

Summary
该协议详细介绍了在4天或更短的时间内生成大肠杆菌基细胞提取物并实施体外蛋白质合成反应所需的步骤、成本和设备。为了将这一平台的灵活性用于广泛的应用, 我们讨论了可以调整和优化的反应条件。
Abstract
在过去的50年里, 无细胞蛋白合成 (cfps) 已成为一种强大的技术, 可以利用试管内细胞的转录和转化能力。通过避免维持细胞活力的需要, 并通过消除细胞屏障, cfps 已成为新兴应用的基础, 在生物制造传统上具有挑战性的蛋白质, 以及应用于快速原型设计代谢工程和功能基因组学。我们实施基于大肠杆菌的 cfps 平台的方法允许新用户访问其中的许多应用程序。在这里, 我们描述了通过使用浓缩介质、挡板烧瓶和可调谐超声基础细胞裂解的可重复方法制备提取物的方法。该提取物可用于蛋白质表达, 能够在从实验设置到数据分析的短短5小时内产生900μg/ml 或更多的超级文件夹绿色荧光蛋白 (sfgfp), 因为事先已经准备好了适当的试剂储备。获得试剂的启动成本估计为 4, 500 美元, 这将维持数千种反应, 估计每产生的蛋白质成本为0.01 美元, 每微米反应0.1919 美元。此外, 蛋白质表达方法反映了由于优化试剂预混料而在市售系统中看到的反应设置的易用性, 成本只是很小的一小部分。为了使用户能够利用 cfps 平台的灵活性, 实现广泛的应用, 我们确定了该平台的各个方面, 这些方面可以根据可用资源和蛋白质表达结果进行调整和优化期望。
Introduction
无细胞蛋白合成 (cfps) 已成为一项技术, 在过去50年中, 在1、2的时间里, 为蛋白质生产、功能基因组学、代谢工程等带来了许多新的机遇。与标准的体内蛋白质表达平台相比, cfps 提供了三个关键优势: 1) 该平台的无细胞性质使其能够产生对细胞有潜在毒性或与细胞无关的蛋白质3,4 ,5,6;2) 基因组 dna 失活, 并引入编码感兴趣基因的模板 dna, 将所有的系统能量引导到对感兴趣的蛋白质产生的反应中;3) 平台的开放性使用户能够实时修改和监测反应条件和成分7、8.这种直接进入反应的方式支持了生物系统的增强, 增加了化学和氧化还原条件, 用于生产新型蛋白质和调整代谢过程2,9,10. 直接访问还允许用户将 cfps 反应与活动检测结合在单锅系统中, 以便更快速地进行设计-构建-测试周期11。在小批量液滴或纸质设备上执行 cfps 反应的能力进一步支持了高通量发现工作和 12、13、14、15的快速原型设计 ,16。由于这些优势和系统的即插即用性质, cfps 独特地促进了各种生物技术应用, 例如在体内难以溶解表达的蛋白质的生产17 18,19,20, 疾病检测 21,22,23, 按需生物制造18,24 25,26, 27,和教育28,29, 所有这些都显示了无细胞平台的灵活性和实用性。
cfps 系统可以从原核细胞和真核细胞系的各种粗裂解物中生成。这使得选择系统中有不同的选择, 每个选择都有优点和缺点, 这取决于利益的应用。cfps 系统在制备时间、成本和生产率方面也有很大差异。最常用的细胞提取物来自小麦胚芽、兔网状细胞、昆虫细胞和大肠杆菌细胞, 后者是迄今为止最具成本效益的, 同时产生的蛋白质体积产量最高。.虽然其他 cfps 系统可以有利于其固有的翻译后修饰机械, 新兴的应用使用大肠杆菌为基础的机械能够通过产生现场-专门磷酸化和糖基化蛋白按需31,32,33,34,35。
cfps 反应可以以批处理、连续交换无细胞 (cecf) 或连续无流动细胞 (cfcf) 格式运行。批次格式是一个封闭的系统, 其反应寿命是有限的, 由于减少的反应物的数量和反应的抑制副产物的积累。cecf 和 cfcf 方法提高了反应的使用寿命, 从而提高了与批处理反应相比的体积蛋白产量。这是通过允许从反应容器中去除蛋白质合成的副产品来实现的, 而在整个反应过程中, 新的反应物都是在2的过程中提供的。在 cfcf 的情况下, 感兴趣的蛋白质也可以从反应室中去除, 而在 cecf 中, 感兴趣的蛋白质仍然存在于由半透膜36,37组成的反应室中。这些方法对于克服感兴趣的难以表达的蛋白质的体积产量低特别有价值,38、39、40、41、42、 43岁实施 cecf 和 cfcf 方法的挑战是: 1) 虽然它们能更有效地利用负责转录和翻译的生物机械, 但它们需要更多的试剂, 从而增加总体成本, 并且增加 2)与批处理格式44相比, 它们需要更复杂的反应设置和专用设备。为了确保新用户的可访问性, 本文所述的协议侧重于15微米反应量的批处理格式, 并就将反应体积增加到毫升规模提出具体建议。
