

Summary
Bu iletişim kuralı adımları, maliyetleri ve ekipman E. colioluşturmak için gerekli ayrıntıları-hücre özleri dayalı ve 4 gün içinde veya daha az vitro protein sentez reaksiyonları uygulamak. Bu platformun geniş uygulamalar için esnek yapısı kullanabilmeniz için biz uyarlanmış ve en iyi duruma getirilmiş reaksiyon koşulları tartışıyorlar.
Abstract
Son 50 yılda, boş hücre Protein sentezi (CFPS) tüp bebek hücrelerde transkripsiyon ve çevirim kapasitesi koşum için güçlü bir teknoloji olarak ortaya çıkmıştır. Hücre canlılığı korumanıza gerek obviating tarafından ve hücresel bariyer ortadan kaldırarak CFPS hızlı prototipleme için geleneksel olarak zorlu proteinlerin biomanufacturing ortaya çıkan uygulamalarda yanı sıra uygulamaları temel olmuştur metabolik Mühendisliği ve fonksiyonel genomik. Bir E. coliuygulamak için bizim yöntemleri-tabanlı CFPS platformu bırakmak bu uygulamaların birçoğu erişmek yeni kullanıcı. Burada, özü zenginleştirilmiş medya, şaşkın şişeler ve akort sonication tabanlı hücre lizis tekrarlanabilir yöntemi kullanılarak hazırlamak için yöntemleri açıklanmaktadır. Uygun reaktif stokları önceden hazırlanmış göz önüne alındığında bu özü sonra protein ifade 900 µg/mL veya daha fazla süper klasör yeşil flüoresan protein (sfGFP) veri analizi, deneysel kurulum sadece 5 h üretebilen için kullanılabilir. Reaktifler elde etme tahmini başlangıç maliyeti 4500 $ $0.021 µg üretilen protein başına veya 0.019 $ µL tepki başına tahmini ücret karşılığında tepkiler binlerce sürdürmek olduğunu. Ayrıca, protein ifade yöntemleri piyasada bulunan sistemlerde reaktif öncesi karışımları, bir kısmını maliyeti duruma getirilmesi nedeniyle görülen tepki kurulum kolaylığı ayna. Geniş uygulamaları için CFPS platform esnek doğası kaldıraç sağlamak için biz bu şekilde ayarlanmış ve kullanılabilir kaynakları ve protein ifadesi sonuçlar bağlı olarak en iyi duruma getirilmiş platformu yönlerini çeşitli belirledik istenen.
Introduction
Boş hücre Protein sentezi (CFPS) protein üretim, fonksiyonel genomik, metabolik Mühendisliği ve son 50 yıl1,2içinde daha fazlası için yeni fırsatlar bir dizi kilidi bir teknoloji olarak ortaya çıkmıştır. Standart vivo içinde protein ifade platformları için karşılaştırıldığında, CFPS üç önemli avantajlar sağlar: 1) platformu boş hücre doğa olası toksik veya dış hücre3,4 olurdu proteinleri sağlar ,5,6; 2) inactivation genomik DNA'ın ve bir şablon kodlama ilgi gene(s) DNA giriş tüm protein(s) üretimi için tepki ilgi içinde sistemik enerji kanal; ve 3) platformu açık yapısı değiştirin ve gerçek zamanlı7,8kompozisyon ve reaksiyon koşulları izlemek kullanıcı sağlar. Biyolojik sistemleri genişletilmiş kimyaları ve Redoks koşullarına roman proteinleri ve metabolik süreçleri2,9, ayarlama için büyütme tepki bu doğrudan erişimi destekler 10. doğrudan erişim de daha hızlı tasarım-yapı-test11döngüleri için bir tek-pot sisteminde CFPS reaksiyon etkinlik deneyleri ile birleştirmek kullanıcı sağlar. Küçük hacimli damlacıkları CFPS reaksiyonu gerçekleştirmek veya kağıt tabanlı cihazlarda daha fazla destekler yüksek üretilen iş bulma çabaları ve hızlı prototipleme12,13,14,15 kapasite ,16. Bu avantajları ve Tak ve Kullan doğa sisteminin bir sonucu olarak, CFPS gibi solubly hızlı vivo içindeiçinzor olan proteinleri biyoteknoloji uygulamaları çeşitli benzersiz olarak sağladı17, 18,19,20, hastalık21,22,23, isteğe bağlı biomanufacturing18,24 tespiti ,25,26,27ve göstermek tüm-in hangi esneklik ve yeni boş hücre platformu eğitim28,29,.
CFPS sistemleri hem prokaryotik ve ökaryotik hücre satırlarından ham lysates çeşitli oluşturulur. Bu seçim, sistemdeki farklı seçenekler her biri avantaj ve dezavantajları bağlı olarak faiz uygulanması vardır sağlar. CFPS sistemleri de hazırlık süresi, maliyet ve verimlilik büyük ölçüde değişir. En sık özleri buğday tohumu, tavşan Retikülosit, böcek hücreleri ve Escherichia coli hücreleri, ikinci en uygun maliyetli protein30 en hacimsel verimi üreten ise bugüne varlık ile üretilen hücre kullanılan . Diğer CFPS sistemleri onların doğuştan gelen translasyonel modifikasyon makine için avantajlı olabileceği gibi E. colikullanan uygulamalar ortaya-tabanlı makine site-specifically fosforile oluşturarak boşluğu edebiliyoruz ve glikozile proteinler talep31,32,33,34,35.
CFPS reaksiyonlar ya da toplu iş iş işlemde, sürekli değişimi-boş hücre (CECF) veya sürekli akış boş hücre (CFCF) biçimleri çalıştırabilirsiniz. Kapalı bir sistem olan reaksiyon ömür boyu azalan miktarda Reaktanları ve inhibitör yan reaksiyon birikimi nedeniyle sınırlıdır Toplu biçimidir. CECF ve CFCF yöntemleri tepki ömrünü artırmak ve böylece toplu tepki göre artan hacimsel protein verimleri neden. Bu yeni Reaktanları tepki2ders boyunca sağlanan süre tepki gemiden kaldırılacak protein sentezi yan sağlayarak gerçekleştirilir. CFCF söz konusu olduğunda, faiz protein tepki odasında CECF iken, protein yarı geçirgen membran36,37oluşan reaksiyon odasında faiz kalıntılarının da kaldırılabilir. Bu yöntemler özellikle zor express proteinler faiz38,39,40,41,42yoksul hacimsel verimi üstesinden değerlidir, 43. CECF ve CFCF yaklaşımlar uygulanmasında zorluklar 1) varken Onlar transkripsiyon ve translasyon için sorumlu biyo makinelerin daha verimli kullanılmasını sonucu, toplam maliyeti ve 2 artırır reaktifler özellikle büyük miktarda gerektirir vardır) Onlar daha karmaşık reaksiyon kurulumları ve özel ekipman için toplu iş biçimi44karşılaştırıldığında gerektirir. Yeni olan kullanıcılar için erişilebilirlik sağlamak için iletişim kuralları burada toplu biçimini mililitre ölçek tepki birime artırmak için özel öneriler ile 15 µL tepki hacimleri, odak nitelendirdi.
