

Summary
Questo protocollo dettaglia i passaggi, i costi e attrezzature necessarie per generare e. coli-base di estratti cellulari e implementare in vitro reazioni di sintesi di proteine entro 4 giorni o meno. Per sfruttare la natura flessibile di questa piattaforma per vaste applicazioni, discutiamo le condizioni di reazione che possono essere adattate e ottimizzate.
Abstract
Negli ultimi 50 anni, sintesi di proteina Cell-Free (CFPS) è emerso come una potente tecnologia per sfruttare la capacità trascrizionale e traslazionale delle cellule all'interno di una provetta. Eliminando la necessità di mantenere la vitalità della cellula ed eliminando la barriera cellulare, CFPS è stato fondamentale per le applicazioni emergenti in bioproduzione di proteine tradizionalmente difficili, nonché applicazioni nella prototipazione rapida per Ingegneria metabolica e genomica funzionale. I nostri metodi per l'implementazione di un e. coli-piattaforma basato su CFPS ai nuovi utenti di accedere a molte di queste applicazioni. Qui, descriviamo i metodi per preparare l'estratto attraverso l'uso di media arricchita, boccette sconcertati e un metodo riproducibile di lisi cellulare basato su sonicazione sintonizzabile. Questo estratto quindi può essere utilizzato per l'espressione della proteina in grado di produrre 900 µ g/mL o più di proteina fluorescente verde super cartella (sfGFP) in appena 5 ore dalla messa a punto sperimentale all'analisi dei dati, dato che le scorte di reattivo appropriato sono state preparate in anticipo. Il costo stimato avvio di ottenere reagenti è $4.500 che sosterrà migliaia di reazioni ad un costo stimato di $0,021 per µ g di proteina prodotta o $0,019 per µ l di reazione. Inoltre, i metodi di espressione della proteina specchio la facilità dell'installazione reazione visto nei sistemi disponibili in commercio a causa dell'ottimizzazione di reagente premiscelati, ad una frazione del costo. Al fine di consentire all'utente di sfruttare la natura flessibile della piattaforma CFPS per vaste applicazioni, abbiamo identificato una varietà di aspetti della piattaforma che possono essere ottimizzate e ottimizzato a seconda delle risorse disponibili e gli esiti di espressione della proteina desiderato.
Introduction
Sintesi di proteine senza cellula (CFPS) è emerso come una tecnologia che ha sbloccato una serie di nuove opportunità per la produzione di proteine, genomica funzionale, ingegneria metabolica e più entro gli ultimi 50 anni1,2. Rispetto alla standard in vivo piattaforme di espressione di proteine, CFPS offre tre vantaggi principali: 1) la natura senza cellula della piattaforma consente la produzione di proteine che sarebbe potenzialmente tossici o stranieri per la cella3,4 ,5,6; 2) inattivazione di DNA genomic e l'introduzione di un modello di DNA codificante il gene di interesse canale tutta l'energia sistemica all'interno della reazione per la produzione della proteina di interesse; e 3) la natura aperta della piattaforma consente all'utente di modificare e monitorare le condizioni di reazione e la composizione in tempo reale7,8. Questo accesso diretto alla reazione supporta l'incremento dei sistemi biologici con composizioni chimiche espanse e condizioni redox per la produzione di nuove proteine e la messa a punto di processi metabolici2,9, 10. diretto accesso consente inoltre all'utente di combinare la reazione CFPS con analisi di attività in un sistema singolo-pot per cicli di test per la compilazione di una progettazione più rapida11. La capacità di eseguire la reazione CFPS nelle goccioline di piccolo volume o su dispositivi basati su carta maggiori azioni di alto-rendimento individuazione di supporti e prototipazione rapida12,13,14,15 ,16. A seguito di questi vantaggi e la natura di plug and play del sistema, CFPS ha permesso in modo univoco una varietà di applicazioni della biotecnologia come la produzione di proteine che sono difficilmente solubile veloce in vivo17, 18,19,20, rilevamento di22,21,malattia23, sulla domanda bioproduzione18,24 ,25,26,27e formazione28,29, i quali mostrano la flessibilità e l'utilità della piattaforma senza cellula.
Sistemi di CFPS possono essere generati da una varietà di greggio lisati da entrambe le linee cellulari procarioti ed eucarioti. Questo permette diverse opzioni nel sistema di scelta, ciascuno dei quali hanno vantaggi e svantaggi a seconda dell'applicazione di interesse. CFPS sistemi variano anche notevolmente in produttività, costi e tempi di preparazione. La maggior parte utilizzati comunemente cella estratti sono prodotti da germe di grano, dei reticolociti di coniglio, cellule di insetto e cellule di Escherichia coli , con quest'ultimo è il più redditizio per data, producendo il massimo rendimento volumetrico della proteina30 . Mentre altri sistemi CFPS possono essere vantaggiosi per le loro macchine di modificazione post-traduzionale innata, emergendo applicazioni utilizzando il e. coli-base macchine sono in grado di colmare il divario generando site-specifically fosforilato e proteine glicosilate su domanda31,32,33,34,35.
Le reazioni CFPS possono essere eseguite in entrambi batch, scambio continuo senza cellula (CECF) o flusso continuo senza cellula (CFCF) formati. Il formato di batch è un sistema chiuso in cui durata di reazione è limitata a causa della diminuzione quantità di reagenti e l'accumulazione dei sottoprodotti inibitori della reazione. Metodi CECF e CFCF aumentano la durata della reazione e quindi causare rendimenti volumetrici aumentata della proteina rispetto alla reazione batch. Questa operazione viene eseguita consentendo i sottoprodotti della sintesi proteica per essere rimosso dal recipiente di reazione, mentre nuovi reagenti sono forniti nel corso della reazione2. Nel caso di CFCF, la proteina di interesse può essere rimosso anche dalla camera di reazione, mentre in CECF, la proteina di interesse rimane nella camera di reazione, costituito da una membrana semi-permeabile36,37. Questi metodi sono particolarmente preziosi nel superare la scarsa resa volumetrica di difficile--express proteine di interesse38,39,40,41,42, 43. Le sfide nell'attuazione degli approcci CECF e CFCF sono che 1) mentre risultano in un uso più efficiente dell'apparato bio responsabile della trascrizione e traduzione, richiedono in particolare più grandi quantità di reagenti che aumenta il costo complessivo e 2) Essi richiedono configurazioni più complesse di reazione e attrezzature specializzate rispetto al lotto formato44. Al fine di garantire l'accessibilità per i nuovi utenti, i protocolli descritti qui messa a fuoco sul formato batch a volumi di reazione di 15 µ l con raccomandazioni specifiche per aumentare il volume di reazione per la scala di millilitro.