本文介绍的方法使具有基本实验室技能的非专家 (如本科生) 能够实现大肠杆菌 cfps 系统的细胞生长、提取制备和批量格式反应设置。与商用套件相比, 这种方法具有成本效益, 而不会牺牲基于风筝的反应设置的易用性。此外, 这种方法还可在实验室和现场应用。在决定实施 cfps 时, 新用户应彻底评估传统蛋白表达系统在启动投资方面的有效性, 因为 cfps 可能并非在所有情况下都优于 cfps。这里描述的 cfps 方法使用户能够直接实现各种应用, 包括功能基因组学、高通量测试、可在体内表达的难解蛋白质的生产以及现场应用包括合成生物学的生物传感器和教育套件。其他应用, 如代谢工程, 调整蛋白表达条件, 疾病检测, 并使用 cecf 或 cfc 方法扩大规模仍然是可能的, 但可能需要经验与 cfps 平台进一步修改反应条件。我们的方法结合了丰富的介质和挡板烧瓶的生长, 通过超声相对快速和可重复的细胞裂解方法, 然后是一个简化的 cfps 反应设置, 利用优化的预混料45。虽然细胞生长方法在这一领域已经有些标准化, 但细胞裂解的方法差别很大。除了超声, 常见的裂解方法包括利用法国印刷机, 均质机, 珠虫, 或溶菌酶和其他生化和物理破坏方法46,47, 48,49. 使用我们的方法, 每1升细胞获得大约2毫升的粗细胞提取物。该细胞提取物可支持 4, 100μl cfps 反应, 每个反应从模板质粒 pjl1-sfgfgfp 中产生 ~ 900μgg\ cfml 蛋白。这种方法的成本为0.021 美元/mfgfp (反应为 0.19μl $), 不包括人工和设备成本 (补充图 1)。从零开始, 此方法可以在4天内由一个人实现, 重复 cfps 反应可以在数小时内完成 (图 1)。此外, 该协议可以批量扩展, 以进行更大批次的试剂制备, 以满足用户的需要。重要的是, 这里介绍的协议可以由实验室培训的非专家 (如本科生) 来实施, 因为它只需要基本的实验室技能。下面描述的过程和随附的视频是专门开发的, 以改善大肠杆菌cfps 平台的可访问性, 供广泛使用。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 培养基制备和细胞生长
-
第1天
- 将大肠杆菌BL21*(DE3) 细胞从甘油库存流到 lb 琼脂板上, 在37°c 下孵育至少18小时。
- 在121°c 的液体循环中准备50毫升 lb 介质和高压灭菌器溶液, 时间为30分钟。在室温下存放。
-
第2天
- 如补充信息所述, 准备750毫升的 2x ytp 介质和250毫升 0.4 m d-葡萄糖溶液。
- 将 2x ytp 介质倒入高压灭菌的 2.5 l 挡板瓶中, d-葡萄糖溶液倒进蒸压500毫升玻璃瓶中。在121°c 的液体循环中, 两个溶液在液体循环中进行30分钟的高压灭菌。
- 如果第二天进行细胞生长, 请确保两种无菌溶液都储存在 37°c, 以便在接种时最大限度地提高生长速度。接种前不要混合溶液。
注: 如果需要, 解决方案可以在4°c 下存储 1-d, 但 2x ytp 介质极易受到污染。 - 通过使用灭菌环路和无菌技术, 为50毫升的 lb 介质接种单池 BL21(DE3), 开始一夜培养 BL21(DE3), 以避免污染。
- 将 BL21*(DE3) 的50毫升) lb 培养物放入 37°c 250 转/分摇床培养器中, 长至15-18小时。
- 准备和消毒第3天和第4天所需的所有材料, 包括: 两个1升离心机瓶, 4x 冷50毫升锥形管 (重量和记录质量为 3个), 和许多 1.5 ml 微泡管。
-
第3天
- 从晃动孵化器中取出 BL21*(DE3 中的50毫升隔夜培养物, 并使用1:10 稀释 lb 介质在分光光度计上测量 od 600。计算为1升增加1升的介质所需的动态培养量, 用于启动 od 600 的 0.1 (例如, 如果 1:10 稀释的 od600被理解为 0.4, 则将未稀释的 od 600 的25毫升接种到2xytp 的1升中 g)。
- 从37°c 孵化器中取出加热的 2x ytp 介质和 d-葡萄糖溶液以及 lb 培养的50毫升。使用无菌技术, 小心地将 d-葡萄糖溶液倒入 2x ytp 介质中 (避免令人困惑的烧瓶两侧)。
注: d-葡萄糖的添加完成了 2倍 ytpg 的1升配方。 - 保持无菌技术, 用适当数量的50毫升培养接种 2x ytpg 溶液的1升, 以 0.1 od 600 的形式开始1升培养.立即将接种的1升培养培养在200转/分的37°c 晃动孵化器中。
- 在生长的第一个小时后进行第一个 od600读数 (滞后阶段典型需要 1小时)。不要冲淡文化。继续进行 od600测量大约每20-30 一次, 直到 od600达到0.6。