Burada sunulan yöntemleri sigara-uzmanlar (lisans öğrencisi gibi) temel laboratuvar becerileri ile hücre büyümesini uygulamak, hazırlık ayıklamak ve biçimi tepki Kur bir E. coliiçin toplu etkinleştirmek-tabanlı CFPS sistemi. Bu yaklaşım için piyasada kitleri tepki kiti tabanlı kurulum kolaylığı ödün vermeden uygun maliyetli karşılaştırılır. Ayrıca, bu yaklaşım uygulamaları laboratuvarında ve alan sağlar. CFPS her durumda üstün olmayabilir olarak CFPS uygulamaya karar verirken, yeni kullanıcılar başlangıç yatırım için geleneksel protein ifade sistemlerinin etkinliğini iyice değerlendirmelidir. CFPS yöntem tanımlamak burada doğrudan uygulayan uygulamaları, fonksiyonel genomik, test, alanının yanı sıra vivo içinde ifade için zorlu proteinleri yüksek üretilen iş de dahil olmak üzere çeşitli sağlamak biyosensörler ve sentetik Biyoloji eğitim setleri gibi uygulamalar. Metabolik Mühendisliği gibi ek uygulamalar protein ifade koşulları, hastalık algılama ve ölçek-up kullanarak CECF veya CFCF yöntemlerini hala mümkün ancak deneyim CFPS platformu ile daha fazla tepki değiştirilmesi için gerektirebilir ayarlama koşullar. Bizim yöntemleri hücre lizis sonication, en iyi duruma getirilmiş premiksler45kullanan bir basitleştirilmiş CFPS reaksiyon Kur tarafından takip aracılığıyla nispeten hızlı ve tekrarlanabilir yöntemleri ile zenginleştirilmiş medya ve şaşkın şişe, büyüme birleştirir. Hücresel büyüme yöntemleri biraz içinde bu alan standart haline iken, hücre lizis için yöntemleri büyük ölçüde değişmektedir. Sonication ek olarak, ortak lizis yöntemleri kullanımı bir Fransız basınında, bir homogenizer, boncuk Tokmaklar, veya lizozim ve diğer biyokimyasal ve fiziksel bozulma yöntemleri46,47,48, içerir. 49. bizim yöntemleri kullanarak, yaklaşık 2 mL ham hücre ekstresinin elde edilir hücre 1 L. Hücre özü bu miktarı dört yüz destekleyebilir 15 µL CFPS reaksiyonlar, her üreten ~ 900 µg/mL muhabir sfGFP protein şablon plazmid pJL1-sfGFP. Bu yöntem maliyeti $0.021/µg, üretilen sfGFP ($.019/µL, tepki), işgücü ve ekipman (tamamlayıcı şekil 1) maliyeti hariç. Sıfırdan başlayarak, bu yöntem 4 gün içinde tek bir kişi tarafından uygulanabilir ve CFPS reaksiyonlar (şekil 1) saat içinde tamamlanabilir yineleyin. Ayrıca, protokol birimdeki kullanıcı ihtiyaçlarına uygun reaktif hazırlık daha büyük toplu işlemler için ölçeklenebilir. Önemlisi, sadece temel laboratuvar becerileri gerektirir gibi burada sunulan Protokolü eğitim laboratuvarı sigara-uzmanlar gibi lisans öğrencileri tarafından uygulanabilir. Aşağıda açıklanan işlemler ve beraberindeki video E. coli CFPS platformu için geniş kullanım erişilebilirliğini geliştirmek için özel olarak geliştirilmiştir.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. medya hazırlanması ve hücre büyümesi
-
Gün 1
- Bir gliserol çizgi E. coli BL21*(DE3) hücrelerden bir LB agar tabağa stok ve en az 18 h 37 ° C'de için kuluçkaya
- Bir sıvı döngüsü için 121 ° C'de 30 dk 50 mL LB medya ve otoklav çözüm hazırlamak Oda sıcaklığında saklayın.
-
2. gün
- 750 mL 2 x YTP medya ve 250 mL 0,4 M D-glukoz çözeltisi içinde ek bilgiler açıklandığı şekilde hazırlayın.
- 2 x YTP kitle iletişim araçları içine bir autoclaved 2.5 L şaşkın şişesi ve D-glikoz çözüm bir autoclaved 500 mL cam şişe içine dökün. Otoklav sıvı döngüsü 121 ° C'de 30 dk için her iki çözümleri
- Hücre büyümesini büyüme oranları aşı üzerine en üst düzeye çıkarmak ertesi gün gerçekleştiriliyorsa steril her iki çözüm de 37 ° C'de depolanan emin olun. Çözümler kadar aşı birleştirmeyin.
Not: 2 x YTP kitle iletişim araçları için kirlenme son derece eğilimli olsa çözümleri için 1-2 gerekirse, d 4 ° C'de muhafaza edilebilir. - BL21(DE3) bir gecede kültür LB Medya 50 mL steril döngü ve steril tekniği kirlenmesini önlemek için kullanarak BL21(DE3) bir koloni ile aşı başlatın.
- 50 mL BL21*(DE3) LB kültürünün 37 ° C 250 kuluçka sallayarak devir/dakika yerleştirin ve gecede 15-18 h için büyür.
- Hazırlamak ve gün için 3 ve 4, dahil olmak üzere gerekli tüm malzemeleri sterilize: iki 1 L santrifüj şişe, 4 x soğuk 50 mL konik tüpler (tartmak ve üç kayıt kitlelerin) ve birçok 1.5 mL microfuge tüpleri.
-
3. gün
- BL21*(DE3) 50 mL gecede kültür LB içinde sallayarak kuluçka makinesi kaldırmak ve bir 1:10 kullanarak bir Spektrofotometre üzerinde OD600 ölçmek seyreltme LB medya ile. Gecede kültür medya 0.1 başlangıç OD600 için 1 L eklemek gerekli hacmi hesaplamak (örneğin, 4.0 gecede kültür 1 L / 2 = 1:10 seyreltme 0,4 okumak bir OD600 aşılamak ise 25 mL su katılmamış OD600 x YTP G).
- Isıtılan 2 x YTP kitle iletişim araçları ve D-glikoz çözümleri ile birlikte 50 mL LB kültürünün 37 ° C kuluçka kaldırın. Steril tekniği kullanarak, dikkatle D-glikoz solüsyonu (şaşkın şişeye kenarlarında kaçınarak) 2 x YTP kitle iletişim araçları dökün.
