

Summary
このプロトコルの詳細手順、コスト、および大腸菌を生成に必要な装置・細胞抽出液をベースし、4 日間以内の in vitroタンパク質合成反応を実装します。広範なアプリケーションは、このプラットフォームの柔軟な性質を活用するには、と最適化することができます反応条件について検討します。
Abstract
最後の 50 年にわたって無細胞タンパク質合成 (当たり) は、試験管内のセルの転写と翻訳の能力を活用する強力な技術として浮上しています。迅速なプロトタイプで当たりを伝統的に挑戦的な蛋白質の製造の新興アプリケーションとしてアプリケーションを基礎されているセルの実行可能性を維持する必要があるし、細胞の障壁を排除することによって、代謝工学、ゲノム機能学。私たちの方法、エシェリヒア属大腸菌の実装-ベース CFPS プラットフォームは、これらのアプリケーションの多くにアクセスする新しいユーザーを許可します。ここでは、我々 は豊かなメディア、困惑してフラスコおよび可変超音波ベースのセル換散の再現可能なメソッドを使用して抽出を準備する方法を説明します。この抽出物は適切な試薬株式を事前に準備されていることを考える 900 μ g/mL 以上のデータ分析、実験のセットアップからちょうど 5 時間でスーパー フォルダー緑色蛍光蛋白質 (sfGFP) を作り出すことができる蛋白質の表現を使用できます。試薬を得る推定のスタートアップのコストは $4,500 を何千もの $0.021 産生される蛋白の μ g あたりの $0.019 μ L 反応のあたりの推定コストで反応を維持します。さらに、蛋白質の表現方法は、コストのほんの一部で、試薬中古ミックスの最適化のための市販のシステムに見られる反応セットアップの容易さをミラーします。CFPS 広範なアプリケーション用の柔軟な性質を活用するユーザーを有効にするためにさまざまな調整し、使用可能なリソースおよび蛋白質の表現の結果に応じて最適化できるプラットフォームの側面を位置付けています。望まれています。
Introduction
無細胞タンパク質合成 (当たり) は、タンパク質の生産、機能ゲノム学、代謝工学と最後の 50 年間1,2以内に詳細のための新しい機会の数をアンロックしている技術として浮上しています。株当たり標準生体内タンパク質式プラットフォームと比較して、3 つの利点を提供します: 1) 携帯無料自然のプラットフォームの潜在的毒性や細胞の3,4 外国語になるタンパク質の製造を可能に ,5,6;2) ゲノム DNA の不活性化および興味の遺伝子を符号化する DNA テンプレートの導入が関心のタンパク質の生産への反応の中で全身のエネルギーのすべてをチャネルします。・ 3) プラットフォームのオープンな性質により、ユーザーが変更したり、反応条件とリアルタイム7,8の組成を監視します。この反応への直接アクセスをサポートしている拡張化学や新規タンパク質の生産と代謝過程2,9,のチューニングのための酸化還元状態の生物学的システムの増強10. 直接のアクセスは、ユーザーがより迅速な設計、ビルド、テスト サイクル11のシングル ポット システムで活性測定法と当たりの反応を組み合わせることもできます。紙ベースのデバイスのさらなるサポート ハイスループット探索努力とラピッドプロトタイピング12,13,14,15 または少量の水滴で当たり反応を実行する能力 ,16。これらの利点は、システムのプラグ アンド プレイの性質の結果として当たりが一意に溶けてエクスプレス体内しにくいタンパク質の生産などのバイオ テクノロジーのアプリケーションのさまざまなを有効に17、 18,19,20, 疾患21,22,23, 需要製造18,24 の検出、25,26,27、および教育28,29、柔軟性と携帯無料プラットフォームのユーティリティを表示します。
CFPS システムは様々 な原核生物と真核生物の細胞から粗野な lysates から生成できます。これにより、対象のアプリケーションによって一長一短があるそれぞれの選択のシステムで多様なオプション。CFPS システムは準備時間、コスト、および生産性にも大きく異なります。最も一般的エキス、小麦胚芽、ウサギ網状赤血球、昆虫細胞、大腸菌細胞、後者はタンパク質30 の最高の容積の生産に最も費用対効果されてから生成されるセルを活用.他の当たりのシステムはその生得的な翻訳後修飾機械に有利であるが、新興の大腸菌を使用してアプリケーション-ベース機械が生成 site-specifically リン酸化によってギャップを埋めることができると需要31,32,33,34,35の糖蛋白質。
株当たりの反応は、いずれかのバッチ、連続交換携帯無料 (展) または連続的なフロー セル無料 (CFCF) 形式で実行できます。バッチ形式は、その反応の有効期間は反応物と反応の抑制の副産物の蓄積の減少量により制限クローズド システムです。展と CFCF 方法、反応の寿命を延ばすし、それによってバッチ反応と比較して増加した体積蛋白質収量の結果します。これは、新しい反応の反応2コース全体で供給している反応容器から削除するタンパク質合成の副産物を許可するで。CFCF の場合は、反応室、展、半透膜36,37から成る反応室のままに興味の蛋白質の中から興味の蛋白質を削除できます。これらのメソッドは特に関心38,39,40,41,42、エクスプレス難しいタンパク質の低体積収率を克服するための貴重です 43。展と CFCF アプローチの実装の課題は、1) 一方、彼らは転写と翻訳を担当バイオ機械のより効率的な使用の結果、特に大きい増加の全体的なコストと 2 試薬量が必要)複雑な反応セットアップとバッチ形式44と比較して特殊な装置が必要です。新しいユーザーのためのアクセシビリティを確保するためにプロトコルに記載フォーカス ミリリットル スケールの反作用ボリュームを増加させるための特定の推奨事項 15 μ L の反応ボリュームでバッチ形式で。
