

Summary
Этот протокол подробно шаги, расходы и оборудование, необходимое для создания E. coli-на основе клеточных экстрактов и реализовать в vitro реакции синтеза белка в течение 4 дней или меньше. Чтобы использовать гибкий характер этой платформы для широкого применения, мы обсуждаем условий реакции, которые могут быть адаптированы и оптимизированы.
Abstract
За последние 50 лет свободных клеток синтез белка (СЛТ) стала мощной технологии осваивать транскрипционный анализ и трансляционная потенциала клеток в пробирке. Устраняя необходимость поддержания жизнеспособности клетки и устраняя сотовой барьер СЛТ был основополагающим для новых приложений в biomanufacturing традиционно сложных белков, а также приложений в быстрого прототипирования для Метаболический инжиниринг и функциональной геномики. Наши методы для реализации E. coli-платформе СЛТ позволяют новым пользователям для доступа к многим из этих приложений. Здесь мы описываем методы для подготовки экстракт обогащенного СМИ, недоумение колбы и воспроизводимый метод лизис перестраиваемый на основе sonication клеток. Этот экстракт может затем использоваться для выражения протеина, способных производить 900 мкг/мл или более супер папку зеленого флуоресцентного белка (sfGFP) в как раз 5 h от экспериментальной установки для анализа данных, учитывая, что соответствующий реагент запасы были подготовлены заранее. По оценкам запуска получения реагентов стоит $4500, которая будет поддерживать тысячи реакций по ориентировочной цене $0.021 за мкг белка, произведенного или $0.019 за мкл реакции. Кроме того методы выражения протеина зеркало легкость установки реакции, видели в коммерчески доступных систем за счет оптимизации готовые смеси реагентов, на долю от стоимости. Чтобы разрешить пользователю использовать гибкий характер СЛТ платформы для широкого применения, мы определили целый ряд аспектов платформы, которые могут быть настроены и оптимизированы в зависимости от имеющихся ресурсов и результатов выражения протеина желаемого.
Introduction
Бесплатный мобильный синтез белка (СЛТ) стала технология, которая разблокирована ряд новых возможностей для производства белка, функциональной геномики, метаболический инжиниринг и более в течение последних 50 лет1,2. По сравнению с стандартным в vivo белка выражение платформ, СЛТ предоставляет три основных преимущества: 1) клетки бесплатный характер платформы позволяет производство белков, которые бы потенциально токсичных или иностранных в ячейке3,4 ,5,6; 2) инактивации геномной ДНК и внедрение шаблона ДНК кодирования gene(s) интерес канал все системные энергии в рамках реакции на производство белки интерес; и 3) открытый характер платформы позволяет пользователю изменять и контролировать условия реакции и композиции в режиме реального времени7,,8. Такой непосредственный доступ к реакции поддерживает увеличение биологических систем с расширенной химия и окислительно-восстановительных условий для производства новых белков и тюнинг метаболических процессов2,9, 10. прямого доступа также позволяет пользователю комбинировать СЛТ реакции с анализов деятельности в системе одного горшок для более быстрого дизайн строить тест циклов11. Способность выполнять СЛТ реакции в малый объем капли или устройств на базе бумаги далее поддерживает усилия обнаружения высокой пропускной способностью и быстрое прототипирование12,13,14,15 ,16. В результате эти преимущества и подключи и играй характер системы, СЛТ однозначно включен разнообразные применения биотехнологии такие как производство белков, которые трудно solubly Экспресс в vivo17, 18,19,20, обнаружение заболевания21,22,23, по требованию biomanufacturing18,24 ,25,26,27и образование28,29, все из которых проявить гибкость и полезность платформы клетки бесплатно.
СЛТ систем могут быть сгенерированы из различных сырой лизатов от обеих линий клеток прокариот и эукариот. Это позволяет для различных вариантов в системе выбор, каждая из которых имеет свои преимущества и недостатки в зависимости от интереса. СЛТ систем также различаются по времени приготовления, стоимости и производительности. Наиболее часто используются ячейки, что экстракты производятся из зародышей пшеницы, кролик ретикулоцитов, насекомое клетки и клетки Escherichia coli , с бытием наиболее экономически эффективных на сегодняшний день при производстве высокий объемный урожайность белка30 . В то время как другие СЛТ системы может быть выгодным для их врожденной столб-поступательные изменения техники, новых приложений с помощью кишечной палочки-на основе техники способны преодолеть разрыв, создавая site-specifically фосфорилированных и гликозилированного белков на спрос31,,3233,34,35.
СЛТ реакции могут выполняться в любом пакете, непрерывный обмен клетки бесплатно (CECF) или непрерывный поток свободных клеток (CFCF) форматы. Формат пакета является закрытой системой, время жизни которого реакции ограничена из-за уменьшения количества реактивов и накопление ингибирующее побочные реакции. Методы CECF и CFCF увеличить время жизни реакции и тем самым привести к повышенной объемной белка урожай по сравнению с пакетной реакции. Это достигается путем предоставления побочных продуктов синтеза белка необходимо удалить из реакции судна, в то время как новые реактивы поставляются в ходе реакции2. В случае CFCF протеин интереса также могут быть удалены из реакционной камеры, в то время как в CECF, протеин интереса остается в реакционной камере, состоящая из36,полупроницаемой мембраны37. Эти методы являются особенно ценными в преодолении бедных объемные урожайности трудно Экспресс белков интерес38,39,40,,4142, 43. Проблемы в реализации CECF и CFCF подходов, что 1) в то время как они приводят к более эффективному использованию механизма био ответственных за транскрипции и трансляции, они требуют особенно больших количеств реагентов, что увеличивает общую стоимость и 2) они требуют более сложных установок реакции и специализированного оборудования, по сравнению с пакетный формат44. Чтобы обеспечить доступность для новых пользователей, протоколы описаного фокус на пакетный формат в реакционных объемов 15 мкл с конкретными рекомендациями для увеличения объема реакции на миллилитр масштаба.