I metodi presentati qui abilitare non esperti con competenze di base di laboratorio (ad esempio studenti universitari) per implementare la crescita delle cellule, estrarre preparazione e installazione di reazione formato in batch per un e. coli-CFPS sistema basato. Questo approccio è conveniente rispetto ai kit disponibili in commercio senza sacrificare la facilità di installazione basato su kit di reazione. Inoltre, questo approccio consente alle applicazioni in laboratorio e in campo. Quando si decide di implementare CFPS, nuovi utenti dovrebbero valutare attentamente l'efficacia di sistemi di espressione di proteine convenzionali per investimento per l'avvio, come CFPS non può essere superiore in ogni caso. I metodi CFPS qui descritti consentono all'utente di implementare direttamente una varietà di applicazioni, tra cui la genomica funzionale, test, la produzione di proteine che sono insolubili per espressione in vivo , come pure il campo ad alta velocità applicazioni tra cui biosensori e kit didattici per biologia sintetica. Le applicazioni aggiuntive come ingegneria metabolica, ottimizzazione delle condizioni di espressione della proteina, individuazione della malattia e scalabilità utilizzando metodi CECF o CFCF sono ancora possibili ma possono richiedere l'esperienza con la piattaforma CFPS per ulteriori modifiche della reazione condizioni. I nostri metodi di combinano una crescita in media arricchita e boccette sconcertati, con metodi relativamente rapidi e riproducibili di lisi cellulare mediante sonicazione, seguita da un'installazione di reazione CFPS semplificata che utilizza ottimizzato premiscele45. Mentre i metodi di crescita cellulare sono diventati un po' standardizzati all'interno di questo campo, metodi per lisi cellulare variano ampiamente. Oltre a sonicazione, metodi di lisi comuni includono l'utilizzo di una pressa francese, un omogeneizzatore, battitori della perla, o lisozima e altri disagi fisici e biochimici metodi46,47,48, 49. usando i nostri metodi, circa 2 mL di estratto grezzo delle cellule sono ottenute per 1 L di cellule. Questa quantità di Estratto di cellule in grado di supportare quattro cento 15 µ l CFPS reazioni, ogni produzione ~ 900 µ g/mL di proteina sfGFP reporter dal plasmide modello pJL1-sfGFP. Questo metodo costa $0,021 / µ g di prodotto sfGFP ($.019/microlitro di reazione), escluso il costo della manodopera e le attrezzature (Supplemental figura 1). A partire da zero, questo metodo può essere implementato in 4 giorni da una sola persona e ripetere le reazioni CFPS possono essere completate entro le ore (Figura 1). Inoltre, il protocollo può essere scalato in volume per grandi lotti di preparazione dei reagenti in base alle esigenze dell'utente. D'importanza, il protocollo qui presentato può essere implementato da laboratorio addestrato non-esperti quali studenti universitari, come richiede solo la capacità di laboratorio di base. Le procedure descritte di seguito e il video di accompagnamento sono stati specificamente sviluppati per migliorare l'accessibilità della piattaforma CFPS Escherichia coli per l'ampio utilizzo.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Media preparazione e crescita cellulare
-
1 ° giorno
- Le cellule di e. coli BL21*(DE3) striscia da un glicerolo stock su una piastra di agar LB e incubare per almeno 18 ore a 37 ° C.
- Preparare 50 mL di LB media e autoclave la soluzione su un ciclo liquido per 30 min a 121 ° C. Conservare a temperatura ambiente.
-
2 ° giorno
- Preparare 750 mL di 2 x YTP media e 250 mL di soluzione di 0,4 M D-glucosio come descritto nelle informazioni supplementari.
- Versare 2 media x YTP in una boccetta di sconcertato 2,5 L sterilizzato nell'autoclave e la soluzione di D-glucosio in una bottiglia di vetro sterilizzati nell'autoclave 500 mL. Autoclave entrambe le soluzioni su un ciclo liquido per 30 min a 121 ° C.
- Verificare che entrambe le soluzioni sterili sono memorizzate a 37 ° C se la crescita cellulare viene eseguita il giorno successivo, per elevare i tassi di crescita al momento dell'inoculazione. Non combinare soluzioni fino a inoculazione.
Nota: Soluzioni possono essere memorizzati a 4 ° C per 1-2 d se necessario, anche se i media x YTP 2 è molto incline alla contaminazione. - Avviare una coltura durante la notte di BL21 (DE3) inoculando 50 mL di LB media con una singola colonia di BL21 (DE3) utilizzando un ciclo sterilizzato e una tecnica sterile per evitare la contaminazione.
- Inserire i 50 mL di LB BL21*(DE3) cultura a 37 ° C 250 giri/min agitazione incubatrice e crescere durante la notte per 15-18 h.
- Preparare e sterilizzare tutti i materiali necessari per i giorni 3 e 4, tra cui: due bottiglie di centrifuga 1 L, 4 x freddo 50 mL provette coniche (pesare e masse record di tre) e molti tubi del microfuge 1,5 mL.
-
3 ° giorno
- Rimuovere la cultura notte 50 mL di BL21*(DE3) in LB dall'incubatrice agitazione e misurare il OD600 su uno spettrofotometro utilizzando un 01:10 diluizione con LB media. Calcolare il volume di notte cultura necessario aggiungere 1 L di media per una partenza OD600 di 0,1 (ad esempio, se un OD600 di un 01:10 diluizione viene letto come 0.4, inoculare 25 mL della non diluito OD600 = 4,0 cultura durante la notte in 1 L di 2 x YTP G).
- Togliere l'incubatore 37 ° C con 50 mL di LB cultura riscaldato 2 x YTP media e soluzioni di D-glucosio. Utilizzando una tecnica sterile, attentamente versare la soluzione di D-glucosio in 2 media x YTP (evitando i lati del matraccio sconcertato).
Nota: Aggiunta di D-glucosio completa la ricetta per 1 L di 2 x YTPG. - Mantenere una tecnica sterile, inoculare il 1 L di 2 x YTPG soluzione con la quantità appropriata della cultura 50ml per iniziare la coltura di 1L a un 0,1 OD600. Posizionare immediatamente la cultura 1L inoculato in un 37 ° C in agitazione incubatore a 200 giri/min.
- Prendere il primo OD600 lettura dopo la prima ora di crescita (fase di ritardo tipico dura 1h). Non diluire la cultura. Continuare a prendere misure600 OD approssimativamente ogni 20-30 min fino a OD600 raggiunge 0,6.
- Dopo aver raggiunto OD600 = 0.6, aggiungere 1 mL di 1 M IPTG (concentrazione finale nella cultura 1 L = 1 mM) alla cultura x YTPG 2.