記載方法は、細胞増殖を実装、エキスの準備、および、エシェリヒア属大腸菌の形式反応セットアップをバッチ (学生) など基本的な実験技術と非専門家を有効にする-ベースの当たりシステム。この方法は、反応のキット ベース セットアップの容易さを損なうことがなく市販のキットと比較して費用対効果です。さらに、このアプローチにより、アプリケーション実験室およびフィールド。当たりを実装するかを決める、新しいユーザーする必要があります当たりはすべてのケースで優れてはないかもしれないスタートアップ投資の従来の蛋白質の表現システムの有効性を評価徹底的に。ここで説明 CFPS 方法機能ゲノミクス、高スループット テスト、フィールドと同様に、体内式の扱いやすいタンパク質の生産を含む、アプリケーションのさまざまなを直接実装するユーザーを有効にします。バイオ センサー、合成生物学の教育キットを含むアプリケーション。代謝工学などの追加アプリケーション チューニングのタンパク質式条件、病気の検出、およびスケール アップを使用して展または CFCF メソッドはまだ可能ですが、さらに反応の編集 CFPS プラットフォームとの経験があります条件。我々 の方法は、超音波処理、次に最適化されたプレミックス45を利用して簡易 CFPS 反応セットアップを介してセル換散の比較的迅速かつ再現性のある方法と成長リッチなメディアと困惑してフラスコを組み合わせます。細胞の増殖方法はややこのフィールド内で標準化になるが、セル換散方法が異なります。一般的な換散方法のほか超音波、フレンチ プレス、ホモジナイザー、ビーズ ビーターやリゾチーム、他生化学的および物理的な中断方法46,47,48,の利用49。 私たちのメソッドを使用すると、セルの 1 L あたり原油細胞抽出液の約 2 mL が得られます。細胞抽出液のこの量は 400 をサポートできる 15 μ L 当たり反応、各生産 〜 900 μ g/mL テンプレート プラスミドの pJL1 sfGFP から記者 sfGFP 蛋白質の。この方法で費用 $0.021/μ g sfGFP 生産 ($.019/μ L 反応)、労働および機器 (補足図 1) のコストを除きます。スクラッチから始めて、このメソッドは、一人で 4 日間で実装することができ、CFPS 反応時間 (図 1) 以内に完了することができますを繰り返します。さらに、ユーザーのニーズに合わせて試薬調製の大きなバッチ量プロトコルを拡張できます。重要なは、それだけ基本的な実験技術を必要とは、ここで提示されたプロトコルを訓練所の非専門家、学生などによって実装できます。以下に説明する手順とそれに伴うビデオは、特に広範な使用のためのエシェリヒア属大腸菌株当たりプラットフォームのアクセシビリティを改善するために開発されています。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. メディアの準備と細胞増殖
-
日 1
- グリセロールから連勝大腸菌BL21*(DE3) 細胞 LB 寒天培地プレート ストックし、37 ° C で少なくとも 18 時間インキュベート
- 121 ° C で 30 分の液体のサイクルで LB 媒体のオートクレーブ ソリューション 50 mL を準備します。常温で保存します。
-
日 2
- 補足情報の説明に従って 0.4 M D-グルコース溶液 250 mL 2 x YTP メディアの 750 mL を準備します。
- オートクレーブの 2.5 L 困惑してフラスコとオートクレーブ 500 mL ガラス瓶の中に D グルコース溶液に 2 x YTP メディアを注ぐ。オートクレーブ 121 ° C で 30 分の液体のサイクルの両方のソリューション
- 両方の滅菌ソリューションは 37 ° c で保存されます細胞増殖で接種時に成長率を最大化するために、次の日に実行する場合を確認します。接種までのソリューションを結合しません。
注: ソリューションは 2 x YTP メディアが汚染しやすいが、1-2 d 必要な場合のための 4 ° C で保存できます。 - 50 mL LB 媒体の汚染を避けるため滅菌ループと生殖不能の技術を使用して BL21(DE3) の単一コロニーを再接種によって BL21(DE3) の一晩かけて培養を開始します。
- インキュベーターの振動 250 rpm 37 ° C に BL21*(DE3) LB 文化の 50 mL を置き、一晩 15 18 h の成長します。
- 準備および日間、3、4 を含む必要なすべての材料を殺菌: 2 1 L 遠心ボトル、4 x 冷 50 mL の円錐管 (重さと 3 つのレコードの固まり)、多くの 1.5 mL microfuge の管。
-
日 3
- 揺れインキュベーターから LB の BL21*(DE3) 50 mL 一晩文化を削除し、測定分光光度計、1:10 を使用して外径600 LB 媒体で希釈。一晩かけて培養開始外径600 0.1 のためメディアの 1 L に追加する必要の体積を計算 (たとえば、1:10 希釈を 0.4 として読むの外径600原液外径600 25 mL を接種する場合は 2 の 1 L に 4.0 一晩文化を = x YTPG)。
