

Summary
Er wordt een protocol gepresenteerd om de tijdsevolutie van het neurale meevoering op externe repetitieve stimuli te beoordelen. Steady-State opnames van dezelfde experimentele conditie worden verworven en gemiddeld in de tijd-domein. De steady-state dynamiek wordt geanalyseerd door de respons amplitude te plotten als functie van de tijd.
Abstract
Neurale meevoering verwijst naar de synchronisatie van neurale activiteit naar de perioverplaats van sensorische stimuli. Deze synchronisatie definieert het genereren van steady-state Evoked responsen (d.w.z. oscillaties in het elektroencephalogram fase-vergrendeld aan de drijvende stimuli). De klassieke interpretatie van de amplitude van de steady-state Evoked reacties veronderstelt een stereotiepe tijd invariante neurale respons plus willekeurige achtergrond schommelingen, zodanig dat het gemiddelde over herhaalde presentaties van de stimulus herstelt de stereotiepe antwoord. Deze benadering negeert de dynamiek van de stationaire toestand, zoals in het geval van de aanpassing die wordt opgewekt door langdurige blootstellingen aan de stimulus. Om de dynamiek van steady-state reacties te analyseren, kan worden aangenomen dat de tijdsevolutie van de respons amplitude hetzelfde is in verschillende stimulatie runs gescheiden door voldoende lange pauzes. Op basis van deze veronderstelling wordt een methode gepresenteerd om de tijds ontwikkeling van steady-state responsen te karakteriseren. Een voldoende groot aantal opnames wordt verkregen als reactie op dezelfde experimentele aandoening. Experimentele runs (opnames) zijn gemiddeld op kolom wijze (d.w.z. runs worden gemiddeld, maar het tijdperk binnen opnames worden niet gemiddeld met de voorgaande segmenten). De kolom-Wise middeling maakt analyse van steady-state responsen in opnames met opvallend hoge signaal-ruis verhoudingen mogelijk. Daarom biedt het gemiddelde signaal een nauwkeurige weergave van de tijdsevolutie van de steady-state respons, die kan worden geanalyseerd in zowel de tijd-als de frequentie domeinen. In deze studie, een gedetailleerde beschrijving van de methode is voorzien, met behulp van steady-state visueel Evoked mogelijkheden een voorbeeld van een reactie. Voordelen en voorbehoud worden geëvalueerd op basis van een vergelijking met enkele trial methoden ontworpen voor het analyseren van neurale entrainment.
Introduction
Wanneer opgenomen van de hoofdhuid, hersenen elektrische activiteit wordt waargenomen als continue en regelmatige veranderingen in spanningen na verloop van tijd. Deze elektrische activiteit heet elektro-encefalogram (EEG) en werd voor het eerst beschreven door Hans Berger in de late twintigers van de vorige eeuw1. Daaropvolgende zaad studies beschreven het EEG als een samengestelde tijdreeks, waarin verschillende ritmische of repetitieve patronen kunnen worden waargenomen2,3,4. Tegenwoordig is het EEG meestal onderverdeeld in vijf gevestigde frequentiebanden, Delta, theta, alpha, Beta en gamma, die worden geassocieerd met het verschillende sensorische en cognitieve proces.
