

Summary
Detta protokoll uppgifter om åtgärder, kostnader och utrustning krävs för att generera E. coli-baserade cell extrakt och genomföra in vitro- proteinsyntes reaktioner inom 4 dagar eller mindre. För att utnyttja flexibiliteten i denna plattform för bred applikationer, diskutera vi reaktionsbetingelser som kan anpassas och optimeras.
Abstract
Under de senaste 50 åren, har Cell-Free Protein syntes (CFPS) framträtt som en kraftfull teknik för att utnyttja den transkriptionell och translationell kapaciteten hos celler i ett provrör. Genom att undanröja behovet av att upprätthålla livskraften i cellen, och genom att eliminera cellulära barriären, har CFPS varit grundläggande för nya tillämpningar inom biomanufacturing av traditionellt utmanande proteiner, samt applikationer i rapid prototyping för metaboliska teknik och funktionsgenomik. Våra metoder för att implementera en E. coli-baserad plattform som CFPS ge nya användare åtkomst till många av dessa program. Här, beskriver vi metoder för att förbereda extrakt med hjälp av berikade media, förbryllad kolvar och en reproducerbar metod för avstämbara ultraljudsbehandling-baserade cellys. Detta extrakt kan sedan användas för proteinuttryck kan producera 900 µg/mL eller mer av super mapp grönt fluorescerande protein (sfGFP) i bara 5 h från experiment till dataanalys, med tanke på att lämpliga reagens bestånd har upprättats i förväg. Beräknad start kostar att erhålla reagenser $4.500 som kommer att stödja tusentals reaktioner till en beräknad kostnad av $0,021 per µg av protein som produceras eller $0,019 per µL av reaktion. Dessutom spegel protein uttryck metoderna användarvänlighet reaktion setup sett i kommersiellt tillgängliga system på grund av optimering av reagens premixer, till en bråkdel av kostnaden. För att möjliggöra för användaren att utnyttja flexibiliteten i CFPS plattformen för breda program, har vi identifierat en mängd olika aspekter av den plattform som kan vara trimmad och optimerad beroende på tillgängliga resurser och protein uttryck utfallen önskas.
Introduction
Cellfria Protein syntes (CFPS) har vuxit fram som en teknik som har låst upp ett antal nya möjligheter för proteinproduktion, Funktionsgenomik, metabola engineering och mer inom de senaste 50 år1,2. CFPS jämfört med standard i vivo protein uttryck plattformar, och ger tre viktiga fördelar: 1) cell-fri natur av plattformen möjliggör produktion av proteiner som skulle vara potentiellt giftiga eller utländska till cell3,4 ,5,6. (2) inaktivering av genomisk DNA och införandet av en mall DNA kodning av gene(s) av intresse kanalisera all den systemiska energin inom reaktionen till produktionen av proteinernas av intresse. och 3) öppna karaktär av plattformen ger användaren möjlighet att ändra och övervaka reaktion villkor och sammansättning i realtid7,8. Denna direkta tillgång till reaktionen stöder augmentation av biologiska system med utökade kemier och redox förutsättningar för produktion av nya proteiner och trimning av metaboliska processer2,9, 10. direkt tillgång tillåter även användaren att kombinera CFPS reaktionen med aktivitet-analyser i ett singel-pot system för snabbare design-build-test cykler11. Kapacitet att utföra CFPS reaktionen i liten volym droppar eller på papper-baserade enheter ytterligare stöder hög genomströmning discovery ansträngningar och prototyper12,13,14,15 ,16. Grund av dessa fördelar och den plug and play typ av systemet, har CFPS unikt möjliggjort en mängd biotekniska tillämpningar såsom produktion av proteiner som är svåra att solubly express i vivo17, 18,19,20, upptäckt av sjukdom21,22,23, på efterfrågan biomanufacturing18,24 ,25,26,27, och utbildning28,29, varav alla Visa flexibilitet och nyttan av cellfria plattformen.
CFPS system kan genereras från en mängd rå lysates från båda prokaryota och eukaryota cellinjer. Detta möjliggör olika alternativ i systemet för val, som alla har för- och nackdelar beroende på tillämpningen av intresse. CFPS system varierar också kraftigt i förberedelsetid, kostnader och produktivitet. Mest utnyttjas vanligen cell extrakt produceras från vetegroddar, kanin retikulocyter, insekt celler och Escherichia coli celler, med den sistnämnda är den mest kostnadseffektiva hittills samtidigt som det producerar högsta volymetriska avkastningarna av protein30 . Medan andra CFPS system kan vara fördelaktiga för deras medfödda posttranslationella modifieringen maskiner, framväxande program med hjälp av E. coli-baserade maskiner ska kunna överbrygga klyftan genom att generera anläggningsvis fosforyleras och Glykosylerat proteiner på efterfrågan31,32,33,34,35.
CFPS reaktioner kan köras i antingen batch, kontinuerlig-exchange cellfria (CECF) eller kontinuerlig flöde cell-fri (CFCF) format. Det batch-formatet är ett slutet system vars reaktion livstid är begränsad på grund av minskande mängder reaktanterna och ansamling av inhiberande biprodukter av reaktionen. CECF och CFCF metoder öka livslängden av reaktionen och resultera därmed i ökat volymetriska protein avkastning jämfört med batch reaktionen. Detta åstadkoms genom att låta biprodukter av proteinsyntesen ska tas bort från reaktionskärlet medan nya reaktanterna levereras under hela den reaktion2. När det gäller CFCF, kan proteinet av intresse också tas bort från reaktionskammaren, medan i CECF, proteinet av intresse förblir i reaktionskammaren består av en halvgenomträngligt membran36,37. Dessa metoder är särskilt värdefullt att övervinna dåliga volymetriska avkastningarna av svårt-att-express proteiner av intresse38,39,40,41,42, 43. Utmaningarna i genomförandet av de CECF och CFCF strategier är att 1) medan de resulterar i effektivare användning av den bio maskinen ansvarar för transkription och translation, de kräver framför allt större mängder reagens som ökar totalkostnad och 2) de kräver mer komplex reaktion uppställningar och specialutrustning jämfört med de batch format44. För att säkerställa tillgänglighet för nya användare, beskrivs protokoll som häri fokus på batch format på reaktion volymer av 15 µL med specifika rekommendationer för att öka volymen reaktion till milliliter skalan.
