

Summary
Dit protocol detailleert de stappen, kosten, en apparatuur die nodig is voor het genereren van E. coli-op basis van cel extracten en uit te voeren in vitro eiwit synthese reacties binnen 4 dagen of minder. Om de invloed van de flexibele aard van dit platform voor brede toepassingen, bespreken we reactie voorwaarden die kunnen worden aangepast en geoptimaliseerd.
Abstract
In de afgelopen 50 jaar, heeft cel-vrij proteïne synthese (GVB) ontpopt als een krachtige technologie om uit te rusten van de transcriptionele en translationele capaciteit van cellen in een reageerbuis. Door het wegnemen van de noodzaak om de levensvatbaarheid van de cel, en door het wegnemen van de cellulaire barrière, is GVB fundamenteel aan opkomende toepassingen in de biomanufacturing van oudsher uitdagende eiwitten, evenals toepassingen, in snelle prototyping voor Metabole engineering en functionele genomica. Onze methoden voor de uitvoering van een e-coli-gebaseerde GVB platform dat nieuwe gebruikers toegang tot veel van deze toepassingen. We beschrijven hier, methoden om te bereiden extract door middel van verrijkte media, verbijsterd kolven en een reproduceerbare methode van afstembare ultrasoonapparaat gebaseerde cellysis. Dit extract kan vervolgens worden gebruikt voor eiwit expressie geschikt voor het produceren van 900 µg/mL of meer van de super map groen fluorescent proteïne (sfGFP) in enkel 5 h van experimentele opstelling van data-analyse, gezien het feit dat geschikt reagens voorraden zijn vooraf opgesteld. De kosten van de geschatte opstarten voor het verkrijgen van reagentia is $4,500 die duizenden reacties op een geraamde kostprijs van $0.021 per µg eiwit geproduceerd of $0.019 per µL van reactie zal ondersteunen. De eiwit expressie methoden spiegel daarnaast het gemak van de setup van de reactie in de verkrijgbare systemen als gevolg van optimalisatie van reagens voormengsels, tegen een fractie van de kosten gezien. Zodat de gebruiker om de invloed van de flexibele aard van het GVB platform voor brede toepassingen, hebben we geconstateerd dat een aantal aspecten van het platform dat kan worden afgestemd en geoptimaliseerd afhankelijk van de beschikbare middelen en de eiwit expressie resultaten gewenste.
Introduction
Cel-vrij proteïne synthese (GVB) heeft ontpopt als een technologie die heeft een aantal nieuwe mogelijkheden voor de productie van eiwitten, functionele genomica, Metabole engineering en meer in de laatste 50 jaar1,2ontgrendeld. In vergelijking met standaard in vivo eiwit expressie platformen, GVB biedt drie belangrijke voordelen: 1) de cel-vrije aard van het platform mogelijk maakt de productie van eiwitten dat zou potentieel giftige of vreemd aan de cel3,4 ,5,6; 2) inactivatie van genomic DNA en de invoering van een sjabloon DNA codering van de gene(s) van belang kanaal alle systemische energie binnen de reactie op de productie van de expressie gebrachte eiwitten van belang; en 3) het open karakter van het platform kan de gebruiker wijzigen en controleren van de omstandigheden en de samenstelling in real-time7,8. Deze directe toegang tot de reactie ondersteunt de vergroting van biologische systemen met uitgebreide oplossingen en redox voorwaarden voor de productie van nieuwe eiwitten en de afstemming van metabole processen2,9, 10. directe toegang maakt het ook mogelijk de gebruiker aan het combineren van de GVB-reactie met activiteit testen in een één-pot-systeem voor een snellere design-build-test cycli11. De capaciteit om te voeren van de GVB-reactie in klein volume druppels of op papier gebaseerde apparaten verder ondersteunt high-throughput ontdekking inspanningen en snelle prototyping12,13,14,15 ,16. Als gevolg van deze voordelen en de plug en play-aard van het systeem, GVB uniek heeft een scala aan biotechnologische toepassingen zoals de productie van eiwitten die moeilijk te solubly snel in vivo17, 18,19,20, opsporing van ziekte21,22,23, op vraag biomanufacturing18,24 ,25,26,27, en onderwijs28,29, die allemaal Toon de flexibiliteit en bruikbaarheid van de cel-vrije platform.
GVB systemen kunnen worden gegenereerd uit een verscheidenheid van ruwe lysates van beide cellijnen prokaryote en eukaryote. Dit zorgt voor diverse opties in het systeem van keuze, die elk voor- en nadelen, afhankelijk van de toepassing van belang zijn. GVB systemen ook variëren sterk in voorbereidingstijd, kosten en productiviteit. De meeste vaak gebruikt cel extracten worden geproduceerd van tarwekiemen, konijn Retikulocyt, insect cellen en Escherichia coli cellen, met de laatste is de meest kosteneffectieve tot nu toe terwijl de productie van de hoogste volumetrische opbrengst van eiwit30 . Terwijl andere systemen GVB kunnen nuttig zijn voor hun aangeboren posttranslationele wijziging-machinery, opkomende toepassingen met behulp van de E. coli-gebaseerde machines zijn in staat de kloof te overbruggen door het genereren van site-specifically gefosforyleerd en geglycosyleerde eiwitten op vraag31,32,33,34,35.
GVB reacties kunnen worden uitgevoerd in een batch, continu-uitwisseling cel-vrij (CECF) of continue-stroom cel-vrij (CFCF) formaten. De batch-indeling is een gesloten systeem waarvan reactie levensduur beperkt als gevolg van de afnemende hoeveelheid reactanten en de accumulatie van remmende bijproduct van de reactie is. CECF en CFCF methoden verhogen de levensduur van de reactie, en daardoor leiden tot verhoogde volumetrische eiwit rendementen ten opzichte van de batch-reactie. Dit wordt bereikt doordat de bijproducten van de eiwitsynthese te worden verwijderd uit het reactievat, terwijl nieuwe reactanten worden geleverd in de loop van de reactie2. In het geval van CFCF, kan de proteïne van belang ook worden verwijderd uit de zaal reactie, terwijl in de CECF, de proteïne van belang blijft in de zaal van de reactie van een semi-permeabel membraan36,37samengesteld. Deze methoden zijn vooral waardevol zijn bij het overwinnen van arme volumetrische opbrengst van moeilijk-aan-express proteïnen van belang38,39,40,41,42, 43. De uitdagingen bij de uitvoering van de CECF en CFCF benaderingen zijn dat 1) terwijl ze leiden een efficiënter gebruik van de bio-machine verantwoordelijk voor transcriptie en vertaling tot, zij vereisen met name grotere hoeveelheden van reagentia die een toename van de totale kosten en 2) ze vereisen meer complexe reactie opstellingen en gespecialiseerde apparatuur ten opzichte van de batch formaat44. Met het oog op toegankelijkheid voor nieuwe gebruikers, beschreven de protocollen hierin focus op het formaat van de partij bij reactie volumes van 15 µL met specifieke aanbevelingen voor het verhogen van het volume van de reactie op de schaal van de milliliter.