Jarenlang was de studie van hersen oscillaties met behulp van EEG beperkt tot ofwel analyse van het spectrum in de lopende activiteit of veranderingen in oscillatoire activiteit opgewekt door niet-periodieke sensorische gebeurtenissen. In de afgelopen decennia zijn verschillende methodologieën geïmplementeerd voor het moduleren van lopende EEG-oscillaties en het onderzoeken van de effecten van dergelijke modulaties op perceptuele en cognitieve processen, waaronder de presentatie van ritmische sensorische stimulatie voor inducerende neurale entrainment. De term neurale meevoering verwijst naar de synchronisatie van neurale activiteit met de periodieke eigenschappen van zintuiglijke stimuli. Dit proces leidt tot de opwekking van steady-state Evoked mogelijkheden (dat wil zeggen, EEG oscillaties vergrendeld aan de periodieke eigenschappen van de drijvende stimuli). Steady-State Evoked mogelijkheden worden meestal opgewekt door visuele, auditieve en vibrotact iele stimulatie, waarbij gebruik wordt gemaakt van voorbijgaande prikkels die worden gepresenteerd met een constante snelheid of continue stimulatie, gemoduleerd in amplitude bij de frequentie van de interesse. Overwegende dat somatosensorische steady-state Evoked mogelijkheden (ssseps) worden geregistreerd als reactie op een repetitieve tactiele stimulatie van5,6, Steady-State visueel Evoked mogelijkheden (ssveps) worden over het algemeen opgewekt door de periodieke presentatie van de luminantie flikkert, Foto's en gezichten7,8. Auditieve steady-state Reacties (assrs) worden meestal gegenereerd door treinen van voorbijgaande akoestische stimuli of door de continue presentatie van amplitude-gemoduleerde tonen9,10.
De extractie van steady-state Evoked mogelijkheden uit de gemeten EEG is in wezen gebaseerd op middeling vervolgens verworven EEG tijdperken tijd vergrendeld aan de stimulus11. Vanwege de periodiciteit van de antwoorden, kunnen ze worden geanalyseerd in zowel tijd-als frequentie domeinen. Na de frequentie-domein transformatie wordt de sensorische respons waargenomen als pieken van amplitude bij de presentatie snelheid of modulatie frequentie van de externe stimuli en hun corresponderende harmonischen. Deze procedures (tijd-domein gemiddelden en de daaropvolgende frequentie-domein transformatie) zijn essentieel voor de ontwikkeling van een hoortest op basis van de detectie van ASSR-methoden met klinische doeleinden12,13,14 ,15,16.
Bovendien, de klassieke tijd-domein gemiddelde van EEG-tijdperken is zeer nuttig voor het analyseren van fysiologische processen zoals het genereren en het uitsterven van ssvep17,18geweest. Met opeenvolgende treinen van lichten en gemiddelde daaropvolgende tijdperken binnen een opname, merkte Wacker et al.19 op dat de fase-vergrendelings index van de SSVEP snel toenam tijdens de eerste 400 MS van stimulatie en daarna hoog bleef . Ze rapporteerden ook dat een robuuste visuele meevoering werd vastgesteld tussen 700-1 100 MS na het begin van de stimulus. Een zekere mate van meevoering bleef werkzaam na de offset van de stimulatie trein, die ongeveer drie perioden van de oscillatoire respons17,19duurde. Dit gedrag is geïnterpreteerd als het boeiende/ontnemende effect van de waargenomen oscillaties, wat een gevolg is van de niet-lineaire informatieverwerking in het menselijke visuele systeem17. Als alternatief, het is bekend dat onder bepaalde experimentele omstandigheden, de flikkering stimulatie kan ontlokken op-reacties aan het begin, en off-reacties aan het einde van de stimulatie treinen in plaats van neurale meevoering18.
De belangrijkste aanname aan de gemiddelde opeenvolgend verworven EEG-tijdperken is dat het EEG-signaal een lineaire combinatie van de sensorische respons en het achtergrondlawaai20vertegenwoordigt. Bovendien wordt verondersteld dat de amplitude, frequentie en fase van de oscillatoire respons stationair zijn, terwijl het achtergrondgeluid wordt beschouwd als een willekeurige activiteit. Echter, in gevallen waarin niet aan deze veronderstelling wordt voldaan, de respons amplitude berekend na verschillende tijdperken niet noodzakelijkerwijs overeenkomen met de momentane amplitude van het opgeroepen potentieel.