Not: D-glikoz ilavesi 2 1 L tarifini tamamlar x YTPG. - Steril tekniği Bakımı, 1 0.1 OD6001 L kültür başlamak için L 2 x YTPG çözüm 50 mL kültürü uygun miktarda aşılamak. Hemen kuluçka 200 devir / dakikada sallayarak 37 ° C inoculated 1 L kültür yerleştirin.
- Büyüme ilk saat sonra okuma ilk OD600 almak (gecikme faz tipik alır 1 h). Kültür sulandırmak değil. OD600 0,6 ulaşana kadar yaklaşık olarak her 20-30 dk OD600 ölçümler alarak devam ediyor.
- OD600 ulaşan üzerine = 0,6, 1 mL 1 m IPTG ekleyin (1 L kültür son konsantrasyonu = 1 mM) 2 x YTPG kültür.
Not: 0,6 Ideal indüksiyon OD600 olduğunu; Ancak, bir dizi 0,6-0,8 kabul edilebilir. İndüksiyon IPTG tarafından endojen T7 RNA polimeraz (T7RNAP) için yapımıdır. - 3.0 ulaşıncaya kadar indüksiyon sonra yaklaşık olarak her 20-30 dk OD600 ölçmek.
Not: Bu süre içinde 4 ° c santrifüj sakin. Soğuk S30 arabellek tamamlayıcı bilgiler detaylı olarak hazırlayın. S30 arabellek önceden hazırlanmış, DTT kullanımı güne kadar eklenmez emin. - OD600 3.0 (şekil 2A) ulaştığında, kültür bir buz-su banyosu içinde soğuk bir 1 L santrifüj şişe içine dökün. Bir su dolu 1 L santrifüj şişe eşit ağırlıklı bir denge santrifüj içinde kullanılmak üzere hazırlayın.
Not: Araç gereç Absorbans değerleri farklı. BL21(DE3) hasat OD600 hassas bir değişken olmasa da, kullanıcı değerlendirmek ve bu değişken sorun giderme bir tedbir olarak optimize önerilir. Daha büyük Spektrofotometreler daha küçük küvet tabanlı Spektrofotometreler göre nispeten düşük OD600 okuma neden olabilir. - 1 L şişe 5000 x g ve hücreleri cips 10 ° C de 10 dakika santrifüj kapasitesi.
- Süpernatant ve bu kurumun biyolojik atık yordamlara göre belirleme yapılacağı atmayın yavaş yavaş dökün. Pelet Buza koyun.
- Steril bir spatula kullanarak, hücre Pelet santrifüj şişeden raspa ve soğuk 50 mL konik tüp transferi.
- Soğuk S30 arabellek 30 mL konik tüp ekleyin ve hücre pelet tarafından tamamen yok parçaları ile resuspended kadar buz öbekler (20-30 s) ve dinlenme süreleri (1 dk) ile vortexing resuspend.
- Pelet tam resuspended sonra 5000 x g ve 10 ° C (4 ° C-önceden soğutmalı) adlı başka bir 50 mL konik tüp su ile bir denge ve 10 dk santrifüj olarak kullanabilirsiniz.
Not: Bu 3 yıkar hücreleri hasat gerekli 1st tamamlar. - Dökmek süpernatant ve bu kurumun biyolojik atık yordamlara göre belirleme yapılacağı atmayın. Pelet 20-25 mL soğuk S30 arabellek ve 5000 x g ve 10 ° C (4 ° C-önceden soğutmalı) 10 dk santrifüj ile resuspend.
Not: Bu 3 yıkar 2nd tamamlar. - Yine, dökmek süpernatant ve bu kurumun biyolojik atık yordamlara göre belirleme yapılacağı atmayın. Tam olarak 30 mL S30 tampon ve girdap tekrar Pelet resuspend ekleyin.
- 3 önceden tartılır, soğuk 50 mL konik boru ve serolojik pipet dolgu steril bir pipet ile aliquot 10 mL resuspended Pelet/S30 arabellek karışımı her 3 Konik tüpler içine kullanarak.
Not: 3 tüpler içine hücreleri bölme gerekli değildir, ancak bu adım daha küçük hücre peletler sonuçlar (~ 1 g) sonraki adımlarında artan rahatınız için. - Tüm tüpler santrifüj kapasitesi, uygun kullanarak gerektiğinde 5000 g ve 10 ° C (4 ° C-önceden soğutmalı) x 10 min için dengeler.
Not: Bu son yıkama adım tamamlar. - Dökmek süpernatant ve bu kurumun biyolojik atık yordamlara göre belirleme yapılacağı atmayın. Aşırı S30 arabellek dikkatle konik tüp ve temiz bir mendil ile kap içine silerek kaldırma; Pelet dokunmaktan kaçının.
- Tüpler bir analitik dengede coorect ve her tüpün üzerinde son Pelet ağırlığı kaydetmek.
Not: Protokol bu noktada duraklatılmış. Granül sıvı azot içinde donmuş ve özü hazırlık için ihtiyaç kadar bir yıl kadar-80 ° c için depolanan flaş olabilir.
2. ham hücre özü hazırlık - 4 gün
- Özü hazırlık için hücreler her adımı sırasında buz soğuk tutmak. 1 mL 1 g Pelet hücre kütlesi başına soğuk S30 arabellek ekleyin. O dithiothreitol (DTT) S30 arabelleğine 2 mM son bir konsantrasyon için desteklenen sağlamak.
Not: Bu süre içinde 4 ° c microcentrifuge sakin. - Hücre Pelet öbekler (20-30 s) ve dinlenme süreleri (1 dk) ile vortexing tamamen resuspended kadar buz tarafından resuspend. Resuspension zor ise, granül defrost için 30 dakika buz üzerinde bırakın.
- Resuspended hücre 1.4 mL 1.5 mL microfuge tüp içine aktarın.
- 1.4 mL resuspended hücre içine bir buz su banyosu bir ölçek içeren bir 1,5 mL tüp yerleştirin. 45 için solüsyon içeren temizleyicide s 59 tarafından takip genlik ile 3 Toplam devir için kapalı s ayarla % 50. Kapatın ve tüpler kapalı dönemlerde karışımı yavaşça tersine çevirin. Toplamda, 800-900 J enerji resuspended hücre (şekil 3A ve 3B) 1.4 mL içeren her 1.5 mL microfuge tüp için teslim.
Not: Bu adım için sonicator türü duyarlıdır ve modeli kullanılmış ve ekipman bu yordam için listelenen daha farklı ise optimize edilmelidir. İki tamamlayıcı yaklaşımlar ölçek-up bu adımı sırasında hazırlanan özü miktarını için kullanılabilir: 1) birden fazla 1.5 mL microfuge tüpler paralel olarak sonicated ve/veya 2) daha büyük birimleri konik tüplerde sonicated (hücre resuspension tüp başına en fazla 15 mL) , teslim enerji miktarı ölçekleme daha önce açıklandığı gibi 29,45. - Sonication tamamlandıktan hemen sonra 1 m (ilave bir ek 2 mM DTT) DTT 4,5 µL 1.4 mL lysate ekleyin ve karıştırmak için birkaç kez tersine çevirin. Tüp Buza koyun. Santrifüjü geçmeden önce resuspended hücrelerinin herhangi bir ek tüpler için 2.4 ve 2.5 adımları yineleyin.