De hier vermelde methoden niet-deskundigen met basic Labor vaardigheden (zoals studenten) inschakelen voor de uitvoering van de celgroei, uitpakken van de voorbereiding en batch formaat reactie setup voor een e-coli-gebaseerde systeem van het GVB. Deze aanpak is rendabel commercieel beschikbare kits t.o.v. zonder in te boeten op het gemak van kit gebaseerde reactie setup. Deze aanpak maakt bovendien toepassingen in het laboratorium en in het veld. Bij de beslissing om de implementatie van GVB, moeten nieuwe gebruikers grondig evalueren de werkzaamheid van conventionele eiwit expressiesystemen voor opstarten investeringen, zoals GVB niet kan superieur in ieder geval. De hier beschreven methoden voor het GVB zodat de gebruiker direct om een verscheidenheid van toepassingen, met inbegrip van functionele genomica, high-throughput testen, de productie van eiwitten die hardnekkige voor in vivo expressie, evenals veld zijn toepassingen met inbegrip van biosensoren en educatieve kits voor synthetische biologie. Extra toepassingen zoals Metabole engineering, afstemming van eiwit expressie voorwaarden, opsporing van de ziekte, en schaal-up met behulp van CECF of CFCF methoden zijn nog steeds mogelijk, maar wellicht ervaring met het GVB-platform voor de verdere wijziging van de reactie voorwaarden. Onze methoden combineren groei in de verrijkte media en verbijsterd kolven, met relatief snelle en reproduceerbare methoden van lysis van de cel via ultrasoonapparaat, gevolgd door een vereenvoudigde GVB reactie setup dat gebruik maakt van geoptimaliseerde premixen45. Terwijl de cellulaire groei methoden hebben enigszins worden gestandaardiseerd op dit gebied, lopen methoden voor lysis van de cel sterk uiteen. Naast ultrasoonapparaat omvatten gemeenschappelijke lysis methodes gebruik van een Franse pers, een homogenizer, kraal drijvers, of lysozym en andere verstoring van de biochemische en fysische methoden46,47,48, 49. met behulp van onze methoden ongeveer 2 mL van ruwe cel extract worden verkregen per 1 liter van cellen. Deze hoeveelheid cel extract kan steunen vier honderd 15 µL GVB reacties, elke producerende ~ 900 µg/mL verslaggever sfGFP eiwit uit de sjabloon plasmide pJL1-sfGFP. Deze methode kost $0.021/µg sfGFP geproduceerd ($.019/µL van reactie), exclusief de kosten van arbeid en uitrusting (aanvullende figuur 1). Vanaf de nul, deze methode toegepast kunnen worden in 4 dagen door een enkele persoon en herhaal GVB reacties kunnen worden voltooid binnen uur (Figuur 1). Bovendien kan het protocol worden opgeschaald in volume voor grotere partijen reagens voorbereiding aan de behoeften van de gebruiker. Nog belangrijker is, kan het protocol gepresenteerd hier toegepast worden door laboratorium opgeleid niet-deskundigen zoals Voorgraadse studente, zoals zij alleen basic Labor vaardigheden vereist. De hieronder beschreven procedures, en de begeleidende video zijn specifiek ontwikkeld ter verbetering van de toegankelijkheid van de E. coli GVB platform voor breed gebruik.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Media voorbereiding en celgroei
-
Dag 1
- Streak E. coli BL21*(DE3) cellen uit een glycerol voorraad op een bord van de agar LB en incubeer gedurende ten minste 18 uur bij 37 ° C.
- 50 mL van LB-media en autoclaaf de oplossing voor te bereiden op een vloeibare cyclus gedurende 30 minuten bij 121 ° C. Bewaren bij kamertemperatuur.
-
Dag 2
- 750 mL 2 x YTP media en 250 mL 0,4 M D-Glucose-oplossing te bereiden zoals beschreven in de aanvullende informatie.
- Giet de 2 x YTP media in een gesteriliseerde met autoclaaf 2.5 L verbijsterd kolf en de D-Glucose-oplossing in een gesteriliseerde met autoclaaf 500 mL glazen fles. Autoclaaf beide oplossingen op een vloeibare cyclus gedurende 30 minuten bij 121 ° C.
- Zorg ervoor dat beide steriele oplossingen bij 37 ° C worden opgeslagen als celgroei wordt uitgevoerd op de volgende dag, naar groeicijfers na inoculatie maximaliseren. Combineer oplossingen niet tot inenting.
Opmerking: Oplossingen kunnen worden opgeslagen bij 4 ° C gedurende 1-2 d indien nodig, hoewel de 2 x YTP media is zeer gevoelig voor besmetting. - Start een overnachting cultuur van BL21(DE3) door het enten van 50 mL van LB-media met een één kolonie van BL21(DE3) met behulp van een gesteriliseerde lus en steriele techniek om verontreiniging te voorkomen.
- Plaats van de 50 mL BL21*(DE3) LB cultuur in een 37 ° C 250 rpm schudden incubator en groeien 's nachts voor 15-18 h.
- Bereiden en steriliseren van alle materialen die nodig zijn voor dagen 3 en 4, met inbegrip van: twee flessen van 1 L centrifuge, 4 x koud 50 mL conische buizen (wegen en record massa's van drie), en vele 1,5 mL microfuge buizen.
-
Dag 3
- Verwijder de 50 mL overnachting cultuur van BL21*(DE3) in LB uit de schudden incubator en meten van de OD600 op een spectrofotometer met behulp van een 1:10 verdunning met LB-media. Berekenen van het volume van overnachting cultuur aan 1 L van media voor een startende OD600 0.1 moet worden toegevoegd (bijvoorbeeld, als een OD600 van een 1:10 verdunning wordt gelezen als 0.4, Beënt 25 mL van de onverdunde OD600 = 4.0 overnachting cultuur in 1 L van 2 x YTP G).
- Verwijder de verwarmde 2 x YTP media en D-Glucose oplossingen van de 37 ° C incubator, samen met de 50 mL LB cultuur. Met behulp van steriele techniek, giet zorgvuldig de D-Glucose-oplossing in de 2 x YTP media (het vermijden van de zijkanten van de verbijsterd kolf).
Opmerking: Toevoeging van D-Glucose voltooit het recept voor 1 liter 2 x YTPG. - Behoud van steriele techniek, Beënt de 1 L van 2 x YTPG oplossing met het juiste bedrag van de 50 mL cultuur om te beginnen met de cultuur van de 1 L op een 0.1 OD600. Onmiddellijk plaats de geënte 1 L-cultuur in een 37 ° C incubator schudden bij 200 omwentelingen per minuut.
- Neem de eerste OD600 lezen na het eerste uur van groei (lag fase typische duurt 1U). De cultuur niet worden verdund. Blijven metingen OD600 ongeveer elke 20-30 min tot OD600 0.6 bereikt.