Nota: Induzione ideale OD600 è 0,6; Tuttavia, una gamma di 0.6-0.8 è accettabile. Induzione di IPTG è per la produzione endogena di T7 RNA polimerasi (T7RNAP). - Dopo l'induzione, misurare il OD600 circa ogni 20-30 min fino a 3.0.
Nota: Raffreddare la centrifuga a 4 ° C durante questo tempo. Preparare il freddo S30 buffer come dettagliato nelle informazioni supplementari. Se il buffer di S30 è preparato in anticipo, è necessario garantire che il DTT non viene aggiunto fino al giorno di utilizzo. - Una volta che il OD600 raggiunge 3.0 (Figura 2A), versare la cultura in una bottiglia fredda di centrifuga 1 L in un bagno di acqua e ghiaccio. Preparare una bottiglia piena d'acqua centrifuga di 1L di uguale peso per essere utilizzato come un equilibrio nella centrifuga.
Nota: I valori di assorbanza variano da strumento a strumento. Mentre il OD600 del raccolto di BL21 (DE3) non è una variabile sensibile, è consigliabile che l'utente valutare e ottimizzare questa variabile come una misura di risoluzione dei problemi. Gli spettrofotometri più grandi possono causare letture di600 OD relativamente più basse rispetto al più piccoli basati su cuvetta spettrofotometri. - Centrifugare le bottiglie di 1 L per 10 min a 5.000 x g e 10 ° C per agglomerare le cellule.
- Versare lentamente il surnatante e smaltirla secondo le procedure dei rifiuti biologici dell'istituzione. Mettere la pallina sul ghiaccio.
- Usando una spatola sterile, raschiare il pellet cellulare dalla bottiglia centrifuga e trasferirlo in una provetta conica da freddo 50 mL.
- Aggiungere 30 mL di tampone di S30 refrigerato a tubo conico e risospendere il pellet cellulare nel Vortex con raffiche brevi (20-30 s) e periodi di riposo (1 min) su ghiaccio fino a quando completamente risospese con senza blocchi.
- Una volta che il pellet è risospeso completamente, utilizzare un'altra provetta conica da 50 mL con acqua come un equilibrio e centrifugare per 10 min a 5000 x g e 10 ° C (pre-raffreddata a 4 ° C).
Nota: Questo completa il 1st di 3 lavaggi necessari durante la raccolta delle cellule. - Riversare il surnatante e smaltirla secondo le procedure dei rifiuti biologici dell'istituzione. Risospendere il pellet con 20-25 mL di freddo S30 tampone e centrifugare per 10 min a 5000 x g e 10 ° C (pre-raffreddata a 4 ° C).
Nota: Questo completa la 2nd di 3 lavaggi. - Ancora una volta, di riversare il surnatante e smaltirla secondo le procedure dei rifiuti biologici dell'istituzione. Aggiungere esattamente 30 mL di tampone di S30 e vortice nuovamente per risospendere il pellet.
- Utilizzando le 3 provette coniche da 50 mL pre-pesati, freddo e un riempitore di pipette sierologiche con una pipetta sterile, aliquota 10 mL di miscela di sedimento pellet/S30 buffer in ciascuna delle 3 provette coniche.
Nota: Dividere le celle in 3 tubi non è necessaria, ma questo passaggio si traduce in piccole palline delle cellule (~ 1 g) per una maggiore comodità alle fasi successive. - Tutte le provette da centrifuga, utilizzando appropriati Saldi come necessario, per 10 min a 5000 x g e 10 ° C (pre-raffreddata a 4 ° C).
Nota: Questo completa la fase di lavaggio finale. - Riversare il surnatante e smaltirla secondo le procedure dei rifiuti biologici dell'istituzione. Rimuovere il tampone in eccesso S30 pulendo accuratamente la parte interna del tubo conico e coperchio con un panno pulito; evitare di toccare il pellet.
- Ripesare i tubi su una bilancia analitica e registrare il peso finale della pallina su ogni tubo.
Nota: Il protocollo può essere sospesa a questo punto. Il pellet può essere flash congelati in azoto liquido e conservato a-80 ° C per fino a un anno fino a quando necessario per la preparazione di estratto.
2. greggio Cell Extract preparazione - giorno 4
- Per la preparazione di estratto, mantenere le cellule fredda su ghiaccio durante ogni passaggio. Aggiungere 1 mL di tampone di S30 refrigerato per 1 g di massa cellulare del pellet. Garantire che ditiotreitolo (DTT) è stato completato il buffer S30 per una concentrazione finale di 2 mM.
Nota: Raffreddare il microcentrifuge a 4 ° C durante questo tempo. - Risospendere il pellet cellulare nel Vortex con raffiche brevi (20-30 s) e periodi di riposo (1 min) su ghiaccio fino a quando completamente in sospensione. Se risospensione è difficile, è possibile lasciare il pellet in ghiaccio per 30 minuti scongelare.
- Trasferimento 1,4 mL di sedimento centrifuga cellule in una provetta da 1,5 mL microfuge.
- Collocare una provetta da 1,5 mL contenente 1,4 mL di sedimento centrifuga cellule in un bagno di ghiaccio d'acqua in un becher. Sonicare per 45 s su seguita da 59 s fuori per 3 cicli totali, con ampiezza fissato al 50%. Chiudere e invertire i tubi a mescolare delicatamente i periodi di fermo. In totale, è necessario consegnare 800-900 J di energia in ogni provetta di microfuge 1,5 mL contenente 1,4 mL di sedimento centrifuga cellule (Figura 3A e 3B).
Nota: Questo passaggio è sensibile al tipo sonicatore e modello utilizzato e deve essere ottimizzato se l'apparecchiatura è diverso da quelli elencati per questa procedura. Due approcci complementari possono essere utilizzati per la quantità di estratto preparato durante questo passaggio la scalabilità: 1) più provette da 1,5 mL microfuge possono essere sonicate in parallelo, e/o 2) più grandi volumi possono essere sonicati in provette coniche (fino a 15 mL di risospensione delle cellule per tubo) , la quantità di energia fornita di scala come descritto in precedenza 29,45. - Subito dopo sonicazione è completa, aggiungere 4,5 µ l di 1 M DTT (completando un'ulteriore 2 mM DTT) nell'1,4 mL di lisato e mescolare capovolgendo. Posizionare il tubo sul ghiaccio. Ripetere i passaggi da 2.4 e 2.5 per qualsiasi altre provette di sedimento cellule prima di procedere alla centrifugazione.
- Microcentrifuga campioni a 18.000 x g e a 4 ° C per 10 min (Figura 3).