- LB 文化の 50 mL と一緒に 37 ° C の定温器から温めた 2 x YTP メディアと D-グルコース ソリューションを削除します。(困惑してフラスコの側面を回避) 2 x YTP メディアに D-グルコース溶液を注ぐ無菌技術を使用して、慎重に。
注: D-グルコース添加完了 2 の 1 L のレシピ x YTPG。 - 生殖不能の技術を維持し、0.1 外径6001 L 文化を開始する 2 x YTPG 溶液 50 mL 文化の適切な量の 1 L を接種します。すぐに 200 rpm でインキュベーターを揺れ 37 ° C に接種の 1 L 文化を配置します。
- 成長の最初の 1 時間後を読んで初めて OD600を取る (典型的な遅れ位相が 1 h)。文化を薄めないでください。外径600に達する 0.6 まで 20-30 分毎に約外径600測定を服用し続けます。
- 外径600に達する = 0.6、1 M IPTG の 1 mL を追加 (1 L 文化の最終濃度 = 1 mM) 2 x YTPG 文化。
注: 最適な誘導外径600は 0.6;ただし、0.6 から 0.8 の範囲は、許容範囲です。IPTG による誘導は内因性生産の T7 RNA ポリメラーゼ (T7RNAP) のためです。 - 誘導後に、、3.0 に達するまで OD600約 20-30 分毎を測定します。
注: は、この時間の間に 4 ° C に遠心分離機を冷やします。詳細の補足情報として冷たい S30 バッファーを準備します。S30 バッファーを事前に準備する場合は、DTT は利用日まで追加されていないことを確認します。 - 外径600 3.0 (図 2 a) に達すると、氷水浴の冷たい 1 L 遠心ボトルに文化を注ぐ。遠心分離機でバランスとして使用される等しい重量の水で満たされたボトルの 1 L 遠心分離機を準備します。
注: 吸光度値は、計測器-計測器によって異なります。BL21(DE3) の収穫の OD600は敏感な変数が、ユーザーの評価し、トラブルシューティング手段としてこの変数を最適化、それをお勧めします。小さいキュベット ベース分光光度計と比較して比較的低い外径600測定値の大きい分光光度計があります。 - 5,000 × g とペレットのセルに 10 ° C で 10 分の 1 L ボトルを遠心します。
- 上清から処分施設の生物学的廃棄物手順に従ってゆっくりと注ぐ。氷の上には、ペレットを配置します。
- 滅菌へらを使用して、遠心ボトルから細胞ペレットをこすりし、冷たい 50 mL の円錐管にそれを転送します。
- 円錐管に 30 mL の冷たい S30 バッファーを追加し、氷なしの塊と完全に再停止されるまでの短いバースト (20-30 秒) と休憩時間 (1 分) ボルテックスによって細胞ペレットを再懸濁します。
- ペレットが完全に再停止される一度、g、10 ° C (4 ° C に冷却済み) × 5000 で水でバランスと 10 分間遠心する別の 50 mL のコニカル チューブを使用します。
注: これは細胞を収穫するときに必要な 3 洗浄の 1stを完了します。 - 上澄みと機関の生物学的廃棄物手順に従って廃棄を注ぐ。寒さの S30 バッファーと 10 ° C (4 ° C に冷却前) 5000 × g で 10 分間遠心の 20-25 mL でペレットを再懸濁します。
注: これは 3 洗浄の 2ndを完了します。 - もう一度、清と機関の生物学的廃棄物手順に従って廃棄を注ぐ。S30 バッファーとペレットを再懸濁しますして渦の丁度 30 mL を追加します。
- 割り切れる滅菌ピペットで 10 mL の再懸濁のペレット/S30 バッファー混合物の 3 円錐管のそれぞれに 3 あらかじめ重量を量られた、冷たい 50 mL の円錐管と血清ピペット注入口使用します。
注: セルを分割 3 管必須ではありませんが、このステップの結果より小さい細胞ペレット (~ 1 g) 後の手順で利便性の向上のため。 - 5000 × g と 10 ° C (4 ° C に冷却済み) で 10 分間、必要に応じて適切なバランスを使用して、すべてのチューブを遠心します。
注: これは最終的な洗浄ステップを完了します。 - 上澄みと機関の生物学的廃棄物手順に従って廃棄を注ぐ。慎重に円錐管とクリーンな組織; とキャップの内側を拭くことによって余分な S30 バッファーを削除します。ペレットを触れないでください。
- 分析用天秤にチューブを reweigh、各チューブに最終的なペレット重量を記録します。
注: プロトコルをこの時点で一時停止できます。ペレットは、液体窒素で凍結し、エキスの準備のために必要になるまでの年まで-80 ° C で保存されているフラッシュすることができます。
2. 原油細胞エキスの準備 - 4 日目
- エキスの準備のため細胞を維持冷たい氷の上各段階。1 mL の細胞ペレットの質量の 1 g あたりの寒さの S30 バッファーを追加します。2 mM の最終的な集中に S30 バッファーに補われたそのジチオトレイトール (DTT) を確認します。
注: は、この時間の間に 4 ° C に遠心を冷やします。 - 完全に再停止されるまで氷の上の短いバースト (20-30 秒) と休憩時間 (1 分) ボルテックスによって細胞ペレットを再懸濁します。巻き上がりが困難である場合は、解凍する 30 分間氷の上ペレットを残します。
- 1.5 mL および microfuge の管に再懸濁細胞の 1.4 mL を転送します。
