى أنبوب جديد. إنشاء العديد من مختبرين ميليلتر 100 من استخراج للتخزين.
ملاحظة: يمكن إيقاف البروتوكول هنا، ويمكن أن يكون الاستخراج فلاش المجمدة في النتروجين السائل وتخزينه في-80 درجة مئوية لتصل إلى سنة حتى بحاجة لردود الفعل الحراجية. يمكن مرت على الأقل 5 تجميد أذاب دورات دون المساس باستخراج الإنتاجية (الشكل 4).
3-خالية من الخلية البروتين التوليف دفعة تنسيق ردود الفعل
- ذوبان الجليد بحلول وب والحمض النووي القالب، BL21(DE3) استخراج (إذا كانت مجمدة)، T7RNAP وقاسمه المياه الصف الجزيئية.
ملاحظة: يمكن الاطلاع على قالب رد فعل الحراجية في المعلومات التكميلية. وترد معلومات تكميلية وصفات الحلول A و B وتتوافق مع تركيزات محددة للعديد من الكواشف لدعم نظام الطاقة بانوكس-ليرة سورية على أساس الحراجية. وكانت دور كل كاشف والاختلاف مقبولاً في هذه التركيزات المتفاعلة التي يمكن أن تدعم الحراجية العزم50. بروتوكول تنقية T7RNAP ويمكن الاطلاع على "معلومات تكميلية"51. T7RNAP تكميلية يمكن أن يزيد الغلة الحجمي ولكن ليس من الضروري إذا هو فعل T7RNAP أثناء نمو الخلايا. يمكن إعداد قالب بلازميد الحمض النووي (pJL1-سفجفب) باستخدام مجموعة أدوات ماكسيبريب مع يغسل اثنين باستخدام المخزن المؤقت الغسيل في هذه المجموعة، تليها بعد تجهيز الحمض النووي--تنظيف باستخدام مجموعة أدوات تنقية بكر (الشكل 2). يمكن أيضا استخدام القوالب الحمض النووي الخطي في ردود الفعل الحراجية. - قم بتسمية المبلغ اللازم من [ميكروفوج] أنابيب بحاجة لردود الفعل الحراجية.
ملاحظة: ردود فعل يمكن أن يؤديها في أحجام مختلفة من السفينة، ولكن سفينة أصغر يمكن إنقاص البروتين الحجمي غلة (الشكل 2). الارتقاء بمستوى رد فعل في نفس حجم السفينة قد يقلل أيضا من غلة الحجمي، كدالة لتناقص تبادل الأوكسجين، نتيجة انخفاض مساحة السطح إلى نسبة الحجم. عند زيادة حجم الرد أعلاه 100 ميكروليتر، من المستحسن استخدام مسطحة القاع جيدا لوحات 31،،من3752. - إضافة ميليلتر 2.2 الحل A، ميليلتر 2.1 من "حل ب"، 5 ميليلتر من BL21*(DE3) استخراج، 0.24 ميكروغرام من T7RNAP (تركيز ميكروغرام/مل 16 النهائي) 0.24 نانوغرام من قالب الحمض النووي (16 نانوغرام/مليلتر تركيز النهائي)، والمياه لجعل وحدة التخزين النهائي 15 ميليلتر.
ملاحظة: الحلول دوامة A و B كثيرا أثناء إعداد رد فعل تجنب ترسب مكونات، وضمان حصول كل رد فعل قاسمة متجانسة لكل حل. تجنب فورتيكسينج الاستخراج، بدلاً من ذلك عكس أنبوب للمزيج. - بعد أن تم إضافة جميع الكواشف إلى رد فعل، خلط كل أنبوبة بيبيتينج صعودا وهبوطاً أو بلطف فورتيكسينج مع ضمان أن الخليط الرد النهائي هو دمج حبة 15 ميليلتر واحد في الجزء السفلي من 1.5 مل [ميكروفوج] أنابيب.
- ضع كل رد فعل إلى حاضنة 37 درجة مئوية دون هز ح 4، أو 30 درجة مئوية بين عشية وضحاها.
ملاحظة: ردود الفعل ناجحة يمكن نوعيا تقييم بصريا استناداً إلى اللون الأخضر للمنتج سفجفب داخل الخليط رد فعل الحراجية (3D الشكل). يمكن أيضا تأكيد التعبير عن بروتين اهتمام بالحزب الديمقراطي الصربي صفحة (تكميلية الشكل 2).
4-تحديد مقدار "البروتين مراسل"، [سفجفب]
- تحميل 48 ميليلتر من 0.05 متر مكعب حبيس، درجة الحموضة 8، إلى كل هناك حاجة أيضا للقياس الكمي (وعادة ما يقوم في ثلاث نسخ كل أنبوب رد فعل).