De metoder som presenteras häri aktivera icke-experter med grundläggande laboratorium färdigheter (t.ex. studenter) att genomföra celltillväxt, extrahera förberedelse och batch-format reaktion setup för en E. coli-baserat CFPS system. Detta tillvägagångssätt är kostnadseffektiv jämfört med kommersiellt tillgängliga kit utan att offra lättheten av kit-baserade reaktion setup. Dessutom kan detta tillvägagångssätt program i laboratorium och i fält. När man beslutar att genomföra CFPS, bör nya användare grundligt utvärdera effekten av konventionell protein uttryck system för start investeringar, eftersom CFPS inte kanske är överlägsen i varje fall. CFPS metoderna som beskrivs här möjligt för användaren att direkt genomföra en mängd olika tillämpningar, inklusive Funktionsgenomik, hög genomströmning provning, produktion av proteiner som är svårlösta för i vivo uttryck, samt fältet applikationer inklusive biosensorer och utbildningspaket för syntetisk biologi. Ytterligare applikationer såsom metabola engineering, trimning av protein uttryck villkor, sjukdom upptäckt och skala upp med CECF eller CFCF metoder är fortfarande möjligt men kan kräva erfarenhet med CFPS plattform för vidare modifiering av reaktion villkor. Våra metoder kombinera tillväxt i berikade media och förbryllad kolvar, med relativt snabba och reproducerbara metoder för cellys genom ultraljudsbehandling, följt av en förenklad CFPS reaktion installation som använder optimerade premixer45. Medan de cellulära tillväxt metoderna har blivit något standardiserat inom fältet, varierar metoder för cellen lysis kraftigt. Utöver ultraljudsbehandling inkluderar vanliga lysis metoder utnyttjandet av en fransk press, en Homogenisatorer, pärla visparna, eller lysozym och andra biokemiska och fysiska störningar metoder46,47,48, 49. med våra metoder, ca 2 mL av rå cell extrakt erhålls per 1 L av celler. Denna kvantitet av cell extrakt kan stödja fyra hundra 15 µL CFPS reaktioner, varje producerande ~ 900 µg/mL reporter sfGFP protein från den mall plasmiden pJL1-sfGFP. Denna metod kostar $ 0,021/µg av sfGFP produceras ($.019/µL av reaktion), exklusive kostnader för arbetskraft och utrustning (kompletterande figur1). Start från scratch, denna metod kan genomföras på 4 dagar av en enda person och upprepa CFPS reaktioner kan slutföras inom timmar (figur 1). Protokollet kan dessutom skalas upp i volym för större partier av REAGENSBEREDNING att passa användarens behov. Ännu viktigare, kan det protokoll som presenteras här genomföras genom laboratoriet utbildade icke-experter såsom studenter, eftersom det kräver endast grundläggande laboratorium färdigheter. De förfaranden som beskrivs nedan, och medföljande videon har utvecklats speciellt för att förbättra tillgängligheten av E. coli CFPS plattform för bred användning.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Media förberedelse och celltillväxt
-
Dag 1
- Strimma E. coli BL21*(DE3) celler från en glycerol lager på en LB agarplattan och Inkubera under minst 18 timmar vid 37 ° C.
- Förbereda 50 mL LB media och autoklav lösningen på en flytande cykel under 30 minuter vid 121 ° C. Förvara i rumstemperatur.
-
Dag 2
- Förbereda 750 mL 2 x YTP media och 250 mL 0,4 M D-glukos lösning som beskrivs i den kompletterande informationen.
- Häll 2 x YTP media i en Ånghärdad 2.5 L förbryllad kolv och D-glukos lösningen till en Ånghärdad 500 mL glasflaska. Autoklav båda lösningarna på en flytande cykel under 30 minuter vid 121 ° C.
- Se till att båda sterila lösningar lagras vid 37 ° C om celltillväxt utförs på nästa dag, för att maximera tillväxten vid inympning. Blanda inte lösningar tills inympning.
Obs: Lösningar kan lagras vid 4 ° C för 1-2 d om det behövs, även 2 x YTP media är mycket benägna att kontaminering. - Starta en övernattning kultur av BL21(DE3) genom ympning 50 mL LB media med en enda koloni av BL21(DE3) med en steriliserad ögla och steril teknik för att undvika kontaminering.
- Placera de 50 mL av BL21*(DE3) LB kultur i en 37 ° C 250 rpm skakar inkubator och växa över natten för 15-18 h.
- Förbereda och sterilisera alla material som krävs för dag 3 och 4, inklusive: två 1 L centrifug flaskor, 4 x kall 50 mL koniska rör (väga och registrera massorna av tre) och många 1,5 mL microfuge rör.
-
Dag 3
- Ta bort 50 mL övernattning kulturen av BL21*(DE3) i LB från skakningar inkubatorn och mäta OD600 på en spektrofotometer med en 1:10 utspädning med LB media. Beräkna volymen av övernattning kultur nödvändigt att lägga till 1 L av media för en start OD600 0,1 (exempelvis om en OD600 av en 1:10 utspädning läses som 0.4, Inokulera 25 mL outspädd OD600 = 4,0 övernattning kultur i 1 L 2 x YTP (G).
- Ta bort de värmde 2 x YTP media och D-glukos lösningar från 37 ° C inkubatorn tillsammans med de 50 mL LB kultur. Med steril teknik, häll försiktigt D-glukos lösning i 2 x YTP media (undvika sidorna av förbryllad kolven).
Obs: Tillägg av D-glukos Slutför receptet för 1 L 2 x YTPG. - Att upprätthålla steril teknik, Inokulera 1 L 2 x YTPG lösning med lämplig mängd 50 mL kulturen för att påbörja 1 L kulturen på en 0,1 OD600. Placera omedelbart inokulerade 1 L kulturen i en 37 ° C skakar inkubator på 200 rpm.
- Ta den första OD600 läsa efter den första timmen av tillväxt (fördröjning fas typiskt tar 1 h). Späd inte kulturen. Fortsätt att ta OD600 mätningar ungefär varje 20-30 min tills OD600 når 0,6.