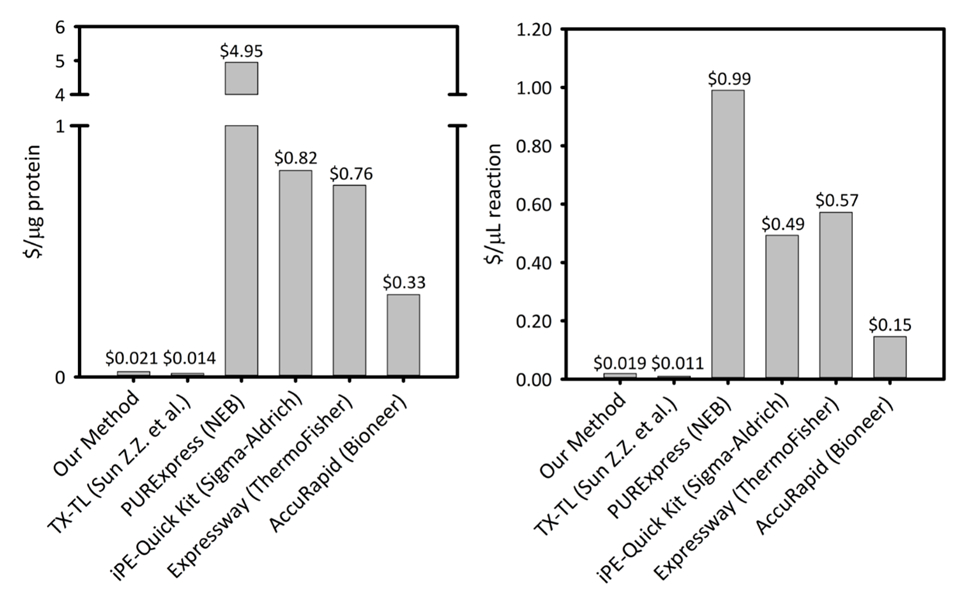
Представленные здесь методы позволяют реализовать рост клеток, извлекать подготовки и пакетная установка реакции формат для е. coliнеспециалистов с основных лабораторных навыков (например, студентов)-на основе системы СЛТ. Этот подход является экономически эффективным по сравнению с коммерчески доступные наборы без ущерба для легкости установки комплекта основе реакции. Кроме того этот подход позволяет приложениям в лаборатории и на местах. При принятии решения о реализации СЛТ, новых пользователей следует тщательно оценить эффективность системах выражения протеина обычных для запуска инвестиций, поскольку СЛТ не могут быть выше в каждом конкретном случае. Описанные здесь методы СЛТ позволяют пользователю напрямую реализовать целый ряд приложений, в том числе функциональной геномики, испытания, производство белков, которые являются неразрешимыми в vivo выражение, а также поле высокой пропускной способности приложения, включая биодатчики и учебных комплектов для синтетической биологии. Дополнительные приложения, такие как метаболический инжиниринг, тюнинг белка выражение условий, обнаружение заболеваний, и масштабирования с помощью методов CECF или CFCF по-прежнему возможны, но может потребовать опыта с платформой СЛТ для дальнейшего изменения реакции условий. Наши методы сочетают в себе рост обогащенного СМИ и озадачен колбы, с относительно быстрым и воспроизводимые методами лизис клеток через sonication, а затем упрощенной установки реакции СЛТ, которая использует оптимизированные премиксы45. Хотя методы клеточного роста стали несколько стандартизированы в этой области, методы для лизиса клеток варьируются в широких пределах. В дополнение к sonication общие лизис методы включают использование французской прессе, гомогенизатор, шарик загонщиков, или лизоцима и другие биохимические и физические нарушения методы46,47,48, 49. с помощью наших методов, примерно 2 мл экстракт сырой клетки получены на 1 Л клеток. Это количество клеток экстракт может поддерживать четыре сотни 15 мкл СЛТ реакции, каждый производства ~ 900 мкг/мл репортер sfGFP белка из шаблона плазмида pJL1-sfGFP. Этот метод по цене $ 0,021/мкг производства sfGFP ($.019/мкл реакции), за исключением стоимости труда и оборудования (Дополнительные рис. 1). Начиная с нуля, этот метод может быть реализован в течение 4 дней одного человека и повторять что СЛТ реакции может быть завершена в течение часов (рис. 1). Кроме того протокол может масштабироваться объема для больших партий реагента подготовки в соответствии с потребностями пользователя. Важно протокола, представленные здесь может быть реализован специалистами лаборатории обучены не-такие как студентов, как он только требует основных лабораторных навыков. Процедуры, описанные ниже и сопровождающие видео были разработаны специально для повышения доступности E. coli СЛТ платформы для широкого использования.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. средства массовой информации подготовка и рост клеток
-
1 день
- Полоска E. coli BL21*(DE3) клетки от глицерина складе на плиту LB агар и проинкубируйте по крайней мере в 18 ч при 37 ° C.
- Подготовка 50 мл LB СМИ и автоклав решение на жидких цикла в течение 30 мин при температуре 121 ° C. Хранить при комнатной температуре.
-
День 2
- Подготовьте 750 мл 2 x YTP СМИ и 250 мл раствора 0.4 М Д-глюкозы, как описано в дополнительной информации.
- Наливайте 2 x YTP СМИ в газобетона 2,5 Л озадачен колбу и D-глюкозы раствор в стеклянной бутылке газобетона 500 мл. Автоклав оба решения на основе жидкого цикла для 30 мин при температуре 121 ° C.
- Убедитесь, что оба стерильные растворы хранятся при 37 ° C, если рост клеток выполняется на следующий день, чтобы увеличить темпы роста после прививки. Не следует сочетать решения до прививки.
Примечание: Решения могут храниться при температуре 4 ° C для 1-2 d при необходимости, хотя 2 СМИ x YTP чрезвычайно подвержен загрязнению. - Запустите на ночь культуры BL21(DE3), прививки 50 мл с одной колонии BL21(DE3), с помощью цикла стерилизации и стерилизации во избежание загрязнения LB СМИ.