- ビーカーの氷水風呂に再懸濁細胞の 1.4 mL を含む 1 つの 1.5 mL チューブを配置します。45 超音波の s に続いて 59 50% で振幅の 3 総サイクル オフ s を設定。閉じ、優しくオフ期間中にミックスするチューブを反転します。合計では、再懸濁細胞 (図 3 a ・ 3 b) の 1.4 mL を含む各 1.5 mL および microfuge の管にエネルギーの 800-900 J を提供します。
注: この手順は超音波発生装置タイプに敏感とモデル使用し機器がこの手順に記載されているよりも異なる場合に最適化する必要があります。この手順で抽出量をスケール アップ 2 つの相補的なアプローチを使用できます: 1) 複数の 1.5 mL microfuge の管は、並列で超音波処理することができますおよび/または 2) より大きなボリュームは、円錐管で超音波処理することができます (最大 15 mL チューブあたり細胞を再懸濁の)、 29,45を前述したエネルギーの供給量をスケーリングします。 - 超音波処理を完了後すぐにライセートの 1.4 mL に 1 M (補足 2 mM DTT) DTT の 4.5 μ L を追加し、ミックスする数回を反転します。氷の上には、チューブを配置します。手順 2.4 および 2.5 を遠心分離に進む前に再懸濁細胞の任意の追加のチューブ。
- 遠心試料 18,000 × g と 4 ° C 10 分 (図 3)。
- 上清を新しい 1.5 mL および microfuge の管にピペットします。ペレット; 操作不可します。収量を最大化する取り組みで、ペレットを混乱させるよりも純度を維持するためにいくつかの上澄みを残すことをお勧めします。
- (これは流出反応) インキュベーターの動揺のプラットホームにチューブをテープで 250 rpm と 37 ° C, 60 分前の手順から上澄みを孵化させなさい。
- 遠心 10,000 × g と 4 ° C 10 分のサンプル。
- ペレットを乱すことがなく上澄みを除去し、新しい管にそれを転送します。ストレージのためのエキスの多くの 100 μ 因数を作成します。
注: プロトコルがここでは、一時停止にできるし、液体窒素で凍結し、株当たり反応のために必要になるまでの年までの-80 ° C で保存抽出物がフラッシュすることができます。抽出 (図 4) の生産性を損なうことがなく、少なくとも 5 凍結融解サイクルを受けてすることができます。
3. 無細胞タンパク質合成バッチ形式の反応
- ソリューション A と B、DNA テンプレート、BL21(DE3) エキス (冷凍) 場合、T7RNAP、および分子グレード水の因数分解しなさい。
注: CFPS 反応テンプレートは、補足情報を見つけることが。ソリューション A と B のレシピは補足情報を提供され、当たりの PANOx SP ベースのエネルギー システムをサポートするための多数の試薬の特定濃度に対応します。各試薬と当たりをサポートすることができますこれらの試薬濃度の許容変動の役割は、決められた50をされています。補足情報51T7RNAP の浄化のプロトコルを見つけることができます。補足 T7RNAP 体積の収量を増やすことができますが、T7RNAP はセル成長の間に誘導される場合は必要ありません。プラスミド DNA のテンプレート (pJL1 sfGFP) は、PCR 精製キット (図 2 b) を使用して処理後の DNA クリーンアップ続いてキットに洗浄バッファーを使用して 2 つの洗浄と maxiprep キットを用いて調製できます。株当たり反応の線形 DNA のテンプレートが使えます。 - 株当たり反応のために必要の microfuge の管の必要な量をラベル付けします。
メモ: 様々 な容器サイズで反応を行うことが、小さい容器の体積蛋白質収量 (図 2) を減らすことができます。同じサイズの容器内で反応をスケーリング場合も少なく体積収量減少体積比表面積の減少により、酸素交換の機能として。100 μ L の上の反作用ボリュームを増加する場合は、底が平らなウェル プレート31,37,52を使用することをお勧めします。 - 溶液の 2.2 μ L を追加、ソリューション B、BL21*(DE3) の 5 μ L の 2.1 μ L エキス、T7RNAP の 0.24 μ g (16 μ g/mL 最終濃度)、0.24 DNA のテンプレート (16 ng/mL 最終濃度)、そして 15 μ L の最終巻をさせる水の ng。
注: 渦解 A と反応セットアップ中によく B 成分の沈降を避けるために、各反応が各ソリューションの均質な因数を受け取ることを確認します。ボルテックス抽出物を避けるために、代わりにミックスするチューブを反転します。 - すべての試薬は反応に追加されている後、最終的な反応混合物が結合されて単一 15 μ L ビーズ 1.5 mL および microfuge の管の下部に確保しながら上下にピペッティングまたは軽くボルテックスによって各チューブをミックスします。
- 4 時間または 30 ° C を一晩振盪せず 37 ° C の定温器にそれぞれの反応を配置します。
注: 正常な反応質的評価できます視覚的に当たり反応混合物 (図 3 D) 内の sfGFP 製品の緑の色に基づいて。興味の蛋白質の表現は、SDS ページ (補足図 2) によっても確認できます。
4. [sfGFP] レポーター蛋白質の定量
- よく定量化 (通常は反応チューブあたり 3 通で実行されます) に必要なそれぞれに 0.05 m HEPES、pH 8、48 μ L をロードします。
