

Summary
Denne protokollen detaljer trinn, kostnader, og nødvendig å generere E. coliutstyr-basert celle ekstrakter og implementere i vitro protein syntese reaksjoner innen 4 dager eller mindre. For å utnytte denne plattformen bred Søknadsfrist fleksibel natur, diskutere vi reaksjonen forhold som kan tilpasses og optimalisert.
Abstract
Over de siste 50 årene, Cell-Free Protein syntese (CFPS) har dukket opp som en kraftig teknologi for å utnytte av transcriptional og translasjonsforskning antall celler i et reagensrør. Obviating behovet for å opprettholde levedyktigheten til cellen, og eliminere mobilnettet barrieren, er CFPS fundamentet til emerging programmer i biomanufacturing av tradisjonelt utfordrende proteiner, samt programmer i rapid prototyping for Metabolsk engineering og funksjonell genomforskning. Våre metoder for å implementere en E. coli-basert CFPS plattform at nye brukere tilgang til mange av disse programmene. Her beskriver vi metoder for å forberede pakke ved bruk av beriket medier, forbløffe flasker og en reproduserbar metode for tunable sonication-baserte celle lysis. Dette ekstrakt kan deretter brukes til protein uttrykk kan produsere 900 µg/mL eller flere super mappen grønne fluorescerende protein (sfGFP) i bare 5 timer fra experimental oppsettet til dataanalyse, gitt at riktig reagens aksjer er utarbeidet på forhånd. Anslått oppstart prisen for innhenting reagenser er $4500 som vil opprettholde tusenvis av reaksjoner til en estimert kostnad på $0.021 per µg protein produsert eller $0.019 per µL av reaksjonen. I tillegg speilet metodene protein uttrykk enkel reaksjon oppsettet i kommersielt tilgjengelige systemer på grunn av optimalisering av reagens pre mikser, til en brøkdel av prisen. For å aktivere brukeren å utnytte CFPS plattformen bred Søknadsfrist fleksibel natur, har vi identifisert en rekke aspekter av plattformen som kan innstilt og optimalisert avhengig av tilgjengelige ressurser og protein uttrykk utfall ønsket.
Introduction
Cellen gratis Protein syntese (CFPS) har dukket opp som en teknologi som har låst opp en rekke nye muligheter for protein produksjon, funksjonell genomforskning, metabolske engineering og mer i de siste 50 år1,2. Sammenlignet med standard i vivo protein uttrykk plattformer, CFPS gir tre viktige fordeler: 1) cellen-fri natur plattformen gjør produksjonen av proteiner som ville være potensielt giftig eller utenlandske til celle3,4 ,5,6; 2) inaktivering av genomisk DNA og innføring av en mal DNA koding gene(s) rundt kanalen alle systemisk energien i reaksjonen til produksjon av protein(s) steder. og 3) den åpne naturen plattformen gjør det mulig å endre og overvåke reaksjonen forhold og komposisjon i sanntid7,8. Denne direkte tilgang til reaksjonen støtter augmentation av biologiske systemer med utvidet kjemikalier og redoks forholdene for produksjon av romanen proteiner og tuning av metabolske prosesser2,9, 10. direkte tilgang gir også brukeren å kombinere CFPS reaksjon med aktivitet analyser i et enkelt-potten system for rask utformingen-bygge-test sykluser11. Kapasitet til å utføre CFPS reaksjonen i små dråper eller på papir-baserte enheter videre støtter høy gjennomstrømming oppdagelsen innsats og rask prototyping12,13,14,15 ,16. Disse fordelene og plug and play natur systemet, CFPS har unikt aktivert en rekke bioteknologi programmer som produksjonen av proteiner som er vanskelige å solubly express i vivo17, 18,19,20, påvisning av sykdom21,22,23, på etterspørsel biomanufacturing18,24 ,25,26,27, og utdanning28,29, som viser fleksibilitet og nytten av celle-fri plattformen.
CFPS systemer kan genereres fra en rekke råolje lysates fra både prokaryotic og eukaryotic linjer. Dette gir forskjellige alternativer i systemet av valg, hver har fordeler og ulemper avhengig av anvendelsen av interesse. CFPS systemer også variere i Forberedelsestid, kostnader og produktivitet. De benyttes vanligvis celle ekstrakter produseres fra hvetespirer, kanin retikulocytt, insekt celler og Escherichia coli celler, sistnevnte er den mest kostnadseffektive hittil mens de høyeste volumetriske gir protein30 . Mens andre CFPS systemer kan være fordelaktig for medfødte post-translasjonell modifikasjon maskiner, nye programmer ved hjelp av E. coli-baserte maskiner kan bygge bro ved å generere site-specifically fosforylert og Glycosylated proteiner på etterspørsel,31,,32,,33,,34,,35.
CFPS reaksjoner kan kjøres i enten bakst, kontinuerlig-utveksling celle-free (CECF) eller kontinuerlig flyt celle-fritt (CFCF) format. Batch-format er et lukket system som reaksjon levetid er begrenset på grunn av avtagende mengder reaktantene og akkumulering av hemmende biprodukter av reaksjonen. CECF og CFCF øke levetiden til reaksjonen og føre dermed til økt volumetriske protein gir sammenlignet med satsvise reaksjonen. Dette gjøres ved at biprodukter av proteinsyntese fjernes fra reaksjonen fartøyet mens nye reaktantene leveres i løpet av reaksjon2. Når det gjelder CFCF, kan protein rundt også fjernes fra reaksjon kammer, i CECF, protein av interesse i reaksjon kammeret består av et halvt gjennomtrengelig membran36,37. Disse metodene er spesielt nyttig i å overvinne dårlig volumetriske gir vanskelig-å-uttrykker proteiner av interesse38,39,40,41,42, 43. Utfordringene med å implementere CECF og CFCF tilnærminger er at 1) mens de resultere i mer effektiv bruk av bio maskiner ansvarlig for transkripsjon og oversettelse, de krever spesielt større mengder reagenser som øker totalkostnad og 2) de krever mer komplekse reaksjon oppsett og spesialisert utstyr forhold til satsvise format44. For å sikre tilgjengelighet for nye brukere, beskrevet protokollene heri fokus på batch-format på reaksjon mengder 15 µL med konkrete anbefalinger for å øke volumet reaksjon til milliliter skala.