- إزالة ردود الفعل من الحاضنة. "الماصة؛" صعودا وهبوطاً لخلط كل رد فعل، ثم نقل 2 ميليلتر من رد الفعل إلى 48 ميليلتر من 0.05 متر مكعب هيبيس، درجة الحموضة 8. بيبيت صعودا وهبوطاً مرة أخرى في البئر للمزيج.
- حالما يتم تحميل جميع ردود الفعل ومختلطة، ضع لوحة جيدا 96 إلى فلوروميتير وقياس fluorescence نقطة النهاية سفجفب.
ملاحظة: أطوال موجية الإثارة وانبعاث سفجفب fluorescence التقدير الكمي هي 485 نانومتر و 510 نيوتن متر، على التوالي. - باستخدام منحنى قياسي الذي تم إنشاؤه مسبقاً، تحديد [سفجفب] من قراءات الأسفار التي تم الحصول عليها.
ملاحظة: تقدم تعليمات لإنشاء منحنى قياسي لتركيز سفجفب مقابل كثافة fluorescence في "المعلومات التكميلية" (تكميلية الشكل 3). سوف يحتاج المستخدمون إنشاء منحنى قياسي للصك حيث قد تختلف حساسية الصك.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
وقد قدمنا على أساس سونيكيشن كولاي استخراج إعداد البروتوكول التي يمكن إنجازها على مدى أربعة أيام، مع الرقم 1 يدل انهيار الإجرائية أكثر من كل يوم. وهناك قابلية تطويع للخطوات التي يمكن أن تنجز في كل يوم مع مختلف نقاط التوقف، ولكن وجدنا سير العمل هذا لتكون الأكثر فعالية لتنفيذ. بالإضافة إلى ذلك، كل خلية الكريات (الخطوة 1.3.18) واستخراج استعداد تام (الخطوة 2.10) مستقرة في-80 درجة مئوية لمدة سنة على الأقل، مما يسمح للمستخدم بإنشاء أكبر مخزون من كل منهما لإنقاذ لاستخدامها في وقت لاحق17. يتم الاستخراج مستقرة على مدى فترات زمنية طويلة، بل يمكن أيضا الخضوع للاستخراج دورات ذوبان التجميد خمسة على الأقل دون خسارة كبيرة للإنتاجية (الشكل 4). هذا يسمح لأكبر مختبرين لاستخراج ليتم تخزينها لاستخدامات متعددة إذا كانت مساحة التخزين المجمد محدودة. على الرغم من ذلك، نوصي مختبرين أصغر متعددة (~ 100 ميكروليتر) لاستخراج كلما كان ذلك ممكناً.
مع كل إعداد استخراج جديدة، نوصي بأن يقوم المستخدم معايرة المغنيسيوم بغية تحديد كمية المغنيسيوم لأن الدفعة من استخراج الأمثل. يمكن تحديد المستخدمين تقلب دفعة إلى دفعة في تركيز البروتين الكلي من استخراج الخلية بمقايسة برادفورد. أعلى أداء مقتطفات، نرى عادة تركيزات البروتين الكلي من 30-50 ملغ/مل، وضمن هذا النطاق هناك لا علاقة متبادلة واضحة بين تركيزات البروتين الكلي والأداء استخراج الخلية. ولذلك، نوصي أن المستخدمين النغمة تركيزات المغنيسيوم وبالتالي لضمان أن يتم تكبير البروتين ووظيفة الحمض النووي لكل دفعة استخراج. مستويات المغنيسيوم هامة للنسخ المتماثل الحمض النووي السليم والنسخ والترجمة، ولكن المستويات المفرطة يمكن أن تكون ضارة لهذه العمليات53. من أجل إثبات هذه التبعية، نحن أدوا معايرة المشارك من المغنيسيوم واستخراج وحدة التخزين لتحديد أفضل مزيج مما يقلل كمية استخراج اللازمة، مع الحفاظ على فعل إنتاجية (الشكل 5). من هذه التجربة، ونحن نوصي باستخدام 5 ميليلتر لاستخراج وملم 10 ملغ2 + لاستخراج بمحتوى بروتين الكلي من 30 ملغ/مل، بغية الحصول على أكثر من 1,000 ميكروغرام/مل من سفجفب.