- Bij het bereiken van de OD600 = 0,6, voeg 1 mL 1 M IPTG (eindconcentratie in 1 L cultuur = 1 mM) aan de 2 x YTPG cultuur.
Opmerking: Ideale inductie OD600 is 0,6; een aantal 0.6-0.8 is echter aanvaardbaar. Inductie door IPTG is voor de endogene productie van T7 RNA Polymerase (T7RNAP). - Na inductie, meten de OD600 ongeveer elke 20-30 min tot het 3.0 bereikt.
Opmerking: Afkoelen in de centrifuge tot 4 ° C gedurende deze tijd. Bereiden van koude S30 buffer zoals beschreven in de aanvullendeinformatie. Als de buffer S30 is voorbereid, ervoor zorgen dat DTT niet tot de dag van gebruik wordt toegevoegd. - Zodra de OD600 bereikt 3.0 (figuur 2A), giet de cultuur in een koude 1 L centrifuge fles in een bad met ijs-water. Bereiden een water gevulde 1 L centrifuge fles gelijk gewicht moet worden gebruikt als een evenwicht in de centrifuge.
Opmerking: Extinctie waarden verschillen van instrument tot instrument. Terwijl de OD600 van oogst van BL21(DE3) niet een gevoelige variabele is, is het aanbevolen dat de gebruiker evalueren en optimaliseren van deze variabele als een probleemoplossing maatregel. Grotere spectrofotometers kunnen leiden tot relatief lagere OD600 lezingen in vergelijking met kleinere cuvette gebaseerde spectrofotometers. - Centrifugeer het 1 L flessen voor 10 min op 5.000 x g en 10 ° C tot pellet cellen.
- Langzaam giet af de bovendrijvende substantie en de afzet van het volgens de instelling van biologisch afval procedures. Plaats de pellet op ijs.
- Met behulp van een steriele spatel, schraap de cel pellet uit de fles centrifuge en overbrengen naar een conische buis van koude 50 mL.
- Voeg 30 mL koud S30 buffer aan de conische buis en resuspendeer de pellet cel door vortexing met korte uitbarstingen (20-30 s)- en rusttijden (1 min) op ijs totdat volledig geresuspendeerde met geen brokken.
- Zodra de pellet is volledig geresuspendeerde, gebruikt u een ander conische tube van 50 mL met water als een evenwicht en Centrifugeer gedurende 10 min op 5000 x g en 10 ° C (vooraf gekoeld tot 4 ° C).
Opmerking: Dit voltooit de 1st 3 wasbeurten vereist tijdens het oogsten van de cellen. - Giet het supernatant en de afzet van het volgens de instelling van biologisch afval procedures. Resuspendeer de pellet met 20-25 mL koud S30 buffer en Centrifugeer gedurende 10 min bij 5000 x g en 10 ° C (vooraf gekoeld tot 4 ° C).
Opmerking: Dit voltooit de 2nd van 3 wasbeurten. - Nogmaals, uitstorten de bovendrijvende substantie en de afzet van het volgens de instelling van biologisch afval procedures. Voeg precies 30 mL S30 buffer en vortex opnieuw naar resuspendeer de pellet.
- Met behulp van de 3 conische buisjes in de vooraf gewogen, koude 50 mL en een serologische pipet-vuller met een steriele Pipet, aliquoot 10 mL geresuspendeerde pellet/S30 buffer mengsel in elk van de 3 conische buisjes.
Opmerking: De cellen te splitsen in 3 buizen is niet vereist, maar deze stap leidt tot kleinere cel pellets (~ 1 g) voor meer gemak op latere stappen. - Centrifugeer alle buizen, met behulp van passende saldi, zo nodig, voor 10 min op 5000 x g en 10 ° C (vooraf gekoeld tot 4 ° C).
Opmerking: Dit is de laatste wassen stap voltooid. - Giet het supernatant en de afzet van het volgens de instelling van biologisch afval procedures. Verwijderen van de overtollige S30 buffer door zorgvuldig het afvegen van de binnenkant van de conische buis en cap met een schone tissue; Raak de pellet.
- Weeg de buizen op een analytische balans en het gewicht van de laatste pellet op elke buis.
Opmerking: Het protocol kan worden onderbroken op dit punt. De pellets kunnen flash in vloeibare stikstof bevroren en opgeslagen bij-80 ° C gedurende maximaal een jaar totdat het nodig is voor de bereiding van het extract.
2. de ruwe cel Extract voorbereiding - dag 4
- Voor de bereiding van het extract, houden cellen koud op ijs tijdens elke stap. Voeg 1 mL koude S30 buffer per 1 g van cel massa van de pellet. Zorg ervoor dat dithiothreitol (DTT) is aangevuld tot de buffer S30 aan een definitieve concentratie van 2 mM.
Opmerking: Afkoelen in de microcentrifuge tot 4 ° C gedurende deze tijd. - Resuspendeer de pellet cel door vortexing met korte uitbarstingen (20-30 s)- en rusttijden (1 min) op ijs totdat volledig geresuspendeerde. Als resuspensie moeilijk is, laat u de pellets op ijs gedurende 30 minuten om te ontdooien.
- Pipetteer 1,4 mL geresuspendeerde cellen in een 1,5 mL microfuge buis.
- Plaats een 1,5 mL tube met 1,4 mL geresuspendeerde cellen in een bad met ijs water in een bekerglas. Bewerk ultrasone trillingen ten voor 45 s op gevolgd door 59 s uit voor 3 totale cycli, met amplitude ingesteld op 50%. Sluit en het omkeren van de buizen voorzichtig mengen off perioden. In totaal 800-900 J van energie te leveren aan elke 1,5 mL microfuge buis met 1,4 mL geresuspendeerde cellen (figuur 3A & 3B).
Opmerking: Deze stap is gevoelig voor het ultrasoonapparaat type en model gebruikt en moet worden geoptimaliseerd als apparatuur anders is dan die voor deze procedure. Twee complementaire benaderingen kunnen worden gebruikt voor schaal-up de hoeveelheid extract bereid tijdens deze stap: 1) meerdere 1,5 mL microfuge buizen parallel kunnen worden sonicated, en/of 2) grotere volumes kunnen worden sonicated in conische buisjes (maximaal 15 mL cel resuspensie per buis) , 29,45schalen van de hoeveelheid energie geleverd zoals hiervoor is beschreven. - Onmiddellijk nadat ultrasoonapparaat voltooid is, 4.5 µL van 1 M DTT (ter aanvulling van een extra 2 mM DTT) toevoegen in de 1,4 mL lysate en omkeren van meerdere keren te mengen. Plaats de buis op ijs. Herhaal stap 2.4 en 2.5 voor elke extra buizen van geresuspendeerde cellen alvorens te centrifugeren.
- Microcentrifuge monsters op 18.000 x g- en 4 ° C voor 10 min (Figuur 3 c).