Het is onlangs gemeld dat de ASSR gegenereerd in de hersenstam van ratten zich aanpast aan de continue presentatie van amplitude-gemoduleerde tonen (dat wil zeggen, de respons amplitude daalt exponentieel na verloop van tijd)21,22. Aanpassing is geïnterpreteerd als een neurale mechanisme dat het verlies van nieuwheid van een monotoon repetitieve zintuiglijke stimulus weerspiegelt, waardoor de gevoeligheid voor relevante schommelingen in de akoestische omgeving wordt vergroot23,24. In de auditieve route kan aanpassing het spraakbegrip in lawaaierige omgevingen verbeteren. Bovendien kan dit proces deel uitmaken van bestaande mechanismen om de auditieve feedback van iemands eigen stem te controleren om de spraakproductie te beheersen.
Het analyseren van de tijd evolutie van de 40 Hz ASSR bij de mens, van Eeckhoutte et al.25 waargenomen een significante maar kleine afname van de respons amplitude na verloop van tijd (rond-0,0002 μv/s op basis van de groeps analyse, bij het aannemen van een lineaire afname in de tijd). Bijgevolg concludeerden deze auteurs dat de 40 Hz ASSR bij mensen zich niet aanpast aan de stimulatie. Bij de mens zijn niet-stationaire gedragingen waargenomen bij het analyseren van de stabiliteit van de SSVEP26. Deze auteurs hebben opgemerkt dat de amplitude van de fundamentele frequentie en de tweede harmonische van de SSVEP stationair waren in slechts 30% en 66,7% van de proefpersonen die ze testten. De fasen van beide SSVEP frequentiecomponenten, hoewel relatief stabiel na verloop van tijd, tentoongesteld kleine Drifts26.
Hoewel de klassieke tijd-domein middeling van vervolgens verworven tijdperken het verkennen van stationaire eigenschappen van de neurale meevoering mogelijk maakt, moet deze methodologie worden herzien wanneer de lange termijn dynamiek van het meevoering de focus is van de onderzoek, of wanneer het gemiddelde van de kortetermijndynamiek is beschadigd door het optreden van lange termijn dynamiek. Om niet-stationaire gedragingen van de steady-state reacties te karakteriseren, moet de opgeroepen reactie die wordt berekend op een bepaald tijdvenster niet worden aangetast door de antwoorden die in de voorgaande EEG-segmenten zijn berekend. Met andere woorden, het opgeroepen potentieel moet worden geëxtraheerd uit het achtergrondgeluid zonder dat tijd-domein wordt gemiddeld met de voorgaande EEG segmenten.
In deze studie, een methode voor het beoordelen van de dynamiek van neurale meevoering wordt gepresenteerd. Steady-State responsen worden herhaaldelijk opgenomen in reactie op dezelfde stimulatie, waarbij opeenvolgende opnames Interleaved zijn door een rustinterval van drie keer de lengte van de experimentele run. Gezien het feit dat als de tijd evolutie van de fysiologische respons hetzelfde is in verschillende onafhankelijke experimentele runs (onafhankelijke opnames), zijn opnames kolom-verstandig gemiddeld. Met andere woorden, de tijdperken die overeenkomen met dezelfde locatie in de verschillende opnames worden gemiddeld, zonder gemiddelde tijdperken binnen een opname. Bijgevolg zal de respons amplitude die bij een stimulatie interval wordt berekend, overeenstemmen met de momentane amplitude van het opgeroepen potentieel. De zintuiglijke reacties kunnen worden geanalyseerd in de tijd-domein of omgezet in de frequentie-domein, afhankelijk van het doel van het experiment. In ieder geval kunnen de amplituden worden uitgezet als een functie van de tijd om de tijd evolutie van de steady-state respons te analyseren. Het genereren en uitsterven van de steady-state Evoked mogelijkheden kan worden beoordeeld door de analyse te beperken tot de eerste en laatste tijdperken van de opnames.