أن تجربتنا مع الحراجية كما أتاحت لنا لتحديد الخطوات في إطار البروتوكول التي يمكن أن تختلف دون المساس بالإنتاجية العامة للنظام، والبعض الآخر تشكل جزءا لا يتجزأ من نظام الحراجية أداء عالية. وأبرزها التطوير التنظيمي النهائي600 من الحصاد الخلية لا تؤثر تأثيراً كبيرا على الناتج النهائي لرد فعل الحراجية، وعملياً يمكن أن تحصد الخلايا في أي مكان من 2.7 4.0 OD600. وهذا يمثل المرحلة الأسية المبكرة من النمو حيث أعلى تركيز الريبوسوم كل خلية وأكثر نشاطا لدعم النمو السريع إليه متعدية الجنسيات. هذه الملاحظة تسمح للمستخدمين المرونة اللازمة لتحسين إجراءاتها الخاصة. نوصي بالحصاد في حوالي 3.0 OD600 بغية التقاط الخلايا التطوير التنظيمي600 أقرب إلى 3.3 بحصاد الوقت الكامل (الشكل 2A). وتشمل المتغيرات التي تؤثر على غلة الحراجية جودة الحمض النووي قالب ورد فعل السفينة حجم الكميات النسبية من الخلية استخراج والمغنيسيوم أيون موجودين في رد فعل. وقد وجدنا الجودة الحمض النووي أن يكون التباين دفعة لدفعة ملحوظة. لحل هذه المشكلة، نوصي أن المستخدمين تنقية الحمض النووي عن طريق ميدي أو ماكسي الإعدادية، تليها خطوة تنظيف إضافية من الحمض النووي أما على عمود تنقية الحمض النووي المستخدمة في ماكسيبريب، أو تنقية بعد استخدام مجموعة إضافية من مواد تنظيف الحمض النووي. وهذا يحسن من إمكانية تكرار نتائج في نوعية الحمض النووي لردود الفعل الحراجية والنتائج في إنتاج البروتين أكثر قوة (الشكل 2). كما يؤثر السفينة رد فعل غلة الحجمي، حيث أن إنتاج البروتين من رد فعل مماثل على الأجهزة في وحدات التخزين السفينة متفاوتة يمكن أن تختلف نسبة تصل إلى 40%. وقد اقتنع أن زيادة في الغلة الحجمي لوحظت في السفن الأكبر حجماً بسبب زيادة منطقة سطح الخليط رد فعل، مما يسمح لتبادل الأوكسجين أفضل (الشكل 2)، وآخرين عززت زيادة غلة الحجمي بتشغيل ردود الحراجية في لوحات مسطحة القاع الكبيرة، التي نوصي لردود الفعل على مدى 100 ميليلتر17،،،من3137،52.

الشكل 1 : الجدول الزمني لنمو الثقافة، والإنتاج لاستخراج الخلية والإعداد والتقدير الكمي لردود الفعل الحراجية. المستخدم يمكن تنفيذ منهاج الحراجية لتطبيقاتها البحثية من خلال سير العمل هذا في أربعة أيام. إعداد كاشف تمثل الوقت الأساسي وتكلفة الاستثمار للجولة الأولى من هذه التجربة، ويقلل إلى حد كبير بعد إنشاء مخزون الكواشف. بالإضافة إلى ذلك، يمكن تخزين حبيبات الخلية واستخراج خلايا المعدة لأكثر من سنة في-80 درجة مئوية، مما يتيح للمستخدم لبدء الفترة الزمنية في الخطوات المختلفة لنتائج أسرع. يمكن أيضا إيقاف المستخدم في مختلف الخطوات لتعديل الجدول الزمني لسير العمل هذا. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الشكل 2 : قابل للتعديل الشروط الحراجية، والآثار على غلة رد فعل الحجمي. ألف- مقتطفات مقارنة إنتاجية تستند إلى حصاد الخلايا BL21(DE3) في مختلف القراءات600 OD. وبناء على هذه الأرض، نوصي بالحصاد OD600 3-3 تنتج على الأقل 1000 ميكروغرام/مل من البروتين المستهدف. ردود الفعل أجريت في نطاق ميكروليتر 15 في 1.5 مل [ميكروفوج] أنابيب. مقارنة ب الحمض النووي ماكسيبريب أغسل بروتوكولين مع أو بدون ما بعد تنقية الحمض النووي-التنظيف. وخضع والبلازميدات pJL1-سفجفب ماكسيبريب مع واحد أو اثنين من يغسل متبوعاً بتنظيف ما بعد تنقية طقم تنقية PCR. لتحقيق ~ 900 ميكروغرام/مل من البروتين التعبير، فإننا نقترح إجراء تنظيف بعد تنقية الحمض النووي بغض النظر عن عدد يغسل ماكسيبريب. ردود الفعل أجريت في نطاق ميكروليتر 15 في 1.5 مل [ميكروفوج] أنابيب. جيم 15 ميليلتر الحراجية من ردود الفعل المنجز في سفن مختلفة تتراوح من 2 مل إلى 0.6 مل [ميكروفوج] أنابيب. يمثل "الراتب" عنصر تحكم سلبية حيث تمت إضافة أي قالب الحمض النووي لرد الفعل. وتمثل كافة أشرطة الخطأ الانحراف المعياري 1 من ثلاثة ردود مستقلة لكل حالة، وكل منها كان كمياً في ثلاث نسخ. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الشكل 3 : استخراج مفتاح الأجهزة الإجرائية ونتائج لإنشاء المنتج. ألف الإعداد السليم لحمام الماء المثلج سونيكيشن لضمان التبريد من عينة بينما يتم إنشاء الحرارة أثناء sonication. ب 1.5 مل [ميكروفوج] أنابيب تحتوي على خلية حراكه بيليه قبل (يسار) ووظيفة (يمين) سونيكيشن. ليستي الناجمة عن ذلك ينبغي عرض لوناً أغمق مقارنة بالخلية حراكه بيليه. جيم- الفصل السليم للمادة طافية وبيليه خلية بعد 18,000 س ز الطرد المركزي. دال الحراجية ردود الفعل بعد ح 4 من الحضانة عند 37 درجة مئوية. 1.5 مل [ميكروفوج] أنابيب على اليمين (رد فعل ناجح) يبين fluorescence مرئية من البروتين مراسل سفجفب ~ 900 ميكروغرام/مل. أنبوب مراقبة سلبية على اليسار، وتفتقر إلى قالب الحمض النووي ومحاكاة فعل غير ناجحة، يعرض حلاً واضحا مع لا الأسفار. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الشكل 4 : استخراج التغيير في التعبير البروتين ما يزيد على 5 تجميد أذاب دورات الحراجية. استخراج إعداد من نفس النمو خضع خمسة تجميد دورات ذوبان الجليد عن طريق النيتروجين السائل فلاش تجميد اتباعها قبل ذوبان الجليد على الجليد. وشوهدت على مدى خمس دورات تجميد أذاب لا تغييرات كبيرة في الإنتاجية استخراج للتعبير عن سفجفب. ردود الفعل أجريت في نطاق ميكروليتر 15 في 1.5 مل [ميكروفوج] أنابيب. يمثل "الراتب" عنصر تحكم سلبية حيث تمت إضافة أي قالب الحمض النووي لرد الفعل. وتمثل كافة أشرطة الخطأ الانحراف المعياري 1 من ثلاثة ردود مستقلة لكل حالة، وكل منها كان كمياً في ثلاث نسخ. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-