- Pipetteer de bovendrijvende substantie in een nieuwe 1,5 mL microfuge buis. Niet storen de pellet; het is beter dan sommige supernatant achterlaten om zuiverheid dan om de pellet te verstoren in inspanningen te maximaliseren van de opbrengst.
- Incubeer de bovendrijvende vloeistof uit de vorige stap bij 250 rpm en 37 ° C gedurende 60 minuten door taping van de buizen aan het schudden platform van de incubator (dit is de reactie van de afvoer).
- Microcentrifuge monsters op 10.000 x g- en 4 ° C gedurende 10 minuten.
- Verwijder de bovendrijvende vloeistof zonder verstoring van de pellet en overbrengen naar een nieuwe buis. Maak veel 100 µL aliquots voor extract voor opslag.
Opmerking: Het protocol hier kan worden onderbroken kan, en het extract flash in vloeibare stikstof bevroren en opgeslagen bij-80 ° C voor maximaal een jaar totdat het nodig is voor GVB reacties. Ten minste 5 bevriezen-ontdooien cycli kunnen worden ondergaan zonder afbreuk te doen aan het uitpakken van de productiviteit (Figuur 4).
3. cel-vrij proteïne synthese Batch formaat reacties
- Oplossingen A en B, DNA sjabloon BL21(DE3) extract (als bevroren), T7RNAP en een aliquoot gedeelte van het moleculaire rang water ontdooien.
Opmerking: GVB reactie sjabloon kan gevonden worden in de Aanvullendeinformatie. Oplossingen A en B recepten vindt u in de Aanvullendeinformatie en corresponderen met bepaalde concentraties voor talrijke reagentia ter ondersteuning van het energiesysteem PANOx-SP gebaseerd voor GVB. De rol van elk reagens en aanvaardbare schommelingen in deze reagens concentraties die GVB ondersteunen kunnen geweest vastberaden50. Een T7RNAP zuivering protocol kan worden gevonden in de aanvullendeinformatie51. Aanvullende T7RNAP volumetrische opbrengst kan verhogen maar is niet nodig als T7RNAP wordt veroorzaakt tijdens de celgroei. Plasmide DNA sjabloon (pJL1-sfGFP) kan bereid worden met behulp van een maxiprep kit met twee wasbeurten met behulp van de was-buffer in de kit, gevolgd door een post-processing DNA-opruiming met behulp van een PCR zuivering kit (figuur 2B). Lineaire DNA-sjablonen kunnen ook worden gebruikt in GVB reacties. - Label de benodigde hoeveelheid microfuge buizen nodig voor GVB reacties.
Opmerking: Reacties kunnen worden uitgevoerd in verschillende maten van het schip, maar een kleiner vaartuig kunt verminderen volumetrische eiwit opbrengst (figuur 2C). Opschalen van een reactie in het dezelfde grootte vaartuig kan volumetrische opbrengst, als een functie van het verlagen van de uitwisseling van zuurstof, als gevolg van een daling in het oppervlak volumeverhouding ook verminderen. Wanneer het volume van de reactie boven 100 μl verhogen, is het aanbevolen om het gebruik van platte bodem goed platen 31,37,52. - Voeg 2.2 µL van oplossing A, 2.1 µL van oplossing B, 5 µL van BL21*(DE3) extract, 0,24 μg T7RNAP (16 μg/mL-eindconcentratie), 0,24 ng van DNA sjabloon (eindconcentratie is 16 ng/mL) en water toe zodat het eindvolume 15 µL.
Opmerking: Vortex oplossingen A en B vaak tijdens de installatie van de reactie te voorkomen van sedimentatie van componenten en ervoor zorgen dat elke reactie een homogene aliquot van elke oplossing ontvangt. Vermijd vortexing het extract, in plaats daarvan het omkeren van de buis te mengen. - Nadat alle reagentia zijn toegevoegd aan de reactie, mengen elke buis door pipetteren omhoog en omlaag of zachtjes vortexing en tegelijkertijd te garanderen dat het uiteindelijke reactiemengsel worden gecombineerd tot een enkel 15 µL kraal aan de onderkant van de 1,5 mL microfuge buis.
- Plaats elke reactie in de 37 ° C incubator zonder schudden voor 4 uur of 30 ° C's nachts.
Opmerking: Succesvolle reacties kunnen worden kwalitatief beoordeeld visueel op basis van de groene kleur van het product van de sfGFP binnen het GVB reactiemengsel (figuur 3D). Uitdrukking van de proteïne van belang kan er ook worden bevestigd door SDS-PAGE (aanvullende figuur 2).
4. kwantificering van de verslaggever proteïne, [sfGFP]
- 48 µL van 0,05 M HEPES, pH 8, in elk goed die nodig zijn voor de kwantificering (meestal uitgevoerd in drievoud per reactiebuis) laden.
- Verwijder reacties van incubator. Pipetteer omhoog en omlaag als u wilt mengen elke reactie, dan breng 2 µL van de reactie in de 48 µL van 0,05 M HEPES, pH 8. Pipetteer op en neer opnieuw in de put te mengen.
- Zodra alle reacties worden geladen en gemengd, plaats de 96 goed plaat in de Fluorimeter en meten van de sfGFP eindpunt fluorescentie.
Opmerking: Excitatie en emissie golflengten voor sfGFP fluorescentie kwantificering zijn 485 nm en 510 nm, respectievelijk. - Met behulp van een eerder gegenereerde standaard curve, bepalen de [sfGFP] van de verkregen fluorescentie-lezingen.
Opmerking: Instructies voor het genereren van een standaard curve van sfGFP concentratie ten opzichte van de intensiteit van de fluorescentie vindt in aanvullendeinformatie (aanvullende figuur 3). Gebruikers zullen moeten vaststellen van een standaard curve voor hun instrument omdat instrument gevoeligheid kan variëren.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Wij hebben voorgesteld een ultrasoonapparaat gebaseerde E. coli extract voorbereiding protocol die met cijfer 1 demonstreren de procedurele verdeling over elke dag gedurende een vier-daagse span, kan worden voltooid. Er is buigbaarheid naar de stappen die in elke dag met diverse onderbreken punten kan worden voltooid, maar we hebben gevonden deze werkstroom het meest effectief uit te voeren. Bovendien zijn zowel de cel pellets (stap 1.3.18) en volledig bereid extract (stap 2.10) stabiel op-80 ° C voor ten minste een jaar, waardoor de gebruiker kan het maken van grotere voorraden van elk op te slaan voor gebruik op een later moment17. Niet alleen is het extract stabiel over lange perioden, maar het extract kan ook ondergaan ten minste vijf bevriezen dooi cycli zonder een aanzienlijk verlies aan productiviteit (Figuur 4). Dit zorgt voor grotere hoeveelheden voor extract voor meermalig gebruik worden opgeslagen als diepvries opslagruimte beperkt is. Wij raden echter aan meerdere kleinere aliquots (~ 100 μl) van extract waar mogelijk.