Metodene presenteres her at ikke-eksperter med grunnleggende laboratorium ferdigheter (for eksempel studentene) kan implementere cellevekst, ekstra forberedelse og satsvis format reaksjon installasjon for en E. coli-basert CFPS system. Denne tilnærmingen er kostnadseffektiv sammenlignet kommersielt tilgjengelig kits uten å ofre enkel kit-baserte reaksjon oppsett. Videre lar denne tilnærmingen programmer i laboratoriet, og i feltet. Når beslutter å implementere CFPS, bør nye brukere grundig vurdere effekten av konvensjonelle protein uttrykk for oppstart investeringer, som CFPS ikke kan bli bedre i alle tilfeller. CFPS metodene beskrevet her at brukeren direkte implementere en rekke programmer, inkludert funksjonell genomforskning, høy gjennomstrømming testing, produksjonen av proteiner som er vanskelige for i vivo uttrykk, i tillegg til feltet programmer inkludert biosensors og pedagogisk kits for syntetisk biologi. Tilleggsprogrammer som metabolske engineering, tuning av protein uttrykk forhold, sykdom gjenkjenning og skalere opp bruke CECF eller CFCF er fortsatt mulig, men kan kreve erfaring med CFPS plattform for videre modifikasjon av reaksjon forhold. Våre metoder kombinere vekst i beriket media og forvirret flasker, med relativt rask og reproduserbar metoder for cellen lysis gjennom sonication, etterfulgt av en forenklet CFPS reaksjon oppsett som utnytter optimalisert premixes45. Mens metodene mobilnettet vekst har blitt noe standardisert innen dette feltet, varierer metoder for cellen lysis mye. I tillegg til sonication omfatter felles lysis metoder bruk av en fransk trykk, en homogenizer, perle beaters, eller lysozyme og andre biokjemiske og fysiske avbrudd metoder46,47,48, 49. bruke våre metoder, ca 2 mL olje celle pakke hentes per 1 L celler. Dette antallet av cellen ekstrakt kan støtte fire hundre 15 µL CFPS reaksjoner, hver produserende ~ 900 µg/mL reporter sfGFP protein fra malen plasmider pJL1-sfGFP. Denne metoden koster $0.021/µg av sfGFP produsert ($.019/µL av reaksjonen), unntatt arbeidskraft og utstyret (supplerende figur 1). Denne metoden starter fra scratch, kan implementeres i 4 dager ved en enkelt person, og gjenta CFPS reaksjoner kan fullføres innen timer (figur 1). I tillegg kan protokollen skaleres i volumet for større batcher av forberedelse av reagenser etter brukerens behov. Viktigere, kan protokollen presenteres her implementeres av laboratoriet trent ikke-eksperter som studenter, som det bare krever grunnleggende laboratorium ferdigheter. Fremgangsmåtene nedenfor og på videoen har blitt spesielt utviklet for å forbedre tilgjengeligheten av E. coli CFPS plattform for bred bruk.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Media forberedelse og cellevekst
-
Dag 1
- Strek E. coli BL21*(DE3) celler fra en glyserol lager på en LB agar plate og ruge minst 18 h på 37 ° C.
- Forbered 50 mL av LB media og autoklav løsningen på en flytende syklus i 30 min på 121 ° C. Lagre ved romtemperatur.
-
Dag 2
- Forberede 750 mL av 2 x YTP media og 250 mL 0,4 M D-glukose løsning som beskrevet i tilleggsinformasjon.
- Hell 2 x YTP media i en autoklaveres 2.5 L forbløffet kolbe og D-glukose løsningen i en autoklaveres 500 mL glassflaske. Autoclave begge løsninger på en flytende syklus i 30 min på 121 ° C.
- Kontroller at begge sterilt løsninger lagres på 37 ° C hvis cellevekst utføres på dagen å maksimere vekstrater på vaksinering. Ikke kombinere løsninger til vaksinering.
Merk: Løsninger kan lagres ved 4 ° C i 1-2 d eventuelt om 2 x YTP media er svært utsatt for forurensning. - Start en overnatting kultur av BL21(DE3) ved å vaksinere 50 mL av LB medier med en enkelt koloni av BL21(DE3) bruker en sterilisert sløyfen og steril teknikk for å unngå forurensning.
- Plasser den 50 mL av BL21*(DE3) LB kultur i en 37 ° C 250 rpm risting inkubator og vokse over natten for 15-18 h.
- Forberede og sterilisere alle materialer som kreves for dager 3 og 4, inkludert: to 1 L sentrifuge flasker, 4 x kaldt 50 mL konisk rør (veier og registrere massene av tre) og mange 1,5 mL microfuge rør.
-
Dag 3
- Fjern 50 mL natten kultur BL21*(DE3) i LB settefiskanlegg risting og måle OD600 på et spektrofotometer ved hjelp av en 1:10 fortynning med LB media. Beregne volumet av natten kultur nødvendig å legge til 1 L medier for en start OD600 0,1 (For eksempel hvis en OD600 på en 1:10 fortynning leses som 0,4, vaksinere 25 mL ufortynnet OD600 = 4.0 natten kultur i 1 L 2 x YTP G).
- Fjerne varmet 2 x YTP media og D-glukose løsninger settefiskanlegg 37 ° C med 50 mL av LB kultur. Bruker steril teknikk, nøye hell D-glukose løsningen i 2 x YTP media (unngå sidene av forbløffet kolbe).
Merk: Tillegg av D-glukose fullfører oppskriften på 1 L 2 x YTPG. - Opprettholde steril teknikk, vaksinere 1 L 2 x YTPG løsning med riktig mengde 50 mL kultur for å starte 1 L kulturen på en 0,1 OD600. Umiddelbart plassere inokulerte 1 L kulturen i en 37 ° C risting inkubator på 200 rpm.
- Ta den første OD600 leser etter den første timen av vekst (lag fase typisk tar 1 time). Ikke utvanne kulturen. Fortsett å ta OD600 målinger ca hvert 20-30 min til OD600 når 0,6.