- インキュベーターから反応を削除します。ピペットの上下各反応を混在させるし、0.05 m HEPES、pH 8 48 μ L に 2 μ L の反応を転送します。ミックスに井戸に再び上下ピペットで移しなさい。
- すべての反応は読み込まれ、混合、一度、蛍光光度計 96 well プレートに配置し、sfGFP エンドポイント蛍光を測定します。
注: sfGFP 蛍光定量化のための励起と放射の波長は、485 nm、510 nm、それぞれ。 - 以前に生成された標準的なカーブを使用すると、得られる蛍光の測定値から [sfGFP] を決定します。
注: sfGFP 蛍光強度と濃度の標準曲線を生成する手順は、補足情報 (補足図 3) で提供されます。ユーザーは、測定器の感度が異なる場合がありますので、それらの楽器の標準曲線を確立する必要があります。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
超音波処理による大腸菌にエキスを提案した各曜日に手続き型の内訳を示す図 1の 4 日間にわたって、完了できる準備のプロトコル。毎日様々 な一時停止ポイントで完了ことができます手順に展性があるが、このワークフローを実行する最も効果的であることがわかった。さらに、細胞ペレット (ステップ 1.3.18) と完全に準備された抽出 (ステップ 2.10) の両方は、-80 ° C で安定して少なくとも 1 年間、後時17で使用するため保存するそれぞれの大きな株式を作成するユーザーを許可します。長時間にわたり安定した抽出物をただ抽出 (図 4) の生産性が大幅に低下することがなく、少なくとも 5 つの凍結融解を受けることができるも。これは冷凍庫の容量が限られている場合、複数の用途に格納される抽出のより大きい因数にできます。ただし、可能な限り抽出物の複数の小さい因数 (~ 100 μ L) を推奨します。
すべての新しい抽出の準備とユーザーがマグネシウムのバッチ抽出のための最適な量を決定するためにマグネシウム滴定を実行することをお勧めします。ユーザーは、ブラッドフォードの試金によって、細胞抽出液の総蛋白濃度のバッチごとに変動を定量化できます。抽出物、高性能の我々 は通常 30-50 mg/mL、総蛋白濃度を見るし、この範囲内で総蛋白濃度と細胞抽出パフォーマンスの明白な相関関係はありません。したがって、ユーザーがマグネシウム濃度を適宜調整し、抽出の各バッチは、タンパク質と核酸機能も最大化することを確認することをお勧めします。マグネシウム濃度は適切な DNA 複製、転写、翻訳のために重要が、過剰なレベルは53これらのプロセスに支障をきたすことができます。この依存関係を示すためには、マグネシウムの共同滴定を行ったし、生産的な反応 (図 5) を維持しながら、必要なエキスの量を最小にする最適な組み合わせを決定するボリュームを抽出します。この実験から、sfGFP の以上 1,000 μ g/mL を得るために 30 mg/ml、総たんぱく抽出物エキスと 10 mM Mg2 +の 5 μ L を使用をお勧めします。
当たりと私たちの経験は、高パフォーマンス当たりのシステムに不可欠なシステムの全体的な生産性を損なうことがなく変化するプロトコルと他の内にある手順を確認することもできました。特に、細胞収穫の最終的な OD600が当たり反応の最終出力を大幅に影響しないため、2.7 4.0 外径600から細胞がどこでも収穫動けなかったことができます。これはセルあたりリボソーム濃度が最も高い並進機械は急速な成長をサポートする最も活発な成長の早い指数フェーズを表します。この観測では、ユーザー独自の手順を最適化する柔軟性をことができます。約 3.0 外径600時間収穫、OD で600近い 3.3 細胞を捕捉するために収穫が完了 (図 2 a) をお勧めします。株当たり収量に影響を与えるか変数は、テンプレート DNA の品質、反応容器のサイズ、および細胞抽出物とマグネシウム イオン反応の存在の相対的な量に含まれます。我々 は DNA の品質は顕著なバッチごとにバリエーションを持って発見しました。この問題を解決するために、ユーザーが DNA 精製カラムのいずれかは、maxiprep で使用される追加の DNA クリーンアップ手順が続くまたは追加の DNA クリーンアップ キットを使用して後の浄化、midi やマキシ準備を介して DNA を浄化することをお勧めします。これは当たり反応の DNA 品質の再現性とより堅牢なタンパク質の生産 (図 2 b) の結果を向上させます。反応容器は、さまざまな容器の容積に同一の反応セットアップの蛋白質の生産は 40% を異なることが、また、体積の利回りに影響を与えます。実行して、体積利回りをさらに強めている他、大きな船で観測された体積の収率でのブーストより良い酸素交換 (図 2) を可能にする、反応混合物の高められた表面積のために学説をたてられました。大型のフラット底板を 100 μ L17,31,37,52以上反応の勧めで当たりの反応。

図 1: 文化の成長、細胞抽出液、セットアップ、当たり反応の定量化の生産のためのタイムライン。ユーザーは、この 4 日間ワークフローを通じて、研究用 CFPS プラットフォームを実装できます。試薬の準備は主な時間を表し、この実験の最初のラウンドのためのコスト投資と試薬株が確立された後、大幅に減少します。また、-80 ° c、高速な結果を得るのためのさまざまな段階でタイムラインを開始するユーザーを許可する 1 年間に、細胞ペレットと準備の細胞抽出液を格納することができます。ユーザーは、このワークフローのタイムラインを変更するさまざまな手順で一時停止もできます。この図の拡大版を表示するのにはここをクリックしてください。