Met elke nieuwe extract voorbereiding, is het raadzaam dat de gebruiker een magnesium titratie uitvoert om te bepalen van de optimale hoeveelheid magnesium voor die partij voor extract. Gebruikers kunnen kwantificeren-charges-variabiliteit in totaal eiwitconcentratie van het extract van de cel door analyse van Bradford. Want hoger uitvoeren extracten, we meestal totaal eiwit concentraties van 30-50 mg/mL zien, en binnen dit gamma er geen voor de hand liggende direct verband tussen totale proteïne concentraties en cel extract prestaties bestaat. Daarom is het raadzaam dat gebruikers magnesium concentraties dienovereenkomstig stemmen om ervoor te zorgen dat proteïnen en nucleic zuur functionaliteit voor elke batch extract worden gemaximaliseerd. Magnesium niveaus zijn belangrijk voor goede DNA-replicatie, transcriptie en vertaling, maar buitensporige niveaus schadelijk kunnen zijn voor deze processen53. Om aan te tonen deze afhankelijkheid, wij hebben uitgevoerd een co titratie van magnesium en haal volume om te bepalen van de optimale combinatie die minimaliseert u de hoeveelheid extract nodig, met behoud van een productieve reactie (Figuur 5). Uit dit experiment raden we 5 µL van het uittreksel en 10 mM Mg2 + voor extract met een totale proteïnegehalte van 30 mg/mL, met het oog op meer dan 1000 μg/mL sfGFP.
Onze ervaring met GVB heeft ook konden we stappen binnen het protocol dat kan worden gevarieerd zonder afbreuk te doen aan de algehele productiviteit van het systeem, en anderen die integraal deel uitmaken voor een hoog presterende GVB systeem bepalen. Meest in het bijzonder, de laatste van de OD600 van cel oogst heeft geen aanzienlijk invloed op de uiteindelijke uitvoer van de GVB-reactie en cellen kunnen redelijkerwijs worden geoogst overal van 2.7-4.0 OD600. Dit vertegenwoordigt de vroege exponentiële fase van groei waar ribosoom concentratie per cel is de hoogste en de vertalende machine is de meest actieve ter ondersteuning van de snelle groei. Deze observatie gebruikers flexibiliteit voor het optimaliseren van hun eigen procedures. Het is raadzaam om de oogsten op ongeveer 3.0 OD600 om te vangen van de cellen in een OD600 dichter aan 3.3 door de tijd te oogsten is voltooid (figuur 2A). Variabelen die hebben invloed op GVB opbrengsten omvatten sjabloon DNA kwaliteit, de reactie vaartuig grootte en de relatieve hoeveelheden van cel extract en magnesium ion aanwezig in de reactie. We hebben DNA kwaliteit op het opmerkelijke-charges-variatie hebben gevonden. Om dit oplossen, is het raadzaam dat gebruikers DNA zuiveren via een midi of maxi prep, gevolgd door een extra DNA opschonen stap op de DNA zuivering kolom in de maxiprep gebruikt, of na reiniging met behulp van een extra DNA opruimen kit. Dit verbetert de reproduceerbaarheid in DNA kwaliteit voor GVB reacties en resultaten in robuustere eiwitproductie (figuur 2B). Het reactievat ook gevolgen volumetrische opbrengst, zodanig dat de productie van eiwitten van identieke reactie opstellingen in verschillende delen van het schip kan oplopen tot 40% verschillen. Het heeft zijn theorie dat de impuls in in volumetrische opbrengst waargenomen in grotere schepen is te wijten aan een verhoogde oppervlakte van het reactiemengsel, waardoor betere uitwisseling van zuurstof (figuur 2C), en anderen hebben volumetrische opbrengst verder gestimuleerd door het uitvoeren van De reacties van het GVB in grote flat-bodem platen, die we voor reacties over 100 µL17,31,37,52 aanbevelen.

Figuur 1 : Tijdlijn voor cultuur groei, productie van cel extract, setup en kwantificering van GVB reacties. De gebruiker kan het GVB platform voor hun onderzoek aanvragen via deze werkstroom vierdaags implementeren. Bereiding van het reagens vertegenwoordigt de primaire tijd en kosten voor de eerste ronde van dit experiment en vermindert aanzienlijk nadat reagentia voorraden zijn gevestigd. Bovendien, kunnen cel pellets en bereid cel extract worden opgeslagen voor meer dan een jaar bij-80 ° C, waardoor de gebruiker om te beginnen de tijdlijn op verschillende stappen voor snellere resultaten. De gebruiker kan ook op verschillende stappen om te wijzigen de tijdlijn van deze workflow onderbreken. Klik hier voor een grotere versie van dit cijfer.

Figuur 2 : Halster voorwaarden voor GVB en de gevolgen voor de opbrengst van de volumetrische reactie. A. Extract productiviteit vergelijking op basis van het oogsten van BL21(DE3) cellen in verschillende OD600 lezingen. Op basis van dit perceel, raden we oogsten op een OD-600 voor 3.3 tot ten minste 1000 µg Mn/mL doel eiwit. Reacties werden uitgevoerd op 15 μL schaal in 1,5 mL microfuge buizen. B. vergelijking van twee maxiprep wassen de protocollen van DNA met en zonder na zuivering DNA-opruimingen. pJL1-sfGFP-plasmiden onderging een maxiprep met één of twee wasbeurten, gevolgd door een opruimen na zuivering door PCR zuivering kit. Om te bereiken ~ 900 µg/mL eiwit expressie, wij stellen voor het uitvoeren van een opruiming DNA na zuivering ongeacht het aantal maxiprep wast. Reacties werden uitgevoerd op 15 μL schaal in 1,5 mL microfuge buizen. C. 15 µL GVB reacties uitgevoerd in verschillende schepen variërend van 2 mL tot 0,6 mL microfuge buizen. "Neg" vertegenwoordigt een negatieve controle waar geen DNA-sjabloon aan de reactie toevoegde. Alle foutbalken vertegenwoordigen 1 standaarddeviatie van drie onafhankelijke reacties voor elke voorwaarde, elk waarvan werd gekwantificeerd in drievoud. Klik hier voor een grotere versie van dit cijfer.

Figuur 3 : Sleutel procedurele opstellingen en resultaten voor het maken van productieve uitpakken. A. juiste instelling van ultrasoonapparaat ijswater bad om koeling van monster terwijl warmte wordt gegenereerd tijdens de ultrasoonapparaat. B. 1,5 mL microfuge buis met cel van de geresuspendeerde pellet pre (links) en post (rechts) ultrasoonapparaat. De resulterende lysate een donkerder tint in vergelijking met de cel van de geresuspendeerde pellet moet worden weergegeven. C. juiste scheiding van de bovendrijvende substantie en pellet van cel lysate na 18.000 x g centrifugeren. D. GVB reacties na 4 uur incubatie bij 37 ° C. 1,5 mL microfuge buis aan de rechterkant (succesvolle reactie) toont zichtbaar fluorescentie van het eiwit sfGFP verslaggever bij ~ 900 μg/mL. De negatieve controle linkertube, ontbreekt sjabloon DNA en simuleren van een mislukte reactie, verschijnt een duidelijke oplossing met geen fluorescentie. Klik hier voor een grotere versie van dit cijfer.