- Ved å nå OD600 = 0,6, Legg 1 mL 1 M IPTG (siste konsentrasjon i 1 L kultur = 1 mM) til 2 x YTPG kultur.
Merk: Ideelle induksjon OD600 er 0,6; men er en rekke 0,6 0,8 akseptabelt. Induksjon av IPTG er for endogene produksjon av T7 RNA Polymerase (T7RNAP). - Etter innledningen, måle OD600 ca hvert 20-30 min før den når 3.0.
Merk: Avkjøl sentrifuge 4 ° c i løpet av denne tiden. Forberede kaldt S30 bufferen som beskrevet i den utfyllende informasjonen. Hvis S30 bufferen er forberedt på forhånd, kontrollerer du at DTT ikke legges helt frem til bruk. - Når OD600 når 3.0 (figur 2A), hell kulturen i en kald 1 L sentrifuge flasken i en is-vannbad. Forberede en vannfylte 1 L sentrifuge flaske av samme vekt som en saldo i sentrifugen.
Merk: Absorbance verdier varierer fra instrument-til-instrument. OD600 av innhøstingen av BL21(DE3) er ikke en følsom variabel, anbefales det at brukeren evaluere og optimalisere denne variabelen som feilsøking mål. Større Spektrofotometrene kan resultere i relativt lavere OD600 opplesninger sammenlignet med mindre cuvette-baserte Spektrofotometrene. - Sentrifuge 1 L flasker for 10 min på 5000 x g og 10 ° C til pellets celler.
- Langsomt helle av nedbryting og i tråd med institusjonens biologisk avfall prosedyrer. Sted pellet på is.
- Bruker en steril slikkepott, skrape celle pellet fra sentrifuge flasken og overføre den til en kald 50 mL konisk rør.
- Legg til 30 mL kaldt S30 bufferen til konisk røret og resuspend celle pellet av vortexing med korte serieopptak (20-30 s) og hvileperioder (1 min) på is til fullt resuspended med ingen biter.
- Når pellet er fullt resuspended, bruke en annen 50 mL konisk tube med vann som balanse og sentrifuge for 10 min på 5000 x g og 10 ° C (pre kjølt ned til 4 ° C).
Merk: Denne fullfører 1st med 3 vasker når høsting cellene. - Hell nedbryting og i tråd med institusjonens biologisk avfall prosedyrer. Resuspend pellets med 20-25 mL kaldt S30 buffer og sentrifuge for 10 min 5000 x g og 10 ° C (pre kjølt ned til 4 ° C).
Merk: Denne fullfører 2nd av 3 vasker. - Igjen, helle ut nedbryting og i tråd med institusjonens biologisk avfall prosedyrer. Legge til nøyaktig 30 mL S30 buffer og vortex igjen å resuspend pellet.
- Bruker 3 pre vektet, kalde 50 mL konisk rør og en serologisk pipette filler med en bakteriefri pipette, aliquot 10 mL av resuspended pellets/S30 buffer blandingen i hver av de 3 koniske rørene.
Merk: Dele cellene i 3 rør er ikke nødvendig, men dette trinnet resulterer i mindre celle pellets (~ 1 g) for økt bekvemmelighet i senere trinn. - Sentrifuge alle rør, bruke aktuelle balanserer nødvendig for 10 min på 5000 x g og 10 ° C (pre kjølt ned til 4 ° C).
Merk: Denne fullfører det endelige wash trinnet. - Hell nedbryting og i tråd med institusjonens biologisk avfall prosedyrer. Fjerne overflødig S30 bufferen ved å nøye tørke innsiden av koniske tunnelbane og lue med en ren vev; unngå å berøre pellet.
- Reweigh rør på en analytical balanse og registrere siste pellet vekten på hver rør.
Merk: Protokollen kan pauses på dette punktet. Pellets kan flash frosset i flytende nitrogen og lagret ved-80 ° C i opp til et år før behov for ekstra klargjøring.
2. råolje celle ekstra forberedelse - dag 4
- For ekstra forberedelser, holde celler kaldt på is på hvert trinn. Legg 1 mL kaldt S30 buffer per 1 g cellemasse pellet. Kontroller at dithiothreitol (DTT) har vært supplert til S30 buffer til en siste konsentrasjon av 2 mM.
Merk: Avkjøl microcentrifuge 4 ° c i løpet av denne tiden. - Resuspend celle pellet av vortexing med korte serieopptak (20-30 s) og hvileperioder (1 min) på is til fullt resuspended. Hvis rørets er vanskelig, la pellets på isen i 30 minutter til defrost.
- Overføre 1,4 mL av resuspended celler i en 1,5 mL microfuge tube.
- Plass en 1,5 mL tube som inneholder 1,4 mL av resuspended celler i et isvann bad i et beaker. Sonicate for 45 s på etterfulgt av 59 s av 3 totalt sykluser, med amplituden satt til 50%. Lukk og invertere rør som bland forsiktig i av perioder. Totalt lever 800-900 J energi til hver 1,5 mL microfuge rør som inneholder 1,4 mL av resuspended celler (figur 3A og 3B).
Merk: Dette trinnet er følsom for sonicator type og modellen og bør optimeres hvis utstyret er annerledes enn oppført for denne prosedyren. To supplerende tiltak kan brukes til å skalere opp hvor mye ekstra utarbeidet under dette trinnet: 1) flere 1,5 mL microfuge rør kan være sonicated parallelt, og/eller 2) større volumer kan være sonicated i konisk rør (opptil 15 mL av cellen rørets per tube) , skalering mengden energi levert som tidligere beskrevet 29,45. - Umiddelbart etter at sonication er fullført, Legg 4,5 µL av 1 M DTT (supplere en ekstra 2 mM DTT) til 1,4 mL av lysate og invertere flere ganger å blande. Sett røret på is. Gjenta trinn 2.4 og 2.5 for noen ekstra rør av resuspended celler før du går videre til sentrifugering.
- Microcentrifuge eksempler på 18 000 x g og 4 ° C i 10 min (Figur 3 c).
- Pipetter nedbryting i en ny 1,5 mL microfuge tube. Ikke forstyrr pellet; Det anbefales å etterlate noen nedbryting for å opprettholde renhet enn å forstyrre pellet i forsøk på å maksimere avkastning.