- Dispensare il surnatante in una nuova provetta microfuge da 1,5 mL. Non disturbare il pellet; si consiglia di lasciare alcuni surnatante per mantenere la purezza di to perturbare il pellet negli sforzi per massimizzare il rendimento.
- Incubare il surnatante dal passaggio precedente a 250 giri/min e 37 ° C per 60 min registrando i tubi per la piattaforma d'agitazione dell'incubatore (questa è la reazione di deflusso).
- Microcentrifuga campioni a 10.000 x g e a 4 ° C per 10 min.
- Rimuovere il sopranatante senza disturbare il pellet e trasferirlo in un nuovo tubo. Creare molti aliquote di 100 µ l di estratto per deposito.
Nota: Il protocollo può essere messo in pausa qui, e l'estratto può essere flash congelati in azoto liquido e conservato a-80 ° C per fino a un anno fino a quando necessario per CFPS reazioni. Almeno 5 cicli di gelo-disgelo possono essere subiti senza detrimento per estrarre la produttività (Figura 4).
3. cell-Free proteina sintesi Batch formato reazioni
- Disgelo soluzioni un e B, DNA modello, BL21 (DE3) Estratto (se congelati), T7RNAP e un'aliquota di acqua di grado molecolare.
Nota: Modello di reazione CFPS può essere trovato nelle Informazioni supplementari. Ricette di soluzioni A e B sono fornite le Informazioni supplementari e corrispondono a concentrazioni specifiche per numerosi reagenti sostenere il sistema di energia PANOx-SP basata per CFPS. Il ruolo di ciascun reagente e variazione accettabile in queste concentrazioni di reagente che può supportare CFPS sono stati determinati50. Un protocollo di purificazione di T7RNAP si trovano le informazioni supplementari51. T7RNAP supplementare può aumentare i rendimenti volumetrici ma non è necessario se T7RNAP è indotta durante la crescita delle cellule. Modello di DNA del plasmide (pJL1-sfGFP) può essere preparato utilizzando un kit di maxiprep con due lavaggi usando il tampone di lavaggio in kit, seguito da una post-elaborazione del DNA-pulitura utilizzando un kit di purificazione di PCR (Figura 2B). Modelli di DNA lineare utilizzabile anche nelle reazioni CFPS. - Etichettare la quantità necessaria di microfuge tubi necessario per CFPS reazioni.
Nota: Reazioni possono essere eseguite in vari formati di nave, ma una nave più piccola può ridurre i rendimenti volumetrici della proteina (Figura 2). La scalabilità di una reazione nella stessa nave di dimensioni può anche ridurre rendimenti volumetrici, in funzione della diminuzione lo scambio di ossigeno, a causa di una diminuzione della superficie in rapporto al volume. Quando si aumenta il volume di reazione sopra 100 μL, si consiglia di utilizzare il fondo piatto piatti ben 31,37,52. - µ L di soluzione A 2,2, 2,1 µ l di soluzione B, 5 µ l di BL21*(DE3) Estratto, 0,24 μg di T7RNAP (16 concentrazione finale μg/mL), 0,24 ng della mascherina del DNA (concentrazione finale di 16 ng/mL) e acqua per portare il volume finale a 15 µ l.
Nota: Vortex soluzioni A e B frequentemente durante l'installazione di reazione per evitare la sedimentazione dei componenti e garantire che ogni reazione riceve un'aliquota omogenea di ogni soluzione. Evitare nel Vortex l'estratto, invece capovolgere la provetta per mescolare. - Dopo che tutti i reagenti sono stati aggiunti alla reazione, mescolare ogni tubo pipettando su e giù o delicatamente nel Vortex, garantendo nel contempo che la miscela di reazione finale è combinata in una singola perlina 15 µ l nella parte inferiore del tubo microfuge 1,5 mL.
- Posizionare ogni reazione in incubatore 37 ° C senza agitare per 4 h, o a 30 ° C durante la notte.
Nota: Reazioni di successo possono essere qualitativamente valutate visivamente basata sul colore verde del prodotto sfGFP all'interno della miscela di reazione CFPS (Figura 3D). Espressione della proteina di interesse può essere confermata anche da SDS-PAGE (Supplemental figura 2).
4. quantificazione della proteina Reporter, [sfGFP]
- Caricare 48 µ l di 0,05 M HEPES, pH 8, in ogni bene necessario per la quantificazione (di solito eseguita in triplice copia per ogni provetta di reazione).
- Rimuovere le reazioni dall'incubatore. Pipettare su e giù per mescolare ogni reazione, quindi trasferire 2 µ l di reazione nel 48 µ l di 0,05 M HEPES, pH 8. Pipetta su e giù di nuovo nel pozzo per mescolare.
- Una volta che tutte le reazioni sono caricate e mescolate, inserire la piastra ben 96 il fluorimetro e misurare la fluorescenza di endpoint sfGFP.
Nota: Lunghezze d'onda di eccitazione e di emissione per la quantificazione di fluorescenza di sfGFP sono 485 nm e 510 nm, rispettivamente. - Utilizzando una curva standard generata in precedenza, determinare il [sfGFP] dalle letture fluorescenza ottenuti.
Nota: Vengono fornite istruzioni per la generazione di una curva standard di concentrazione sfGFP contro l'intensità di fluorescenza in informazioni supplementari (complementare figura 3). Gli utenti dovranno stabilire una curva standard per il loro strumento, poiché possono variare la sensibilità dello strumento.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Abbiamo presentato una base di sonicazione Escherichia coli Estratto protocollo di preparazione che può essere completato in un arco di quattro giorni, con Figura 1 dimostrando la ripartizione procedurale sopra ogni giorno. C'è malleabilità ai passaggi che possono essere completati in ogni giorno con vari punti di sospensione dell'esecuzione, ma abbiamo trovato questo flusso di lavoro per essere il più efficace eseguire. Inoltre, sia il pellet cellulare (passo 1.3.18) ed estratto completamente pronto (punto 2.10) sono stabili a-80 ° C per almeno un anno, consentendo all'utente di creare più grandi scorte di ciascuno per salvare per l'utilizzo in un tempo successivo17. Non solo è l'Estratto stabile per lunghi periodi di tempo, ma l'estratto può anche subire freeze almeno cinque cicli di scongelamento senza una significativa perdita di produttività (Figura 4). Questo permette maggiori aliquote di estratto deve essere conservato per molteplici usi, se lo spazio di archiviazione del congelatore è limitato. Tuttavia, si consiglia di diverse aliquote più piccole (~ 100 μL) di estratto quando possibile.