Figuur 4 : Verandering in de eiwit expressie, meer dan 5 bevriezen-ontdooien cycli voor GVB halen. Uittreksel bereid uit de dezelfde groei onderging vijf bevriezen dooi cycli via vloeibare stikstof flash bevriezing gevolgd door ontdooien op ijs. Geen significante veranderingen in uittreksel productiviteit voor het uitdrukken van sfGFP werden gezien over de vijf cycli bevriezen-ontdooien. Reacties werden uitgevoerd op 15 μL schaal in 1,5 mL microfuge buizen. "Neg" vertegenwoordigt een negatieve controle waar geen DNA-sjabloon aan de reactie toevoegde. Alle foutbalken vertegenwoordigen 1 standaarddeviatie van drie onafhankelijke reacties voor elke voorwaarde, elk waarvan werd gekwantificeerd in drievoud. Klik hier voor een grotere versie van dit cijfer.

Figuur 5 : GVB voor reacties met variërende [Mg2+] en extract volumes versus [sfGFP]. [Mg2 +] varieerden van 8 mM tot 14 mM met 2 mM stappen en pak volumes varieerde van 3 µL tot 7 µL met 1 µL stappen. De kleur code vertegenwoordigt [sfGFP] geproduceerd van hoog (rood) naar laag (paars). Reagens efficiëntie maximaliseren terwijl het handhaven van hoge eiwitproductie, raden we 5 µL van extract en 10 mM Mg2+ voor extracten die een totaal eiwitgehalte van ~ 30 mg/mL, hebben zoals bepaald door analyse van Bradford. Originele punten voor het genereren van de contour plot waren gebaseerd uit eindpunt fluorescentie van drie onafhankelijke reacties voor elke voorwaarde, elk waarvan werd gemeten in drievoud. Reacties werden uitgevoerd op 15 μL schaal in 1,5 mL microfuge buizen. Klik hier voor een grotere versie van dit cijfer.
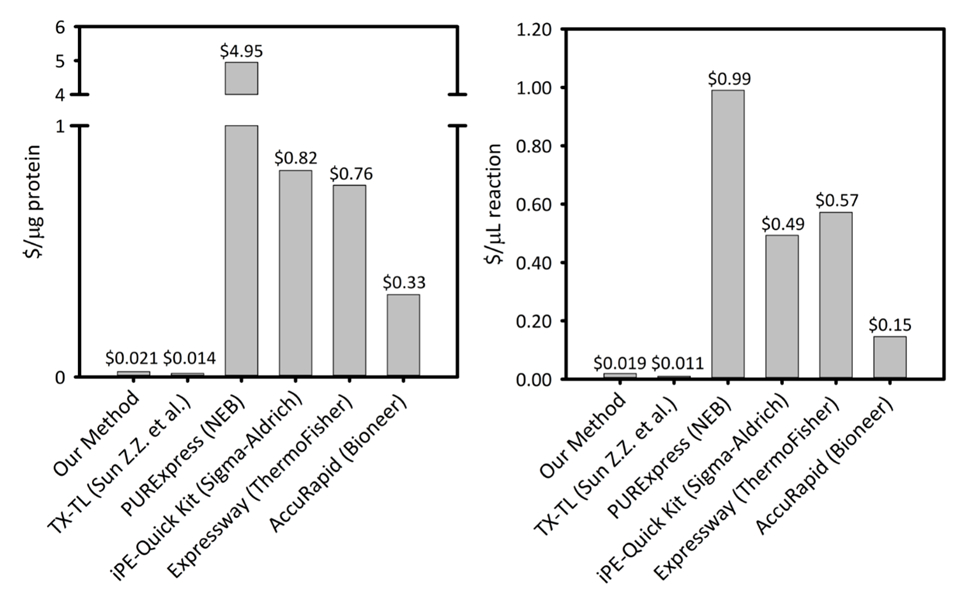
Aanvullende Figuur 1: kost per microgram eiwit geproduceerd en per microliter van reactie op zes cel-vrij proteïne synthese platformen. Ons platform wordt vergeleken met vijf andere cel gratis eiwit synthese kits/platformen met variërend van productiviteit en prijsstelling. Onze ultrasoonapparaat gebaseerd GVB platform is kosteneffectiever in beide $/ µg voor eiwit en $/ µL van reactie dan de meeste commerciële kits en het gemak van een kit voor de installatie van de reactie, terwijl de resterende kosten vergelijkbaar met andere academische GVB-platformen biedt. Klik hier voor een grotere versie van dit cijfer.
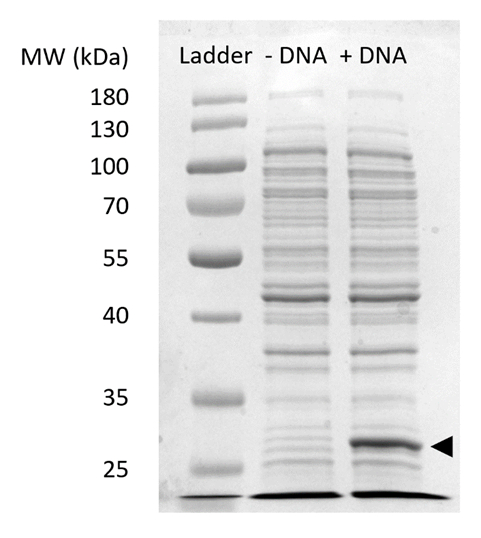
Aanvullende Figuur 2: SDS-pagina van sfGFP expressie in GVB. Cel-gratis eiwit synthese reacties met (+ DNA) en zonder (-DNA) DNA sjabloon voor sfGFP werden uitgevoerd op een 12% SDSPAGE gel om aan te tonen van de expressie van sfGFP waargenomen bij 27 kDa (zwarte pijl). Traditionele SDS-pagina technieken werden gebruikt. Elk monster geladen op de gel opgenomen 18 µg totaal eiwit op basis van Bradford assay kwantificering van totaal eiwit in de cel halen. Op basis van fluorescentie intensiteit metingen en onze standaard curve, schatten we dat het "+ DNA" lane bevat 0,42 µg voor sfGFP. Met het oog op deze monsters, werden GVB reacties uitvoeren op een schaal van 15 µL in 1,5 mL microfuge buizen produceren volumetrische opbrengst conform Figuur 3 c Klik hier voor een grotere versie van dit cijfer.