- Место 50 мл BL21*(DE3) LB культуры в 37 ° C 250 об/мин, пожимая инкубатора и расти на ночь для 15-18 ч.
- Подготовить и простерилизуйте все материалы, необходимые для дней 3 и 4, в том числе: две бутылки 1 Л центрифуги, 4 x холодной 50 мл конические трубы (весят и записи масс трех) и многие 1.5 мл пробирок отцентрифугировать.
-
День 3
- Удаление 50 мл на ночь культуры BL21*(DE3) в LB от тряски инкубатора и измерить ОД600 на спектрофотометр с помощью 1:10 разрежения с LB СМИ. Рассчитать объем ночь культуры необходимо добавить 1 Л средств массовой информации для начала ОД600 0.1 (к примеру, если ОД600 1:10 разрежения читается как 0.4, прививать 25 мл неразбавленного ОД600 = 4.0 ночь культуры в 1 Л 2 x YTP G).
- Удаление утепленные 2 x YTP средства массовой информации и решения D-глюкозы из инкубатора 37 ° C наряду с 50 мл LB культуры. Тщательно используя стерильную методику, Залейте раствор Д-глюкозы в 2 x YTP СМИ (избегая сторон озадачен колбу).
Примечание: Добавление D-глюкозы завершает рецепт для 1 Л 2 x YTPG. - Поддержании стерильных, прививать 1 Л раствора 2 x YTPG с соответствующим объемом 50 мл культуры начать 1 Л культуры 0.1 ОД600. Немедленно переведите привитых культуры 1 Л в 37 ° C, пожимая инкубатор на 200 об/мин.
- Возьмите первый ОД600 чтения после первого часа роста (запаздывание фазы типичный занимает 1 час). Не разбавлять культуры. Продолжить, замеры ОД600 примерно каждые 20-30 мин до600 ОД 0,6.
- По достижении ОД600 = 0,6, добавляют 1 мл 1 м IPTG (конечная концентрация в культуре 1 Л = 1 мм) 2 x YTPG культуры.
Примечание: Идеальный индукции ОД600 -0,6; Однако широкий спектр 0,6-0,8 является приемлемым. Индукции с IPTG — для внутреннего производства T7 РНК-полимеразы (T7RNAP). - После индукции Измерьте ОД600 примерно каждые 20-30 мин до тех пор, пока он достигает 3.0.
Примечание: Охлаждения центрифуги до 4 ° C в течение этого времени. Подготовка холодной S30 буфера, как подробно указано в дополнительной информации. Если буфер S30 готовится заранее, убедитесь, что DTT не добавляется до дня использования. - Как только ОД600 достигает 3.0 (рисунок 2A), залейте культуры в холодной центрифуги бутылка 1 Л в ледяной воде ванны. Подготовьте заполненные водой бутылка центрифуге 1 Л равный вес использоваться как баланс в центрифуге.
Примечание: Значения поглощения варьируются от инструмент инструмент. Хотя ОД600 урожай BL21(DE3) не является чувствительным переменной, рекомендуется, что пользователь оценивать и оптимизировать эту переменную как мера устранения неполадок. Больших спектрофотометры может привести к относительно низкой ОД600 чтений по сравнению с небольших основанных на кювету спектрофотометры. - Центрифуга бутылки 1 Л за 10 мин на 5000 x g и 10 ° C до Пелле клетки.
- Медленно слить супернатант и утилизируйте его в соответствии с института биологических отходов процедур. Место гранул на льду.
- С помощью стерильных шпателя, скрести клетки гранулы из бутылки центрифуги и перенести его на холодной 50 мл Конические трубки.
- Добавьте 30 мл холодного S30 буфера Конические трубки и Ресуспензируйте Пелле клеток, vortexing с короткими очередями (20-30 сек) и периодов отдыха (1 мин) на льду до тех пор, пока полностью высокомобильна с не куски.
- После того, как гранулы полностью высокомобильна, используйте другой 50 мл Конические трубки с водой как баланс и центрифуги для 10 мин на 5000 x g и 10 ° C (предварительно охлаждается до 4 ° C).
Примечание: Это завершает 1st 3 моет, необходимых при уборке клетки. - Вылейте супернатант и утилизируйте его в соответствии с института биологических отходов процедур. Ресуспензируйте лепешка с 20-25 мл холодного S30 буфера и центрифуги для 10 мин на 5000 x g и 10 ° C (предварительно охлаждается до 4 ° C).
Примечание: Это завершает 2nd 3 стирок. - Опять же вылейте супернатант и утилизируйте его в соответствии с института биологических отходов процедур. Добавьте ровно 30 мл буфера S30 и вихрь снова, чтобы Ресуспензируйте гранулы.
- Использование 3 предварительно взвешенный, холодная 50 мл конические трубы и Серологические Пипетки наполнитель с стерильной пипеткой, кратные 10 мл ресуспензированы гранул/S30 буферной смеси в каждой из 3 конические трубы.
Примечание: Разделение клетки на 3 трубы не требуется, но этот шаг приводит к меньше ячейка окатышей (~ 1 g) для повышения удобства в последующих шагах. - Центрифуга все трубы, используя соответствующие остатки, при необходимости, за 10 мин на 5000 x g и 10 ° C (предварительно охлаждается до 4 ° C).
Примечание: Это завершает шаг окончательной стирки. - Вылейте супернатант и утилизируйте его в соответствии с института биологических отходов процедур. Удалите избыток S30 буфера, тщательно протирая внутри конические пробки и крышки с чистой ткани; не прикасайтесь гранулы.
- Reweigh трубы на аналитический баланс и запишите вес окончательной гранул на каждой трубе.
Примечание: Протокол может быть приостановлена на данный момент. Гранулы может быть заморожен в жидком азоте и температуре-80 ° C на срок до одного года, пока она не понадобится для приготовления экстракта.