الشكل 5 : الحراجية لردود الفعل مع اختلاف [مغ2+[واستخراج كميات مقابل [سفجفب]. [Mg2 +] تتراوح من 8 مم إلى 14 مم مع زيادات 2 مم واستخراج كميات تتراوح 3 ميليلتر ميليلتر 7 مع 1 ميليلتر زيادات. يمثل رمز اللون [سفجفب] ينتج من عالية (الأحمر) إلى الأدنى (الأرجواني). لتعظيم كفاءة الكاشف مع المحافظة على نسبة عالية من البروتين الإنتاج، نوصي باستخدام 5 ميليلتر لاستخراج وملم 10 ملغ2+ للمقتطفات التي يكون محتوى بروتين الكلي ~ 30 ملغ/مل، كما يحددها المقايسة برادفورد. واستندت النقاط الأصلي لتوليد هذه المؤامرة كفاف قبالة fluorescence نقطة النهاية من ثلاثة ردود مستقلة لكل حالة، وكل منها تم قياسها في ثلاث نسخ. ردود الفعل أجريت في نطاق ميكروليتر 15 في 1.5 مل [ميكروفوج] أنابيب. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
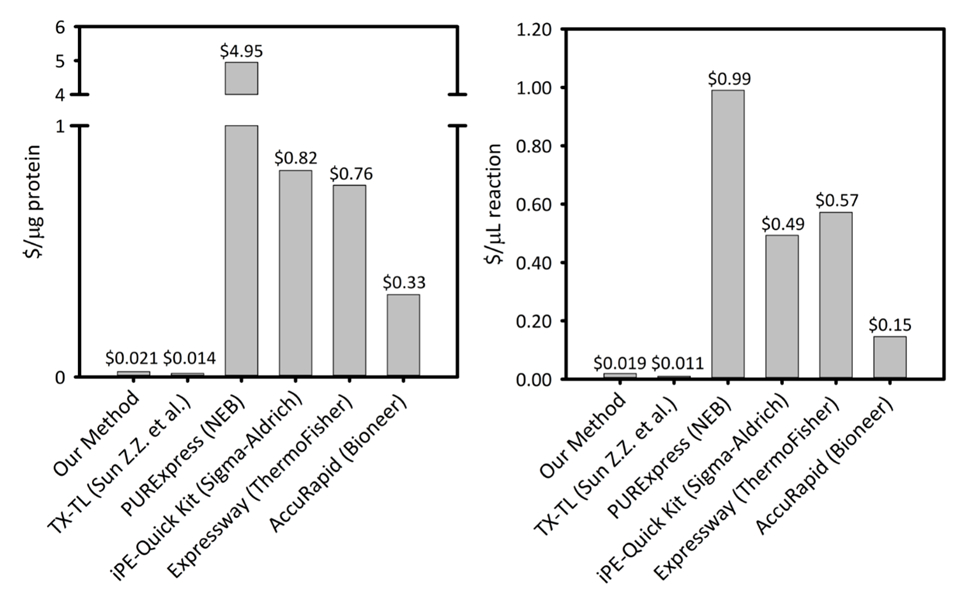
التكميلية رقم 1: تكلفة كل ميكروغرام بروتين التي تنتجها وكل ميكروليتير رد الفعل عبر ست منصات توليف بروتين الخلية الحرة- برنامجنا هو مقارنة بخمس خلايا مختلفة البروتين حرة توليف مجموعات/منصات مع اختلاف الإنتاجية والتسعير. برنامجنا الحراجية sonication على أساس أجدى في كلا $/ميكروغرام من البروتين و $/ميليلتر من رد فعل من معظم التجارية ومجموعات ويوفر سهولة مجموعة لإعداد رد الفعل، بينما التكلفة المتبقية مماثلة لغيرها من منصات الحراجية الأكاديمية. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
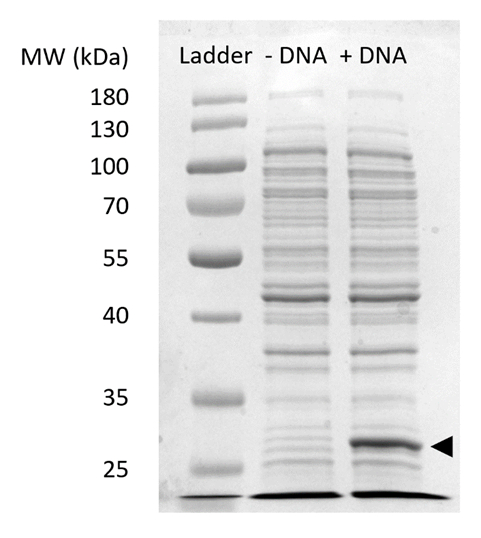
الإضافي رقم 2: الحزب الديمقراطي الصربي صفحة التعبير سفجفب في الحراجية. خلية حرة البروتين التوليف التفاعلات مع (+ الحمض النووي) ودون (-DNA) تم تشغيل قالب الحمض النووي سفجفب على نسبة 12% جل سدسباجي لإظهار التعبير عن سفجفب ولاحظ في كاتشين 27 (السهم الأسود). وقد استخدمت التقنيات التقليدية للحزب الديمقراطي الصربي صفحة. تحميل كل عينة على جل استخراج شملت ميكروغرام 18 من مجموع البروتين استناداً إلى برادفورد التحليل الكمي لمجموع البروتين في الخلية. استناداً إلى قياسات كثافة الأسفار ولدينا المنحنى المعياري، فإننا نقدر أن "+ الحمض النووي" يتضمن لين 0.42 ميكروغرام من سفجفب. من أجل الحصول على هذه العينات، تم تشغيل الحراجية ردود الفعل في نطاق 15 ميليلتر في 1.5 مل [ميكروفوج] أنابيب إنتاج الغلال الحجمي متسقة مع الرقم 3. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
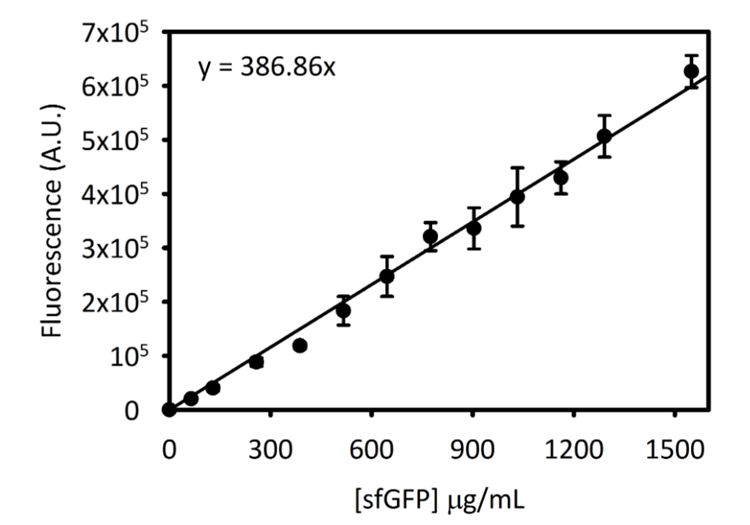
التكميلية الرقم 3: المنحنى المعياري سفجفب على 5 سيتاتيون- وتقرر هذا المنحنى باستخدام الأساليب المذكورة أعلاه. جميع البيانات التي تم جمعها لهذه المخطوطة تم تحويلها من نقطة النهاية قراءات الأسفار إلى [سفجفب] في ميكروغرام/مل تستخدم هذا المنحنى المعياري. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
Subscription Required. Please recommend JoVE to your librarian.