- Inkuber nedbryting fra det tidligere trinnet på 250 rpm og 37 ° C for 60 min ved taping rør for å riste plattform inkubator (dette er avrenning reaksjonen).
- Microcentrifuge eksempler på 10.000 x g og 4 ° C i 10 min.
- Fjern nedbryting uten å forstyrre pellet og overføre den til en ny tube. Opprette mange 100 µL dele pakke for lagring.
Merk: Protokollen kan pauses her, og ekstrakt kan flash frosset i flytende nitrogen og lagret ved-80 ° C i opptil et år inntil for CFPS reaksjoner. Minst 5 fryse-Tin sykluser kan være gjennomgått uten skade for ekstra produktivitet (Figur 4).
3. cell-Free Protein syntese Batch Format reaksjoner
- Tine Solutions en og B, DNA mal, BL21(DE3) ekstrakt (hvis frossen), T7RNAP og en aliquot av molekylære klasse vann.
Merk: CFPS reaksjon malen finnes i Tilleggsinformasjon. Løsninger A og B oppskrifter finnes i Tilleggsinformasjon og tilsvarer spesifikke konsentrasjoner for mange reagenser til støtte PANOx-SP basert energisystem for CFPS. Rollen som hver reagens og akseptabel variasjon i disse reagens konsentrasjonene som støtter CFPS har vært bestemt50. En T7RNAP rensing protokoll kan finnes i tilleggsinformasjon51. Ekstra T7RNAP kan øke volumetriske gir, men er ikke nødvendig hvis T7RNAP er indusert i cellevekst. Plasmider DNA mal (pJL1-sfGFP) kan være forberedt med en maxiprep kit med to vasker ved å vaske bufferen i pakken, etterfulgt av en etterbehandling DNA-opprydding bruker en PCR rensing kit (figur 2B). Lineær DNA maler kan også brukes i CFPS reaksjoner. - Merke den nødvendige mengden av microfuge rør nødvendig for CFPS reaksjoner.
Merk: Reaksjoner kan utføres i ulike fartøy størrelser, men en mindre fartøy kan redusere volumetriske protein avkastning (figur 2C). Skalere opp en reaksjon i samme størrelse fartøy kan også redusere volumetriske avkastning, som en funksjon av avtagende oksygen utveksling, grunnet en nedgang i overflaten området som volumkontrollen. Når økende reaksjon volum over 100 μL, anbefales det å bruke flat bunn bra plater 31,37,52. - Legge til 2,2 µL løsning A, 2.1 µL av løsning B, 5 µL av BL21*(DE3) ekstra, 0,24 μg av T7RNAP (16 μg/mL siste konsentrasjon), 0.24 ng DNA mal (16 ng/mL siste konsentrasjon) og vann for å bringe det siste bindet til 15 µL.
Merk: Vortex løsninger A og B ofte under installasjonen av reaksjon for å unngå sedimentering komponenter og sikre at hver reaksjon mottar en homogen aliquot av hver løsning. Unngå vortexing ekstrakt, i stedet Inverter røret å blande. - Etter reagenser har blitt lagt til reaksjonen, bland hver rør av pipettering opp og ned eller forsiktig vortexing samtidig som man sikrer at siste reaksjonsblandingen er kombinert i en enkelt 15 µL perle nederst på 1,5 mL microfuge røret.
- Plass hver reaksjon i 37 ° C inkubator uten risting 4 h eller 30 ° C over natten.
Merk: Vellykket reaksjoner kan kvalitativt vurderes visuelt basert på den grønne fargen på sfGFP produktet innenfor CFPS reaksjonsblandingen (figur 3D). Uttrykk for protein rundt kan også bli bekreftet av SDS-side (supplerende figur 2).
4. kvantifisering av Reporter Protein, [sfGFP]
- Legg 48 µL 0,05 m HEPES, pH 8 i hver også nødvendig for kvantifisering (vanligvis utføres i tre eksemplarer per reaksjon rør).
- Fjern reaksjoner fra inkubator. Pipetter opp og ned for å blande hver reaksjon, deretter overføre 2 µL av reaksjonen til de 48 µL 0,05 m HEPES, pH 8. Pipetter opp og ned igjen i brønnen å blande.
- Når alle reaksjoner er lastet og blandet 96 godt platen i fluorometer og sfGFP endepunkt fluorescens.
Merk: Eksitasjon og utslipp bølgelengder for sfGFP fluorescens kvantifisering er 485 nm og 510 nm, henholdsvis. - Bruker en tidligere genererte standardkurve, finne ut [sfGFP] fra innhentet fluorescens målingene.
Merk: Instruksjonene for å generere en standardkurve sfGFP konsentrasjon versus fluorescens intensitet finnes i tilleggsinformasjon (supplerende figur 3). Brukernes ville nød å opprette en standardkurven for sine instrument siden instrument følsomhet kan variere.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Vi har presentert en sonication-baserte E. coli ekstrakt forberedelse protokoll som kan gjennomføres over en firedagers, med figur 1 demonstrere fremgangsmåter for nedbryting over hver dag. Det er malleability til fremgangsmåten som kan gjennomføres på hver dag med ulike pause poeng, men vi har funnet denne arbeidsflyten til å være den mest effektive kjøre. I tillegg både celle pellets (trinn 1.3.18) og fullt forberedt ekstrakt (trinn 2.10) er stabilt på-80 ° C i minst ett år, tillater brukeren å opprette større bestander av hver lagre for bruk på et senere tidspunkt17. Ikke bare er ekstraktet stabil over lange tidsperioder, men ekstrakt kan også gjennomgå minst fem fryse tine sykluser uten en betydelig tap av produktiv (Figur 4). Dette gir større dele ekstrakt lagres for flergangsbruk hvis fryser lagringsplass er begrenset. Vi anbefaler imidlertid mindre Quidel (~ 100 μL) av ekstrakt mulig.