- När du når OD600 = 0,6, tillsätt 1 mL 1 M IPTG (slutlig koncentration i 1 L kultur = 1 mM) till 2 x YTPG kultur.
Obs: Idealisk induktion OD600 är 0,6; en rad 0,6-0,8 är dock godtagbart. Induktion av IPTG är för endogen produktion av T7 RNA-polymeras (T7RNAP). - Efter induktion, mäta OD600 ungefär varje 20-30 min tills den når 3.0.
Obs: Svalka centrifugen till 4 ° C under denna tid. Förbereda kalla S30 buffert som detaljerad kompletterande uppgifter i. Om S30 bufferten är förberett i förväg, kontrollera att DTT inte har lagts fram till användning. - När OD600 når 3.0 (figur 2A), häll kulturen i en kall flaska 1 L centrifug i ett is-vatten-bad. Förbereda en vattenfylld 1 L centrifug flaska lika stor vikt som ska användas som en balans i centrifugen.
Obs: Absorptionsvärden variera från instrument till instrument. OD600 av skörden av BL21(DE3) är inte en känslig variabel, rekommenderas det att användaren utvärdera och optimera denna variabel som en åtgärd för felsökning. Större spektrofotometrar kan resultera i relativt lägre OD600 avläsningar jämfört med mindre cuvette-baserade spektrofotometrar. - Centrifugera i 1 L-flaskor i 10 min på 5 000 x g och 10 ° C till pellet celler.
- Häll långsamt bort supernatanten och kassera den enligt institutets biologiska avfall förfaranden. Placera pelleten på is.
- Med hjälp av en steril spatel, skrapa cellpelleten från flaskan med centrifug och överföra den till en kall 50 mL koniska tub.
- Tillsätt 30 mL kall S30 buffert till konisk röret och resuspendera cellpelleten av vortexa med korta skurar (20-30 s) och viloperioder (1 min) på is tills fullt resuspended med inga bitar.
- När pelleten är fullt resuspended, använda en annan 50 mL koniska tub med vatten som en balans och Centrifugera i 10 min på 5000 x g och 10 ° C (pre kylas till 4 ° C).
Obs: Detta avslutar 1st 3 tvättar krävs när skörden cellerna. - Häll ut supernatanten och kassera den enligt institutets biologiska avfall förfaranden. Återsuspendera pelleten med 20-25 mL kall S30 buffert och Centrifugera i 10 min på 5000 x g och 10 ° C (pre kylas till 4 ° C).
Obs: Detta avslutar 2nd 3 tvättar. - Igen, häll ut supernatanten och kassera den enligt institutets biologiska avfall förfaranden. Tillsätt exakt 30 mL S30 buffert och vortex igen för att resuspendera pelleten.
- Använda 3 pre vägde, kall 50 mL koniska rören och en serologisk pipett filler med en steril pipett alikvotens 10 mL återsuspenderad pellet/S30 buffert blandning i var av 3 koniska rören.
Obs: Dela upp cellerna i 3 rör krävs inte, men detta steg resulterar i mindre cell pellets (~ 1 g) för ökad bekvämlighet vid senare steg. - Centrifugera alla rör, använder lämpliga saldon som behövs, i 10 min på 5000 x g och 10 ° C (pre kylas till 4 ° C).
Obs: Detta avslutar sista tvättningen. - Häll ut supernatanten och kassera den enligt institutets biologiska avfall förfaranden. Ta bort överflödigt S30 bufferten genom att försiktigt torka insidan av koniska röret och keps med en ren trasa; Undvik att vidröra pelleten.
- Väg om dem rören på en Analysvåg och den slutliga pellet vikten på varje rör.
Obs: Protokollet kan pausas vid denna punkt. Pellets kan vara flash fryst i flytande kväve och lagras vid-80 ° C i upp till ett år tills den behövs för extrahera förberedelse.
2. råolja Cell extrahera förberedelse - dag 4
- För utdrag förberedelse, hålla celler kall på is under varje steg. Tillsätt 1 mL kall S30 buffert per 1 g av cellmassan av pelleten. Säkerställa att Ditiotreitol (DTT) har kompletterats till S30 bufferten till en slutkoncentration av 2 mM.
Obs: Svalka i mikrocentrifug till 4 ° C under denna tid. - Återsuspendera cellpelleten av vortexa med korta skurar (20-30 s) och viloperioder (1 min) på is tills fullt resuspended. Om resuspension är svårt, lämna pellets på is för 30 min att Tina.
- Över 1,4 mL återsuspenderade celler till ett 1,5 mL mikrofugrör.
- Placera en 1,5 mL tub som innehåller 1,4 mL återsuspenderade celler till en is vattenbad i en bägare. Sonikera för 45 s på följt av 59 s off för 3 totala cykler, med amplitud inställd på 50%. Stäng och Invertera rören för att blanda försiktigt under off perioder. Totalt leverera 800-900 J av energi till varje 1,5 mL mikrofugrör med 1,4 mL återsuspenderade celler (figur 3A & 3B).
Obs: Detta steg är känsliga för någon sonikator typ och modell används och bör optimeras om utrustning är annorlunda än som anges för detta förfarande. Två kompletterande metoder kan användas för att skala upp mängden extrakt beredd under detta steg: (1) flera 1,5 mL microfuge rör kan vara sonicated parallellt, eller 2) större volymer kan vara sonicated i koniska rör (upp till 15 mL cell resuspension per rör) , skalning mängden energi som levereras som tidigare beskrivs 29,45. - Omedelbart efter ultraljudsbehandling är klar, tillsätt 4,5 µL av 1 M DTT (komplettera en ytterligare 2 mM DTT) i de 1,4 mL lysat och Invertera flera gånger för att blanda. Placera röret på is. Upprepa steg 2.4 och 2.5 för eventuella ytterligare tuber av återsuspenderade celler innan centrifugering.
- Mikrocentrifug prover på 18.000 x g och 4 ° C i 10 min (figur 3 c).