Con ogni nuova preparazione di estratto, si consiglia che l'utente esegue una titolazione di magnesio al fine di determinare la quantità ottimale di magnesio per quella partita di estratto. Gli utenti possono quantificare lotto variabilità nella concentrazione nella proteina totale dell'estratto cellulare di analisi di Bradford. Per alte prestazioni estratti, vediamo in genere le concentrazioni di proteina totale di 30-50 mg/mL, e all'interno di questa gamma non c'è correlazione evidente fra le concentrazioni della proteina totale e prestazioni estratto cellulare. È pertanto consigliabile che gli utenti ottimizzare le concentrazioni di magnesio di conseguenza per garantire che proteine e acidi nucleici funzionalità sono massimizzati per ogni batch di estratto. I livelli del magnesio sono importanti per la corretta replica del DNA, trascrizione e traduzione, ma livelli eccessivi possono essere dannosi per questi processi53. Per dimostrare questa dipendenza, ci hanno eseguito una co-titolazione di magnesio ed estrarre il volume per determinare la combinazione ottimale che riduce al minimo la quantità di Estratto necessario, pur mantenendo una reazione produttiva (Figura 5). Da questo esperimento, si consiglia di utilizzare 5 µ l di estratto e mM 10 Mg2 + per estratto con un contenuto di proteina totale di 30 mg/mL, al fine di ottenere oltre 1.000 μg/mL di sfGFP.
La nostra esperienza con CFPS ha anche permesso di determinare i passaggi all'interno del protocollo che può essere variato senza pregiudicare la produttività complessiva del sistema e altri che sono parte integrante di un sistema a elevate prestazioni CFPS. In particolare, il finale OD600 di colture di cellule non influisce in modo significativo l'output finale della reazione CFPS, e cellule tenerne possono essere raccolto ovunque da 2,7-4,0 OD600. Questo rappresenta la prima fase esponenziale di crescita dove ribosoma concentrazione per la cellula è la più alta e il macchinario traslazionale è il più attivo per sostenere la rapida crescita. Questa osservazione permette agli utenti flessibilità ottimizzare le proprie procedure. Si consiglia la raccolta a circa 3,0 OD600 al fine di catturare le cellule a un OD600 più vicino a 3.3 raccogliendo il tempo è completa (Figura 2A). Variabili che hanno un impatto CFPS rendimenti includono modello DNA qualità, dimensioni del vaso di reazione e le relative quantità di celle agli ioni di estratto e di magnesio presenti nella reazione. Abbiamo trovato qualità del DNA ad avere notevole variazione lotto. Per risolvere questo problema, si consiglia che gli utenti purificano il DNA tramite una preparazione midi o maxi, seguita da un ulteriore passaggio di pulitura di DNA sia sulla colonna di purificazione di DNA utilizzati nella maxiprep, o post-purificazione utilizzando un kit di pulizia aggiuntivo di DNA. Questo migliora la riproducibilità in qualità di DNA per reazioni CFPS e risultati nella produzione di proteine più robusta (Figura 2B). Il recipiente di reazione impatti anche rendimenti volumetrici, tale che la produzione di proteine delle configurazioni di reazione identici in diversi volumi di nave può differire fino al 40%. È stato teorizzato che la spinta nella resa volumetrica osservata in vasi più grandi è a causa di una superficie maggiore della miscela di reazione, consentendo per un miglior scambio di ossigeno (Figura 2), e altri hanno ulteriormente rafforzato rendimenti volumetrici eseguendo Reazioni di CFPS in grandi piatti di fondo piatto, che si consiglia per le reazioni sopra 100 µ l17,31,37,52.

Figura 1 : Timeline di sviluppo della cultura, produzione di estratti della cellula, la configurazione e la quantificazione delle reazioni CFPS. L'utente può implementare la piattaforma CFPS per le loro applicazioni di ricerca attraverso questo flusso di lavoro di quattro giorni. Preparazione dei reagenti rappresenta il primario tempo e costo di investimento per il primo turno di questo esperimento e diminuisce sostanzialmente dopo sono costituite scorte di reagenti. Inoltre, palline delle cellule ed estratto preparato delle cellule può essere conservati per più di un anno a-80 ° C, permettendo all'utente di iniziare la sequenza temporale a vari passi per risultati più rapidi. L'utente può inoltre sospendere a vari passi per modificare la sequenza temporale del flusso di lavoro. Clicca qui per visualizzare una versione più grande di questa figura.

Figura 2 : Condizioni modificabile per CFPS e gli effetti sui rendimenti volumetrici reazione. A. estrarre il confronto della produttività basato su raccolta delle cellule BL21 (DE3) alle varie letture di600 OD. Basato su questo terreno, si consiglia di raccolta presso un OD600 di 3.3 per produrre almeno 1000 µ g/mL di proteina bersaglio. Le reazioni sono state eseguite a una scala di 15 μL in provette da 1,5 mL microfuge. B. confronto dei due protocolli del DNA maxiprep lavare con e senza post-purificazione DNA-Ripuliture. pJL1-sfGFP plasmidi hanno subito una maxiprep con uno o due lavaggi, seguite da una pulitura post-purificazione da kit di purificazione di PCR. Per raggiungere ~ 900 µ g/mL di espressione della proteina, suggeriamo di eseguire una pulizia del DNA di post-purificazione indipendentemente dal numero di lavaggi maxiprep. Le reazioni sono state eseguite a una scala di 15 μL in provette da 1,5 mL microfuge. C. 15 µ l CFPS reazioni condotte in vari vasi che vanno da 2 mL a tubi del microfuge 0,6 mL. "Neg" rappresenta un controllo negativo in cui è stato aggiunto alcun modello di DNA per la reazione. Tutte le barre di errore rappresentano 1 deviazione standard di tre reazioni indipendenti per ogni condizione, ognuno dei quali è stato quantificato in triplice copia. Clicca qui per visualizzare una versione più grande di questa figura.

Figura 3 : Chiave procedurali configurazioni e risultati per la creazione di produttiva estraggono. R. una corretta impostazione del bagno di acqua ghiacciata di sonicazione per garantire il raffreddamento del campione, mentre il calore è generato durante la sonicazione. B. 1,5 mL microfuge provetta contenente risospese cella pellet pre (a sinistra) e post (a destra) sonicazione. Il lysate risultante dovrebbe visualizzare una tonalità più scura rispetto al pellet sedimento cellulare. C. la corretta separazione del surnatante e pellet di cella lysata dopo 18.000 x centrifugazione g. D. reazioni CFPS dopo 4 h di incubazione a 37 ° C. tubo da 1,5 mL microfuge sulla destra (reazione di successo) Mostra visibile fluorescenza della proteina reporter sfGFP ~ 900 μg/mL. La provetta di controllo negativo sulla sinistra, manca il DNA del modello e simulando una reazione infruttuosa, Visualizza una soluzione limpida senza fluorescenza. Clicca qui per visualizzare una versione più grande di questa figura.