Med hver ny pakke forberedelse anbefaler vi at brukeren utfører en magnesium titrering for å bestemme den optimale mengden av magnesium for at batch av ekstrakt. Brukere kan kvantifisere parti til parti variasjon i totale protein konsentrasjon av cellen ekstrakt av Bradford analysen. For høyere utføre ekstrakter, vi vanligvis ser totale protein konsentrasjoner av 30-50 mg/mL, og innenfor dette området er det ingen åpenbar sammenheng mellom totalt protein konsentrasjoner og celle ekstrakt ytelse. Derfor anbefaler vi at brukerne finjustere magnesium konsentrasjoner tilsvarende for å sikre at protein og nukleinsyre funksjonalitet maksimeres for hvert ekstra sett. Magnesium nivåer er viktig for riktig DNA-replikasjon, transkripsjon og oversettelse, men overdreven nivåer kan være skadelig for disse prosesser53. For å vise denne avhengigheten, vi har utført en co titrering magnesium og ekstra volum for å fastslå den optimale kombinasjonen som minimaliserer ekstra nødvendig, samtidig opprettholde en produktiv reaksjon (figur 5). Fra dette eksperimentet anbefaler vi bruke 5 µL av ekstrakt og 10 mM Mg2 + for ekstra med et totalt proteininnhold på 30 mg/mL, for å oppnå over 1000 μg/mL av sfGFP.
Vår erfaring med CFPS har også tillatt oss å bestemme trinn i protokollen som kan varieres uten skade produktivitet generelt av systemet, og andre som er integrert for en høy ytelse CFPS system. Særlig den siste OD600 for cellen påvirker ikke betydelig det endelige resultatet av CFPS reaksjon, og cellene kan feasibly høstes overalt 2.7-4.0 OD600. Dette representerer den tidlige eksponentielle fasen av veksten der ribosom konsentrasjon per celle er den høyeste og translasjonsforskning maskiner er den mest aktive støtte rask vekst. Denne observasjonen brukerne fleksibilitet til å optimalisere sine egne prosedyrer. Vi anbefaler høsting på omtrent 3.0 OD600 for å fange cellene på et OD600 nærmere til 3.3 av tiden høsting er fullført (figur 2A). Variabler som påvirker CFPS gir inkluderer mal DNA kvalitet, reaksjon fartøyet størrelse og relative antall celle ekstrakt og magnesium ion i reaksjonen. Vi har funnet DNA kvalitet å ha bemerkelsesverdige parti til parti variant. For å løse dette problemet, anbefales det at brukere rense via en midi eller maxi prep, etterfulgt av et ekstra DNA rengjøring trinn enten på DNA rensing kolonnen brukes i maxiprep eller etter rensing med en ekstra DNA opprydding kit. Dette forbedrer reproduserbarhet i DNA kvalitet for CFPS reaksjoner og resulterer i mer robust protein produksjon (figur 2B). Reaksjonen fartøyet også påvirker volumetriske avkastning, slik at protein produksjon av identiske reaksjon oppsett i varierende fartøyet volumer kan variere opp til 40%. Det har blitt sagt at økning i i volumetriske avkastning i større fartøy skyldes en økt areal reaksjonsblandingen, tillater for bedre oksygen exchange (figur 2C), og andre har ytterligere styrket volumetriske avkastning ved å kjøre CFPS reaksjoner i store flat bunn plater, som vi anbefaler for reaksjoner over 100 µL17,31,37,52.

Figur 1 : Tidslinje for kultur vekst, produksjon av cellen ekstrakt, oppsett og kvantifisering av CFPS reaksjoner. Brukeren kan implementere CFPS plattformen for forskning programmer gjennom denne fire dagers arbeidsflyten. Forberedelse av reagenser representerer primære tid og kostnader investeringen i første runde av dette eksperimentet og reduserer betydelig etter reagenser aksjer er etablert. I tillegg kan celle pellets og forberedt celle ekstrakt lagres i over et år på-80 ° C, tillater brukeren å begynne tidslinjen trinnene for raskere resultater. Brukeren kan også pause på trinnene endre tidslinjen i denne arbeidsflyten. Klikk her for å se en større versjon av dette tallet.

Figur 2 : Kan endres forhold for CFPS og virkningene på volumetrisk reaksjonen gir. A. pakke produktivitet sammenligning basert på høsting BL21(DE3) cellene på ulike OD600 målinger. Basert på denne tomten, anbefaler vi høsting på et OD600 3,3 å produsere minst 1000 µg/mL målet protein. Reaksjoner ble utført på 15 μL skala i 1,5 mL microfuge rør. B. sammenligning av to DNA maxiprep vask protokoller med og uten etter rensing DNA-opprydding. pJL1-sfGFP plasmider gjennomgikk en maxiprep med en eller to vasker etterfulgt av en etter rensing opprydding av PCR rensing kit. For å oppnå ~ 900 µg/mL av protein uttrykk, foreslår vi utfører en etter rensing DNA opprydding uavhengig av antall maxiprep vasker. Reaksjoner ble utført på 15 μL skala i 1,5 mL microfuge rør. C. 15 µL CFPS reaksjoner utført i ulike fartøy mellom 2 mL 0,6 mL microfuge rør. "Neg" representerer en negativ kontroll der ingen DNA mal ble lagt til reaksjonen. Alle feilfelt representerer 1 standardavvik av tre uavhengige reaksjoner for hvert vilkår, hver ble kvantifisert i tre eksemplarer. Klikk her for å se en større versjon av dette tallet.

Figur 3 : Nøkkel fremgangsmåter for oppsett og resultater for oppretting av produktiv ekstrakt. A. riktig oppsett av sonication is vannbad å sikre kjøling av prøven mens varme genereres under sonication. B. 1,5 mL microfuge rør som inneholder resuspended celle pellet før (til venstre) og post (høyre) sonication. Den resulterende lysate skal vise en mørkere nyanse sammenlignet resuspended celle pellets. C. riktig separasjon av nedbryting og pellets cellen lysate etter 18 000 x g sentrifugering. D. CFPS reaksjoner etter 4 h med inkubering på 37 ° C. 1,5 mL microfuge rør til høyre (vellykket reaksjon) viser synlig fluorescens sfGFP reporter protein ~ 900 μg/mL. Negativ kontroll røret på venstre, mangler mal DNA og simulerer en mislykket reaksjon, viser en klar løsning med ingen fluorescens. Klikk her for å se en større versjon av dette tallet.