De dynamiek van de neurale meevoering kan worden geanalyseerd met behulp van andere benaderingen, zoals smalbandfiltering enkelvoudige proef metingen rond de frequentie van belangstelling en het berekenen van de envelop van het stroomsignaal met behulp van low-pass filtering25 en de Hilbert-transformatie27. In vergelijking met deze methodologieën maakt de kolom-Wise middeling van tijdperken het mogelijk om stationaire parameters te berekenen op basis van signalen met de hogere signaal-ruis verhouding (SRV). Onlangs is Kalman filtering ontstaan als een veelbelovende techniek voor de schatting van 40-Hz ASSR amplitudes28,29,30. De implementatie van Kalman filtering kan de detectie van steady-state responsen dichter bij de elektrofysiologische drempel verbeteren en de tijd van de gehoortest29verminderen. Bovendien zijn stationaire responsen niet nodig om te worden verondersteld wanneer een Kalman filtering benadering wordt gebruikt om de ASSR amplitude30te schatten. Niettemin heeft slechts één studie de tijd evolutie van ASSRs geanalyseerd met Kalman filtering25. De conclusie van de studie is dat de 40-Hz ASSR amplitude stabiel is over het stimulatie interval. Daarom moet Kalman filtering worden getest onder omstandigheden waaronder de ASRR niet stilstaat.
Hoewel tijdrovend, de kolom-Wise middelaging methode is model-vrij en hoeft geen initialisatie waarden en/of a priori definities van het lawaai gedrag. Bovendien, aangezien het geen convergentie tijden inhoudt, kan het gemiddelde van de kolom wijzen een betrouwbaardere weergave van het ontstaan van neurale entrainment bieden. Daarom kunnen de resultaten die met de kolom-Wise Middelings methode worden verkregen, worden beschouwd als de grond waarheid voor het analyseren van de dynamiek van het neurale meevoering met behulp van Kalman-filtering.
Deze beschrijving van het protocol is gebaseerd op een voorbeeld van SSVEP. Echter, het is belangrijk op te merken dat de hier gepresenteerde methode is modaliteit-onafhankelijk, zodat het kan ook worden gebruikt voor het analyseren van de tijd evolutie van SSSEP en ASSR.
Protocol
De huidige studie werd uitgevoerd onder goedkeuring van het onderzoeks-en ethisch comité van de Universidad de Valparaíso, Chili (beoordelings verklaring code CEC170-18), bevestigd aan de nationale richtlijnen voor onderzoek met menselijke proefpersonen.
1. voorbereiding
- Verwelkom het onderwerp.
- Leg de doelstellingen en relevantie van de studie uit. Geef een beschrijving van de relevante technische details. Beantwoord alle vragen grondig.
- Expliciet vermelden dat zij/hij de experimentele sessie op elk gewenst moment kan onderbreken indien gewenst.
- Vraag de vrijwilliger om het onderwerp geïnformeerde toestemming te lezen en het bijbehorende formulier te ondertekenen. De experimentele sessie onderbreken als de geïnformeerde toestemming niet wordt verkregen.
2. voorbereiding van het onderwerp
- Vraag het onderwerp om in een comfortabele positie in een laboratorium stoel te zitten.
- Reinig de hoofdhuid met ethanol (een oplossing bij 95%) om de laag dode huidcellen en talg die het bedekken te verwijderen. Deze stap is belangrijk om de impedantie tussen de elektroden en de hoofdhuid te verminderen.
- Meet de hoofd omtrek met een meetlint om de grootte van de te gebruiken elektrode dop te bepalen.
- Vraag het onderwerp om de elektrode dop te dragen. Geef de instructies voor een comfortabele, maar correcte positionering van de dop.
- Meet de afstand tussen de neus (NZ, het middelste punt van de nasofrontale hechting, die kan worden geïdentificeerd door de depressie tussen de ogen en de bovenkant van de neus) en de INION (iz, de prominentie van het occipitale bot) met behulp van een meetlint.
- Meet de afstand tussen de linker-en rechter Pre-auriculaire punten (geïdentificeerd als de depressie net voor de oorschelp van de oren) met behulp van een meetlint.