Figura 4 : Estratto di cambiamento nell'espressione della proteina oltre 5 cicli di gelo-disgelo per CFPS. Estratto preparato dalla stessa crescita ha subito cinque congelare cicli di scongelamento tramite azoto liquido congelamento seguita dallo scioglimento sul ghiaccio flash. Nessun cambiamento significativo nella produttività estratto per esprimere sfGFP sono stato veduto sopra i cinque cicli di gelo-disgelo. Le reazioni sono state eseguite a una scala di 15 μL in provette da 1,5 mL microfuge. "Neg" rappresenta un controllo negativo in cui è stato aggiunto alcun modello di DNA per la reazione. Tutte le barre di errore rappresentano 1 deviazione standard di tre reazioni indipendenti per ogni condizione, ognuno dei quali è stato quantificato in triplice copia. Clicca qui per visualizzare una versione più grande di questa figura.

Figura 5 : CFPS per reazioni con diversi [Mg2+] ed estrarre volumi contro [sfGFP]. [Mg2 +] ha variato da 8 mM a 14 mM con incrementi di 2 mM ed estrarre volumi hanno variati da 3 µ l a 7 µ l con incrementi di 1 µ l. Il colore codice rappresenta [sfGFP] prodotto da alta (rosso) a bassa (viola). Per massimizzare l'efficienza di reagente pur mantenendo la produzione ad alta percentuale proteica, si consiglia di utilizzare 5 µ l di estratto e mM 10 Mg2+ per gli estratti che hanno un contenuto proteico totale di ~ 30 mg/mL, come determinato mediante l'analisi di Bradford. Punti originali per generare il grafico di contorno sono stati basati fuori fluorescenza endpoint di tre reazioni indipendenti per ogni condizione, ognuno dei quali è stata misurata in triplice copia. Le reazioni sono state eseguite a una scala di 15 μL in provette da 1,5 mL microfuge. Clicca qui per visualizzare una versione più grande di questa figura.
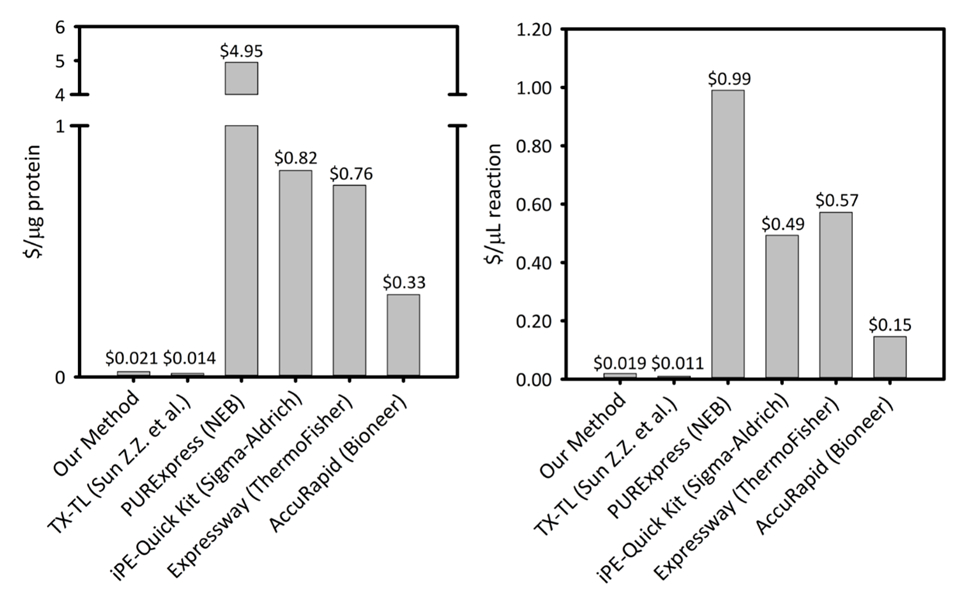
Complementare Figura 1: costi per microgrammo di proteina prodotta e per microliter di reazione su sei piattaforme di sintesi di proteine senza cellula. La nostra piattaforma è paragonato a cinque differenti delle cellule proteina libera sintesi Kit/piattaforme variando la produttività e i prezzi. La nostra piattaforma CFPS sonicazione basata è più conveniente in entrambi $/ µ g di proteina e $/ µ l di reazione rispetto la maggior parte delle imprese kit e fornisce la facilità di un kit per l'installazione di reazione, mentre il rimanente costo paragonabile ad altre piattaforme CFPS accademici. Clicca qui per visualizzare una versione più grande di questa figura.
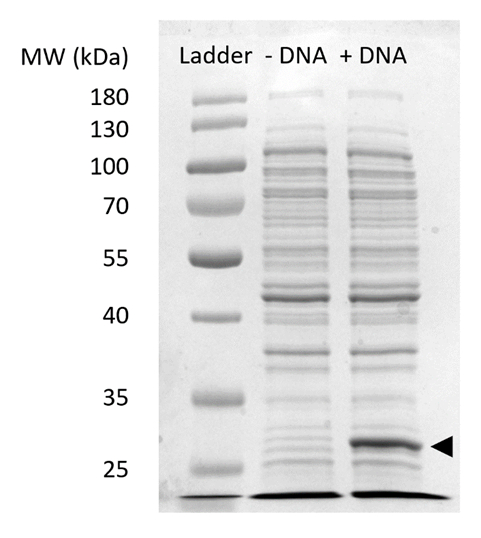
Complementare Figura 2: SDS-PAGE di sfGFP espressione in CFPS. Senza cellula reazioni di sintesi di proteine con (+ DNA) e senza (-DNA) modello di DNA per sfGFP sono stati eseguiti su un 12% gel SDSPAGE per dimostrare l'espressione del sfGFP osservato al 27 kDa (freccia nera). Sono state usate tecniche tradizionali di SDS-PAGE. Ogni campione caricato sul gel incluso 18 µ g di proteina totale basato sulla quantificazione di analisi di Bradford di proteine totali nella cella estrarre. Basato su misure di intensità di fluorescenza e la nostra curva standard, stimiamo che il "+ DNA" lane contiene 0.42 µ g di sfGFP. Al fine di ottenere questi esempi, le reazioni CFPS sono state eseguite in una scala di 15 µ l in tubi del microfuge 1,5 mL producendo rendimenti volumetrici coerenti con Figura 3. Clicca qui per visualizzare una versione più grande di questa figura.