- Microcentrifuge örnekleri, 18.000 x g ve 4 ° C'de 10 dakika (şekil 3 c).
- Süpernatant yeni 1.5 mL microfuge tüpüne pipette. Pelet rahatsız etmeyin; bazı süpernatant saflık verimi en üst düzeye çıkarmak için çabalarında Pelet bozabilir daha korumak için geride bırakmak tercih edilir.
- Süpernatant 250 devir/dakika ve 37 ° C 60 dk için önceki adımdaki tüpler (Bu ikinci tur reaksiyondur) kuluçka sallayarak platformu taping tarafından kuluçkaya.
- 10.000 x g ve 10 dk 4 ° C'Microcentrifuge örnekleri.
- Ve yeni bir tüp transfer süpernatant Pelet bozmadan çıkarın. Birçok 100 µL aliquots Muhafazası için ekstresinin oluşturun.
Not: Burada protokol duraklatılmış ve sıvı azot içinde donmuş ve CFPS reaksiyonlar için ihtiyaç kadar bir yıl kadar-80 ° c için depolanan özü flaş olabilir. En az 5 donma-çözülme çevrimleri verimlilik (şekil 4) ayıklamak için zararına geçirmiş.
3. hücre-Alerjik Protein sentezi toplu biçimi tepkiler
- Tezcan çözümleri A ve B, DNA şablon, BL21(DE3) Eğer (dondurulmuş) özü, T7RNAP ve moleküler sınıf su bir aliquot.
Not: CFPS reaksiyon şablonu Ek bilgibulunabilir. Çözümler A ve B tarifleri Tamamlayıcı bilgiler sağlanır ve CFPS için PANOx-SP dayalı enerji sistemi desteklemek çok sayıda reaktifler için belirli konsantrasyonları karşılık gelir. Rolü her reaktif ve kabul edilebilir değişkenlik CFPS destekleyebilir Bu reaktif konsantrasyonlarda kararlı50olmuştur. T7RNAP arıtma Protokolü tamamlayıcı bilgiler51de bulunabilir. Ek T7RNAP Volumetrik verimleri artırabilir ama eğer T7RNAP sırasında hücre büyümesini indüklenen gerekli değildir. Plazmid DNA şablonu (pJL1-sfGFP) ile iki yıkama yıkama tampon bir son işlem DNA-PCR arıtma Kiti (şekil 2B) kullanarak Temizleme tarafından takip seti kullanarak bir maxiprep kit kullanarak hazırlanabilir. Doğrusal DNA şablonları CFPS tepkileri de kullanılabilir. - CFPS reaksiyonlar için gerekli microfuge tüpler gerekli miktarda etiketleyin.
Not: Tepkileri çeşitli kap boyutlarında yapılabilir, ancak küçük bir gemi hacimsel protein verimleri (şekil 2C) düşürebilir. Aynı boyutu gemi bir reaksiyon yukarı ölçekleme de hacimsel verimi, oksijen Satım, hacim oranı için yüzey alanındaki bir düşüş nedeniyle azalan bir fonksiyonu olarak azaltabilir. Reaksiyon cilt 100 μL yukarıda artan zaman, düz dipli iyi plakaları 31,37,52kullanılması önerilir. - Çözüm A 2.2 µL eklemek için çözüm b, BL21*(DE3) 5 µL 2.1 µL ayıklamak, T7RNAP 0,24 μg (16 μg/mL nihai toplama), 0,24 ng DNA şablonu (16 ng/mL nihai toplama) ve son hacim için 15 µL getirmek için su.
Not: Girdap çözümleri A ve B reaksiyon Kur sırasında sık sık sedimantasyon bileşenlerinin önlemek ve her tepki her çözüm homojen bir aliquot almasını sağlamak. Vortexing özü önlemek için bunun yerine karıştırmak için tüp ters çevir. - Tüm reaktifler için tepki ekledikten sonra her tüpün yukarı ve aşağı pipetting karıştırmak ya da vortexing tarafından hafifçe son tepki karışımı bir tek 15 µL boncuk alt kısmındaki 1.5 mL microfuge tüp içine birleştirilmiştir sağlarken.
- Gecede 4 h için veya 30 ° C sallayarak olmadan her reaksiyon 37 ° C kuluçka makinesi içine yerleştirin.
Not: Başarılı tepkiler niteliksel görsel olarak bağlı sfGFP ürün CFPS reaksiyon karışımı (şekil 3D) içinde yeşil renkli olarak değerlendirilebilir. İfade ilgi proteinin SDS-sayfa (tamamlayıcı Şekil 2) tarafından onaylanabilir.
4. miktar muhabir protein, [sfGFP]
- 0,05 m HEPES, pH 8, 48 µL her miktar (genellikle nüsha tepki tüp başına yapılan) için de gerekli içine yükleyin.
- Tepkiler kuluçka kaldırın. Her reaksiyon karıştırmak için yukarı ve aşağı pipet, sonra tepki 2 µL 0,05 M HEPES, pH 8 48 µL aktarın. Tekrar iyi karıştırın damlalıklı yukarı ve aşağı.
- Bir kez tüm tepkiler yüklü ve karışık, 96 iyi plaka yer yerleştirin ve sfGFP bitiş noktası Floresans ölçmek.
Not: 485 sfGFP floresan miktar için uyarma ve emisyon dalga boylarında olan nm ve 510 nm, anılan sıraya göre. - Önceden oluşturulan bir standart eğri kullanarak, [sfGFP] elde edilen floresan okuma belirlemek.
Not: ek bilgi (ek şekil 3) sfGFP konsantrasyon floresan yoğunluğu karşı standart bir eğri oluşturmak için yönergeler sağlanmıştır. Kullanıcıların araç duyarlılık değişebilir beri kendi araç için standart bir eğri oluşturmak için gereken.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Biz özü sonication tabanlı E. coli sundu dört gün span boyunca her gün üzerine yordam dökümü gösteren şekil 1 ile tamamlanabilir hazırlık protokolü. Her gün çeşitli duraklatma puan ile tamamlanabilir adımlara yumuşaklık, ama biz yürütmek en etkili olduğu için bu iş akışını bulduk. Ayrıca, hücre granül (Adım 1.3.18) ve tam olarak hazırlanan özü (Adım 2.10) en az bir yıl için kullanıcı her bir sonraki saat17kullanmak için kaydetmek için daha büyük hisse senetleri oluşturmak için izin-80 ° C'de stabildir. Sadece özü uzun süreler stabil ama özü de verimlilik (şekil 4) önemli bir kayıp olmadan en az beş donma çözülme döngüleri uygulayabilir. Bu özü daha büyük aliquots için dondurucu depolama alanı sınırlı ise birden fazla kullanım için saklanmasına olanak. Ancak, birden çok daha küçük aliquots (~ 100 μL) özü mümkün olduğunda önerilir.