Discussion
تخليق البروتين الخلية الحرة برز كتقنية قوية وملائمة لمجموعة متنوعة من التطبيقات تتراوح التحويلية سريع نموذجي الأنظمة البيوكيميائية. ويدعم القدرة على رصد ومعالجة وزيادة الأجهزة الخلوية في الوقت الحقيقي إلى اتساع نطاق التطبيقات. وعلى الرغم من تأثير هذه التكنولوجيا منصة الآخذة في التوسع، ظلت التكيف الواسع بطيئا بسبب الفروق التقنية في تنفيذ الأساليب. من خلال هذا الجهد، ونحن نهدف إلى توفير البساطة والوضوح لوضع هذه التكنولوجيا في مختبرات جديدة. تحقيقا لهذه الغاية، لدينا بروتوكول كولاي-منصة توليف البروتين استناداً إلى الخلية الحرة يمكن أن يتحقق في وقت بدء تشغيل لأربعة أيام بغير المختبر بتدريب الخبراء، مثل الطلبة الجامعيين (الشكل 1). بالإضافة إلى ذلك، حالما يتم إنتاج مخزون من الكواشف واستخراج، ردود دفعة الحراجية لاحقة يمكن إعداد، المحتضنة، وكمياً في ح 5 فقط. واحد، يمكن أن يؤدي نمو الخلايا 1 لتر في استخراج ما يكفي لردود الفعل الحراجية أربعمائة 15 ميليلتر، أثناء الأعمال التحضيرية دفعة واحدة من الكواشف الأخرى خالية من الخلايا يمكن أن تدعم الآلاف من ردود الفعل. الاستعدادات الكاشف يمكن أيضا الارتقاء إذا كانت هناك حاجة الأوراق المالية أكبر. يمكن أن ردود الفعل الحراجية الإعداد بطريقة الفائق، باستخدام لوحة 96-جيدا أو أنابيب PCR لاختبار مجموعة متنوعة من الظروف في نفس الوقت. سوف انخفاض غلة الحجمي عند استخدام سفن أصغر حجماً كما هو مبين في الشكل 2. ردود الفعل الحراجية يمكن أيضا الارتقاء من ميكروليتيرس إلى عشرات ملليلتر من حجم رد فعل مجموع بغية زيادة إنتاج البروتين الكلي لشرط واحد. عند رفع مستوى وحدة التخزين، أن الاعتبار الرئيسي أن رد فعل الحجمي غلة نقصان كما يقلل نسبة مساحة للتخزين السطحي لرد فعل37،52. بغية الارتقاء مع المحافظة على غلات الحجمي مشابهة للتعبير البروتين، المستخدمين يجب تقسيم حجم رد فعل العديد من أوعية التفاعل و/أو زيادة حجم السفينة. لرد فعل جداول تتراوح بين 15 ميليلتر-100 ميليلتر في وحدة التخزين، ينصح العديد من 15 ردود الفعل ميليلتر في نفس الوقت. لردود فعل تتجاوز 100 ميليلتر في وحدة التخزين، يوصي بلوحات مسطحة القاع 24-جيدا، وينصح بلوحات 12-جيدا لحجم رد فعل يتجاوز 600 ميليلتر. مثل هذه الأزواج من حجم رد الفعل وسفن توفير الاتساق في غلة الحجمي رد فعل على زيادة17،31،،من3752. الارتقاء بمستوى يتجاوز وحدات التخزين هذه يمكن أن يتحقق عن طريق استخدام الآبار متعددة للوحة بالتوازي. باستخدام هذا التنسيق، يمكن تحجيم رد فعل لما يزيد على 10 مل الحجم الإجمالي. تحسين تركيبة سفينة تخزين--رد فعل رد فعل يمكن دعم التطبيقات التحويلية دون التضحية بالإنتاجية من رد فعل.