- 达到 od600 = 0.6 后, 在 2x ytpg 培养中加入 1M iptg (最终浓度为1升培养 = 1 mm)。
注: 理想感应 od600为 0.6;然而, 0.6-0.8 的范围是可以接受的。iptg 的诱导是用于 t7 rna 聚合酶 (t7rnap) 的内源性生产。 - 诱导后, 大约每20-30 测量一次 od600 , 直到达到3.0。
注: 在此期间将离心机冷却至4°c。准备冷 s30 缓冲区, 详见补充资料。如果 s30 缓冲区是预先准备好的, 请确保直到使用当天才添加 dtt。 - 一旦 od600达到 3.0 (图 2a), 将培养物倒入冰水浴池中的冷1升离心机瓶中。准备一个重量相等的充水1升离心机瓶, 作为离心机中的平衡。
注: 吸波值因仪器到仪器而异。虽然 BL21(DE3) 收获的 od600不是一个敏感变量, 但建议用户评估和优化此变量, 作为故障排除措施。与较小的基于立方的分光光度计相比, 较大的分光光度计可能会导致更低的 od600读数。 - 在 5, 000 x g 和10°c 的温度下将1升瓶离心 10分钟, 使其产生颗粒细胞。
- 慢慢地把上清剂倒出来, 按照机构的生物废物程序进行处理。把颗粒放在冰上。
- 使用无菌铲子, 刮掉离心机瓶中的细胞颗粒, 并将其转移到一个冷的50毫升锥形管。
- 将30毫升的冷 s30 缓冲液加入锥形管, 并通过短爆裂 (20-30秒) 和在冰上的休息时间 (1分钟) 循环重新悬浮细胞颗粒, 直到完全重新悬浮, 没有块。
- 一旦颗粒完全重新悬浮, 使用另一个50毫升锥形管与水作为平衡和离心10分钟在 5000 x g 和 10°c (预冷到 4°c)。
注: 这就完成了收获细胞时所需的3次洗涤的1次。 - 将上清液倒出, 并根据机构的生物废物程序进行处理。用 20-25 ml 的冷 s30 缓冲液和离心机在 5000 x 克和 10°c (预冷至 4°c) 下重新使用颗粒10分钟。
注: 这完成了3次洗涤中的第2个. - 再次, 倒出上清液, 并根据机构的生物废物程序进行处置。再次加入30毫升的 s30 缓冲液和涡旋, 重新悬浮颗粒。
- 使用3个预称、冷的50毫升锥形管和带有无菌移液器的血清学移液器填料, 将重新悬浮的颗粒颗粒的 aliquot 10 ml/s30 缓冲液混合物放入3个锥形管中的每一个。
注: 不需要将电池拆分为3个管, 但此步骤会产生较小的电池颗粒 (~ 1 g), 以便在以后的步骤中增加便利性。 - 在 5000 x g 和 10°c (预冷至 4°c) 下, 使用所需的适当平衡, 离心所有管10分钟。
注: 这将完成最后的清洗步骤。 - 将上清液倒出, 并根据机构的生物废物程序进行处理。用干净的纸巾仔细擦拭锥形管和帽的内部, 取出多余的 s30 缓冲液;避免接触颗粒。
- 在分析天平上重新称重管, 并记录每个管的最终颗粒重量。
注: 此时可以暂停协议。颗粒可以在液氮中闪烁冷冻, 并在-80°c 下储存长达一年, 直到提取制备所需。
2. 粗细胞提取物制备-第4天
- 为了提取准备, 在每一步都要让细胞在冰上保持低温。每1克细胞质量加入1毫升冷 s30 缓冲液。确保二硫醇 (dtt) 已补充到 s30 缓冲液, 最终浓度为 2 mm。
注: 在此期间将微型离心机冷却至4°c。 - 通过短时间 (20-30秒) 和在冰上休息 (1分钟) 的循环, 重新悬浮细胞颗粒, 直到完全重新悬浮。如果重新悬浮是困难的, 让颗粒在冰上 3 0分钟解冻。
- 将1.4 毫升的再悬浮细胞转移到 1.5 ml 微泡管中。
- 将一个含有 1.4 ml 重新悬浮细胞的 1.5 ml 管放入烧杯中的冰水浴中。在45秒上使用, 在3个总周期中, 以59秒为秒, 振幅设置为50%。关闭并反转管, 在关闭期间轻轻混合。总共向每一个含有 1.4 ml 重悬浮细胞的 1.5 ml 微泡管提供 800-900 j 的能量 (图 3 a & 3A)。
注: 此步骤对使用的声纳器类型和型号非常敏感, 如果设备与此过程中列出的设备不同, 则应对此步骤进行优化。两种互补的方法可以用来扩大在这一步骤中制备的提取物的数量: 1) 多个 1.5 ml 微泡管可以平行超声对,/或 2) 较大的体积可以在锥形管中超声 (每个管的细胞再悬浮液最多15毫升), 以扩展前面所述的 29 、45 所述的交付能量。 - 在超声完成后, 立即在 1.4 ml 的裂解液中加入4.5μl 的 1 m dtt (补充另外的 2 mm dtt), 并多次反转混合。把管子放在冰上。对任何额外的重新悬浮细胞管重复步骤2.4 和 2.5, 然后再进行离心。
- 在 18, 000 x g 和4°c 的微离心机样品中, 时间为 10分钟 (图 3c)。