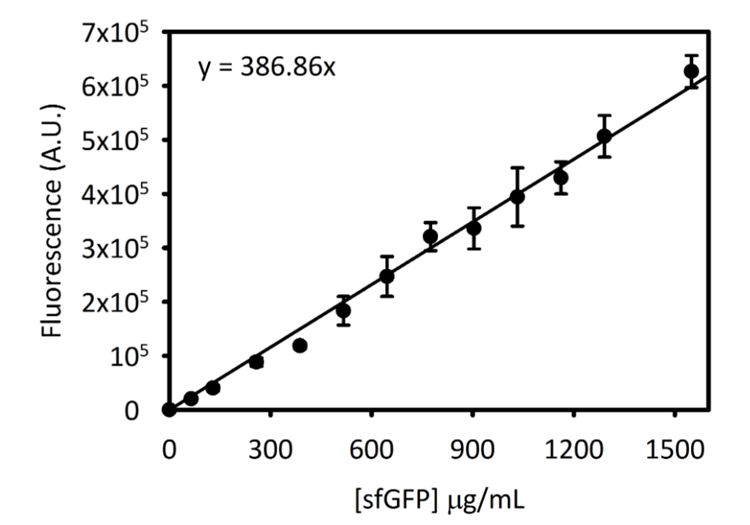
Complementare Figura 3: curva Standard per sfGFP il Cytation 5. Questa curva è stata determinata utilizzando i metodi descritti sopra. Tutti i dati raccolti per questo manoscritto è stato convertito da letture di fluorescenza di endpoint in [sfGFP] in µ g/mL utilizzando questa curva standard. Clicca qui per visualizzare una versione più grande di questa figura.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Sintesi proteica senza cellula è emerso come una tecnologia potente e abilitazione per una varietà di applicazioni che vanno dalla bioproduzione di prototipazione rapida di sistemi biochimici. L'ampiezza delle applicazioni è supportato dalla capacità di monitorare, modificare e aumentare il macchinario cellulare in tempo reale. Nonostante l'impatto di espansione di questa tecnologia di piattaforma, è rimasto ampio adattamento lento a causa di sfumature tecniche nell'implementazione dei metodi. Grazie a questo sforzo, miriamo a fornire chiarezza e semplicità per la costituzione di questa tecnologia nei nuovi laboratori. Verso la fine, il nostro protocollo per un e. coli-piattaforma di sintesi della proteina senza cellula base può essere raggiunto entro un tempo di avvio di quattro-giorni di laboratorio addestrato non-esperti, come ad esempio gli studenti universitari (Figura 1). Inoltre, una volta che uno stock di reagenti ed estratto sono prodotte, successive CFPS batch reazioni possono essere istituite, incubate e quantificate in appena 5 ore. Un singolo, la crescita delle cellule di 1 L può causare abbastanza estratto per reazioni di quattrocento 15 µ l CFPS, mentre preparazioni singolo batch di altri reagenti senza cellula in grado di supportare migliaia di reazioni. Preparazioni di reagente possono essere scalati anche se è necessaria un'azione ancora più grande. Le reazioni di CFPS possono essere configurato in un modo ad alta velocità, utilizzando una piastra a 96 pozzetti o provette PCR per il testing di una varietà di condizioni in parallelo. Rendimenti volumetrici diminuirà quando si utilizzano più piccoli vasi come si vede nella Figura 2. Reazioni CFPS possono anche essere scalate da microliters a decine di millilitri di volume di reazione totale per aumentare il rendimento di proteina totale per una singola condizione. Quando si ridimensiona il volume, la considerazione principale è che i rendimenti volumetrici reazione diminuiscono man mano che diminuisce il rapporto superficie-a-volume della reazione37,52. Per scalabilità mantenendo simili rendimenti volumetrici dell'espressione della proteina, gli utenti dovrebbero dividere il volume di reazione in numerosi vasi di reazione e/o aumentare le dimensioni del vaso. Per reazione scale che vanno da 15 µ l - 100 µ l di volume, numerose reazioni di 15 µ l in parallelo sono raccomandate. Per reazioni superiori a 100 µ l di volume, piastre da 24 pozzetti a fondo piatto sono consigliate e 12 pozzetti sono raccomandati per i volumi di reazione superiore a 600 µ l. Tali associazioni di volumi di reazione e navi forniscono consistenza volumetrica reazione rendimenti su scale up17,31,37,52. Scalabilità verticale di là di questi volumi è possibile utilizzando più pozzetti della piastra in parallelo. Utilizzando questo formato, la reazione può essere scalata a oltre 10 mL di volume totale. Ottimizzare la combinazione di vaso di reazione volume-reazione può supportare applicazioni di bioproduzione senza sacrificare la produttività della reazione.
Quando si esegue questo protocollo, ci sono alcune considerazioni chiave che influenzare i rendimenti volumetrici reazione, nonché indicatori associati a scarso rendimento Estratto. Al fine di garantire adeguata lisi e per evitare la denaturazione del macchinario della trascrizione/traduzione funzionale, è importante mitigare il calore prodotto durante la lisi. Immergere la risospensione delle cellule in un bagno di ghiaccio d'acqua durante la sonicazione per dissipare rapidamente il calore durante la sonicazione (Figura 3A). Un indicatore della lisi cellulare efficace è l'emergere di un aspetto più scuro della cella lysata confrontato ai campioni pre-sonicato (Figura 3B). Per flessibilità all'utente, il sonicatore e la sonda mostrata in Figura 3A è adattabile a una gamma di volumi da 100 µ l a 15 mL di sedimento centrifuga cellule. A tale scopo, l'utente può regolare il numero di Joule consegnato per lisi del volume desiderato delle cellule. Inoltre, grandi volumi di Estratto possono essere preparati attraverso due approcci complementari. Utenti possono sottoporre ad ultrasuoni tubi multipli in parallelo e/o sottoporre ad ultrasuoni più grandi volumi di risospensione delle cellule, scalare la quantità di energia proporzionalmente con il volume come descritto in precedenza29,45. Un altro passo che indica Estratto di qualità è il passo di centrifugazione dopo lisi cellulare. Post lisi cellulare, si consiglia di centrifugazione a 18.000 x g per fornire una chiara divisione tra il surnatante (macchinario di trascrizione/traduzione, frammentazione DNA genomico che non funziona più modello trascrizione/traduzione) e il pellet ( indesiderati componenti cellulari come la membrana cellulare e proteine precipitate) (Figura 3). Abbiamo trovato quel centrifugazione a 18.000 x g migliora la separazione, con conseguente migliore riproducibilità rispetto alla gira a velocità inferiori come 12.000 x g. Per comodità, si consiglia di utilizzare una centrifuga refrigerata da appoggio, in grado di raggiungere un minimo di 12.000 x g. Questo passaggio è anche comunemente eseguita a 30.000 x g, che dovrebbe essere considerata se l'attrezzatura appropriata è disponibile54,55,56,57,58, 59 , 60. Estratto di prestazioni non sono interessata dalla velocità di centrifugazione a questo punto dato che la corretta separazione è raggiunto. Quando si rimuove il surnatante desiderato, è meglio evitare qualsiasi materiali nuvolosi che esistono al confine tra il surnatante e pellet poiché questa contaminazione ridurrà la produttività dell'estratto. Con l'obiettivo per la purezza del surnatante si traduce in estratti più produttivi e vale la pena della riduzione della quantità di estratto ottenuto per i nuovi utenti.