図 2:当たりの体積反応収率に及ぼす変更条件。A.エキスの生産性の比較に基づいて様々 な外径600朗読で BL21(DE3) セルを収穫します。このプロットに基づいて、ターゲット蛋白質の少なくとも 1000 μ G/ml を生成する 3.3 の OD600収穫ことをお勧めします。反応は、1.5 mL の microfuge の管に 15 μ L スケールで行われました。B. 2 つの DNA maxiprep 洗浄のプロトコル後精製 DNA クリーンアップとの比較。pJL1 sfGFP プラスミド PCR 精製キット後浄化クリーンアップによって続く 1 つまたは 2 つの洗浄、maxiprep を施行しました。~ 900 μ g/mL タンパク質発現を実現、maxiprep 洗浄の数にかかわらず後精製 DNA クリーンアップを実行するをお勧めします。反応は、1.5 mL の microfuge の管に 15 μ L スケールで行われました。C. 15 μ L 当たり反応は 2 mL から 0.6 mL の microfuge の管に至るまで様々 な容器で実行されます。"Neg"は、反応する DNA テンプレートが追加されたない否定的なコントロールを表します。すべての誤差範囲は、3 通で定量化されたそれぞれのそれぞれの条件の 3 つの独立した反作用の 1 の標準偏差を表します。この図の拡大版を表示するのにはここをクリックしてください。

図 3:キー手順セットアップや生産性を作成するための結果を抽出します。A.超音波氷水風呂超音波処理中に熱を生成しながらサンプルの冷却を確保するための適切なセットアップ。B. 1.5 mL および microfuge の管再懸濁細胞ペレットの前 (左) とポスト (右) 超音波を含みます。その結果ライセート再懸濁細胞ペレットに比べて暗い色相が表示されます。C.上清と細胞ライセート 18,000 × g の遠心分離後のペレットの適切な分離。37 ° C で培養 4 h 後D. CFPS 反応右 (正常反応) に 1.5 mL および microfuge 管 〜 900 μ g/mL で sfGFP レポーター蛋白質の可視蛍光を示します。テンプレート DNA を欠けていると失敗反応をシミュレートする、左側のネガティブ コントロール チューブ no 蛍光で明確な解決策が表示されます。この図の拡大版を表示するのにはここをクリックしてください。

図 4: 蛋白質の表現 5 以上の凍結融解サイクル当たりの変更を抽出します。同じ成長から調製した抽出物を受けた 5 液体窒素による融解凍結融解氷の上でフラッシュを凍結します。5 凍結融解サイクルで sfGFP を表現するため抽出生産性に大きな変化は見られなかった。反応は、1.5 mL の microfuge の管に 15 μ L スケールで行われました。"Neg"は、反応する DNA テンプレートが追加されたない否定的なコントロールを表します。すべての誤差範囲は、3 通で定量化されたそれぞれのそれぞれの条件の 3 つの独立した反作用の 1 の標準偏差を表します。この図の拡大版を表示するのにはここをクリックしてください。

図 5: さまざまな反応に当たり [Mg2+] と [sfGFP] 対ボリュームを抽出。[Mg2 +] 8 mM 〜 14 mM 2 mM 単位で、1 μ L 単位で 7 μ 3 μ L から 〜 ボリュームを抽出します。色コードを表す [sfGFP] 生産高 (赤) (紫) を低から。高蛋白質の生産を維持しながら試薬の効率を最大にするエキスと 10 mM Mg 5 μ L の使用をお勧めします。2+ 〜 30 mg/ml、総たんぱく量を持つブラッドフォードの試金によって定められるようにエキスのため。コンター プロットを生成する元の点は 3 通で測定したそれぞれの条件ごとに 3 つの独立した反作用のエンドポイント蛍光を基づいていた。反応は、1.5 mL の microfuge の管に 15 μ L スケールで行われました。この図の拡大版を表示するのにはここをクリックしてください。
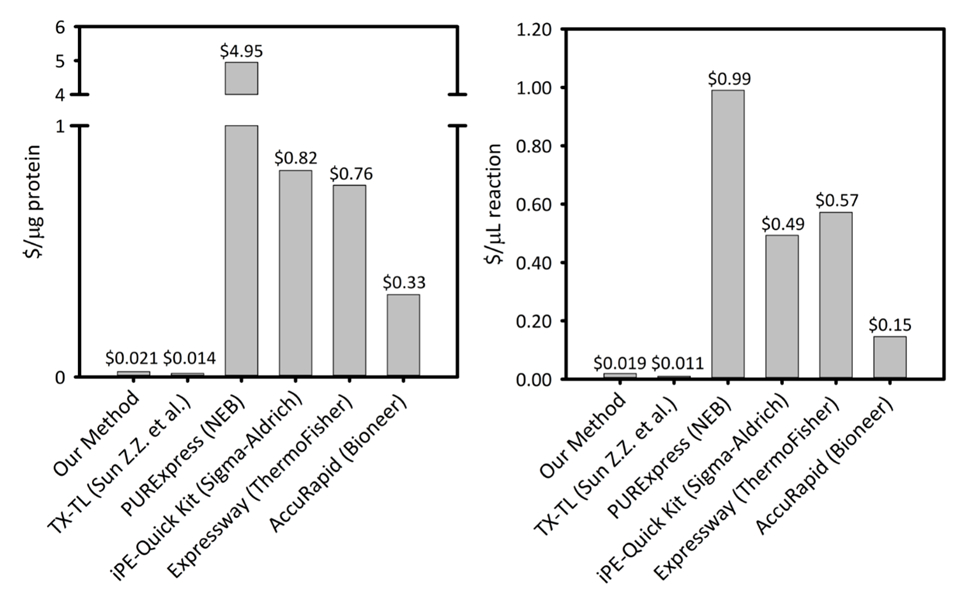
補足図 1: 6 無細胞タンパク質合成プラットフォーム間で産生される蛋白の 1 マイクログラムあたりと反応 1 マイクロリットルあたりのコストです。当社のプラットフォームは、生産性の変化と価格で 5 異なる細胞無細胞タンパク質合成キット/プラットフォームと比較されます。当社の超音波による当たりプラットフォームはほとんど商業キット、キットの使いやすさは、残存コストその他学術 CFPS プラットフォームに匹敵する中の反応セットアップよりも蛋白質の両方の $/μ g で $/μ L 反応の費用対効果です。この図の拡大版を表示するのにはここをクリックしてください。
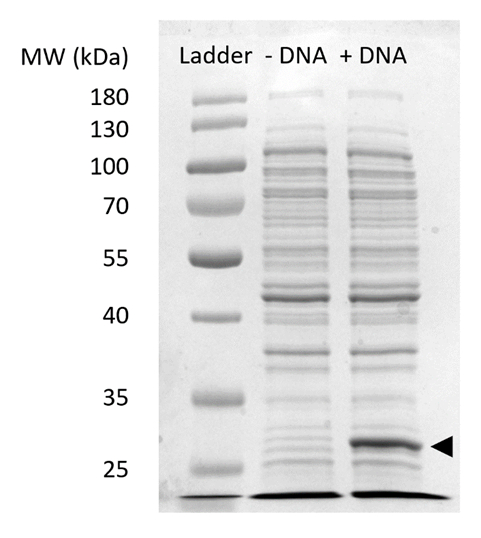
補足図 2: 当たりの sfGFP 式の SDS-PAGE 。無細胞タンパク質合成反応 (+ DNA)、なし (-DNA) sfGFP のための DNA のテンプレート 12% 上 27 kDa (黒矢印) で観測された sfGFP の式を示す SDSPAGE ゲルで実行されました。伝統的な SDS ページの技術が使用されました。各サンプルはセル全体蛋白質のブラッドフォードの試金の定量化に基づく総蛋白 18 μ g 含まれている抽出ゲルにロード。蛍光強度の測定と私達の標準曲線に基づいて、予想では、「+ DNA」レーンには sfGFP 0.42 μ g が含まれています。これらのサンプルを得るために当たり反応は、図 3と一貫性のある容積生産 1.5 mL の microfuge の管に 15 μ L スケールで実行しました。この図の拡大版を表示するのにはここをクリックしてください。