2. сырой сотовый экстракт подготовка - день 4
- Для приготовления экстракта держите клетки холодной на льду во время каждого шага. Добавьте 1 mL холодной S30 буфера на 1 г клеточной массы таблетки. Убедитесь, что Дитиотреитол (DTT) была дополнена S30 буфер до конечной концентрации 2 мм.
Примечание: Остыть microcentrifuge до 4 ° C в течение этого времени. - По vortexing с короткими очередями (20-30 сек) и периодов отдыха (1 мин) на льду до тех пор, пока полностью высокомобильна Ресуспензируйте Пелле клеток. Если ресуспендирования трудно, оставьте окатышей на льду за 30 мин до размораживания.
- Перевод 1.4 мл ресуспензированы клеток на 1,5 мл отцентрифугировать.
- Поместите один 1.5 мл пробирку, содержащую 1.4 мл ресуспензированы клеток в ледяной водой ванну в стакан. Sonicate 45 s на следуют 59 s покинуть на 3 всего цикла, с амплитудой установлен на 50%. Закройте и инвертировать трубы осторожно смешать периоды выкл. В общей сложности доставить 800-900 J энергии каждой трубы отцентрифугировать 1,5 мл, содержащих 1.4 мл ресуспензированы клеток (рис. 3A и 3B).
Примечание: Этот шаг чувствителен к sonicator тип и модель используется и должны быть оптимизированы, если оборудование отличается от перечисленных для этой процедуры. Два взаимодополняющих подхода может использоваться для масштабирования количество экстракта, подготовленные в ходе этого шага: 1) несколько 1,5 мл отцентрифугировать трубы может быть sonicated параллельно, или 2) больших томов может быть sonicated в конические трубы (до 15 мл ресуспендирования клеток в трубу) , масштабирования количества энергии доставлены как описано 29,45. - Сразу же после завершения sonication 4.5 мкл 1 М DTT (дополняющий дополнительные 2 мм DTT) 1.4 мл lysate и перевернуть несколько раз перемешать. Место труб на льду. Повторите шаги 2.4 и 2.5 для любых дополнительных трубок ресуспензированы клеток, прежде чем приступать к центрифугирования.
- Microcentrifuge образцы на 18,000 x g и 4 ° C на 10 минут (рис. 3 c).
- Пипетка супернатант в новой трубки отцентрифугировать 1,5 мл. Не беспокоить гранулы; Это предпочтительнее оставить некоторые супернатант поддерживать чистоту чем нарушить гранулы в усилия по максимизации доходности.
- Инкубируйте супернатант из предыдущего шага в 250 об/мин и 37 ° C 60 мин, лентой трубы для встряхивания платформы инкубатора (это реакция стока).
- Microcentrifuge образцы на 10000 x g и 4 ° C на 10 мин.
- Удалить супернатант не нарушая гранулы и перенести его на новой трубки. Создание многих 100 мкл аликвоты экстракта для хранения.
Примечание: Протокол может быть приостановлена здесь, и экстракт может быть заморожен в жидком азоте и температуре-80 ° C для до одного года, пока она не понадобится для реакций СЛТ. По крайней мере 5 циклов замораживания оттаивания можно прошел без ущерба для извлечения производительности (рис. 4).
3. Сотовый бесплатно белка синтез пакетный формат реакции
- Оттепель решения A и B, ДНК шаблон, BL21(DE3) экстракт (если замороженные), T7RNAP и Алиготе молекулярного уровня воды.
Примечание: СЛТ реакции шаблон можно найти в Дополнительной информации. Рецепты решения A и B приводятся в Дополнительной информации и соответствуют конкретным концентрации для многочисленных реагентов для поддержки PANOx-SP на основе энергетической системы СЛТ. Роль каждого реагента и приемлемым вариации концентраций этих реагентов, поддерживающие СЛТ были определены50. Протокол T7RNAP очистки можно найти в дополнительной информации51. Дополнительные T7RNAP может увеличить объёмный урожайность, но не является необходимым, если T7RNAP индуцируется во время роста клеток. Плазмида ДНК шаблон (pJL1-sfGFP) может быть подготовлен с использованием комплекта maxiprep с двумя моет, используя буфер умывальник в комплекте, следуют постобработки ДНК очистка с помощью ПЦР очистка kit (рис. 2B). Линейная ДНК шаблоны могут также использоваться в реакциях СЛТ. - Ярлык необходимое количество отцентрифугировать трубы, необходимые для реакций СЛТ.
Примечание: Реакции могут быть выполнены в различных размеров судна, однако меньше судно может уменьшить объемные белка урожай (рис. 2 c). Масштабирование реакцию в того же размера судна может также уменьшить объемные урожайности, как функция снижения обмена кислорода, за счет снижения в отношение площади поверхности к тома. При увеличении объема реакции выше 100 мкл, рекомендуется использовать плоское дно хорошо пластин 31,37,52. - 2.2 мкл раствора A, 2.1 мкл раствора B, 5 мкл BL21*(DE3) экстракт, 0,24 мкг T7RNAP (16 мкг/мл конечная концентрация), 0,24 ng шаблона дна (16 конечная концентрация нг/мл), и воды довести окончательный объем до 15 мкл.
Примечание: Во время установки реакции во избежание оседания компонентов и обеспечить, чтобы каждый реакции Vortex решения A и B часто получает однородных Алиготе каждого решения. Избегайте vortexing экстракт, вместо инвертировать трубки смешивать. - После того, как были добавлены все реагенты реакции, mix каждую пробирку закупорить вверх и вниз или слегка vortexing обеспечивая, что окончательный реакционную смесь объединяется в один шарик 15 мкл в нижней части трубки отцентрифугировать 1,5 мл.
- Место каждой реакции в инкубатор 37 ° C без встряхивания за 4 ч или 30 ° C на ночь.