È importante notare che mentre i metodi che abbiamo presentato sono riproducibili e possono essere eseguiti dagli scienziati con minimo grado di esperienza, ci può essere lotto e variazione di reazione a reazione. Ciò può essere attribuito alla variazione della composizione di proteomica dell' lisato post-sonicazione61. La variabilità di lotto che abbiamo osservato è generalmente diminuita dopo il completamento con T7RNAP e ottimizzazione delle concentrazioni di magnesio. Aggiunta esogena di T7RNAP è comune tra reazioni CFPS per sostenere l'espressione della proteina ottimale, e troviamo che avendo due fonti di T7RNAP - espressione endogena in BL21*(DE3) e il T7RNAP supplementare ad una concentrazione finale di 16 µ g/mL - migliora lotto riproducibilità per nuovi utenti45,46. Con esperienza, gli utenti possono modificare i loro esperimenti per utilizzare solo una singola fonte di T7RNAP se lo si desidera. Quantificazione del contenuto proteico totale di un nuovo lotto di estratto e opportuna regolazione della concentrazione di Mg2 + può anche aiutare a diminuire-lotto variazione dei rendimenti di espressione volumetrica della proteina. Variazioni nell'espressione della proteina possono anche essere dovuto le differenze nella dimensione e nella struttura della proteina di interesse, l'utilizzo di codone del gene e il suo sito di legame ribosoma corrispondente del gene di interesse, come pure il tipo di vettore di espressione utilizzato62 ,63. Per queste ragioni, alcune proteine non possono esprimere così come il modello della proteina sfGFP, con conseguente diminuzione della resa volumetriche da reazioni CFPS.
Le limitazioni della tecnica CFPS presentata comprendono che non può essere direttamente adatto a tutte le applicazioni del cellulare-free, come ingegneria metabolica e ottimizzazione delle condizioni di espressione, senza ulteriori modifiche ai protocolli. Tuttavia, riteniamo che questo protocollo fornirà una base per stabilire la piattaforma CFPS in nuovi laboratori e fornire non esperti con la possibilità di implementare introduttive senza cellula reazioni nei loro laboratori. Dopo l'implementazione iniziale, i ricercatori possono sperimentare con la piattaforma per fare le proprie modifiche per applicazioni più specifiche basate su altra letteratura nel campo.
La proteina CFPS piattaforma costi $0,021 / µ g (escluso il costo della manodopera e attrezzature), rendendo il nostro sistema a prezzi competitivi con kit commerciali senza compromettere la facilità d'installazione di reazione. Le valutazioni dei costi comparativi per µ l di reazione mostrano un andamento simile (Supplemental Figura 1). Si stimano costi di avvio per essere ~ $4.500 per tutti i reagenti e un ulteriore $3.200 per attrezzature specializzate, come ad esempio un sonicatore. Ore di persona per completare questa procedura sono stimati essere ~ 26 h per tutti reagente preparazione da terra fino. Tuttavia, una volta che sono state preparate grandi scorte di reagenti, richieste sul lavoro diminuiscono sostanzialmente. Inoltre, come si guadagna esperienza con la piattaforma, si consiglia di scalabilità verticale la dimensione della crescita cellulare, estrarre preparazione e preparazione dei reagenti per massimizzare l'efficienza del tempo. Visti i costi di avvio, si consiglia la piattaforma CFPS per applicazioni in biologia sintetica, gli sforzi di alto-rendimento, e condizioni di espressione di proteine che sono incompatibili con le piattaforme di espressione tradizionale proteina causa in conflitto con la cella vincoli di biochimica e di vitalità. In questi casi speciali dove la tecnica desiderata è attivata dalla piattaforma CFPS, è giustificato il costo maggiore del CFPS sopra espressione in vivo .
Costante sviluppo della piattaforma CFPS rischia di fornire il più ampio programma di utilità per gli sforzi di biotecnologia come l'ingegneria metabolica delle vie enzimatiche, produzione e caratterizzazione di proteine tradizionalmente insolubile, non standard dell'amminoacido incorporazione e l'espressione della proteina innaturale, manufacturing della medicina stratificata ed espansione oltre il laboratorio in aula per STEM educazione64,65,66. Questi sforzi saranno ulteriormente supportati dagli sforzi in corso per la caratterizzazione dettagliata della piattaforma CFPS. Una migliore comprensione della composizione dell'estratto cellulare porterà al perfezionamento continuo verso una migliore reazione rendimenti e flessibilità nelle condizioni di reazione61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Gli autori dichiarano di avere nessun concorrenti interessi finanziari o altri conflitti di interesse.
Acknowledgments
Gli autori si desidera ringraziare Dr. Jennifer VanderKelen, Andrea Laubscher e Tony Turretto per il supporto tecnico, Wesley Kao, Layne Williams e Christopher Hight per utili discussioni. Autori riconoscono anche finanziamento da Bill e Linda Frost Fund, centro per le applicazioni in Chevron biotecnologia applicata ricerca Endowment Grant di biotecnologia, Cal Poly ricerca, da studioso e Creative attività Grant Program (RSCA 2017), e la National Science Foundation (NSF-1708919). MZL riconosce la California State University Graduate Grant. MCJ riconosce l'esercito ricerca ufficio W911NF-16-1-0372, National Science Foundation concede MCB-1413563 e MCB-1716766, l'Air Force Research laboratorio centro di eccellenza Grant FA8650-15-2-5518, la concessione di agenzia di difesa minaccia riduzione HDTRA1-15-10052/P00001, il David e Lucile Packard Foundation, il programma di Camille Dreyfus Teacher-Scholar, il dipartimento di energia BER Grant DE-SC0018249, il programma di scienza umana frontiere (RGP0015/2017), la concessione ETOP DOE Joint Genome Institute, e il Consorzio biomedica di Chicago con il sostegno dei fondi di Searle presso la comunità di Chicago Trust per il supporto.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}