Figur 4 : Endring i protein uttrykk 5 fryse-Tin sykluser for CFPS pakke. Ekstra forberedt fra samme vekst gjennomgikk fem fryse tine sykluser via flytende nitrogen flash fryse etterfulgt av tining på is. Ingen vesentlige endringer i ekstrakt produktivitet for å uttrykke sfGFP ble sett over fem fryse-Tin sykluser. Reaksjoner ble utført på 15 μL skala i 1,5 mL microfuge rør. "Neg" representerer en negativ kontroll der ingen DNA mal ble lagt til reaksjonen. Alle feilfelt representerer 1 standardavvik av tre uavhengige reaksjoner for hvert vilkår, hver ble kvantifisert i tre eksemplarer. Klikk her for å se en større versjon av dette tallet.

Figur 5 : CFPS for reaksjoner med varierende [Mg2+] og ekstra volumer versus [sfGFP]. [Mg2 +] varierte fra 8 mM til 14 mM med 2 mM trinn og ekstra volumer varierte fra 3 µL til 7 µL med 1 µL trinn. Fargen koden representerer [sfGFP] produsert fra høy (rød) til lav (lilla). For å maksimere reagens effektivitet samtidig opprettholde høy protein produksjon, anbefaler vi å bruke 5 µL av ekstrakt og 10 mM Mg2+ for ekstrakter som har et totalt proteininnhold på ~ 30 mg/mL, som bestemmes av Bradford analysen. Opprinnelige poeng å generere kontur handlingen var basert på sluttpunktet fluorescens tre uavhengige reaksjoner for hvert vilkår, hver ble målt i tre eksemplarer. Reaksjoner ble utført på 15 μL skala i 1,5 mL microfuge rør. Klikk her for å se en større versjon av dette tallet.
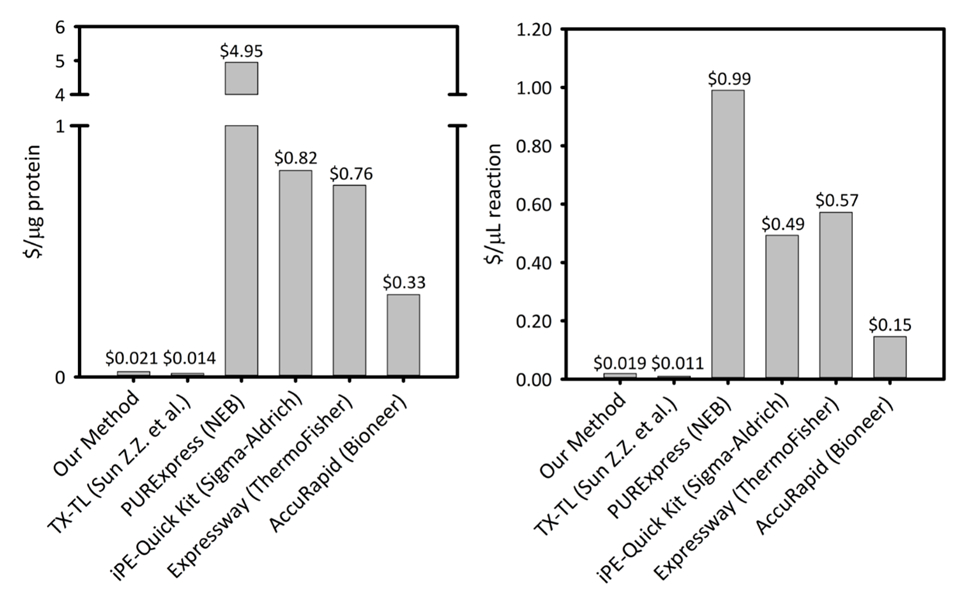
Supplerende figur 1: koster per mikrogram protein produsert og per mikroliter av reaksjon på seks celle-fri protein syntese plattformer. Vår plattform sammenlignes fem forskjellige cellen gratis protein syntese kits/plattformer med varierende produktivitet og priser. Vår sonication basert CFPS plattform er mer kostnadseffektive i begge $/ µg protein og $/ µL reaksjon enn de fleste kommersielle kits og gir enkel en kit for reaksjon oppsett, mens Gjenstående kostnader sammenlignes med andre akademiske CFPS plattformer. Klikk her for å se en større versjon av dette tallet.
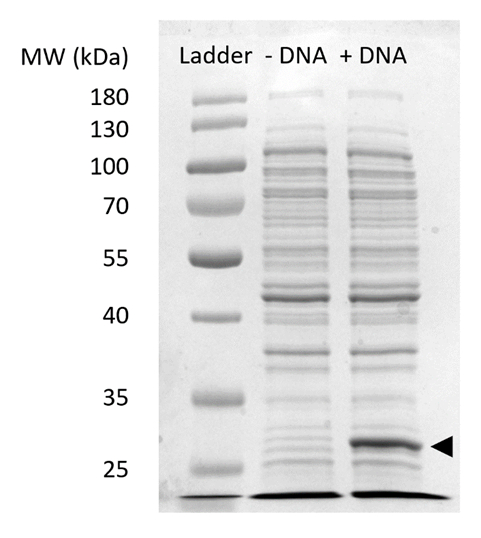
Supplerende figur 2: SDS-siden på sfGFP uttrykk i CFPS. Cellen gratis protein syntese reaksjoner med (+ DNA) og uten (-DNA) DNA mal for sfGFP ble kjørt på en 12% SDSPAGE gel å demonstrere uttrykk for sfGFP observert på 27 kDa (svart pil). Tradisjonelle SDS side teknikker ble brukt. Hvert utvalg lastet inn gel inkludert 18 µg totale protein basert på Bradford analysen kvantifisering totale protein i cellen pakke. Basert på fluorescens intensitet mål og vår standardkurve, vi anslå verdi det av "+ DNA" lane inneholder 0.42 µg av sfGFP. For å få disse prøvene, ble CFPS reaksjoner kjørt på 15 µL skala i 1,5 mL microfuge rør produsere volumetriske gir samsvar med Figur 3 c. Klikk her for å se en større versjon av dette tallet.