- 将上清液移注到一个新的 1.5 ml 微泡管中。不要打扰颗粒;最好是留下一些上清液来保持纯度, 而不是破坏颗粒, 以最大限度地提高产量。
- 将上一步的上清液以250转/分和37°c 孵育 60分钟, 将管道贴在孵化器的晃动平台上 (这是径流反应)。
- 在 10, 000 x g 和4°c 的微离心机样品中, 10分钟。
- 在不干扰颗粒的情况下取出上清液, 并将其转移到新的管中。创建许多100μl 的提取物, 用于存储。
注: 该协议可以在这里暂停, 提取物可以在液氮中闪烁冻结, 并在-80°c 下储存长达一年, 直到 cfps 反应所需。至少可以进行5个冻融循环, 而不会影响提取生产率 (图 4)。
3. 无细胞蛋白合成批式反应
- 解冻溶液 a 和 b, dna 模板, BL21(DE3) 提取物 (如果冷冻), t7rnap, 和分子级水的一个脂肪。
注: cfps 反应模板可在补充信息中找到。补充资料中提供了解决方案 a 和 b 配方, 与众多试剂的特定浓度相对应, 以支持 cfps 基于 panox-sp 的能源系统。每种试剂的作用和这些能够支持 cfps 的试剂浓度的可接受变化已被确定为 50。t7rnap 净化协议见补充资料 51。补充 t7rnap 可以增加体积产量, 但如果在细胞生长过程中诱导 t7rnap, 则没有必要。质粒 dna 模板 (pjl1-sfgffp) 可以使用 maxiacpreckit (pjl1-sfgffp) 进行制备, 该试剂盒中的洗涤缓冲液可进行两次洗涤, 然后使用 pcr 纯化试剂盒进行后处理 dna 清理 (图 2b)。线性 dna 模板也可用于 cfps 反应。 - 标记 cfps 反应所需的必要的微泡管量。
注: 反应可以在不同的容器大小, 但较小的容器可以减少体积蛋白质产量 (图 2c)。由于表面积与体积比的降低, 在同一尺寸的容器中扩大反应也可能会减少体积产率, 这是减少氧交换的函数。当反应体积增加到100μl 以上时, 建议使用平底井板31、37、52. - 加入2.2 微米的溶液 a、2.1 微米的溶液 b、5μl 的 BL21*(DE3) 提取物、0.24 微克的 t7rnap (16μgml 最终浓度)、0.24 ng 的 dna 模板 (16ng/ml 最终浓度) 和水, 使最终体积达到15μl。
注: 涡流溶液 a 和 b 在反应设置过程中经常发生, 以避免成分的沉淀, 并确保每个反应都得到每个溶液的均匀电荷。避免对提取物进行涡流处理, 而是反转管混合。 - 在反应中添加了所有试剂后, 通过上下移液或轻轻涡流混合每个管, 同时确保最终反应混合物在 1.5 ml 微泡管底部的单个15微米珠中混合。
- 将每个反应放入37°c 孵化器, 不晃动 4小时, 或在夜间30°c。
注: 成功的反应可以根据 cfps 反应混合物中 sfgfp 产品的绿色进行定性评估 (图 3d)。感兴趣的蛋白质的表达也可以通过 sds-page 得到证实 (补充图 2)。
4. 记者蛋白的定量, [sfgfp]
- 将 0.05 m hepes 的48μl 加载到定量所需的每口井中 (通常每根反应管以三分位的方式进行)。
- 去除孵化器中的反应。上下移液器混合每个反应, 然后将2μl 的反应转移到 48μl 0.05 m hepes, ph 值8。在井里再上下移水混合。
- 一旦所有反应被加载和混合, 将96井板放入荧光计, 并测量 sfgfp 端点荧光。
注: sfgfp 荧光定量的激发和发射波长分别为485纳米和510纳米。 - 使用先前生成的标准曲线, 从获得的荧光读数中确定 [sfgfp]。
注: 补充资料 (补充图 3) 提供了生成 sfgfp 浓度与荧光强度的标准曲线的说明。用户将需要为其仪器建立一个标准曲线, 因为仪器的灵敏度可能会有所不同。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
我们提出了一个基于声纳的大肠杆菌提取物制备协议, 可以在四天的时间内完成,图 1显示了每天的程序故障。有可利用的步骤, 可以在每天完成与各种暂停点, 但我们发现这个工作流是最有效的执行。此外, 细胞颗粒 (步骤 1.3.18) 和完全准备好的提取物 (步骤 2.10) 在-80°c 下稳定至少一年, 允许用户创建每个的较大库存,以便在以后使用17。提取物不仅在很长一段时间内稳定, 而且还可以进行至少五次冻融循环, 而不会显著降低生产率 (图 4)。这使得在冰柜存储空间有限的情况下, 可以将较大的提取物的等价物存储用于多种用途。但是, 我们建议尽可能使用多个较小的提取物 (~ 100μl)。