Her yeni özü hazırlıkla öneririz kullanıcı magnezyum ekstresinin toplu iş için en uygun miktarını belirlemek için bir magnezyum titrasyon gerçekleştirir. Kullanıcıların hücre ekstresinin toplam protein konsantrasyonu toplu iş toplu iş değişkenlik Bradford tahlil tarafından ölçmek. İçin daha yüksek özleri performans, genelde 30-50 mg/mL toplam protein konsantrasyonları görmek ve bu Aralık içinde toplam protein konsantrasyonları ve hücre özü performans arasında bariz hiçbir bağlantı yoktur. Bu nedenle, protein ve nükleik asit işlevselliği her özü toplu işlem için maksimize sağlamak için kullanıcıların magnezyum konsantrasyonlarının nağme buna göre önerilir. Magnezyum düzeyleri uygun DNA çoğaltma, transkripsiyon ve translasyon için önemlidir, ancak aşırı düzeyleri bu süreçleri53için zararlı olabilir. Bu bağımlılık göstermek için bir eş titrasyon magnezyum gerçekleştirdik ve özü üretken reaksiyonu (şekil 5) korunarak gerekli miktarını en aza indirir en iyi kombinasyon belirlemek için birim ayıklayın. Bu deneyden özü ve 10 mM Mg2 + 5 µL özü 30 mg/mL, bir toplam protein içerikli için 1000'den fazla μg/mL sfGFP elde etmek için kullanmanızı öneririz.
Deneyimlerimiz CFPS ile de bize sistem genel verimliliğini zarar vermeden değiştirilebilir Protokolü ve diğerleri içinde yüksek performans gösteren CFPS sistemi için ayrılmaz adımlarını belirlemek izin verdi. En önemlisi, hücre hasat son OD600 CFPS tepki son çıktı önemli ölçüde etkilemez ve hücreleri feasibly her yerde 2.7-4.0 OD600hasat edilebilir. Bu erken üssel faz nerede ribozom konsantrasyonu hücre başına en yüksek olduğu ve çevirim makine en hızlı büyümeyi desteklemek için etkin olan büyüme gösterir. Bu gözlem kullanıcıların kendi işlemleri en iyi duruma getirmek için esneklik sağlar. Biz zaman hasat tarafından bir OD600 daha yakın 3.3 için hücreleri yakalamak için yaklaşık 3,0 OD600 hasat tam (şekil 2A) öneririz. CFPS verimi etkileyen değişkenler şablon DNA kalite, reaksiyon gemi boyutu ve hücre özü ve magnezyum iyon tepki mevcut göreli miktarlarda bulunmaktadır. DNA kalite önemli toplu iş toplu iş değişim için bulduk. Bu sorunu çözmek için kullanıcıların bir MIDI veya maxi hazırlık, DNA arıtma sütununda maxiprep içinde kullanılan ek bir DNA Temizleme adım ardından veya sonrası arıtma ek bir DNA Temizleme kiti kullanılarak DNA arındırmak öneririz. Bu CFPS reaksiyonlar için DNA kalitesinde tekrarlanabilirlik ve sonuçları daha sağlam protein üretim (2B rakam) artırır. Öyle ki aynı reaksiyon kurulumları değişen gemi cilt olarak protein üretimi % 40 farklı olabilir tepki gemi Volumetrik verimleri, da etkiler. Bu reaksiyon karışımı daha iyi oksijen alışverişi (şekil 2C) için izin artan bir yüzey alanı daha büyük gemiler gözlenen hacimsel verim artırmak kaynaklanmaktadır ve diğerleri daha fazla çalıştırarak hacimsel verimi arttırdı teorisini Biz 100 µL17,31,37,52üzerinde reaksiyonlar için tavsiye büyük düz-alt levha, CFPS reaksiyonlar.

Resim 1 : Zaman çizelgesi kültür büyüme, hücre özü, kurulum ve miktar CFPS tepkiler üretimi için. Kullanıcı bu dört günlük iş akışı aracılığıyla onların araştırma uygulamaları için CFPS platform uygulayabilirsiniz. Reaktif hazırlık birincil saat ve bu denemenin ilk tur için maliyet yatırım temsil eder ve reaktifler stokları oluşturulduktan sonra önemli ölçüde azalır. Ayrıca, hücre topakları ve hazırlanan hücre özü için bir yıl daha hızlı sonuç için çeşitli adımlar timeline başlamak kullanıcı izin-80 ° C'de depolanabilir. Kullanıcı bu iş akışının zaman çizelgesi değiştirmek için çeşitli adımlar da duraklatabilirsiniz. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Resim 2 : CFPS ve hacimsel tepki verimleri üzerinde etkileri için değiştirilebilir koşullar. A. BL21(DE3) hücreler çeşitli OD600 okuma, hasat üzerine dayalı verimlilik karşılaştırma ayıklayın. Bu arsa üzerinde bağlı olarak, bir OD600 3.3 en az 1000 µg/mL hedef proteini üretmek için hasat öneririz. Tepkiler 1.5 mL microfuge tüpler 15 μL ölçekte gerçekleştirilmiştir. B. iki DNA maxiprep yıkama protokol ve sonrası arıtma DNA-temizliğini olmadan karşılaştırılması. pJL1-sfGFP Plasmid'ler ile bir veya iki yıkama sonrası arıtma Temizleme tarafından PCR arıtma paketi tarafından takip bir maxiprep uygulandı. ~ 900 µg/mL protein ifade elde etmek için maxiprep yıkar sayısından bağımsız olarak sonrası arıtma DNA temizlemesi gerçekleştirmek öneririz. Tepkiler 1.5 mL microfuge tüpler 15 μL ölçekte gerçekleştirilmiştir. C. çeşitli damarlarının 2 mL 0.6 mL microfuge tüpler için arasında değişen 15 µL CFPS reaksiyonlar gerçekleştirilen. "Negatif" DNA şablon için tepki eklendiği bir negatif kontrol temsil eder. Tüm hata çubukları her hangi bir nüsha sayısal her koşul için üç bağımsız reaksiyonları 1 standart sapması temsil eder. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Şekil 3 : Anahtar yordam kurulumları ve üretken oluşturmak için sonuçlar ayıklayın. A. sonication buz su banyosu sırasında ısı sonication sırasında oluşturulan örnek soğutma sağlamak için uygun Kur. B. 1.5 mL microfuge tüp resuspended hücre Pelet öncesi (solda) ve yazı (sağda) sonication içeren. Lysate kaynaklanan resuspended hücre Pelet göre daha koyu bir renk tonu görüntülenmelidir. C. uygun ayırma süpernatant ve Pelet hücrenin 18.000 x g aralıklarla sonra lysate. Kuluçka 37 ° C'de 4 h sonra ö CFPS reaksiyonları 1.5 mL microfuge tüp (başarılı reaksiyon) sağda sfGFP muhabir protein görünür floresan ~ 900 μg/mL gösterir. Negatif kontrol tüp şablon DNA eksik ve başarısız bir reaksiyon simüle soldaki net bir çözüm yok floresan ile görüntüler. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Şekil 4 : CFPS için 5 donma-çözülme çevrimleri protein ifadesindeki değişiklik ayıklamak. Aynı büyüme hazırlanan özü uygulandı beş çözülme çevrimleri sıvı azot ile buz çözdürme tarafından izlenen buz gibi flaş dondurma. Herhangi bir önemli değişiklik özü verimlilik sfGFP ifade etmek için beş donma-çözülme çember üzerinde görüldü. Tepkiler 1.5 mL microfuge tüpler 15 μL ölçekte gerçekleştirilmiştir. "Negatif" DNA şablon için tepki eklendiği bir negatif kontrol temsil eder. Tüm hata çubukları her hangi bir nüsha sayısal her koşul için üç bağımsız reaksiyonları 1 standart sapması temsil eder. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Şekil 5 : CFPS değişen reaksiyonlar için [Mg2+] ve [sfGFP] karşı birimleri hulâsa. [Mg2 +] 8 mM 2 mM artışlarla 14 mM arasında değişiyordu ve 3 µL 1 µL artışlarla 7 µL değişmekteydi birimleri ayıklayın. Renk kodu yüksek (kırmızı)-(mor) düşük için üretilen [sfGFP] temsil eder. Yüksek protein üretim koruyarak reaktif verimliliği en üst düzeye çıkarmak için 5 µL özü ve 10 mM Mg kullanmanızı öneririz2+ Bradford tahlil tarafından belirlenen ~ 30 mg/mL, bir toplam protein içeriği var özleri için. Dağılım çizim oluşturmak için orijinal puan bitiş noktası Floresans üç bağımsız reaksiyonların her hangi bir nüsha ölçüldü her koşul için kapalı dayalı idi. Tepkiler 1.5 mL microfuge tüpler 15 μL ölçekte gerçekleştirilmiştir. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
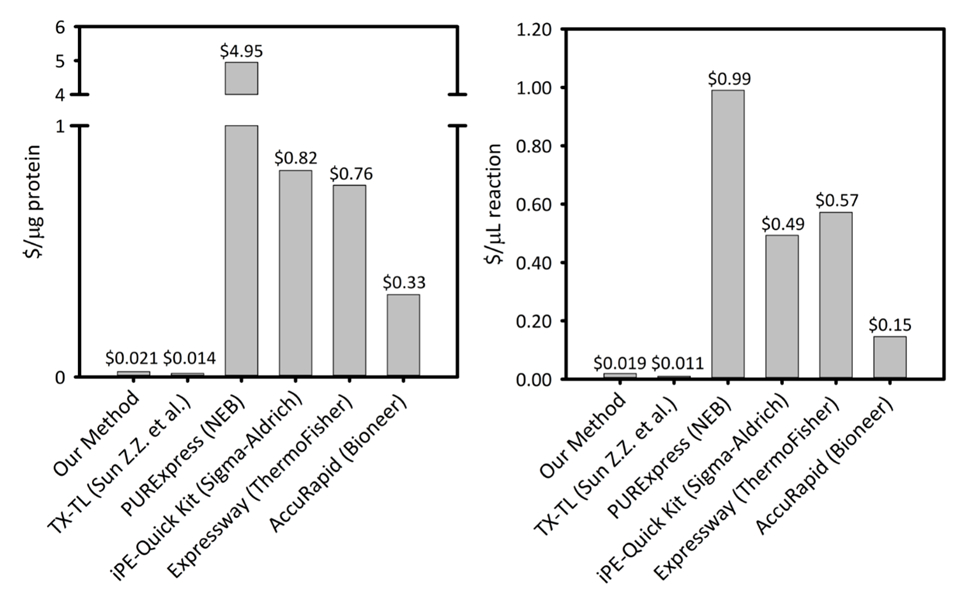
Ek Resim 1: microliter tepki ve mikrogram üretilen protein başına altı boş hücre protein sentezi platformlar arasında mal. Bizim platformu beş farklı hücre ücretsiz protein sentezi kitleri/araçları için verimlilik değişen ve fiyatlandırma ile karşılaştırılır. Bizim temel sonication CFPS platformu daha çoğu ticari kitleri ve kalan maliyet diğer akademik CFPS platformlar için karşılaştırılabilir ise tepki kurulum için bir kit kolaylığı sağlar daha düşük maliyetli her iki $/ µg protein ve $/ µL tepki olduğunu. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
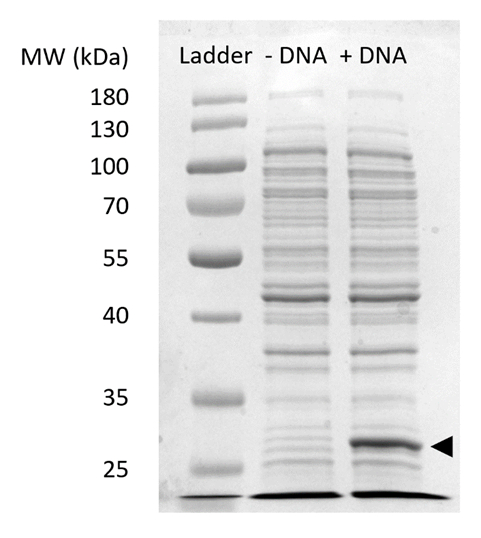
Ek Resim 2: SDS-sayfa CFPS ifadede sfGFP. Protein sentez reaksiyonları ile (+ DNA) boş hücre ve olmadan (-DNA) sfGFP için DNA şablon 27 kDa (siyah ok) gözlenen sfGFP ifade göstermek için SDSPAGE jel % 12 üzerinde çalıştırmak. Geleneksel SDS-sayfa teknikleri kullanılmıştır. Her örnek dahil 18 µg Bradford tahlil miktar toplam protein hücredeki temel alan toplam protein özü jel yüklendi. Floresans yoğunluk ölçümleri ve standart sürecimizde dayalı, biz tahmin "+ DNA" lane sfGFP 0,42 µg içerir. Bu örnekleri edinmek için CFPS reaksiyonlar Volumetrik verimleri şekil 3 cile tutarlı üreten 1.5 mL microfuge tüpler 15 µL ölçekte işletilmiştir. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
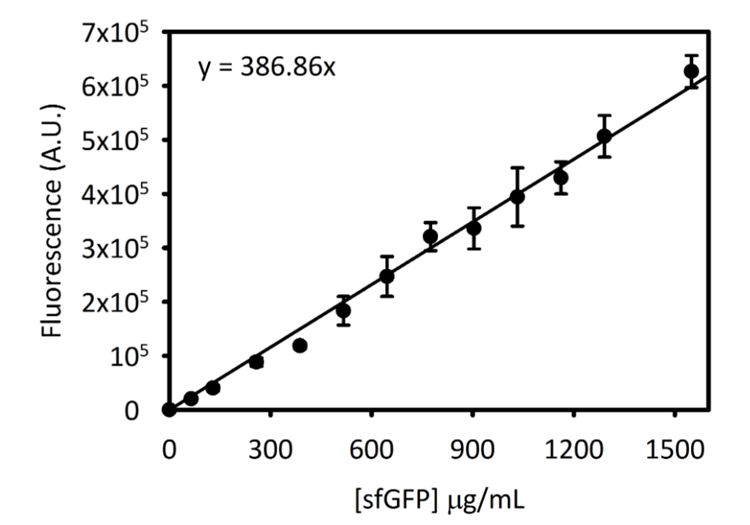