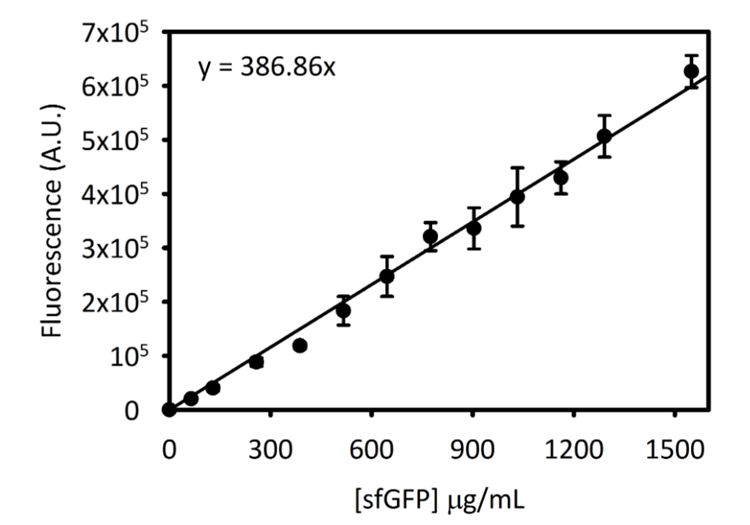
補足図 3: Cytation 5 sfGFP の標準曲線。この曲線は、上記の方法を使用して決定しました。この標準曲線を用いた μ g/mL でこの原稿は [sfGFP] にエンドポイントの蛍光測定値から変換された用に収集されるすべてのデータ。この図の拡大版を表示するのにはここをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
無細胞タンパク質合成は、様々 な生化学システムのラピッドプロトタイピング製造に至るアプリケーションに強力な有効にする技術として浮上しています。広範なアプリケーションは、監視、操作、およびリアルタイムでの細胞機械装置を強化する能力によってサポートされます。このプラットフォーム技術の拡大の影響から、広範な適応は推移しているメソッドの実装の技術的なニュアンスのため遅い。この取り組みを通じて、新しいラボでこの技術を確立するため、シンプルさと明快さを提供するために目指しています。これに向けて、私達のプロトコル、エシェリヒア属大腸菌の-ベースの無細胞タンパク質合成プラットフォームを 4 日の起動時間訓練所非専門家、大学生 (図 1) などが実現できます。また、試薬および抽出物の在庫が生産され、一度それ以降当たりバッチ反応を設定、抱卵でき 5 h だけで定量化します。単一の 1 の L 細胞の増殖につながります 400 15 μ L 当たり反応のため十分に抽出無細胞試薬の 1 つのバッチの準備が何千もの反応をサポートできますが。試薬の準備はさらに大きなストックが必要な場合上げもすることができます。CFPS 反応は、さまざまな並列で条件のテスト 96 ウェル プレートまたは PCR チューブを使用して高スループット方法でセットアップをすることができます。図 2に見られるように小型の船を使用するとき、体積の利回りが低下します。CFPS 反応もスケール アップが可能マイクロリットルから全反応量のミリリットル万単一条件の総蛋白質収量を高めるために。スケール ボリューム アップ、主な考慮事項は、体積反応収率低下反応の表面の区域の容積比減少37,52です。蛋白質の表現のような体積収量を維持しながらスケール アップするためにユーザーは多数反応容器に反作用ボリュームを分割や容器のサイズを増やす必要があります。反応スケール 15 μ ・ 100 μ L のボリュームに至るまで並行して多数の 15 μ L 反応が推奨されます。ボリュームに 100 μ L を超える反応、24 ウェルのフラット底板をお勧めします、12 ウェル プレート 600 μ L を超える反応ボリュームをお勧めします。反応ボリュームと船のような組み合わせは、スケール アップ17,31,37,52時体積反応収率の一貫性を提供します。平行板の複数の井戸を利用してこれらのボリュームを超えるスケール アップを実現できます。この形式を使用すると、反応は以上 10 mL 容量に拡張できます。反応量反応容器の組み合わせの最適化、反応の生産性を損なうことがなく製造のアプリケーションをサポートできます。
このプロトコルを実行すると、うまく抽出を実行に関連付けられているインジケーターと同様、体積反応収率に影響を与える、いくつか重要な考慮事項があります。適切な換散を確保し、機能性転写/翻訳機械の変性を防ぐためには、溶解中に生成される熱を軽減するために重要です。(図 3 a) 超音波処理中に急速に熱を放散する超音波処理中に氷の水のお風呂に携帯巻き上がりを浸します。効果的なセル換散のインジケーターは、事前に超音波処理のサンプル (図 3 b) と比較して細胞ライセートの暗い外観の登場です。ユーザーの柔軟性、超音波発生装置とプローブを図 3 aに示すように、適応ボリュームの範囲に再懸濁細胞の 15 mL に 100 μ L から。これを行うには、ユーザーがセルの音量の換散のため配信ジュール数を調整できます。さらに、2 つの相補的なアプローチを通じて大量のエキスを準備できます。ユーザーは並列で複数のチューブを超音波および超音波セル巻き上がり、大量ボリューム前述29,45に比例してエネルギーの量をスケーリングできます。エキスの品質を示す別のステップは遠心分離のステップ次のセル換散。セル換散、清 (テンプレート転写/翻訳に機能しなくする断片化された DNA 転写/翻訳機械) 間の明確な区分を提供する 18,000 × g で遠心分離を推奨しますポストとペレット (沈殿したタンパク質や細胞膜などの細胞成分を望ましくない) (図 3)。我々 は、遠心分離を発見した g 18,000 x で分離、12,000 × g などの低速時にスピンと比較して再現性が向上の結果を向上させます。便利なテーブル トップ冷却遠心機、12,000 × g の最小値を達成するための能力を使用することをお勧めします。また、一般的で 30,000 × g、適切な機器が使用可能な54,55,56,57,58,の場合に考慮すべき手順は59,60. 抽出パフォーマンスは受けませんこのステップで遠心分離速度適切な分離が達成されることを考える。目的の上澄みを削除する場合は、この汚染は抽出物の生産性が低下しますので、上澄みとペレット間の境界に存在する任意の曇り材料を避けるためにことをお勧めします。上澄みの純度を目指してより生産的な抽出の結果し、新しいユーザーの得られた抽出物の減らされた量の価値があります。