对于每一种新的萃取制剂, 我们建议用户进行镁滴定, 以确定该批次提取物的最佳镁含量。用户可以通过布拉德福德法量化细胞提取物总蛋白质浓度的批次间变化。对于性能较高的提取物, 我们通常会看到总蛋白浓度为 30-50 mgml, 在此范围内, 总蛋白质浓度与细胞提取物性能之间没有明显的相关性。因此, 我们建议用户相应调整镁浓度, 以确保每个提取批次的蛋白质和核酸功能最大化。镁水平对于适当的 dna 复制、转录和翻译非常重要, 但过量的水平可能对这些过程有害53。为了证明这种依赖性, 我们对镁和提取量进行了共滴定, 以确定最佳组合, 最大限度地减少必要的萃取量, 同时保持有效反应 (图 5)。从这个实验中, 我们建议使用5μl 的提取物和 10mm mg2 +的提取物, 总蛋白质含量为 30 mg/ml, 以获得超过 1, 000μgml 的 sfgfp。
我们在 cfps 方面的经验也使我们能够确定协议中可以在不损害系统整体生产力的情况下进行更改的步骤, 以及其他对高性能 cfps 系统不可或缺的步骤。最值得注意的是, 细胞收获的最后 od600并不显著影响 cfps 反应的最终产出, 细胞可以从 2.7-4.0 od600的任何地方收获。这代表了生长的早期指数阶段, 每个细胞核糖体浓度最高, 平移机械是支持快速生长的最活跃的。此观察允许用户灵活地优化自己的过程。我们建议在大约 3.0 od600 处采集, 以便在收获完成时在接近3.3 的 od600处捕获细胞 (图 2a)。影响 cfps 产量的变量包括模板 dna 质量、反应容器大小以及反应中存在的细胞提取物和镁离子的相对数量。我们发现 dna 质量有显著的批次间变化。为了解决这个问题, 我们建议用户通过 midi 或 maxxi 准备技术纯化 dna, 然后在 max-p门前使用的 dna 纯化柱上执行额外的 dna 清理步骤, 或者使用额外的 dna 清理工具包进行纯化后。这提高了 cfps 反应的 dna 质量的重现性, 并使蛋白质生产更加稳健 (图 2b)。反应容器还会影响体积产量, 因此, 在不同容器体积的相同反应装置中, 相同反应装置的蛋白质产量可能相差40%。据推测, 在较大的容器中观察到的体积产率的增加是由于反应混合物的表面积增加, 从而实现了更好的氧交换 (图 2c), 而其他材料则通过运行进一步提高了体积产率cfps 反应在大型平底板, 我们建议超过 100μl17,31,37,52的反应。

图 1: 培养生长、细胞提取物的产生、cfps 反应的设置和定量的时间表.用户可以通过这四天的工作流程为他们的研究应用程序实现 cfps 平台。试剂制备是本实验第一轮的主要时间和成本投资, 在试剂库存建立后大幅减少。此外, 细胞颗粒和准备好的细胞提取物可以存储在-80°c 超过一年, 允许用户在不同的步骤开始时间线, 以更快的结果。用户还可以在不同的步骤中暂停修改此工作流的时间线。请点击这里查看此图的较大版本.

图 2: cfps 的可修改条件及其对体积反应产率的影响.a.在各种 od600 读数中, 根据收获 BL21(DE3) 细胞提取生产力比较。基于此图, 我们建议在 od600的3.3 下进行收获, 以产生至少1000μgl 的目标蛋白。反应在 1.5 ml 微泡管中以15μl 的比例进行。两种dna 最大清洗方案的比较, 包括和没有纯化后的 dna 清理。pjl1-sfgffp 质粒经过一次或两次清洗后, 通过 pcr 纯化试剂盒进行了净化后的工作。为了达到 ~ 900μgl 的蛋白质表达, 我们建议进行纯化后 dna 清理, 而不考虑最大洗液的数量。反应在 1.5 ml 微泡管中以15μl 的比例进行。c. 15μl cfps 反应在各种容器中进行, 范围从2毫升到0.6 毫升微泡管不等。"neg" 代表一种负面的控制, 在这种控制中没有在反应中添加 dna 模板。所有误差条代表每个条件的三个独立反应的1个标准偏差, 每个误差点都被量化为一式三份。请点击这里查看此图的较大版本.

图 3: 创建高效提取物的关键过程设置和结果。a.正确设置超声冰水浴, 以确保样品冷却, 而在超声过程中产生热量。b. 1.5 ml 微泡管, 含有重新悬浮细胞颗粒前 (左) 和后 (右) 超声。与重新悬浮的细胞颗粒相比, 产生的裂解液应显示出更深的色调。c.在 18, 000 x 克离心后, 适当分离细胞裂解液的上清液和颗粒。d.在37°c 孵育4小时后的 cfps 反应。右侧 1.5 ml 微泡管 (成功反应) 显示 sfgfp 报告蛋白在 ~ 900μg/ml 处可见荧光。左侧的负控制管, 缺乏模板 dna, 模拟不成功的反应, 显示一个清晰的解决方案, 没有荧光。请点击这里查看此图的较大版本.