- Pipettera supernatanten till en ny 1,5 mL mikrofugrör. Inte störa pelleten; Det är bättre att lämna vissa supernatanten bakom för att upprätthålla renhet än för att störa pelleten i strävan att maximera avkastningen.
- Inkubera supernatanten från föregående steg vid 250 rpm och 37 ° C i 60 min genom att tejpa rören för att de skakar plattformen av inkubator (detta är avrinning reaktionen).
- Mikrocentrifug prover på 10.000 x g och 4 ° C i 10 min.
- Ta bort supernatanten utan att störa pelleten och överföra den till en ny tub. Skapa många 100 µL portioner av extrakt för lagring.
Obs: Protokollet kan pausas här, och extraktet kan vara flash fryst i flytande kväve och lagras vid-80 ° C i upp till ett år tills den behövs för CFPS reaktioner. Minst 5 frysning-tining cykler kan vara genomgått utan nackdel att extrahera produktivitet (figur 4).
3. cell-Free proteinsyntes Batch Format reaktioner
- Tina lösningar A och B, DNA mall, BL21(DE3) extrakt (om de är frysta), T7RNAP och en alikvot av molekylärbiologisk kvalitet vatten.
Obs: CFPS reaktion mall kan hittas i den Kompletterande Information. Lösningar A och B recept finns i Kompletterande uppgifter och motsvarar specifika koncentrationer för många reagenser att stödja PANOx-SP baserat energisystemet för CFPS. Varje reagens och acceptabel variation i dessa reagens koncentrationer som kan stödja CFPS roll har varit beslutsam50. Ett T7RNAP rening protokoll kan hittas i tilläggsinformation51. Kompletterande T7RNAP kan öka volymetriska avkastningen men är inte nödvändigt om T7RNAP induceras under celltillväxt. Plasmid DNA template (pJL1-sfGFP) kan förberedas med ett maxiprep kit med två tvättar med tvättlösning i kit, följt av en efterbearbetning DNA-sanering med hjälp av en PCR-rening kit (figur 2B). Linjära DNA-mallar kan också användas i CFPS reaktioner. - Märk den nödvändiga mängden microfuge rör behövs för CFPS reaktioner.
Obs: Reaktioner kan utföras i olika storlekar för fartyg, men ett mindre fartyg kan minska volymetriska protein avkastning (figur 2 c). Skala upp en reaktion i samma storlek fartyg kan också minska volymetriska avkastning, som en funktion av minskande syre utbyte, på grund av en minskning av ytan volym-förhållande. När ökar reaktionsvolym ovan 100 μL, är det rekommenderat att använda platt botten plattor med 31,37,52. - Tillsätt 2.2 µL av lösning A, 2.1 µL av lösning B, 5 µL BL21*(DE3) extrakt, 0,24 µg av T7RNAP (16 μg/mL slutliga koncentration), 0,24 ng DNA mall (16 ng/mL koncentration) och vatten för att få den slutliga volymen till 15 µL.
Obs: Vortex lösningar A och B ofta under reaktion installationen för att undvika sedimentering av komponenter och säkerställa att varje reaktion får en homogen alikvot av varje lösning. Undvika vortexa extraktet, istället Invertera röret att blanda. - Efter alla reagens har lagts till reaktion, blanda varje rör genom pipettering upp och ner eller försiktigt vortexa samtidigt att slutliga reaktionsblandningen kombineras till en enda 15 µL pärla längst ned på den 1,5 mL mikrofugrör.
- Placera varje reaktion till 37 ° C inkubatorn utan skakningar i 4 h eller 30 ° C över natten.
Obs: Framgångsrika reaktioner kan kvalitativt bedömas visuellt utifrån den gröna färgen på sfGFP produkten inom CFPS reaktionsblandningen (figur 3D). Uttryck för proteinet av intresse kan också bekräftas av SDS-PAGE (kompletterande figur 2).
4. kvantifiering av proteinet Reporter, [sfGFP]
- Läsa in 48 µL av 0,05 M HEPES, pH 8, i varje väl behövs för kvantifiering (vanligtvis utförs i tre exemplar per reaktionsröret).
- Ta bort reaktioner från inkubator. Pipettera upp och ner för att blanda varje reaktion, sedan överföra 2 µL av reaktion till den 48 µL av 0,05 M HEPES, pH 8. Pipettera upp och ner igen i bra att blanda.
- När alla reaktioner är laddad och blandat, placera 96 väl plattan i fluorometer och mäta sfGFP slutpunkt fluorescensen.
Obs: Excitation och utsläpp våglängder för sfGFP fluorescens kvantifiering är 485 nm och 510 nm, respektive. - Använda en tidigare genererade standardkurvan, bestämma den [sfGFP] från erhållna fluorescens avläsningar.
Obs: Instruktioner för att generera en standardkurva sfGFP koncentration kontra fluorescensintensiteten finns i tilläggsinformation (kompletterande diagram 3). Användare måste upprätta en standardkurva för sina instrument eftersom instrumentet känslighet kan variera.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Vi har lagt fram en ultraljudsbehandling-baserade E. coli extrahera förberedelse protokoll som kan kompletteras under en fyra dagars span, med figur 1 visar den processuella fördelningen över varje dag. Det finns formbarhet med steg som kan genomföras på varje dag med olika pausa punkter, men vi har funnit detta arbetsflöde ska vara den mest effektiva att utföra. Dessutom är både cell pellets (steg 1.3.18) och fullt beredd extrakt (steg 2.10) stabila vid-80 ° C i minst ett år, så att användaren kan skapa större bestånd av var och en att spara för användning vid en senare tid17. Inte bara är extraktet stabil över långa tidsperioder, men extraktet kan också genomgå minst fem frysa tö cykler utan en betydande förlust av produktivitet (figur 4). Detta möjliggör större portioner av extrakt kan lagras för flera användningsområden om frys lagringsutrymme är begränsat. Vi rekommenderar dock flera mindre portioner (~ 100 μL) av extrakt som möjligt.