バッチ間が提案した方法は再現性のある、最低限の専門知識を持つ科学者によって実行することができることができる注意し反応の反応の変化が重要です。これは、溶解後超音波61のプロテオームの組成の変化に起因する可能性があります。我々 が観察しているバッチ間変動は一般的に T7RNAP の補充とマグネシウム濃度の最適化時に減少します。T7RNAP の外因性加算に最適なタンパク質発現をサポートする当たり反応の間で一般的な我々 は向上させる - BL21*(DE3) と 16 μ G/ml の最終的な集中に補足 T7RNAP の内因性表現 - T7RNAP の 2 つのソースを持っていることを見つける新しいユーザー45,46のバッチ間の再現性。経験を持つユーザーは、必要な場合にのみ T7RNAP の単一のソースを活用する彼らの実験を変更できます。抽出の新しいバッチと Mg2 +濃度の適切な調整の総たんぱく量の定量化は、体積蛋白質表現利回りのバッチ間変動を減少させるまた助けるかもしれない。サイズおよび興味の蛋白質、遺伝子のコドン使用頻度、発現ベクター使用62 のタイプと同様、興味の遺伝子の対応するリボソーム結合部位の構造の違いによるタンパク質発現の変化することができます。 ,63。これらの理由から、当たり反応から減らされた容積収率で生じるモデル蛋白質 sfGFP だけでなく、いくつかの蛋白質を表現できないこと。
紹介した当たりテクニックの制限には、直接代謝工学とプロトコルに追加変更することがなく、式の条件の調整など、携帯無料のすべてのアプリケーションに適していない可能性がありますがあります。ただし、このプロトコルを新しい所で当たりプラットフォームを確立するための基礎を提供し、彼らのラボで導入細胞反応を実装する能力を持つ非専門家を提供と考えています。初期実装後研究者は、フィールドの他の文献に基づく具体的なアプリケーションの独自の変更を行うプラットフォームで実験できます。
(労働および機器のコストを除く)、当たりプラットフォーム コスト $0.021/μ g タンパク質を作る反応セットアップの容易さを損なうことがなく市販キットで価格競争力のある体制。反応の μ L あたり比較コストの評価は、似たような傾向 (カスタマーインフォメーション センターの障害図 1) を表示します。推定するスタートアップのコスト 〜 すべての試薬の 4500 ドルと、超音波発生装置などの特殊な装置の追加 $3,200。この手順を完了する時間を地面からすべての試薬の準備のための ~ 26 h になると推定されます。ただし、試薬の大型株が準備されると、労働の需要を大幅に減少させます。また、プラットフォームと経験を得られるよう我々 は細胞の成長のサイズをスケール アップをお勧めします、抽出の準備、および時間効率を最大化する試薬調製。スタートアップのコストを考えると、高スループットの努力、合成生物学のアプリケーションに当たりプラットフォームをお勧めし、タンパク質発現条件ないため伝統的なタンパク質式プラットフォームと互換性のあるセルのと競合するには生化学および実行可能性の制約。CFPS プラットフォームによって目的のテクニックが有効になっているような特殊な場合、体内式の上当たりの大きいコストが正当化されます。
CFPS プラットフォームの継続的な開発は従来難治性性蛋白質、非標準アミノ酸の酵素経路、生産および代謝工学などバイオ テクノロジーの努力により広範なユーティリティを提供する可能性が定款と不自然なタンパク質発現・成層薬製造・幹教育64,65,66の教室、研究室を超えて進出。これらの努力は、株当たりプラットフォームの詳細な特性評価のための継続的な努力によってさらにサポートされます。細胞抽出液の組成のより良い理解は、改善反応収率と反応条件61,,6768柔軟性に向けて継続的な改良をもたらすでしょう。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者はない競合する金銭的な利益やその他の利害の関係があることを宣言します。
Acknowledgments
著者は、博士ジェニファー VanderKelen、アンドレア ・ Laubscher とテクニカル サポートのトニー Turretto、ウェズリー ・花王、Layne ウィリアムズ役に立つ議論のためのクリストファー ・高さを確認したいと思います。著者も認める法案とリンダ霜基金、バイオ テクノロジーの創造活動助成プログラム (RSCA 2017) Scholarly、シェブロン バイオ テクノロジー応用研究基金助成、Cal Poly 研究応用センター資金・国立科学財団 (NSF-1708919)。MZL は、カリフォルニア州立大学大学院助成を認めています。MCJ は、陸軍研究所 W911NF-16-1-0372 を認めている、全米科学財団助成金 MCB 1413563 と MCB 1716766、空軍研究所優秀グラント FA8650 センター-15-2-5518、防衛脅威削減局許可ダビデとルシール パッカード財団、カミーユ ドレイファス先生学者プログラム部門のエネルギー BER グラント デ-SC0018249、人間のフロンティア科学プログラム (RGP0015/2017 年)、エネルギー省の共同ゲノム研究所 ETOP グラント HDTRA1-15-10052/P00001 とサポートのためシカゴのコミュニティ信頼でサール資金からのサポートとシカゴ医療コンソーシアム。
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}