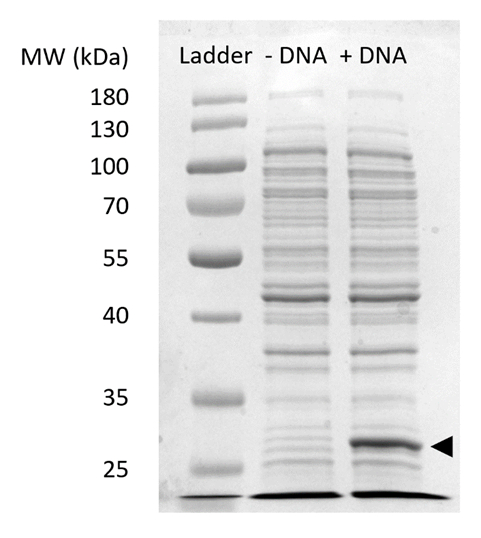
Примечание: Успешное реакции можно качественно оценить визуально основаны на зеленый цвет sfGFP продукта в пределах СЛТ реакционную смесь (рис. 3D). Выражение протеина интереса может быть подтверждена SDS-PAGE (дополнительный рисунок 2).
4. Количественная оценка репортер белка, [sfGFP]
- Загрузите 48 мкл 0,05 М HEPES, рН 8, в каждый хорошо необходимы для количественной оценки (обычно выполняется в трех экземплярах на реакции трубки).
- Удаление реакции из инкубатора. Пипетка вверх и вниз, чтобы смешать каждой реакции, а затем перевести 2 мкл реакции в 48 мкл 0,05 М HEPES, рН 8. Накапайте вверх и вниз снова в хорошо перемешать.
- После того, как все реакции загружаются и смешанные, место пластину 96 хорошо в флуориметр и измерения флуоресценции конечной точки sfGFP.
Примечание: Волны возбуждения и выбросов для квантификации флуоресценции sfGFP находятся 485 Нм и 510 нм, соответственно. - С помощью ранее созданного калибровочной кривой, определите [sfGFP] от полученных флуоресценции чтений.
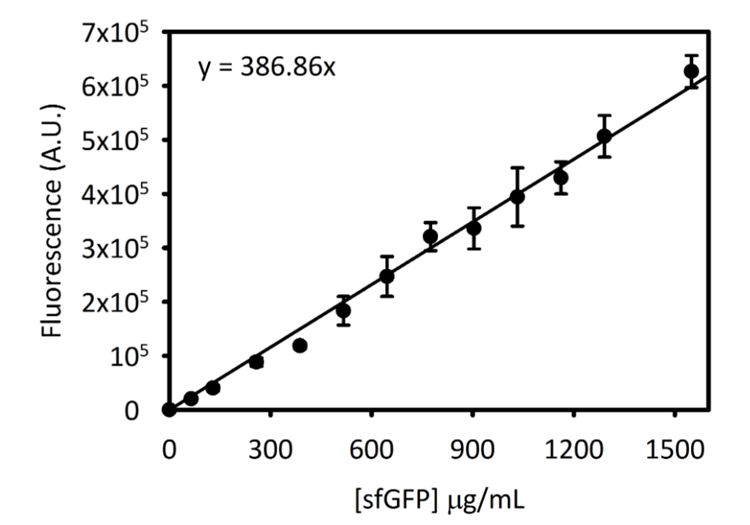
Примечание: Инструкции для создания стандартной кривой концентрации sfGFP против интенсивности флуоресценции приводятся в дополнительной информации (дополнительный рис. 3). Пользователям потребуется установить калибровочной кривой для своего инструмента, поскольку чувствительность инструмента может отличаться.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Мы представили на основе sonication E. coli экстракт подготовки протокола, который может быть завершен в течение четырех дней, с Рисунок 1 демонстрирует процедурные разбивка на каждый день. Есть податливость шаги, которые могут быть завершены в каждый день с различными приостановка очков, но мы нашли этот рабочий процесс будет наиболее эффективным для выполнения. Кроме того клетки окатышей (шаг 1.3.18) и полностью готова экстракт (шаг 2.10) стабильны-80 ° c для по крайней мере год, позволяя пользователю создавать большие запасы каждого, чтобы сохранить для использования в позднее время17. Не только это экстракт стабильное течение длительного времени, но экстракт также можно пройти по меньшей мере пять заморозить циклов оттаивания без значительной потери производительности (рис. 4). Это позволяет для большего аликвоты экстракта храниться для многократного использования, если Морозильная Камера хранения пространство ограничено. Однако мы рекомендуем несколько меньших аликвоты (~ 100 мкл) экстракта, когда это возможно.
С каждой новой экстракт подготовки мы рекомендуем, что пользователь выполняет магния титрования для того, чтобы определить оптимальное количество магния для пакетной выписки. Пользователи могут количественно партии партии изменчивость общего белка концентрацию экстракта клеток в assay Брадфорд. Для выше выполняет экстракты, мы обычно видим концентрации общего белка 30-50 мг/мл, и в этом диапазоне нет очевидных корреляции между концентрациями общего белка и экстракт производительность ячейки. Поэтому мы рекомендуем пользователям настраивать концентрации магния соответственно для обеспечения максимальной белков и нуклеиновых кислот функциональность для каждого пакета экстракт. Уровни магния имеют важное значение для надлежащей репликации, транскрипции и трансляции ДНК, но чрезмерный уровень может быть пагубным для этих процессов53. Чтобы продемонстрировать эту зависимость, мы провели совместное титрования магния и извлекать тома для определения оптимального сочетания, что минимизирует количество экстракта необходимо, сохраняя при этом продуктивной реакции (рис. 5). Из этого эксперимента для мы рекомендуем использовать 5 мкл концентрированного экстракта и 10 мм Mg2 + экстракт с содержанием общего белка 30 мг/мл, для того чтобы получить более 1000 мкг/мл sfGFP.