Med varje ny extrakt förberedelse rekommenderar vi att användaren utför en magnesium titrering för att fastställa den optimala mängden magnesium för den batchen Extract. Användare kan kvantifiera sats till sats variabilitet i total proteinkoncentration av cell extrakt av Bradford assay. För högre utför extrakt, ser vi vanligtvis totalprotein koncentrationer på 30-50 mg/mL, och inom detta spänna finns det inget uppenbart samband mellan totalt protein koncentrationer och cell extrakt prestanda. Därför rekommenderar vi att användare samklang magnesium koncentrationer med detta för att säkerställa att proteiner och nukleinsyra funktionalitet maximeras för varje extrakt parti. Magnesium är viktigt för korrekt DNA-replikering, transkription och translation, men alltför höga halter kan vara skadligt för dessa processer53. För att demonstrera detta beroende, vi har utfört en samtidig titrering av magnesium och extrahera volym för att bestämma den optimala kombinationen som minimerar mängden extrakt nödvändigt, samtidigt som en produktiv reaktion (figur 5). Från detta experiment rekommenderar vi att du använder 5 µL av extraktet och 10 mM Mg2 + för extrakt med en total proteinhalt på 30 mg/mL, för att få över 1000 μg/mL sfGFP.
Vår erfarenhet med CFPS har också tillåtit oss att bestämma steg inom det protokoll som kan varieras utan negativa följder för den totala produktiviteten av systemet, och andra som är integrerad i ett system med hög presterande i CFPS. Framför allt, den slutliga OD600 av cellen skörd påverkar inte signifikant slutresultatet av CFPS reaktionen, och cellerna kan rimligen skördas någonstans från 2,7-4,0 OD600. Detta representerar den tidiga exponentiella fasen av tillväxt där Ribosomen koncentration per cell är den högsta och den translationella maskinen är de mest aktiva att stödja snabb tillväxt. Denna observation användarna flexibilitet att optimera sina egna förfaranden. Vi rekommenderar skörd på cirka 3,0 OD600 för att fånga cellerna på en OD600 närmare för 3,3 av tid skörd är komplett (figur 2A). Variabler som påverkar hur CFPS avkastning inkluderar mallen DNA kvalitet, reaktion fartygens storlek och de relativa mängder cell extrakt och magnesium ion i reaktionen. Vi har hittat DNA kvalitet att ha anmärkningsvärda sats till sats variation. För att lösa detta, rekommenderar vi att användare rena DNA via midi eller maxi prep, följt av en ytterligare DNA cleanup steg antingen på kolumnen för DNA-rening används i maxiprep, eller efter rening med hjälp av ett ytterligare DNA rengöring kit. Detta förbättrar reproducerbarhet i DNA kvalitet för CFPS reaktioner och resultat i mer robust proteinproduktion (figur 2B). Reaktionskärlet påverkar också volymetriska avkastning, sådan att proteinproduktion av identiska reaktion uppställningar i varierande fartyget volymer kan skilja sig upp till 40%. Det har en teori att uppsving i volymetriska avkastning observerade i större fartyg på grund av en ökad yta av reaktionsblandningen, möjliggör bättre syre utbyte (figur 2 c), och andra har ytterligare ökat volymetriska avkastning genom att köra CFPS reaktioner i stora platt-botten pläterar, som vi rekommenderar för reaktioner över 100 µL17,31,37,52.

Figur 1 : Tidslinjen för kultur tillväxt, produktion av cell extrakt, setup och kvantifiering av CFPS reaktioner. Användaren kan implementera CFPS plattformen för deras forskningstillämpningar genom fyra dagars arbetsflödet. REAGENSBEREDNING representerar den primära tid och kostnadseffektiv investering för den första omgången av detta experiment och minskar avsevärt efter reagenser bestånd är etablerade. Cell pellets och beredda cell extrakt kan dessutom lagras för över ett år vid-80 ° C, så att användaren kan påbörja tidslinjen vid olika steg för snabbare resultat. Användaren kan också pausa vid olika steg att ändra tidslinjen av detta arbetsflöde. Klicka här för att se en större version av denna siffra.

Figur 2 : Kan ändras villkoren för CFPS och effekterna på volymetriska reaktion avkastning. A. extrakt produktivitet jämförelse utifrån skörd BL21(DE3) celler på olika OD600 avläsningar. Baserat på denna tomt rekommenderar vi skörd på en OD600 3,3 att producera minst 1000 µg/mL målprotein. Reaktioner framfördes på en 15 μL skala i 1,5 mL microfuge rör. B. jämförelse av två DNA maxiprep tvätta protokoll med och utan efter rening DNA-rensningar. pJL1-sfGFP plasmider genomgick en maxiprep med en eller två tvättar följt av en rensning av efter rening av PCR-rening kit. För att uppnå ~ 900 µg/mL proteinuttryck, föreslår vi att utföra en efter rening DNA rensning oavsett antalet maxiprep tvättar. Reaktioner framfördes på en 15 μL skala i 1,5 mL microfuge rör. C. 15 µL CFPS reaktioner uppträdde i olika fartyg allt från 2 mL till 0,6 mL microfuge rör. ”Neg” representerar en negativ kontroll där ingen DNA-Mall lades till reaktionen. Alla felstaplar representera 1 standardavvikelse av tre oberoende reaktioner för varje villkor, som alla kvantifierades i tre exemplar. Klicka här för att se en större version av denna siffra.

Figur 3 : Nyckel processuella uppställningar och utfall för att skapa produktiva extrakt. A. korrekt inställning av ultraljudsbehandling ice vattenbad för kylning av prov medan värme genereras under ultraljudsbehandling. B. 1,5 mL mikrofugrör med återsuspenderade cell pellets pre (vänster) och efter (höger) ultraljudsbehandling. Den resulterande lysate ska visa en mörkare nyans jämfört återsuspenderade cellpelleten. C. särskilja av supernatanten och pellet av cell lysate efter 18 000 x g centrifugering. D. CFPS reaktioner efter 4 h inkubation vid 37 ° C. 1,5 mL mikrofugrör till höger (framgångsrik reaktion) visar synlig fluorescens av proteinet sfGFP reporter vid ~ 900 μg/mL. Negativ kontroll röret till vänster, saknar mall DNA och simulera en misslyckad reaktion, visar en klar lösning med ingen fluorescens. Klicka här för att se en större version av denna siffra.