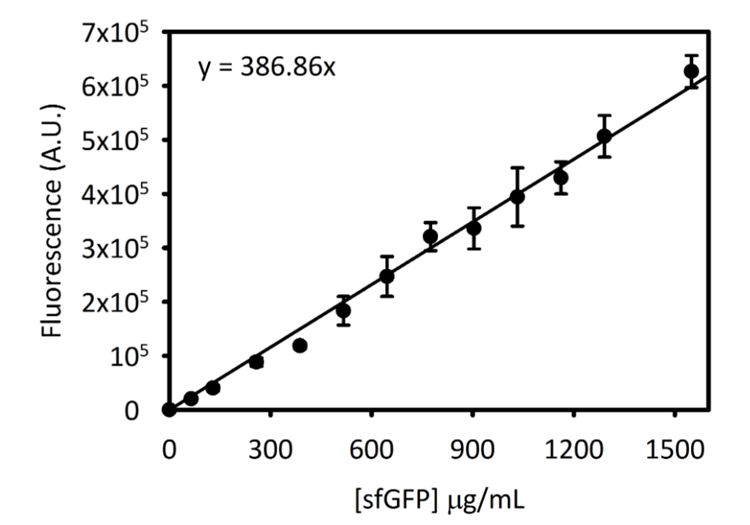
Aanvullende Figuur 3: standaard curve voor sfGFP op Cytation-5. Deze curve werd bepaald met behulp van de methoden die hierboven beschreven. Alle gegevens worden verzameld voor dit manuscript was omgezet van eindpunt fluorescentie lezingen [sfGFP] in µg/mL met behulp van deze standaard curve. Klik hier voor een grotere versie van dit cijfer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Cel-vrije eiwitsynthese heeft ontpopt als een krachtige en activerende technologie voor een verscheidenheid van toepassingen variërend van biomanufacturing tot rapid prototyping van biochemische systemen. De breedte van de toepassingen wordt ondersteund door de capaciteit te controleren, manipuleren, en vergroten van cellulaire machines in real-time. Ondanks de groeiende impact van deze platformtechnologie, brede aanpassing gebleven traag als gevolg van technische nuances bij de uitvoering van de methoden. Door middel van deze inspanning streven we naar eenvoud en duidelijkheid voor de vaststelling van deze technologie in nieuwe labs. Tegen dit einde, onze protocol voor een e-coli-gebaseerde cel-vrij proteïne synthese platform kan worden bereikt binnen een opstarttijd van vier dagen door laboratorium opgeleid niet-deskundigen, zoals Voorgraadse studente (Figuur 1). Bovendien, zodra een voorraad van reagentia en extract worden geproduceerd, kunnen latere GVB batch reacties worden ingesteld, geïncubeerd en gekwantificeerd in enkel 5 h. Een honkslag, 1 L celgroei kan resulteren in genoeg extract voor vierhonderd 15 µL GVB reacties, terwijl één partij voorbereidingen van de andere cel-vrije reagentia duizenden reacties kunnen ondersteunen. Reagens preparaten kunnen ook worden opgeschaald als een nog grotere voorraad nodig is. De GVB-reacties kunnen worden ingesteld op een hoge-doorvoer wijze, met behulp van een 96-wells-plaat of PCR buizen voor het testen van een verscheidenheid van omstandigheden in parallel. Volumetrische opbrengst zal afnemen bij het gebruik van kleinere schepen zoals te zien in figuur 2C. GVB reacties kunnen ook worden opgeschaald van microliters tientallen milliliter van totale reactie volume teneinde het rendement van de totale proteïne voor één voorwaarde opgeeft. Tijdens het schalen volume, is de primaire overweging dat volumetrische reactie opbrengsten afnemen als het oppervlakte gebied-te-volumeverhouding van de reactie37,,52 vermindert. Teneinde schaal-up met behoud van soortgelijke volumetrische opbrengst van eiwit expressie, moeten gebruikers het volume van de reactie gesplitst talrijke reactie vaartuigen en/of vaartuig vergroten. Voor reactie scales variërend van 15 µL - 100 µL in volume, worden talrijke 15 µL reacties parallel aanbevolen. Flat-bodem 24-Wells-platen worden aanbevolen voor reacties meer dan 100 µL in volume, en 12-Wells-platen worden aanbevolen voor reactie volumes meer dan 600 µL. Dergelijke koppelingen reactie volumes en schepen bieden consistentie in volumetrische reactie rendementen op schaalvergroting17,31,37,,52. Buiten deze volumes schaalvergroting kan worden bereikt door gebruik te maken van meerdere putjes van de plaat in parallel. Deze indeling gebruikt, kan de reactie worden geschaald naar meer dan 10 mL totale volume. Optimaliseren van de reactie volume-reactie vaartuig combinatie kan ondersteunen toepassingen van biomanufacturing zonder in te boeten op de productiviteit van de reactie.
Bij het uitvoeren van dit protocol, zijn er een paar essentiële factoren die van invloed zijn volumetrische reactie opbrengsten, alsmede de indicatoren die zijn gekoppeld aan de slecht presterende extract. Met het oog op een juiste lysis en om te voorkomen dat denaturatie van functionele transcriptie/vertaling machines, is het belangrijk te verzachten van de warmte die wordt geproduceerd tijdens cellysis. Dompel de cel hersuspensie in een bad van ijswater tijdens ultrasoonapparaat snel om hitte te verdrijven tijdens ultrasoonapparaat (figuur 3A). Een indicator voor effectieve cellysis is de opkomst van een donkere uitstraling van de cel lysate ten opzichte van pre-sonicated monsters (figuur 3B). Voor de gebruiker de flexibiliteit is de ultrasoonapparaat en sonde weergegeven in figuur 3A aangepast naar een bereik van volumes van 100 µL aan 15 mL geresuspendeerde cellen. Om dit te bereiken, kan de gebruiker het aantal joules geleverd voor lysis van de gewenste hoeveelheid cellen aanpassen. Bovendien kunnen grotere volumes voor extract door twee complementaire benaderingen worden voorbereid. Gebruikers kunnen Bewerk ultrasone trillingen ten meerdere buizen parallel en/of grotere volumes van cel resuspensie, bewerk de schalen van de hoeveelheid energie proportioneel met het volume als eerder beschreven29,45ultrasone trillingen ten. Een andere stap die extract kwaliteit aangeeft is het centrifugeren stap na lysis van de cel. Post lysis van de cel, is het raadzaam centrifugeren op 18.000 x g om een duidelijke scheiding tussen het supernatant (transcriptie/vertaling machines, gefragmenteerde genomic DNA die niet meer sjabloon transcriptie/vertaling functioneert) en de pellet ( ongewenste cellulaire componenten zoals de celmembraan en de neergeslagen eiwitten) (Figuur 3 c). We hebben gevonden dat centrifugeren op 18.000 x g verbetert scheiding, wat resulteert in betere reproduceerbaarheid in vergelijking met draaiingen bij lagere snelheden zoals 12.000 x g. Voor het gemak raden we een tafelblad gekoelde centrifuge, staat voor het bereiken van een minimum van 12.000 x g. Deze stap wordt ook vaak uitgevoerd bij 30.000 x g, die dienen te worden beschouwd als de juiste apparatuur is beschikbaar54,55,56,,57,58, 59 , 60. extract prestaties wordt niet beïnvloed door centrifugeren snelheden bij deze stap, gezien het feit dat goede scheiding wordt bereikt. Wanneer u het gewenste supernatant verwijdert, is het best om te voorkomen dat bewolkt materialen die aanwezig zijn op de grens tussen de bovendrijvende substantie en pellet aangezien deze besmetting de productiviteit van het extract vermindert. Streven naar zuiverheid van het supernatans dat leidt tot meer productieve extracten en is de moeite waard de verminderde hoeveelheid extract verkrijgbaar voor nieuwe gebruikers.