Наш опыт работы с СЛТ также позволило нам определить шаги в рамках протокола, которые могут быть изменены без ущерба для общей производительности системы и другие, которые являются неотъемлемой частью для высокой исполняющей системы СЛТ. Прежде всего окончательный ОД600 клеток урожая существенно не влияет на конечный результат реакции СЛТ, а клетки реально могут быть собраны где-нибудь от 2.7-4.0 ОД600. Это представляет раннего экспоненциальной фазе роста где рибосома концентрация на ячейку является самым высоким и трансляционного оборудования является наиболее активной поддержки быстрого роста. Это наблюдение позволяет пользователям гибкость для оптимизации своих собственных процедур. Мы рекомендуем что уборки приблизительно 3,0 ОД600 для того, чтобы захватить клетки на од600 ближе к 3.3, время сбора урожая является полным (рисунок 2A). Переменные, которые влияют на урожайность СЛТ включают шаблон ДНК качество, размер судна реакции и относительные количества клеток экстракт и магния иона в реакции. Мы нашли ДНК качества иметь заметные различия партии к партии. Чтобы решить эту проблему, мы рекомендуем пользователям очистить ДНК через Миди или макси prep, следуют дополнительного шага очистки ДНК, либо на столбце очистки ДНК используется в maxiprep, или после очистки с помощью дополнительного комплекта очистки ДНК. Это улучшает воспроизводимость качества ДНК для СЛТ реакций и результаты в более надежного производства белка (Рисунок 2Б). Реакционный сосуд также влияет объемные урожайности, таким образом, что белок производства установок одинаковых реакции в различных томах судно может отличаться до 40%. Он предположил, что импульс в объемном урожайности наблюдается в более крупных судов из-за увеличения площади поверхности реакционной смеси, позволяющие улучшить обмен кислорода (рис. 2 c), и другие дальнейшего повышенный объемный урожайности, запустив СЛТ реакции в большие плоскодонные пластины, которые мы рекомендуем для реакций над 100 мкл17,,3137,52.

Рисунок 1 : График роста культуры, производство выдержки клетки, установки и количественной оценки реакций СЛТ. Пользователь может реализовать СЛТ платформы для их исследования приложений через четыре дня рабочего процесса. Подготовка реагента представляет основного времени и стоимости инвестиций для первого раунда этого эксперимента и существенно уменьшается после того, как создаются запасы реагентов. Кроме того клетки окатышей и подготовленные ячейки экстракт может храниться более года в-80 ° C, позволяя пользователю начать временной шкалы на различные шаги для более быстрых результатов. Пользователь может также приостановить на различные шаги, чтобы изменить сроки этого рабочего процесса. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 2 : Изменяемые условия для СЛТ и воздействие на объемные реакция урожайности. A. извлечение сравнения производительности на основе сбора урожая BL21(DE3) клеток на различных чтений600 ОД. Основываясь на этом участке, мы рекомендуем, уборки в ОД600 3.3 производить по крайней мере 1000 мкг/мл целевого белка. Реакции были выполнены в масштабе 15 мкл в 1,5 мл пробирок отцентрифугировать. Б. сравнение двух протоколов мыть maxiprep ДНК с и без после очистки ДНК очистке. pJL1-sfGFP плазмид претерпел maxiprep с одним или двумя моет, следуют после очистки очистки, очистки комплект ПЦР. Для достижения ~ 900 мкг/мл экспрессии белков, мы предлагаем, независимо от количества maxiprep смывки ДНК после очистки очистки. Реакции были выполнены в масштабе 15 мкл в 1,5 мл пробирок отцентрифугировать. C. 15 мкл СЛТ реакции выступал в различных судов от 2 мл до 0,6 мл отцентрифугировать трубы. «Отрицательная» представляет отрицательный контроль, где шаблон не ДНК был добавлен к реакции. Все планки погрешностей представляют 1 стандартное отклонение трех независимых реакций для каждого условия, каждый из которых был количественно в трех экземплярах. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 3 : Ключ процедурных установок и результаты для создания продуктивных экстракт. A. правильной установки sonication ледяной воде ванны для обеспечения охлаждения образца, в то время как тепло генерируется во время sonication. B. 1,5 мл отцентрифугировать трубка содержащие высокомобильна клеток Пелле pre (слева) и sonication пост (справа). Результате lysate должен отображать более темный оттенок, по сравнению с ресуспензированы клеток Пелле. C. надлежащего разделения супернатант и Пелле ячейки lysate после 18 000 x g центрифугирования. D. СЛТ реакции после 4 ч инкубации при температуре 37 ° C. 1,5 мл отцентрифугировать справа (успешный реакции) показывает видимый флуоресценции белков репортер sfGFP ~ 900 мкг/мл. Отрицательный контроль труб на левом, отсутствует шаблон ДНК и имитация неудачного реакции, показывает ясное решение с не флуоресценции. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 4 : Извлекать изменения экспрессии белка свыше 5 циклов замораживания оттаивания СЛТ. Экстракт, приготовленный из же роста прошли пять заморозить циклов оттаивания через жидкого азота флеш замораживание после оттаивания на льду. Никаких существенных изменений в производительности экстракт для выражения sfGFP были замечены над пяти циклов замораживания оттаивания. Реакции были выполнены в масштабе 15 мкл в 1,5 мл пробирок отцентрифугировать. «Отрицательная» представляет отрицательный контроль, где шаблон не ДНК был добавлен к реакции. Все планки погрешностей представляют 1 стандартное отклонение трех независимых реакций для каждого условия, каждый из которых был количественно в трех экземплярах. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.