Figur 4 : Förändring i proteinuttryck över 5 frysning-tining cykler för CFPS extrahera. Extrakt beredd från samma tillväxt genomgick fem frysa tö cykler via flytande kväve flash frysa följt av upptining på is. Inga signifikanta förändringar i extraktet produktivitet för att uttrycka sfGFP sågs över de fem frysning-tining cyklerna. Reaktioner framfördes på en 15 μL skala i 1,5 mL microfuge rör. ”Neg” representerar en negativ kontroll där ingen DNA-Mall lades till reaktionen. Alla felstaplar representera 1 standardavvikelse av tre oberoende reaktioner för varje villkor, som alla kvantifierades i tre exemplar. Klicka här för att se en större version av denna siffra.

Figur 5 : CFPS för reaktioner med varierande [Mg2+] och extrahera volymer jämfört med [sfGFP]. [Mg2 +] varierade från 8 mM till 14 mM med 2 mM intervall och extrahera volymer varierat från 3 µL till 7 µL med 1 µL inkrement. Den färg kod representerar [sfGFP] produceras från hög (röd) till låg (lila). För att maximera reagens effektivitet bibehållen hög proteinhalt produktion, vi rekommenderar att du använder 5 µL av extraktet och 10 mM Mg2+ för extrakt som har en totalt proteininnehåll ~ 30 mg/ml, som bestäms av Bradford assay. Ursprungliga punkter att generera kontur tomten var baserad på endpoint fluorescens av tre oberoende reaktioner för varje villkor, vilka mättes i tre exemplar. Reaktioner framfördes på en 15 μL skala i 1,5 mL microfuge rör. Klicka här för att se en större version av denna siffra.
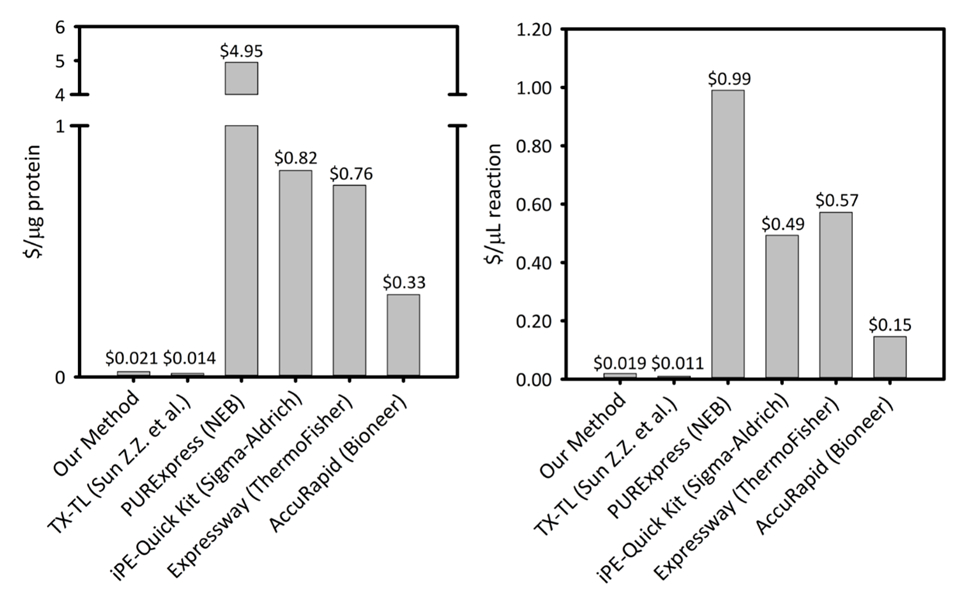
Kompletterande bild 1: kostnad per mikrogram av protein som produceras och per mikroliter reaktion på sex cellfria proteinsyntes plattformar. Vår plattform är jämfört med fem olika cell gratis proteinsyntes kit/plattformar med varierande produktivitet och prissättning. Vår ultraljudsbehandling baserat CFPS plattform är mer kostnadseffektiv i båda $/ µg av protein och $/ µL av reaktion än de flesta kommersiella kit och ger användarvänlighet ett kit för reaktion setup, medan återstående kostnad jämförbar med andra akademiska CFPS plattformar. Klicka här för att se en större version av denna siffra.
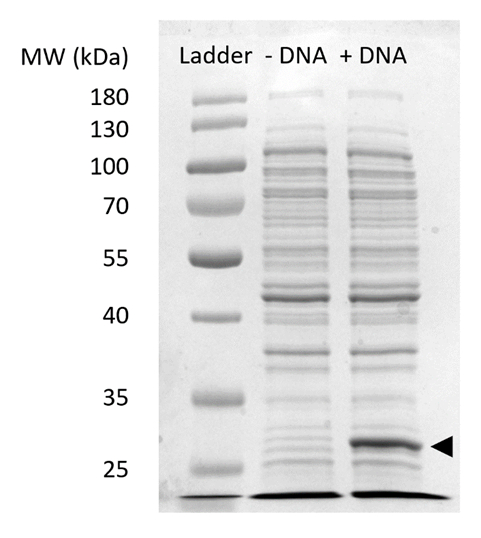
Kompletterande bild 2: SDS-PAGE sfGFP uttryck i CFPS. Cellfria proteinsyntes reaktioner med (+ DNA) och utan (-DNA) DNA mall för sfGFP kördes på 12% SDSPAGE gel att Visa uttrycket av sfGFP observerades vid 27 kDa (svart pil). Traditionella SDS-PAGE tekniker användes. Varje prov lastas på gelen ingår 18 µg totalt protein baserat på Bradford assay kvantifiering av totalt protein i cellen extrahera. Baserat på fluorescens intensitet mätningar och våra standardkurvan, vi uppskattar att den ”+ DNA” lane innehåller 0,42 µg av sfGFP. För att erhålla dessa prover, drevs CFPS reaktioner på en 15 µL skala i 1,5 mL microfuge rör producerar volymetriska uttag som är förenligt med figur 3 c. Klicka här för att se en större version av denna siffra.