图 4: cfps 提取物在5个冻融周期内蛋白质表达的变化.从同一生长中提取的物经过五个冷冻解冻周期, 通过液氮闪光冻结, 然后在冰上解冻。在五个冻融周期中, 表达 sfgfp 的提取物生产率没有显著变化。反应在 1.5 ml 微泡管中以15μl 的比例进行。"neg" 代表一种负面的控制, 在这种控制中没有在反应中添加 dna 模板。所有误差条代表每个条件的三个独立反应的1个标准偏差, 每个误差点都被量化为一式三份。请点击这里查看此图的较大版本.

图 5: cfps 用于不同 [mg2+] 的反应和提取体积与 [sfgfp].[mg2 +] 从 8 mm 到 14 mm 不等, 增量为 2 mm, 提取量从3μl 到 7μl, 增量为1μl。颜色代码表示 [sfgfp] 从高 (红色) 到低 (紫色) 生成。为了在保持高蛋白生产的同时最大限度地提高试剂效率, 我们建议对总蛋白质含量约为 30 mg/ml 的提取物使用5μl 和 10mm mg 2 +, 由 bradford 法测定. 生成等高线图的原始点基于每个条件的三个独立反应的端点荧光, 每个反应都是一式三份。反应在 1.5 ml 微泡管中以15μl 的比例进行。请点击这里查看此图的较大版本.
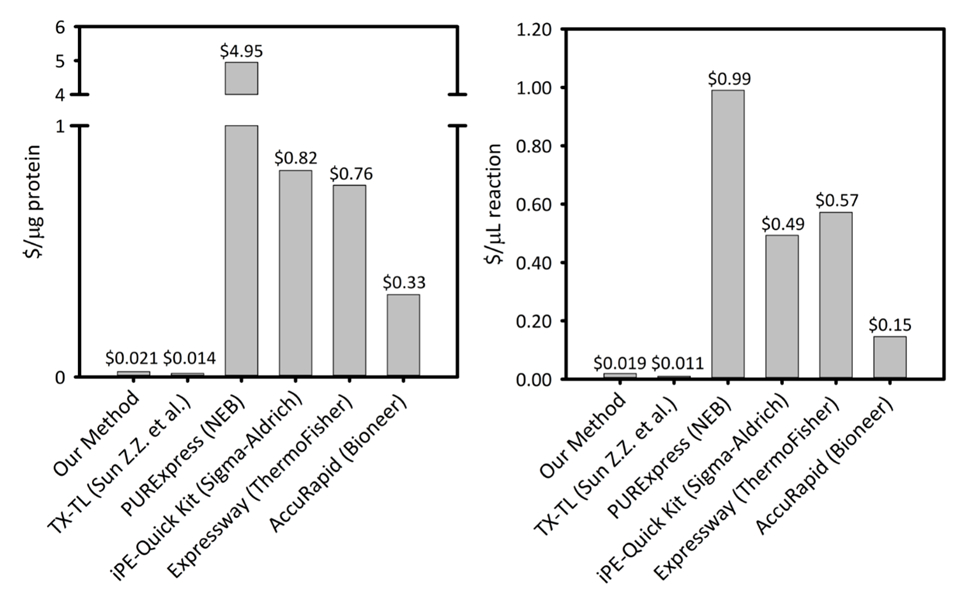
补充图 1: 六个无细胞蛋白质合成平台每微克产生的蛋白质和每微升的反应成本.我们的平台与五种不同的无细胞蛋白合成小猫平台进行了比较, 这些平台具有不同的生产率和定价。我们基于声纳的 cfps 平台在蛋白质和反应中都比大多数商业试剂盒更具成本效益, 并为反应设置提供了更方便的试剂盒, 同时剩余的成本与其他学术 cfps 平台相当。请点击这里查看此图的较大版本.
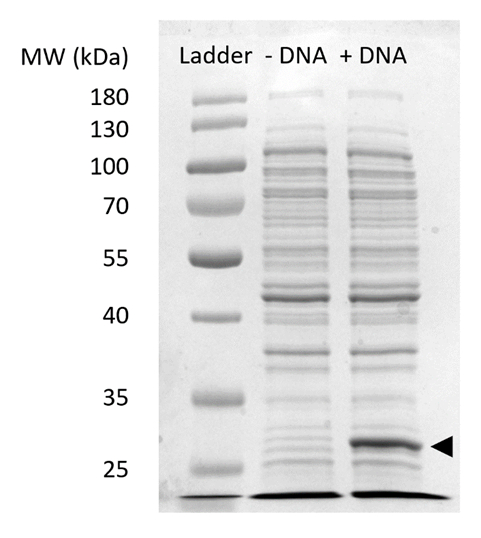
补充图 2: cfps 中 sfgfp 表达式的 sds-page.用 (+ dna) 和不 (-dna) dna 模板在 12% sfGFP 凝胶上进行无细胞蛋白合成反应, 以证明在 27 kda (黑色箭头) 观察到的 sfgfp 表达。采用了传统的 sds-page 技术。每个样品加载到凝胶包括18微克的总蛋白基于布拉德福德分析定量的总蛋白在细胞提取物。根据荧光强度测量和我们的标准曲线, 我们估计 "+ dna" 车道包含0.42 微克的 sfgfp。为了获得这些样品, cfps 反应在 1.5 ml 微泡管中以15μl 的比例运行, 产生的体积产率与图 3c一致。请点击这里查看此图的较大版本.