- Corrigeer de positie van de elektrode dop, zodat de doorsnede tussen de imaginaire lijnen die in de vorige stappen zijn gedefinieerd, overeenkomt met het hoekpunt van het hoofd. Zorg ervoor dat het onderwerp comfortabel is na de aanpassingen.
- Plaats geleidende gel in de elektrode houders, volgens de locaties die voor het experiment worden overwogen.
- Gebruik 64 hoofdhuid locaties na het International 10-20 System31 om de uitkomst van het protocol te gebruiken om bron lokalisatie analyse uit te voeren. Het hogere aantal elektrode locaties (128) op de hoofdhuid kan indien nodig worden gebruikt.
- Implementeren van klinische of ambulante instellingen (met slechts een paar elektroden) als bron lokalisatie analyse is niet gepland. Gebruik occipitale locaties voor het opnemen van SSVEP, tijdelijke locaties voor het verwerven van ASSR en pariëtale locaties voor het opnemen van SSSEP.
- Duw de elektroden in de elektrode houders. Zorg ervoor dat het label van de elektrode overeenkomt met het label van de locatie in de dop.
- Begeleiden de vrijwilliger naar de experimentele kamer (bij voorkeur, een afgeschermde, geluiddempd kamer). Vraag het onderwerp om in een stoel in de kamer te zitten, in een comfortabele positie.
- Plaats externe elektroden op de neus en oorlellen als een fysieke Referentie (verschillend van de hoofdhuid elektroden) zal worden gebruikt voor het opnieuw verwijzen naar de EEG-opname (in stap 3.8.1).
- Plaats externe elektroden in peri oculaire locaties.
- Plaats de elektroden op de Wang en de voorste regio van het hoofd, ongeveer 1 cm boven de wenkbrauw, om op te nemen knipperen (in stap 2.6.1).
- Plaats de elektroden op de buitenste kanthus van de ogen, ongeveer 1 cm boven/onder de middenlijn, om oogbewegingen op te nemen (in stap 2.6.1).
Opmerking: het electrooculogram (EOG) zal worden gebruikt in de stap 3.8.5 voor het verwijderen van EIARTEFACTEN geïnduceerd door knipperen en oogbewegingen.
- Schakel het EEG-acquisitie systeem in en controleer de elektrode impedantie als een systeem met lage impedantie wordt gebruikt voor het opnemen van het EEG. Corrigeer de impedantie indien nodig volgens de aanwijzingen van de fabrikant. De impedantie moet onder 10 kΩ 32 worden gehouden.
- Vraag het onderwerp om te knipperen en beweeg de ogen in verschillende richtingen om ervoor te zorgen dat EOG correct wordt opgenomen.
- Om de dynamiek van SSVEP te analyseren, past u de locatie van het scherm in de verticale richting aan, zodat deze overeenkomt met de weergavehoek van het onderwerp. Dim de lichten van de kamer tot een comfortabel niveau wordt bereikt. Stel het luminantieniveau van het scherm in op de bovengrens van het comfortniveau van de deelnemer.
- Om de dynamiek van ASSR te analyseren, plaatst u de oortelefoon met de juiste schuim wisselplaten, zodat de oortelefoon op de gehoorgang past. Controleer of geluiden worden afgeleverd op de gewenste intensiteit (bijv. een psychofysische comfortabel niveau33).
3. EEG overname en voor verwerking
- Stel de stimulerende parameters in die zijn gedefinieerd in het experimentele ontwerp. Raadpleeg de gebruikershandleiding die door de fabrikant van het stimulatie systeem is verstrekt voor meer informatie over de software.
Opmerking: voor uitgebreide uitleg van de stimulus gebruikt voor het genereren van SSVEP en ASSR, zie Norcia et al.8 en Rance34, respectievelijk. - Instrueer het onderwerp om aandacht te besteden aan de stimulatie, in het geval dat visuele meevoering het onderwerp van het experiment is.