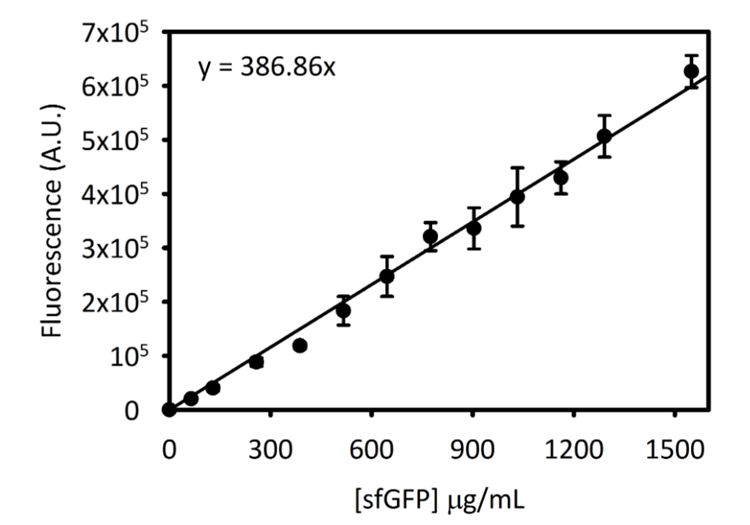
Kompletterande figur 3: Standardkurvan för sfGFP på Cytation 5. Denna kurva bestämdes med hjälp av metoder som beskrivs ovan. Alla data som samlats in för denna manuskriptet var konverterat från endpoint fluorescens avläsningar till [sfGFP] i µg/mL med hjälp av denna standardkurvan. Klicka här för att se en större version av denna siffra.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Cellfria proteinsyntesen har vuxit fram som en kraftfull och möjliggörande teknik för en mängd tillämpningar som sträcker sig från biomanufacturing till prototyper av biokemiska system. Bredden av program stöds av kapaciteten att övervaka, manipulera och öka cellulära maskineriet i realtid. Trots den växande effekten av denna plattform teknik, bred anpassning har varit långsam på grund av tekniska nyanser i genomförandet av metoder. Genom denna insats vill vi att enkelhet och tydlighet för att upprätta denna teknik i nya övningar. Mot därför våra protokoll för en E. coli-baserade cellfria proteinsyntes plattform kan uppnås inom en starttiden för fyra dagar av laboratoriet utbildade icke-experter, såsom studenter (figur 1). Dessutom, när ett lager av reagenser och extrakt produceras, kan efterföljande CFPS batch reaktioner vara ställa in, inkuberas och kvantifieras i bara 5 h. En enda, 1 L celltillväxt kan resultera i tillräckligt extrakt för fyra hundra 15 µL CFPS reaktioner, medan parti beredning av andra cellfria reagensen kan stödja tusentals reaktioner. Reagens preparat kan också skalas om ett ännu större lager behövs. CFPS reaktionerna kan ställas in på ett sätt som hög genomströmning, med en plattan med 96 brunnar eller PCR-rören för provning av en mängd olika förhållanden parallellt. Volymetrisk avkastning kommer att minska när du använder mindre fartyg som kan ses i figur 2 c. CFPS reaktioner kan också skalas upp från mikroliter till tiotals milliliter av total reaktionsvolym för att öka den totala protein avkastningen för ett enda villkor. När skalning upp volym, är det främsta övervägandet att volymetriska reaktion avkastning minskar eftersom förhållandet yta området till volym av reaktionen minskar37,52. För att skala upp bibehållen liknande volymetriska avkastningarna av proteinuttryck, bör användare dela volymen reaktion i talrika reaktionskärl eller öka fartygets storlek. För reaktion skalor alltifrån 15 µL - 100 µL i volym, rekommenderas många 15 µL reaktioner parallellt. För reaktioner överstiger 100 µL i volym, platt botten 24 brunnar rekommenderas, och 12-väl plattor rekommenderas för reaktion volymer överstigande 600 µL. Sådana motståndare av reaktion volymer och fartyg ger konsekvens i volymetriska reaktion avkastning vid skala upp17,31,37,52. Skala upp bortom dessa volymer kan åstadkommas genom att utnyttja flera brunnar i plattan parallellt. Använder detta format, kan reaktionen skalas till över 10 mL total volym. Optimera kombinationen reaktion volym-reaktion fartyget kan stödja tillämpningar av biomanufacturing utan att offra produktiviteten i reaktionen.
När du utför detta protokoll, finns det några viktiga faktorer som påverkar volymetriska reaktion avkastning samt indikatorer som är associerad med dåligt utför extrakt. För att säkerställa korrekt lysis och förhindra denaturering av funktionella transkription och översättning maskiner, är det viktigt att mildra den värme som produceras under Lys. Fördjupa cell resuspension i en ice vattenbad under ultraljudsbehandling till snabbt avleda värme under ultraljudsbehandling (figur 3A). En indikator på effektiv cellys är uppkomsten av ett mörkare utseende av cellen lysate jämfört med pre-sonicated prover (figur 3B). För användaren flexibilitet är och någon sonikator sond som visas i figur 3A anpassningsbar till ett antal volymer från 100 µL till 15 mL återsuspenderade celler. För att åstadkomma detta, kan användaren justera antalet joule levereras för lys av den önskade volymen av celler. Större volymer av extrakt kan dessutom förberedas genom två kompletterande metoder. Användare kan Sonikera flera rör samtidigt, och sonikera större volymer av cell resuspension, skalning mängden energi proportionellt med volymen som tidigare beskrivits29,45. En annan som visar extrakt kvalitet är det centrifugeringssteget efter cellys. Inlägget cellys, vi rekommenderar centrifugering vid 18.000 x g att ge en tydlig uppdelning mellan supernatanten (transkription och översättning maskiner, fragmenterade genomiskt DNA som inte längre fungerar mallen transkription/översättning) och pellet ( oönskade cellulära komponenter såsom cellmembranet och utfällda proteiner) (figur 3 c). Vi har funnit att centrifugering på 18.000 x g förbättrar separation, vilket resulterar i förbättrad reproducerbarhet jämfört spins vid lägre hastigheter som 12 000 x g. Vi rekommenderar att du använder en bordsskiva kyld centrifug, kan uppnå ett minimum av 12 000 x g för bekvämlighet. Detta steg utförs också vanligen vid 30 000 x g, som bör övervägas om lämplig utrustning är tillgängliga54,55,56,57,58, 59 , 60. extrakt prestanda inte påverkas av centrifugering hastigheter vid detta steg med tanke på att särskilja uppnås. När du tar bort önskad supernatanten, är det bäst att undvika grumlig material som finns vid gränsen mellan supernatanten och pellets eftersom denna kontaminering minskar produktiviteten i extraktet. Siktar på renhet av supernatanten resulterar i mer produktiva extrakt och är värt den reducera mängden extrakt som erhålls för nya användare.