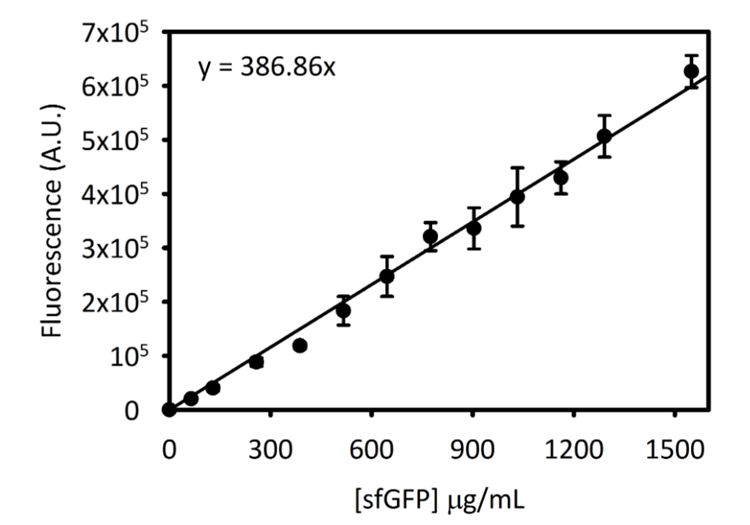
补充图3:cycycycypyp 5 上的 sfgfp 标准曲线.这条曲线是用上面概述的方法确定的。为这份手稿收集的所有数据都使用此标准曲线从端点荧光读数转换为μg/ml 中的 [sfgfp]。请点击这里查看此图的较大版本.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
无细胞蛋白合成已成为一种强大的和有利的技术, 可用于从生物制造到生化系统的快速原型设计等各种应用。应用程序的广度由实时监视、操作和扩充蜂窝机械的能力提供支持。尽管这一平台技术的影响越来越大, 但由于在实施这些方法方面存在技术细微差别, 广泛的适应仍然缓慢。通过这一努力, 我们的目标是为在新实验室中建立这一技术提供简单和清晰。为此, 我们的基于大肠杆菌的无细胞蛋白合成平台的协议可以在四天内由受过实验室培训的非专家 (如本科生) 实现 (图 1)。此外, 一旦产生了试剂和提取物的库存, 随后的 cfps 批处理反应可以在短短5小时内建立、孵化和量化。单次 1 l 细胞生长可产生足够的提取物, 用于 4, 1015μl cfps 反应, 而其他无细胞试剂的单批制剂可支持数千种反应。如果需要更大的库存, 试剂制剂也可以扩大规模。cfps 反应可以通过使用96孔板或 pcr 管同时测试各种条件, 以高通量的方式进行设置。使用较小的容器时, 体积产率将下降, 如图2c 所示。cfps 反应也可以从微升扩大到每千毫升的总反应量, 以增加单一条件下的总蛋白质产量。当缩放体积时, 主要考虑的是体积反应产生下降, 因为反应的表面积与体积的比率降低37,52。为了在保持蛋白质表达量相似的同时扩大规模, 用户应将反应体积分成多个反应容器, 或增加血管大小。对于体积从15μl-100μl 不等的反应尺度, 建议同时进行大量的15μl 反应。对于体积超过100μl 的反应, 建议采用平底24孔板, 对于超过600μl 的反应体积, 建议使用12孔板。这种反应体积和容器的配对在放大17、31、37、52时提供了体积反应产率的一致性。通过并行利用板的多个井, 可以实现超出这些体积的扩展。使用这种格式, 反应可以扩大到超过10毫升的总体积。优化反应体积反应容器组合可以在不影响反应生产率的情况下支持生物制造的应用。
执行此协议时, 有几个关键的注意事项会影响体积反应产生率, 以及与性能不佳的提取相关的指标。为了保证正确的裂解和防止功能转录/翻译机械的变性, 重要的是要减轻裂解过程中产生的热量。在超声过程中将细胞重新悬浮在冰水浴中, 以便在超声过程中迅速散热 (图 3 a)。有效细胞裂解的一个指标是出现了一个更黑暗的外观细胞裂解物相比, 前超声样品 (图 3b)。为了用户的灵活性,图 3 a 所示的声纳和探头适用于从100μl 到 15 ml 的重新悬浮细胞的体积范围。为此, 用户可以调整为对所需的细胞体积进行裂解而交付的焦耳数量。此外, 可以通过两种互补的方法制备更多的提取物。用户可以并行对多个管进行充电, 并且/或将较大体积的细胞再悬浮物压缩, 从而按前面所述的体积按比例缩放能量量。另一个表明提取质量的步骤是细胞裂解后的离心步骤。在细胞裂解后, 我们建议在 18, 000 x 克离心, 以便在上清液 (转录/翻译机制、不再起作用于模板转录/翻译的基因组 dna) 和颗粒 (转录/翻译) 之间进行明确的划分。不需要的细胞成分, 如细胞膜和沉淀蛋白) (图 3c)。我们发现, 18, 000 x 克离心可改善分离, 与 12, 000 x g 等低速旋转相比, 可更好地提高重现性。为方便起见, 我们建议使用桌面冷冻离心机, 能够达到最小的 12, 000 x 克。这一步骤通常也以 30, 000 x g 执行, 如果有适当的设备, 则应考虑到这一步骤:54、55、56、57、58、59,60. 考虑到实现了适当的分离, 在这一步骤中, 提取性能不受离心速度的影响。当去除所需的上清液时, 最好避免在上清液和颗粒之间的边界上存在任何多云的物质, 因为这种污染会降低提取物的生产率。为了提高上清液的纯度, 可以获得更高效的提取物, 并为新用户减少提取物的数量是值得的。