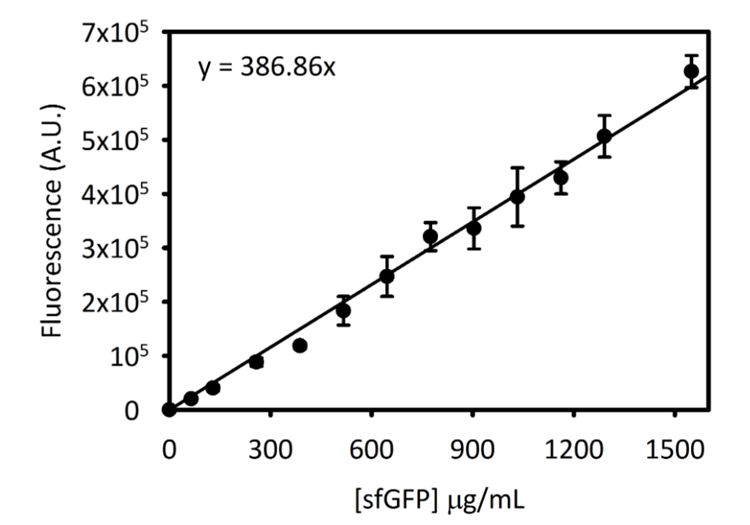
Supplerende Figur 3: standardkurven for sfGFP på Cytation 5. Denne kurven ble bestemt ved hjelp av metodene beskrevet ovenfor. Alle data samlet inn for dette manuskriptet ble konvertert fra endepunktet fluorescens målingene til [sfGFP] i µg/mL bruker denne standardkurve. Klikk her for å se en større versjon av dette tallet.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Cellen gratis proteinsyntese har dukket opp som en kraftig og aktivere teknologi for en rekke applikasjoner, fra biomanufacturing til rask prototyping biokjemiske systemer. Bredden av programmer støttes av evnen til å overvåke, manipulere og øke cellulære maskiner i sanntid. Til tross for den voksende effekten av denne plattform-teknologi, bred tilpasning har vært langsom på grunn av tekniske nyansene i gjennomføringen av metodene. Gjennom dette arbeidet, som mål å tilby enkelhet og klarhet for å etablere denne teknologien i nye labs. Mot dette formål, våre Protokoll for en E. coli-basert celle-fri protein syntese plattformen kan oppnås innen oppstartstid av fire dager ved laboratoriet trent ikke-eksperter, som studenter (figur 1). I tillegg når et lager av reagenser og ekstrakt er produsert, kan påfølgende CFPS satsvise reaksjoner være definert ruges og kvantifisert i bare 5 h. Én 1 L cellevekst kan resultere i nok ekstra for fire hundre 15 µL CFPS reaksjoner, enkelt satsvis forberedelser av andre celle-fri reagensene kan støtte tusenvis av reaksjoner. Reagens preparater kan også skaleres hvis en enda større lager er nødvendig. CFPS reaksjoner kan være oppsett på en høy gjennomstrømming måte, ved hjelp av en 96-brønns plate eller PCR rør for testing av en rekke forhold parallelt. Volumetrisk gir reduseres når du bruker mindre fartøyer som vist i figur 2C. CFPS reaksjoner kan også skaleres fra microliters titalls ml av totale reaksjon for å øke den totale protein avkastningen for én enkelt betingelse. Når skalere opp volum, er den primære vurderingen at volumetriske reaksjonen gir redusere som overflate område-til-volum forholdet mellom reaksjonen senker37,52. For å skalere opp samtidig lignende volumetriske gir protein uttrykk, bør brukere dele reaksjon volumet i mange reaksjon fartøy eller øke størrelsen på fartøyet. For reaksjon vekter fra 15 µL - 100 µL i volum, anbefales mange 15 µL reaksjoner parallelt. Flat-24-og bunnplater anbefales for reaksjoner overstiger 100 µL i volum, og 12-vel plater anbefales for reaksjon volumer over 600 µL. Slike motstandere reaksjon volumer og fartøy gir konsistens volumetriske reaksjonen gir ved oppskalering17,31,37,52. Skalere opp utover disse volumene kan oppnås ved å benytte flere brønner av platen parallelt. Med dette formatet, kan reaksjonen skaleres til over 10 mL totalt volum. Optimalisere reaksjon volum-reaksjon fartøyet kombinasjonen kan støtte programmer som biomanufacturing uten å ofre produktiviteten i reaksjonen.
Når du utfører denne protokollen, er det noen viktige hensyn som påvirker volumetriske reaksjonen gir samt indikatorer forbundet med dårlige resultater ekstrakt. For å sikre riktig lyse og hindre denaturering av funksjonelle transkripsjon/oversettelse maskiner, er det viktig å redusere heten produsert under lysis. Fordype celle rørets i en isen vannbad under sonication å raskt spre varmen under sonication (figur 3A). Indikator for effektiv celle lysis er en mørkere utseende i cellen lysate forhold til pre-sonicated prøver (figur 3B). Brukeren fleksibilitet er sonicator og probe vist i figur 3A tilpasningsdyktige for volumer fra 100 µL til 15 mL av resuspended celler. Dette kan brukeren justere antall joule levert lysis av ønsket antall celler. I tillegg kan større mengder ekstrakt tilberedes gjennom to supplerende tiltak. Brukere kan sonicate flere rør parallelt, eller sonicate større mengder celle rørets, skalering mengden energi proporsjonalt med volumet som beskrevet tidligere29,45. Et skritt som angir ekstra kvalitet er sentrifugering skritt etter celle lysis. Innlegg celle lyse, vi anbefaler sentrifugering 18.000 x g å gi et klart skille mellom nedbryting (transkripsjon/oversettelse maskiner, fragmenterte genomisk DNA som ikke lenger fungerer mal transkripsjon/oversettelse) og pellets ( uønsket cellulære komponenter som cellemembranen og utfelt proteiner) (Figur 3 c). Vi har funnet at sentrifugering på 18 000 x g forbedrer separasjon, noe som resulterer i bedre reproduserbarhet sammenlignet spinn ved lavere hastigheter som 12.000 x g. For bekvemmelighet anbefaler vi en bord nedkjølt sentrifuge, oppnå et minimum av 12.000 x g. Dette trinnet er også utført på 30.000 x g, som bør vurderes hvis utstyr er tilgjengelig54,55,56,57,58, 59 , 60. ekstra ytelsen påvirkes ikke av sentrifugering hastigheter på dette trinnet gitt at riktig separasjon er oppnådd. Når du fjerner ønsket nedbryting, er det best å unngå skyet materiale som finnes på grensen mellom nedbryting og pellets siden denne forurensningen vil redusere produktiviteten av ekstraktet. Satsing for renhet nedbryting av fører til mer produktive ekstrakter og er verdt redusert antall pakke hentes for nye brukere.