- Presenteer een ondertitelde film met het geluid uit wanneer auditieve meevoering het onderwerp van het experiment is.
Opmerking: presentatie van een stomme film maakt het mogelijk om de aandacht van de akoestische stimulatie te trekken terwijl het opwinding niveau25wordt gehandhaafd.
- Presenteer een ondertitelde film met het geluid uit wanneer auditieve meevoering het onderwerp van het experiment is.
- Presenteer stimuli langer dan 90 s, zoals is gedaan om de tijd evolutie van ssveps en ASSR in zowel mens en diermodellen21,22,25,26te onderzoeken.
Opmerking: presenteer stimuli korter in duur als een pilot-studie is uitgevoerd. - Pauzeer de stimulatie gedurende 2 minuten als er slechts één experimentele aandoening wordt getest. Communiceer met het onderwerp om het bewustzijn te controleren.
Opmerking: de duur van de pauze is afhankelijk van de duur van de stimulatie. Pauzeert 3x langer dan de stimulatie intervallen zorgt ervoor dat een respons die wordt opgewekt door een stimulus niet wordt beïnvloed door de vorige stimulatie. Langere pauzes zijn toegestaan als het onderwerp daarom verzoekt.- Pauzeer de stimulatie gedurende ten minste 10 sec. Wanneer verschillende experimentele omstandigheden worden getest, aangezien alternerende stimulatie met pauzes van 10 s is voorgesteld om extra aanpassings effecten te verminderen en de lengte van het experiment te reduceren25.
- Herhaal de presentatie stappen (stappen 3.3-3.4) ten minste 30x om te zorgen voor de hoge SNR van de metingen na het gemiddelde van de tijdperken (stap 4,4).
- Registreer het EEG met behulp van de standaardprocedures35. Maak een afzonderlijk EEG-bestand voor elke experimentele uitvoering.
Opmerking: Raadpleeg de gebruikershandleiding van het acquisitie systeem voor meer informatie over de software. - Bewaak de EEG-opname om slaap perioden te detecteren op basis van het niveau van de Alfa-activiteit en de frequentie waarmee knipperende artefacten worden weergegeven. Pauzeer het experiment wanneer verhoogde Alfa niveaus gepaard gaan met verlaagde knipperende frequenties worden gedetecteerd, wat indicatief is voor slaperigheid. Weiger de experimentele uitvoering van verdere analyse wanneer slaap perioden worden gedetecteerd.
- Bereken de amplitude van de steady-state respons aan het einde van elke experimentele uitvoering, volgens de instructies in de gebruikershandleiding van de acquisitie software die in het experiment wordt gebruikt.
- Bewaak het aandachtsniveau van het onderwerp door de amplitude van de steady-state responsen te vergelijken die aan het einde van elke experimentele run zijn behaald. Stel de steady-state amplitude die is verkregen in de eerste experimentele runs als een referentie-amplitude.
- Stel een afwijzings drempel in (een afname van de respons amplitude van 5% met betrekking tot de referentie-amplitude). Verwerp de experimentele runs waarin de amplitude van de steady-state respons voldoet aan het afwijzings criterium.
- Voltooi de experimentele sessie na het verkrijgen van het aantal runs dat is gedefinieerd in het experimentele ontwerp.
- Pre-process de EEG gegevens offline met behulp van standaard EEG procedures35 beschreven in de volgende stappen volgens de instructies van de fabrikant.
- Herverwijs de opname met een gemiddelde referentie (gemiddelde van alle opname elektroden) of het gemiddelde van een subset van elektroden. U ook een fysieke referentie gebruiken (bijv. externe elektroden die op de neus en oorlellen worden geplaatst zoals beschreven in stap 2,4).
- Converteer de elektrode coördinaten naar het internationale 10/20-systeem als het radiale coördinatensysteem werd gebruikt tijdens de EEG-overnames. Raadpleeg de handleiding van de fabrikant voor meer informatie over de conversie.