Det är viktigt att notera att medan de metoder som vi har presenterat är reproducerbara och kan utföras av forskare med minimal kompetens, det kan finnas sats till sats och reaktion-till-reaktion variation. Detta kan hänföras till variation i proteomiska sammansättningen av den lysate efter ultraljudsbehandling61. Sats till sats variabiliteten som vi har observerat är generellt minskat vid komplettering med T7RNAP och optimering av magnesium koncentrationer. Exogena tillägg av T7RNAP är vanligt bland CFPS reaktioner att stödja optimal proteinuttryck, och vi finner att ha två källor av T7RNAP - endogen uttryck i BL21*(DE3) och den kompletterande T7RNAP till en slutlig koncentration på 16 µg/mL - förbättrar sats till sats reproducerbarhet för nya användare45,46. Med erfarenhet, kan användare ändra sina experiment för att utnyttja endast en enda källa till T7RNAP om så önskas. Kvantifiering av totala proteininnehållet i en ny sats av extrakt och lämplig anpassning av Mg2 + koncentration kan också hjälpa för att minska sats till sats variation i volymetriska protein uttryck avkastning. Variationer i proteinuttryck kan också bero på skillnader i storlek och struktur av proteinet av intresse, kodon användningen av genen och dess motsvarande Ribosomen bindningsställe av genen av intresse, liksom vilken typ av uttryck vektorns62 ,63. Av dessa skäl kan vissa proteiner inte express samt den modell protein sfGFP, vilket resulterar i minskad volymetriska avkastning från CFPS reaktioner.
Begränsningar av presenterade CFPS tekniken är att det inte kanske direkt lämpar sig för alla ansökningar av cell-fri, såsom metabola engineering och trimning av uttrycket villkor, utan ytterligare ändringar till protokollen. Vi anser dock att detta protokoll kommer att ge en grund för att fastställa CFPS plattformen i nya laboratorier och tillhandahålla icke-experter med förmågan att genomföra inledande cellfria reaktioner i deras labb. Efter inledande genomförandet, kan forskare experimentera med plattformen att göra egna ändringar för mer specifika program baserat på annan litteratur på området.
CFPS plattform kostnader $ 0,021/µg proteinet (exklusive kostnader för arbetskraft och utrustning), gör vårt system konkurrenskraftigt prissatta med kommersiella kit utan att kompromissa med användarvänlighet reaktion setup. Bedömningar av jämförande kostnader per µL av reaktion visar liknande trender (Supplemental figur 1). Vi uppskattar startkostnader för att vara ~ $4,500 för alla reagenser och en ytterligare $3,200 för specialutrustning, såsom en någon sonikator. Personen timmar att slutföra den här proceduren bedöms vara ~ 26 h för alla reagens prep från marken upp. Men när stora lager av reagenser har förberetts, minskar krav på arbetskraft avsevärt. Dessutom som med plattformen är erfarenheter, vi rekommenderar skala upp storleken på celltillväxten, extrahera förberedelse och REAGENSBEREDNING att maximera tidseffektivitet. Med tanke på kostnaderna som uppstått, vi rekommenderar CFPS plattformen för tillämpningar inom syntetisk biologi, hög genomströmning ansträngningar, och protein uttryck villkor som är oförenliga med traditionella protein uttryck plattformar på grund strider mot cellens biokemi och livskraft begränsningar. I dessa specialiserade fall där önskad tekniken är aktiverat av CFPS plattformen, är den största kostnaden för CFPS över i vivo uttryck motiverad.
Fortsatt utveckling av CFPS plattformen är sannolikt att ge bredare nytta till bioteknik insatser såsom metabola konstruktion av enzymatisk stigar, produktion och karakterisering av traditionellt svårlösta proteiner, icke-standard aminosyra införlivande och onaturliga proteinuttryck, stratifierat medicin tillverkning och expanderar bortom laboratoriet i klassrummet för STEM utbildning64,65,66. Dessa ansträngningar kommer att stödjas ytterligare av de pågående insatserna för detaljerad karakterisering av CFPS plattformen. En bättre förståelse av sammansättningen av cell extraktet kommer leda till fortsatt förfining mot förbättrad reaktion avkastning och flexibilitet i reaktion villkor61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna förklarar att de har inga konkurrerande ekonomiska intressen eller andra intressekonflikter.
Acknowledgments
Författarna vill erkänna Dr Jennifer VanderKelen, Andrea Laubscher, och Tony Turretto för teknisk support, Wesley Kao, Layne Williams, och Christopher Hight för bra diskussioner. Författarna erkänner också finansiellt stöd från Bill och Linda Frost fonden, Center för applikationer inom bioteknikens Chevron bioteknik tillämpas Endowment forskningsbidrag, Cal Poly forskning, Scholarly och kreativa aktiviteter Grant Program (RSCA 2017), och National Science Foundation (NSF-1708919). MZL erkänner California State University Graduate bidraget. MCJ erkänner den armén forskning Office W911NF-16-1-0372, beviljar National Science Foundation MCB-1413563 och MCB-1716766, de flygvapen forskning laboratorium Center av Excellence Grant FA8650-15-2-5518, Defense Threat minskning byrån bidraget HDTRA1-15-10052/P00001, David och Lucile Packard Foundation, programmet Camille Dreyfus lärare-forskare, Institutionen för energi BER Grant DE-SC0018249, den mänskliga gränser Naturvetenskapsprogrammet (RGP0015/2017), DOE Joint Genome Institute ETOP bidraget, och Chicago biomedicinsk konsortiet med stöd från Searle fonderna på the Chicago Community Trust för stöd.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}