عند تنفيذ هذا البروتوكول، وهناك بعض الاعتبارات الأساسية التي تؤثر على رد فعل الحجمي الغلال، فضلا عن المؤشرات المرتبطة بسوء أداء استخراج. ضمانا لتحلل السليم ومنع تمسخ آلات النسخ/الترجمة الفنية، من المهم للتخفيف من الحرارة تنتج أثناء تحلل. تزج استثارة الخلية في حمام الماء المثلج أثناء سونيكيشن سرعة تبديد الحرارة خلال سونيكيشن (الشكل 3A). مؤشر لتحلل الخلية فعالة هو ظهور مظهر أكثر قتامة من الخلية ليستي مقارنة بعينات قبل سونيكاتيد (الشكل 3B). للمستخدم مرونة، سونيكاتور والتحقيق هو موضح في الشكل 3 ألف التكيف مع مجموعة من وحدات التخزين من 100 ميليلتر إلى 15 مل خلايا حراكه. ولتحقيق ذلك، يمكن للمستخدم ضبط عدد جولات تسليمها لتحلل الحجم المطلوب للخلايا. بالإضافة إلى ذلك، يمكن إعداد كميات أكبر من استخراج من خلال نهجين متكاملين. المستخدمين يمكن أن sonicate أنابيب متعددة في نفس الوقت، و/أو sonicate كميات أكبر من تعليق خلية، قياس كمية الطاقة متناسب مع الحجم كما هو موضح سابقا29،45. هو خطوة أخرى تشير إلى نوعية استخراج خطوة الطرد المركزي عقب تحلل الخلية. بعد تحلل الخلية، نوصي باستخدام الطرد المركزي في 18,000 س ز تقديم تقسيما واضحا بين المادة طافية (النسخ/الترجمة الآلية، تجزئة الحمض النووي الذي لم يعد يعمل قالب النسخ/الترجمة) و (بيليه غير مرغوب فيها المكونات الخلوية مثل غشاء الخلية والبروتينات سرع) (الشكل 3). وقد وجدنا أن الطرد المركزي في س 18,000 ز يحسن الانفصال، أدى إلى تحسين إمكانية تكرار نتائج المقارنة ليدور بسرعة أقل مثل س 12,000 ز. لمزيد من الراحة، نوصي باستخدام الطرد مبردة أعلى الجدول قادرة على تحقيق الحد أدنى من 12,000 س ز. يتم تنفيذ هذه الخطوة أيضا عادة في 30,000 س ز، التي ينبغي أن تعتبر حالة المعدات المناسبة المتاحة54،55،56،،من5758، 59 , 60-لا يتأثر الأداء استخراج بالطرد المركزي بسرعة في هذه الخطوة نظراً إلى أن يتحقق الفصل السليم. عند إزالة المادة طافية المرجوة، فمن الأفضل لتجنب أي مواد الملبدة بالغيوم التي توجد على الحدود بين المادة طافية وبيليه منذ هذا التلوث سوف يقلل من إنتاجية الاستخراج. تهدف إلى تحقيق نقاء المادة طافية ينتج مقتطفات أكثر إنتاجية ويستحق تخفيض كمية استخراج الحصول عليها للمستخدمين الجدد.
من المهم ملاحظة أنه بينما الأساليب التي قدمنا استنساخه ويمكن تنفيذها بواسطة العلماء مع الحد الأدنى من الخبرة، يمكن أن يكون هناك دفعة لدفعة وتباين رد فعل إلى رد فعل. وقد يعزى هذا إلى الاختلاف في تكوين البروتين ليستي سونيكيشن بعد61. تقلب دفعة إلى الدفعة التي لاحظنا يتناقص عموما عند إضافة الفيتامين a T7RNAP والاستغلال الأمثل لتركيزات المغنيسيوم. إضافة خارجية من T7RNAP شائع بين ردود الفعل الحراجية لدعم التعبير الأمثل من البروتين، ونجد أن وجود مصدرين من مصادر T7RNAP-التعبير الذاتية في BL21*(DE3) و T7RNAP تكميلية لتركيز نهائي من 16 ميكروغرام/ملليلتر-يحسن إمكانية تكرار نتائج دفعة لدفعة للمستخدمين جديدة45،46. مع التجربة، يمكن للمستخدمين تعديل التجارب التي تقوم بها الاستفادة من مصدر واحد فقط من T7RNAP إذا رغبت في ذلك. التحديد الكمي لمحتوى البروتين الكلي دفعة جديدة لاستخراج والتكيف المناسب Mg2 + التركيز قد يساعد أيضا التقليل من التباين دفعة إلى دفعة في غلة التعبير البروتين الحجمي. يمكن أيضا أن تكون الاختلافات في التعبير البروتين نظراً للاختلافات في حجم وهيكل بروتين الفائدة، واستخدام كودون الجينات وموقعها ملزمة الريبوسوم المقابلة من الجينات للفائدة، فضلا عن نوع ناقل التعبير يستخدم62 ،63. ولهذه الأسباب، قد لا يعرب عن بعض البروتينات فضلا عن سفجفب البروتين النموذجي، أدى إلى انخفاض غلة الحجمي ردود الفعل الحراجية.