Ek şekil 3: Cytation 5 sfGFP için standart eğri. Bu eğri yukarıda belirtilen yöntemlerle tespit edilmiştir. Bu el yazması bitiş noktası Floresans okuma [sfGFP] dönüştürüldü için µg/mL standart bu eğri kullanarak toplanan tüm verileri. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Boş hücre protein sentezi çeşitli biomanufacturing biyokimyasal sistemleri hızlı prototipleme için değişen uygulamalar için güçlü ve etkinleştirme teknolojisi olarak ortaya çıkmıştır. Uygulamalar genişliğini izlemek, yönetmek ve gerçek zamanlı olarak hücresel makine çoğaltmak için kapasite tarafından desteklenmektedir. Bu platform teknolojisi genişleyen etkisi rağmen geniş adaptasyon kalmıştır yöntemleri uygulanması teknik nüanslar nedeniyle yavaş. Bu çaba ile biz bu teknoloji yeni laboratuvarları kurmak için basitlik ve netlik sağlamayı amaçlamaktadır. Bu amaçla, bizim iletişim kuralı bir E. coliiçin doğru-tabanlı boş hücre protein sentezi platform elde edilebilir dört gün açılışı içinde sigara-uzmanlar, laboratuvar eğitimi lisans öğrencileri (şekil 1) gibi tarafından. Ayrıca, reaktifler ve özü bir hazır giyim üretilen sonra sonraki CFPS toplu tepkiler ayarlayabilir, inkübe ve sadece 5 h sayılabilir. Diğer boş hücre reaktifleri tek toplu iş iş hazırlıkları reaksiyonlar binlerce destekleyebilir iken tek bir yeterli özü için dört yüz 15 µL CFPS reaksiyonlar, 1 L hücre büyümesini sonuçlanabilir. Daha büyük bir hisse senedi gerekirse reaktif hazırlıklar da ölçeklenebilir. CFPS reaksiyonlar bir yüksek-den geçerek şekilde koşullara paralel olarak çeşitli test etmek için bir 96-şey plaka veya PCR tüpleri kullanılarak kurulabilir. Volumetrik verimleri şekil 2Cgörüldüğü gibi daha küçük damarları kullanırken azalır. CFPS reaksiyonlar da microliters toplam reaksiyon birim mililitre onlarca için tek bir koşul toplam protein verimini artırmak için ölçeklenebilir. Sesini ölçeklerken birincil yüzey alanı birimi oranı reaksiyon37,52azaldıkça hacimsel tepki verimleri azaltmak noktadır. Ölçek-up protein ifade benzer hacimsel verimi korurken amacıyla, kullanıcılar tepki birimi çok sayıda tepki damarlarının bölme ve/veya gemi boyutunu artırın. 15 µL - cildinde, 100 µL ranging tepki ölçekler için çok sayıda 15 µL reaksiyonlar paralel olarak tavsiye edilir. Birimdeki 100 µL aşan tepkiler, düz-alt 24-şey plakaları tavsiye edilir ve 12-şey plakaları 600 µL aşan tepki birimleri için tavsiye edilmektedir. Reaksiyon birimleri ve damarları böyle eşleştirmelerini hacimsel tepki verimleri ölçek-up17,31,37,52üzerine tutarlılık sağlar. Bu birimler yükseltme plaka paralel olarak birden çok kuyu kullanarak gerçekleştirilebilir. Bu biçimi kullanarak, reaksiyon 10 mL Toplam hacim için ölçeklendirilebilir. Tepki birim-reaksiyonu gemi kombinasyonu en iyi duruma getirme biomanufacturing uygulamaları tepki verimlilik ödün vermeden destekleyebilir.
Bu iletişim kuralı gerçekleştirirken, hacimsel tepki verimleri ve ekstresi performansı ile düşük ilişkili göstergeleri etkisi birkaç kritik noktalar vardır. Uygun lizis sağlamak ve fonksiyonel transkripsiyon/çeviri makine denatürasyon önlemek için lizis sırasında üretilen ısı azaltmak önemlidir. Hızlı bir şekilde (şekil 3A) sonication sırasında ısı dağılımı için sonication sırasında hücre resuspension bir buz su banyosu içinde bırakın. Hücrenin lysate pre-sonicated örnekleri (şekil 3B) göre daha koyu bir görünümünü ortaya çıkması etkili hücre lizis bir göstergesidir. Kullanıcı esneklik için sonicator ve şekil 3A içinde gösterilen yoklama uyarlanabilir birimleri aralığına resuspended hücre 15 ml 100 µL üzerinden. Bunu yapmak için kullanıcı istenen birim hücre lizis için teslim Joule sayısını ayarlayabilirsiniz. Ayrıca, özü sayıda iki tamamlayıcı yaklaşımlar ile hazırlanabilir. Kullanıcılar birden çok paralel tüplerde solüsyon içeren temizleyicide veya yukarıda açıklanan29,45hacmi ile orantılı olarak enerji miktarı ölçekleme sayıda hücre resuspension, solüsyon içeren temizleyicide. Özü kalite gösterir başka bir adım aralıklarla adımdır hücre lizis takip. Sonrası hücre lizis, biz aralıklarla süpernatant (transkripsiyon/çeviri makinaları, şablon transkripsiyon/çeviri için çalışmamaya başlayan parçalanmış genomik DNA) arasında açık bir bölünme sağlamak için 18.000 x g de tavsiye ve Pelet ( Hücre zarı ve zarlarını proteinler gibi hücresel bileşenleri istenmeyen) (şekil 3 c). O Santrifüjü bulduk 18.000 x g ayırma, spin 12.000 x g gibi daha düşük hızlarda kıyasla geliştirilmiş tekrarlanabilirlik sonuçlanan geliştirir. Kolaylık sağlamak için en az 12.000 x g ulaşma yeteneğine sahip, Masa üstü soğutmalı santrifüj kullanmanızı öneririz. Bu adımı da yaygın uygun ekipman mevcut54,55,56,57,58, ise düşünülmesi gereken 30.000 x g gerçekleştirilir 59 , 60. ekstresi performansı etkilenmez bu adımı hızlarda Santrifüjü tarafından verilen bu uygun ayırma elde edilir. İstenen süpernatant kaldırırken bu kirlenme özü verimlilik azaltacaktır bu yana süpernatant ve Pelet arasındaki sınır, mevcut herhangi bir bulutlu malzemeleri kaçınmak en iyisidir. Süpernatant saflığı için amaçlayan daha üretken özler ve yeni kullanıcılar için elde edilen özü azaltılmış miktarı değer olan.