需要注意的是, 虽然我们提出的方法是可复制的, 并且可以由专业知识最少的科学家执行, 但可能会有批次之间的批次和反应反应的变化。这可能是由于裂解物的蛋白质组组成的变化, 661.我们观察到的批次间的可变性通常在补充 t7rnap 和优化镁浓度时降低。外源添加 t7rnap 在 cfps 反应中很常见, 以支持最佳的蛋白质表达, 我们发现, 有两个来源的 t7rnap-内生表达在 BL21*(DE3) 和补充 t7rnap 到最后浓度为 16μgml-改善新用户的批量重现性为45,46。根据经验, 用户可以修改其实验, 以便在需要时只使用 t7rnap 的单个源。对一批新的提取物的总蛋白质含量进行定量, 并适当调整 mg2+浓度, 也有助于减少体积蛋白质表达产量的批次间变化。蛋白质表达的变化也可能是由于感兴趣的蛋白质的大小和结构的差异, 该基因的密码子使用及其相应的核糖体结合位点的兴趣, 以及使用的表达载体的类型 62 ,63岁。由于这些原因, 一些蛋白质可能无法表达以及模型蛋白 sfgfp, 导致减少体积产量从 cfps 反应。
所介绍的 cfps 技术的局限性包括, 它可能并不直接适用于无细胞的所有应用, 如代谢工程和表达条件的调整, 而无需对协议进行额外的修改。然而, 我们相信, 该协议将为在新实验室建立 cfps 平台提供基础, 并为非专家提供在其实验室实施无细胞介绍反应的能力。在初步实施后, 研究人员可以对该平台进行实验, 根据该领域的其他文献, 对更具体的应用进行自己的修改。
cfps 平台的成本为0.021 美元/μg 蛋白 (不包括人工和设备成本), 使我们的系统在不影响轻松的反应设置的情况下, 通过商业套件具有竞争力的价格。对每μl 反应的比较成本的评估显示了类似的趋势 (补充图 1)。我们估计所有试剂的启动成本为 4, 500 美元, 声纳等专用设备的额外成本为 3, 200 美元。完成这个过程的人的时间估计是 ~ 26小时的所有试剂准备从地面上。然而, 一旦准备好了大量的试剂, 对劳动力的需求就会大幅减少。此外, 随着平台经验的积累, 我们建议扩大细胞生长、提取物制备和试剂制备的规模, 以最大限度地提高时间效率。考虑到启动成本, 我们建议使用 cfps 平台, 用于合成生物学、高通量工作和蛋白质表达条件, 这些应用与传统的蛋白质表达平台不兼容, 因为这些平台与细胞的生物化学和生存能力的限制。在 cfps 平台启用所需技术的这些特殊情况下, cfps 在体内表达的更高成本是合理的。
cfps 平台的持续发展可能为生物技术工作提供更广泛的效用, 例如酶途径的代谢工程、传统难治性蛋白质、非标准氨基酸的生产和表征纳入和非天然蛋白的表达, 分层医学制造, 并扩大到实验室的 stem 教育 64, 65,66. 66. 66.正在进行的对 cfps 平台进行详细定性的努力将进一步支持这些努力。更好地了解细胞提取物的组成将导致在反应条件下提高反应产率和灵活性的持续改进 61,67,68。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
提交人声明, 他们没有相互竞争的经济利益或其他利益冲突。
Acknowledgments
作者希望感谢詹妮弗·范德凯伦博士、安德烈·劳布舍和托尼·图雷托的技术支持, 他们是高卫理、莱恩·威廉姆斯和克里斯托弗·海特的有益讨论。作者还感谢比尔和琳达·弗罗斯特基金、雪佛龙生物技术应用研究捐赠赠款中心、cal poly 研究、学术和创意活动赠款方案 (rsca 2017) 提供的资金支持,和国家科学基金会 (nsf-1708919)。mzl 承认加州州立大学研究生助学金。mcj 感谢陆军研究办公室 w911nf-16-1-072、国家科学基金会提供 mcb-1413563 和 mcb-1716766, 空军研究实验室英才中心赠款 fa8500-15-2-51818, 国防威胁减少机构赠款hdtra1-15-15-10052/p00001, david 和 lucile packard 基金会、camille dreyfus 教师-学者方案、能源部 ber grant de-sc0018249、人类前沿科学方案 (rgp152017)、doe 联合基因组 etop 和芝加哥生物医学联合会, 得到芝加哥社区信托基金的支持。
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}