وتشمل القيود تقنية الحراجية المقدمة أنه قد لا يكون مباشرة مناسبة لكافة التطبيقات للخلية الحرة، مثل الهندسة الأيضية وضبط شروط التعبير، دون إدخال تعديلات إضافية على البروتوكولات. ومع ذلك، نعتقد أن هذا البروتوكول سوف توفر أساسا لوضع منهاج الحراجية في مختبرات جديدة وتزويد غير الخبراء بالقدرة على تنفيذ استهلالي تفاعلات الخلية الحرة في معاملهم. بعد التنفيذ الأولى، تجربة منهاج العمل لجعل التعديلات الخاصة بهم لتطبيقات محددة أكثر استناداً إلى الكتابات الأخرى في الميدان الباحثين.
تكاليف منصة الحراجية دولار 0.021/ميكروغرام البروتين (باستثناء تكاليف العمالة والمعدات)، مما يجعل نظامنا بأسعار تنافسية مع مجموعات تجارية دون المساس بسهولة لإعداد رد فعل. تقييمات للتكاليف النسبية كل ميليلتر من رد الفعل تظهر اتجاهات مماثلة (التكميلية الشكل 1). نحن تقدير تكاليف بدء التشغيل لتكون ~ $4,500 لجميع المواد الكاشفة، و 3,200 دولار إضافية للمعدات المتخصصة، مثل سونيكاتور. ساعات الشخص لإكمال هذا الإجراء يقدر أن ح ~ 26 لكل كاشف الإعدادية من الألف إلى الياء حتى. لكن متى تم إعداد مخزونات كبيرة من المواد الكاشفة، يقلل الطلب على العمالة إلى حد كبير. بالإضافة إلى ذلك، اكتساب الخبرة مع منهاج العمل، ونحن يوصي بالارتقاء بمستوى حجم نمو الخلايا، استخراج الإعداد، وإعداد كاشف لتعظيم كفاءة استخدام الوقت. ونظرا لتكاليف بدء التشغيل، نوصي بمنهاج الحراجية للتطبيقات في البيولوجيا التركيبية، جهود الفائق، وشروط التعبير البروتين التي تتنافى مع منابر التعبير التقليدية البروتين الواجب للصراع مع الخلية القيود الكيمياء الحيوية وقدرتها على البقاء. وفي هذه الحالات المتخصصة حيث تم تمكين تقنية المرجوة من منصة الحراجية، له ما يبرره أكبر تكلفة الحراجية على التعبير في فيفو .
التطوير المستمر للمنهاج الحراجية يرجح أن يوفر فائدة أوسع للجهود الحيوية مثل الهندسة الأيضية للمسارات الانزيمية، إنتاج وتوصيف المستعصية تقليديا والبروتينات، والأحماض الأمينية غير قياسي التأسيس والتعبير بروتين غير طبيعي وصناعة الأدوية طبقية والتوسع خارج المختبر في الفصول الدراسية لوقف التعليم64،،من6566. وستدعم هذه الجهود كذلك بالجهود الجارية من أجل توصيف مفصل لمنهاج الحراجية. فهم أفضل للتكوين لاستخراج الخلية يؤدي إلى التحسين المستمر تجاه رد فعل تحسين غلة المحاصيل والمرونة في رد فعل الظروف61،،من6768.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
الكتاب يعلن أن لديهم لا تضارب المصالح المالية أو غيرها تضارب في المصالح.
Acknowledgments
المؤلف تود أن تقر الدكتورة جنيفر فانديركيلين، أندريا لوبسشير، وتوني تارتو للدعم التقني ويسلي كاو، لين وليامز وهايت كريستوفر لإجراء مناقشات مفيدة. المؤلف يعترفون أيضا بدعم تمويلي من بيل وليندا الصقيع الصندوق، مركز التطبيقات في شركة شيفرون التكنولوجيا الحيوية تطبيق البحوث الهبات المنح للتكنولوجيا الحيوية والبحوث بولي كال، علمية وبرنامج منح الأنشطة الإبداعية (رسكا 2017)، ومؤسسة العلوم الوطنية (NSF-1708919). [مزل] تسلم "المنحة خريج جامعة كاليفورنيا الدولة". MCJ يعترف الجيش بحوث مكتب W911NF-16-1-0372، تمنح "المؤسسة الوطنية للعلوم" 1413563 المجلس والمجلس-1716766، والقوة الجوية البحوث مختبر مركز التميز منحة FA8650-15-2-5518، "منحة وكالة الحد من تهديد الدفاع" HDTRA1-15-10052/P00001، وديفيد ولوسيل باكارد، البرنامج المعلم-الباحث كاميل دريفوس، الإدارة للطاقة البر المنح دي-SC0018249، برنامج العلوم البشرية الحدود (RGP0015/2017)، منحة أتوب معهد الجينوم المشترك الفلاني، و شيكاغو الكونسورتيوم الطبية بدعم من "أموال سيرل" في "ثقة المجتمع شيكاغو" للدعم.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}