Bu iken biz sundu yöntemleri tekrarlanabilir ve en az uzmanlığı ile bilim adamları tarafından yürütülen toplu iş toplu iş olabilir Not ve reaksiyon reaksiyon varyasyon için önemlidir. Bu varyasyon lysate sonrası sonication61proteomik bileşiminde ilişkilendirilebilir. Biz gözlemledim toplu iş toplu iş değişkenlik takviyesi ile T7RNAP ve magnezyum konsantrasyonlarının duruma getirilmesi genellikle azalır. Eksojen eklenmesi T7RNAP arasında CFPS tepkiler en iyi protein ifade desteklemek için yaygındır ve bulduğumuz iki kaynak T7RNAP sahip - BL21*(DE3) ve 16 µg/mL nihai bir konsantrasyon için tamamlayıcı T7RNAP endojen ifade - geliştirir toplu iş toplu iş tekrarlanabilirlik için yeni kullanıcılar45,46. Deneyim ile isterseniz T7RNAP sadece tek bir kaynaktan yararlanmak için deneylerini kullanıcılar değiştirebilir. Miktar toplam protein içeriği özü yeni bir dizi ve Mg2 + konsantrasyonunun uygun ayarı da toplu iş toplu iş varyasyon hacimsel protein ifade verimleri azaltmak için yardımcı olabilir. Protein ifade değişimler aynı zamanda boyut ve faiz protein, geni kodon kullanımı ve onun karşılık gelen ribozom bağlama yer ilgi, hem de kullanılan ifade vektör62 tür gen yapısı farklılıkları nedeniyle olabilir ,63. Bu nedenlerden dolayı CFPS reaksiyonlar üzerinden düşük hacimsel verimi sonuçlanan modeli protein sfGFP yanı sıra bazı proteinler ifade.
Doğrudan hücre ücretsiz metabolik mühendislik ve iletişim kuralları için ek değişiklik olmadan ifade koşullardan ayarlama gibi tüm uygulamalar için uygun olmayabilir sunulan CFPS teknik sınırlamaları içerir. Ancak, bu iletişim kuralı yeni laboratuarlarında CFPS platformu oluşturmak için bir temel sağlamak ve onların Labs'de tanıtım boş hücre reaksiyonları uygulamak için uzmanlar sigara alabilmeniz inanıyoruz. İlk uygulama sonra araştırmacılar daha belirli uygulamalar diğer edebiyat alanındaki temel için kendi değişiklikler yapmak için platform deneyebilirsiniz.
(İşgücü ve ekipman maliyeti hariç), CFPS platform maliyeti $0.021/µg protein sistemimiz tepki kurulum kolaylığı ödün vermeden ticari kitleri ile rekabetçi fiyatlı yapım. Karşılaştırmalı maliyeti µL tepki ilişkin Değerlendirmeler benzer eğilimlerini (Supplemental şekil 1) gösterir. Biz olmak için başlangıç maliyeti tahmin ~ 4500 $ için tüm reaktifler ve ek 3.200 $ bir sonicator gibi özel ekipman için. Bu yordamı tamamlamak için kişi saat kadar ~ 26 h sıfırdan tüm reaktif hazırlık için olduğu tahmin edilir. Ancak, bir kez reaktifler, büyük hisse senedi hazırladık, talepleri işçilikte önemli ölçüde azaltır. Ayrıca, platformu ile deneyim kazanmış gibi biz hücre büyüme boyutu kadar ölçekleme tavsiye, hazırlık ve zaman verimliliği maksimize etmek için reaktif hazırlık ayıklamak. Başlangıç maliyeti göz önüne alındığında, biz CFPS platformu sentetik biyoloji, yüksek üretilen iş çabaları, uygulamalar için tavsiye ve protein ifade koşulları bu hücre ile çatışma geleneksel protein ifade platformları ile uyumlu değildir Biyokimya ve canlılığı kısıtlamaları. Bu özel durumlarda, nerede istenen teknik CFPS platformu tarafından etkin CFPS daha fazla maliyet vivo içinde ifade üzerinde haklı olduğunu.
CFPS platformu sürekli gelişimi biyoteknoloji çabalarına enzimatik yolları, üretimi ve karakterizasyonu geleneksel olarak dirençli proteinler, standart olmayan amino asit metabolik Mühendisliği gibi daha geniş yarar sağlar muhtemeldir birleşme ve doğal olmayan protein ifade, tabakalı ilaç üretim ve kök Eğitim64,65,66için sınıf içine laboratuvar genişletilmesi. Bu çabaların daha fazla devam eden çabaları için CFPS platformu detaylı karakterizasyonu tarafından desteklenecektir. Hücre özü kompozisyonu daha iyi anlaşılmasını sürekli iyileştirme gelişmiş tepki verimleri ve esneklik reaksiyon koşulları61,67,68doğru yol açar.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarlar onlar rakip hiçbir mali çıkarlarının veya diğer çatışmaları var bildirin.
Acknowledgments
Yazar Dr Jennifer VanderKelen, Andrea Laubscher ve Tony Turretto teknik destek için Wesley Kao, Layne Williams ve Christopher Hight yararlı tartışmalar için kabul etmek istiyorum. Yazarlar ayrıca Bill ve Linda Frost Fonu, Merkezi biyoteknoloji'nın Chevron biyoteknoloji uygulanan araştırma bağış Grant, Cal Poly araştırma, Scholarly ve yaratıcı faaliyetleri hibe programı (RSCA 2017), uygulamalar için destek fon kabul ve Ulusal Bilim Vakfı (NSF-1708919). MZL Kaliforniya Devlet Üniversitesi lisansüstü hibe eder. MCJ Ordu Araştırma Office W911NF-16-1-0372 kabul eder, Ulusal Bilim Vakfı Hibe MCB-1413563 ve MCB-1716766, Hava Kuvvetleri araştırma laboratuvarı Merkezi mükemmellik Grant FA8650-15-2-5518, savunma tehdit azaltma ajansı hibe HDTRA1-15-10052/P00001, David ve Lucile Packard Vakfı, Camille Dreyfus öğretmen-akademik Program bölümü, enerjisi BER Grant DE-SC0018249, insan sınır bilim programı (RGP0015/2017), DOE Joint genom Enstitüsü ETOP Grant ve Chicago Biyomedikal Konsorsiyumu Chicago toplum güven destek için Searle fon desteği ile.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}