Det er viktig å merke til at mens metodene vi har presentert er reproduserbare og kan utføres av forskere med minimal kompetanse, kan det være parti til parti og reaksjon-til-reaksjon variasjon. Dette kan tilskrives variasjon i proteomic sammensetningen av lysate etter sonication61. Parti til parti variasjon som vi har observert er vanligvis redusert tilskudd med T7RNAP og optimalisering av magnesium konsentrasjoner. Eksogene tillegg av T7RNAP er vanlig blant CFPS reaksjoner å støtte optimal protein uttrykk, og vi finner at har to kilder til T7RNAP - endogene uttrykk i BL21*(DE3) og den ekstra T7RNAP å en final konsentrasjon av 16 µg/mL - forbedrer parti til parti reproduserbarhet for nye brukere45,46. Med erfaring, kan brukerne endre sine eksperimenter for å bruke bare én kilde av T7RNAP hvis ønsket. Kvantifisering av totale proteininnholdet i en ny gruppe med ekstrakt og riktig justering av Mg2 + konsentrasjon kan også bidra til å redusere parti til parti variant volumetriske protein uttrykk gir. Variasjoner i protein uttrykk kan også være forskjeller i størrelsen og strukturen av proteinet rundt, codon bruken av genet og dens tilsvarende ribosom binding området av genet av interesse, i tillegg til typen uttrykk vektor brukes62 ,63. For disse grunner, kan noen proteiner ikke uttrykke og modell protein sfGFP, som resulterer i redusert volumetriske avkastning fra CFPS reaksjoner.
Begrensninger av presentert CFPS teknikken omfatter at det ikke kan være egnet for alle programmer av celle-fri, som metabolske engineering og tuning av uttrykket forhold, uten ytterligere modifiseringer til protokoller. Vi tror imidlertid at denne protokollen vil gi grunnlag for å etablere CFPS plattformen i nye laboratorier og gi ikke-eksperter kan implementere innledende celle-fri reaksjoner i labs. Etter første implementeringen, kan forskere eksperimentere med plattformen å gjøre egne endringer for mer spesifikke programmer basert på annen litteratur i feltet.
CFPS plattform kostnader $0.021/µg protein (unntatt arbeidskraft og utstyr), gjør systemet konkurransedyktig med kommersielle kits uten at enkel reaksjon oppsett. Vurderinger av komparative kostnadene per µL av reaksjonen viser lignende trender (Supplemental figur 1). Vi anslår oppstart kostnader å være ~ $4500 for reagenser, og en ekstra $3200 for spesialisert utstyr, som en sonicator. Arbeidstimer å gjennomføre denne prosedyren er anslått til ~ 26 h for alt reagens prep fra grunnen opp. Men når store bestander av reagenser er utarbeidet, avta krav til arbeidskraft betydelig. I tillegg som erfaring med plattformen er oppnådd, vi anbefaler skalere opp størrelsen på celle-vekst, ekstra forberedelser og forberedelse av reagenser å maksimere tiden effektiviteten. Gitt de oppstart kostnadene, vi anbefaler CFPS plattformen for programmer i syntetisk biologi, høy gjennomstrømming innsats, og protein uttrykk betingelser som er kompatible med tradisjonelle protein uttrykk plattformer grunn til konflikt med cellens biokjemi og levedyktighet begrensninger. I disse spesialiserte tilfeller der ønsket teknikken er aktivert av CFPS plattformen er større kostnadene ved CFPS over i vivo uttrykk begrunnet.
Videreutvikling av CFPS plattformen er sannsynlig å gi bredere verktøyet til bioteknologi innsats som metabolske prosjektering av enzymatisk veier, produksjon og karakterisering av tradisjonelt uløselige proteiner, aminosyre innlemmelse og unaturlig protein uttrykk, lagdelt medisin produksjon og utvide utover laboratoriet til klasserommet for STEM utdanning64,65,66. Dette arbeidet vil bli videre støttet av det pågående arbeidet for detaljert karakterisering av CFPS plattformen. En bedre forståelse av sammensetningen av cellen ekstrakt vil føre til fortsatte finpussingen mot forbedret reaksjonen gir og fleksibilitet i reaksjonen forhold61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne erklærer at de har ingen konkurrerende økonomiske interesser eller andre interessekonflikter.
Acknowledgments
Forfattere ønsker å erkjenne Dr. Jennifer VanderKelen, Andrea Laubscher, og Tony Turretto kundestøtte, Wesley Kao, Layne Williams og Christopher Hight for nyttig diskusjoner. Forfatterne også erkjenner finansiering støtte fra Bill og Linda Frost fondet, senter for programmer i Bioteknologis Chevron bioteknologi brukt legat forskningsstipend, Cal Poly forskning, Scholarly og kreative aktiviteter Grant Program (RSCA 2017), og National Science Foundation (NSF-1708919). MZL erkjenner California State University Graduate stipendet. MCJ erkjenner den Army Research Office W911NF-16-1-0372, gir National Science Foundation kalt MCB-1413563, kalt MCB-1716766 og den Luftforsvaret forskning laboratorium Center for Excellence Grant FA8650-15-2-5518, forsvar trussel reduksjon Agency stipendet HDTRA1-15-10052/P00001, David and Lucile Packard Foundation, Camille Dreyfus lærer-forsker programmet, Institutt for energi BER Grant DE-SC0018249, programmet for menneskelig grenser Science (RGP0015/2017), DOE Joint Genome Institute ETOP Grant, og Chicago biomedisinsk konsortiet med støtte fra Searle midlene i Chicago samfunnet Trust for støtte.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}