Рисунок 5 : СЛТ для реакции с различной [мг2+] и извлекать тома против [sfGFP]. [Мг]2 +варьировались от 8 мм до 14 мм с шагом 2 мм и извлекать тома, варьировались от 3 мкл до 7 мкл с шагом в 1 мкл. Цветовой код представляет [sfGFP] изготовлены из высокого (красный) к низким (фиолетовый). Чтобы максимизировать эффективность реагента при сохранении производства высоким содержанием белка, мы рекомендуем использовать 5 мкл концентрированного экстракта и 10 мм Mg2+ для выдержки, которые имеют содержание общего белка ~ 30 мг/мл, как определяется assay Брадфорд. Исходные точки для создания контурное были основаны от конечной точки флуоресценции трех независимых реакций для каждого условия, каждая из которых была измерена в трех экземплярах. Реакции были выполнены в масштабе 15 мкл в 1,5 мл пробирок отцентрифугировать. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
Дополнительные Рисунок 1: стоимость с мкг белка, произведенного в микролитр реакции на шесть клеток бесплатно белка синтез платформах. Наша платформа сравнивается с пяти различных клеток бесплатно белка синтеза наборы/платформы с различной производительности и цены. Наша платформа СЛТ sonication на основе более экономически эффективным в обоих $/ мкг белка и $/ мкл реакции, чем большинство коммерческих комплекты и обеспечивает легкость комплект для установки реакции, а оставшиеся затраты сопоставимы с других научных платформ СЛТ. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
Дополнительные Рисунок 2: SDS-PAGE sfGFP выражения в СЛТ. Ячейки бесплатно реакции синтеза белка с (+ ДНК) и без (-ДНК) ДНК шаблон для sfGFP были запущены на 12% гель SDSPAGE для демонстрации выражение sfGFP, на 27 кДа (черная стрелка). Были использованы традиционные методы SDS-PAGE. Каждый пример загружены на геле экстракт включены 18 мкг, общего белка, основанный на Брэдфорд пробирного количественного определения общего белка в клетке. На основании измерения интенсивности флуоресценции и нашей стандартной кривой, мы оцениваем, что «+ ДНК» Лейн содержит 0,42 мкг sfGFP. Чтобы получить эти образцы, СЛТ реакции были запущены в масштабе 15 мкл в 1,5 мл отцентрифугировать трубы производства объемных урожайности в соответствии с рис. 3 c. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
Дополнительная цифра 3: калибровочной кривой для sfGFP на Cytation-5. Эта кривая было определено с помощью методов, изложенных выше. Все данные, собранные для этого манускрипта был преобразован из конечной точки флуоресценции чтений [sfGFP] в мкг/мл, используя этот калибровочной кривой. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Синтез белков, клеток бесплатно стала мощной и благоприятных технологий для различных приложений, начиная от biomanufacturing быстрое прототипирование биохимических систем. Спектру приложений поддерживают способность контролировать, управлять и увеличить клеточными в режиме реального времени. Несмотря на расширение влияния этой технологии платформы, остается широкий адаптации медленно из-за технических нюансов в осуществлении методов. Благодаря этим усилиям мы стремимся обеспечить простоту и ясность для установления эту технологию в новой лаборатории. С этой целью наш протокол для е. coli-платформа синтез белка на основе свободных клеток может быть достигнуто в время запуска четырех дней лаборатории обучены не-эксперты, например студентов (рис. 1). Кроме того, после того, как производятся запас реагентов и экстракт, последующие СЛТ пакетного реакции может быть создана, инкубировали и количественно в как раз 5 h. Единый, 1 Л рост клеток может привести к достаточно экстракт для четырехсот 15 мкл СЛТ реакций, в то время как единый пакет препаратов других свободных клеток реагентов может поддерживать тысячи реакций. Реагент препараты также могут быть расширены необходимости даже больших запасов. СЛТ реакции может быть установка в духе высокой пропускной способности, с помощью 96-луночных плиту или ПЦР трубы для тестирования различных условий параллельно. Объемные урожайность будет уменьшаться при использовании небольших судов, как показано на рисунке 2 c. СЛТ реакции может также быть расширена от микролитров десятки миллилитров объема общей реакции для того чтобы увеличить урожайность общего белка для одного условия. При масштабировании до объема, главным соображением является, что объемный реакция урожайности уменьшаться как коэффициент поверхности площадь и объем реакции уменьшается37,52. Для масштабирования при сохранении подобных объемных урожайности экспрессии белков, пользователям следует разделить том реакции на многочисленные реакции сосудов и/или увеличить размер судна. Для реакции весы, начиная от 15 мкл - 100 мкл в объеме, рекомендуются многочисленные 15 мкл реакции параллельно. Для реакций, превышающей 100 мкл в объеме рекомендуется 24-ну пластины с плоским дном, и 12-ну плиты рекомендуются для реакции томов свыше 600 мкл. Такие комбинации томов реакции и судов обеспечивают согласованность в объемном реакция урожайности после масштабирования17,31,37,52. Расширение масштабов за пределами этих томов может быть достигнуто путем использования нескольких скважин пластины параллельно. Используя этот формат, реакции может быть расширена до более чем 10 мл общего объема. Оптимизация реакции объем реакции судна комбинация может поддерживать приложения biomanufacturing без ущерба для производительности реакции.
При выполнении этот протокол, есть несколько ключевых соображений, которые влияют объемные реакция урожайности, а также показатели, связанные с неэффективные экстракт. Для обеспечения надлежащего лизис и избежание денатурации функциональных транскрипция/перевод механизма, важно для уменьшения производства во время лизис тепла. Погружайте ресуспендирования клеток в ледяной воде ванны во время sonication быстро рассеивать тепло во время sonication (рис. 3A). Показателем эффективного клеток лизис является появление темнее появление lysate клетки по сравнению с pre-sonicated пробы (рис. 3B). Для пользователя гибкости sonicator и зонд, показано на рисунке 3A адаптируется к ряду томов из 100 мкл до 15 мл ресуспензированы клеток. Для этого, пользователь может настроить количество джоули доставлены для лизиса желаемого объема клеток. Кроме того большие объемы экстракт может быть подготовлен через два взаимодополняющих подхода. Пользователи могут sonicate несколько труб параллельно, и/или sonicate большего количества клеток взмучивания, масштабирование количество энергии пропорционально с томом как описано29,45. Другой шаг, который указывает на качество экстракт является центрифугирования шаг после лизиса клеток. После лизиса клеток, мы рекомендуем центрифугирования в 18000 x g обеспечить четкое разделение между супернатант (транскрипция/перевод машин, раздробленной геномной ДНК, которая больше не функционирует шаблон транскрипция/перевод) и Пелле ( нежелательные клетчатых компонентов клеточной мембраны и осажденный белки) (рис. 3 c). Мы нашли что центрифугирования в 18000 x g улучшает разделения, что приводит к Улучшенная воспроизводимость, по сравнению с спинов на более низких скоростях, например g 12000 x. Для удобства мы рекомендуем использовать столешницы охлажденных центрифугу, способным достичь не менее 12000 x г. Этот шаг обычно выполняется на 30000 x g, которые должны быть рассмотрены, если соответствующее оборудование доступны54,55,56,,5758, 59 , 60. Экстракт производительность не зависит от центрифугирования скорости на этот шаг, учитывая, что надлежащее разделение достигается. При удалении желаемого супернатанта, это лучше избегать любых облачно материалов, которые существуют на границе между супернатант и Пелле, поскольку это загрязнение снизит производительность экстракта. Стремясь за чистоту супернатант приводит к более продуктивной экстрактов и стоит уменьшение количество экстракта, полученные для новых пользователей.