Het is belangrijk om het merk op dat terwijl de methoden die we hebben ingediend reproduceerbaar zijn en kunnen worden uitgevoerd door wetenschappers met minimale deskundigheid, kan er charges- en reactie-naar-reactie variatie. Dit kan worden toegeschreven aan de variatie in de proteomic samenstelling van de lysate na ultrasoonapparaat61. De-charges-variabiliteit die we hebben waargenomen is over het algemeen verminderd bij suppletie met T7RNAP en optimalisatie van magnesium concentraties. Exogene toevoeging van T7RNAP is gemeenschappelijk onder GVB reacties ter ondersteuning van de optimale eiwit expressie, en we vinden dat twee bronnen van T7RNAP - endogene expressie in BL21*(DE3) en de aanvullende T7RNAP om een eindconcentratie van 16 µg/mL - verbetert charges-reproduceerbaarheid voor nieuwe gebruikers45,46. Met ervaring, kunnen de gebruikers hun experimenten voor het gebruik van slechts een enkele bron van T7RNAP desgewenst wijzigen. Kwantificering van het totale eiwitgehalte van een nieuwe batch extract en adequate aanpassing van Mg2 + concentratie kan ook helpen verminderen-charges-variatie in volumetrische eiwit expressie opbrengsten. Variaties in eiwit expressie kunnen ook worden veroorzaakt door verschillen in de omvang en structuur van de proteïne van belang, het codon gebruik van het gen en zijn bijbehorende ribosoom bindende plaats van het gen van belang, evenals de aard van de gebruikte expressie vector62 ,63. Om deze redenen, kunnen sommige eiwitten niet uitdrukken evenals de model eiwit sfGFP, wat resulteert in verminderde volumetrische opbrengst van GVB reacties.
Beperkingen van de gepresenteerde GVB techniek opnemen dat het wellicht niet direct geschikt voor alle toepassingen van cel-vrij, zoals Metabole engineering en het afstemmen van expressie voorwaarden, zonder extra aanpassingen aan de protocollen. Wij zijn echter van mening dat dit protocol zal de basis vormen voor de vaststelling van de GVB-platform in nieuwe laboratoria en niet-experts voorzien in de mogelijkheid om inleidende cel-vrije reacties in hun labs. Na de eerste implementatie, kunnen onderzoekers experimenteren met het platform hun eigen wijzigingen voor meer specifieke toepassingen op basis van andere literatuur in het veld te brengen.
Het GVB platform kosten $0.021/µg eiwit (met uitzondering van de kosten van arbeid en apparatuur), maken ons concurrerend geprijsd met commerciële kits zonder afbreuk te doen aan gebruiksgemak reactie installatie systeem. Beoordeling van vergelijkende kost per µL van reactie laten zien soortgelijke trends (Supplemental Figuur 1). Wij schatten opstartkosten te ~ $4.500 voor alle reagentia, en een extra $3200 voor gespecialiseerde apparatuur, zoals een ultrasoonapparaat. Persoon om deze procedure te voltooien uren worden geschat op ~ 26 h voor alle reagens prep uit de grond omhoog. Echter zodra grote voorraden van reagentia zijn opgesteld, de eisen op arbeid aanzienlijk afnemen. Bovendien, zoals ervaring met het platform is opgedaan, we raden schaalvergroting van de grootte van de celgroei, uitpakken van de voorbereiding en bereiding van het reagens om tijd productiviteit te maximaliseren. Gezien de opstartkosten, is het raadzaam het GVB platform voor toepassingen in de synthetische biologie, hoog-productie inspanningen, en eiwit expressie voorwaarden die niet compatibel zijn met traditionele eiwit expressie platformen verschuldigde in conflict met van de cel Biochemie en levensvatbaarheid beperkingen. In deze speciale gevallen waar de gewenste techniek is ingeschakeld door het GVB-platform, is de grotere kosten van GVB over in vivo expressie gerechtvaardigd.
Verdere ontwikkeling van het GVB-platform is kans op bredere utility om biotechnologie inspanningen zoals de metabole engineering van enzymatische trajecten, productie en karakterisering van traditioneel onoplosbare eiwitten, afwijkende aminozuur opneming en onnatuurlijke eiwit expressie, gestratificeerd geneeskunde productie en uit te breiden buiten het laboratorium in de klas voor STEM onderwijs64,65,66. Deze inspanningen zullen verder worden gesteund door de voortdurende inspanningen voor gedetailleerde karakterisering van de GVB-platform. Een beter begrip van de samenstelling van het extract van de cel zal leiden tot een verdere verfijning naar betere reactie opbrengst en flexibiliteit in reactie voorwaarden61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs verklaren dat zij geen concurrerende financiële belangen of andere belangenconflicten.
Acknowledgments
Auteurs wil erkennen van Dr. Jennifer VanderKelen, Andrea Laubscher, en Tony Turretto voor technische ondersteuning, Wesley Kao, Layne Williams en Christopher Hight voor nuttige discussies. Auteurs erkennen ook financiering van de steun van de Bill en Linda Frost Fonds, Center for Applications in biotechnologie van Chevron biotechnologie toegepast onderzoek Endowment Grant, Cal Poly onderzoek Scholarly en creatieve activiteiten Grant programma (RSCA 2017), en de National Science Foundation (NSF-1708919). MZL erkent de California State University Graduate Grant. MCJ erkent het leger onderzoek Office W911NF-16-1-0372, verleent National Science Foundation MCB-1413563 en MCB-1716766, de Air Force Research Laboratorium Center van Excellence Grant FA8650-15-2-5518, de Defense bedreiging vermindering Agentschap subsidie HDTRA1-15-10052/P00001, de David en Lucile Packard Foundation, het programma van de leraar-Scholar Camille Dreyfus, het departement van energie BER Grant DE-SC0018249, de menselijke grenzen Science Program (RGP0015/2017), de DAMHINDE gezamenlijk genoom Instituut ETOP Grant, en het Chicago biomedische Consortium met steun uit de fondsen Searle op de Chicago Gemeenschap Trust voor steun.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}