Важно отметить, что хотя методы, которые мы представили воспроизводимость и может быть выполнена учеными с минимальным опытом, может быть партии партии и вариация реакции и реакции. Это может объясняться различия в составе proteomic lysate после sonication61. Партии партии изменчивости, которую мы наблюдали обычно уменьшается после добавок с T7RNAP и оптимизации концентрации магния. Экзогенных T7RNAP является распространенным среди СЛТ реакции для поддержки оптимального белков, и мы считаем, что наличие двух источников T7RNAP - эндогенного выражение в BL21*(DE3) и дополнительные T7RNAP до конечной концентрации 16 мкг/мл - улучшает воспроизводимость партии партии для новых пользователей45,46. С опытом работы пользователи могут изменять свои эксперименты, чтобы использовать только один источник T7RNAP, при желании. Количественное определение содержания общего белка новой партии экстракт и соответствующую корректировку мг2 + концентрации может также помочь уменьшить различия партии партии в объемный белка выражение урожайности. Вариации в выражении протеина также может быть из-за различий в размерах и структуре протеина интереса, использование кодон гена и его соответствующий сайт связывания рибосома гена интереса, а также тип вектора выражения используется62 ,63. По этим причинам некоторые белки могут не выразить а также модель белка sfGFP, обусловило снижение объемной урожайности от СЛТ реакций.
Ограничения метода представленных СЛТ включают что оно не может быть непосредственно подходит для всех приложений клеток-Free, таких как метаболический инжиниринг и тюнинг выражение условий, без дополнительных изменений к протоколам. Однако мы считаем, что этот протокол будет служить основой для создания платформы СЛТ в новой лаборатории и обеспечивают неспециалистов с возможностью осуществлять вводный реакции свободных клеток в их лабораториях. После первоначального внедрения исследователи могут экспериментировать с платформы вносить собственные изменения для более конкретных приложений, основанных на других литературы в области.
СЛТ платформы по цене $ 0,021/мкг белка (за исключением стоимости труда и оборудования), что делает наша система, конкурентоспособным ценам с коммерческие наборы без ущерба для простоты установки реакции. Оценки сравнительной стоимости мкл реакции показывают аналогичные тенденции ( рис. 1дополнительный ). Оценим затраты запуска для ~ $4500 для всех реактивов и дополнительные $3200 для специализированного оборудования, например sonicator. Человеко-часов для завершения этой процедуры оцениваются в ~ 26 h для всех prep реагент от земли вверх. Однако после того, как были подготовлены большие запасы реагентов, спрос на труд существенно уменьшаются. Кроме того как накопленный опыт с платформы, мы рекомендуем масштабирования размер роста клеток, экстракт подготовка и подготовка реагента для максимизации эффективности время. Учитывая расходы на запуск, мы рекомендуем СЛТ платформы для приложений в синтетической биологии, высок объём усилия, и белка выражение условия, несовместимы с традиционными белка выражение платформы из-за конфликта с ячейкой Биохимия и жизнеспособности ограничения. В этих специализированных случаях, когда желаемый метод включена платформой СЛТ большую стоимость СЛТ в vivo выражение является оправданным.
Дальнейшее развитие платформы СЛТ может обеспечить более широкие утилита усилиям биотехнологии такие как метаболический Инжиниринг ферментативные пути, производства и характеристика традиционно сложных белков, нестандартные аминокислоты Включение и неестественным белков, производства слоистых медицины и расширяется за пределы лаборатории в классе для стволовых образование64,,6566. Эти усилия будут подкрепляться продолжающиеся усилия для подробного описания платформы СЛТ. Лучшее понимание состава выдержки клетки приведет к дальнейшее уточнение к Улучшенная реакция урожайности и гибкость в реакции условий61,,6768.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Авторы заявляют, что они имеют не конкурирующих финансовых интересов или другие конфликты интересов.
Acknowledgments
Авторы хотели бы признать д-р Дженнифер VanderKelen, Андреа Лаубшер и Тони Turretto технической поддержки, Уэсли Kao, Layne Уильямс и Кристофер Hight полезные обсуждения. Авторы также признают финансовой поддержки от Билла и Линда Фрост фонд, центр для приложений в биотехнологии в Chevron биотехнологии применяются исследовательский фонд Грант, Cal Поли исследовательских, научных и творческой деятельности грантовой программы (RSCA 2017), и Национальный научный фонд (NSF-1708919). MZL признает Калифорнийский государственный университет выпускников Грант. MCJ признает армии исследований бюро W911NF-16-1-0372, Национальный научный фонд предоставляет MCB-1413563 и MCB-1716766, военно-воздушные силы исследований лаборатории центра передового опыта Грант FA8650-15-2-5518, обороны угрозы сокращения агентства грант HDTRA1-15-10052/P00001, Дэвида и Люсиль Пакард, Camille Дрейфус преподаватель стипендиат программы, Департамент из энергии BER Грант де SC0018249, человека границ научной программы (RGP0015/2017), Грант ЭТОП DOE Объединенный институт генома, и Чикаго биомедицинских консорциум с поддержкой от Searle фондов в Чикаго доверия сообщества для поддержки.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}