- Band-pass filter het EEG-signaal tussen 0,5-300 Hz. Stel, indien nodig, een inkeping-filter in (gecentreerd op 50 Hz of 60 Hz).
- Down-sample het EEG-signaal om de uitvoeringstijd van het algoritme dat is geselecteerd voor het verwijderen van oculaire artefacten te verminderen (stap 3.8.5).
Opmerking: een bemonsteringsfrequentie van 512 Hz is voldoende om de hersen oscillaties van de frequentie onder 40 Hz35te analyseren. - Verwijder de oculaire artefacten.
Let op: hiervoor kunnen verschillende technieken worden gebruikt (Zie Urigüen en Garcia-Zapirain35 voor een uitgebreide beoordeling van algoritmes voor het verwijderen van artefacten). Onder hen is onafhankelijke component analyse een van de meest uitgebreide methodologieën en wordt geïmplementeerd in zowel commerciële als gratis analyse software37,38,39. - Segmenteer de EEG-gegevens in tijdperken vergrendeld op stimulatie. Selecteer de lengte van de tijdperken volgens het doel van het experiment.
NB: de tijdperken moeten voldoende lang zijn om de steady-state respons in het frequentiedomein te kunnen analyseren met een adequate spectrale resolutie. - Voer geen artefact afwijzing algoritmen in deze fase om te detecteren en verwijderen van tijdperken met artefacten.
Opmerking: het verwijderen van Epoch in deze fase zal fouten veroorzaken wanneer de gegevensset is georganiseerd voor het uitvoeren van het kolom-verstandig middeling van tijdperken (stappen 4,2 en 4,4). Afwijzings algoritmen worden geïmplementeerd bij een latere verwerkingsstap (stap 4.1.4). - Voer de DC-detrend functie voor het berekenen van DC-trends in afzonderlijke EEG-tijdperken en corrigeer deze.
- Voer de basislijn correctiefunctie uit om de basislijn van de opname te corrigeren. Selecteer tijdintervallen van vóór de stimulus langer dan 200 MS.
Opmerking: basislijn correctie bestaat uit het gemiddelde van de gegevens in het geselecteerde tijdsinterval. Het gemiddelde wordt berekend voor elk kanaal en afgetrokken van elk gegevenspunt in elk tijdperk.
4. berekening van de respons amplituden
- Voer de parameters in die nodig zijn voor de berekening van de steady-state responsen (Figuur 1a).
Opmerking: de in-House code die wordt gebruikt voor het verwerken van de gegevens is vrij beschikbaar op < https://62573 >. Raadpleeg de Help-tekst in de code voor verdere instructies. Evenzo is een subset van de gegevens die in deze studie worden gebruikt beschikbaar.- Voer het aantal opnamen (experimentele runs) van het experiment in.
- Voer de lengte van de tijdperken in om de afzonderlijke opnamen te segmenteren.
- Voer de bemonsteringsfrequentie van het experiment in.
- Selecteer algoritmen voor het afwijzen van artefact om te detecteren en verwijderen van tijdperken met artefacten. De beschikbare selectiecriteria zijn 1) gradiënt (absoluut verschil tussen twee opeenvolgende monsters), 2) max-min (het verschil tussen de maximale en minimale amplitude in het tijdperk), en 3) amplitude (absolute maximale en minimale amplituden).
- Voer de verwerkings code uit.
Opmerking: stappen 4.2-4.7 worden automatisch uitgevoerd wanneer deze optie is geselecteerd. Voer de stappen zo nodig handmatig uit.
- Regel de tijdperken opnieuw in een gegevensmatrix van n rijen en m kolommen, waarin n het aantal opnamen (experimentele runs) en m het aantal tijdperken (Figuur 1b) weergeeft.
- Gewicht van de epoche te verzachten van het effect van beweging en gespierde artefacten.
Opmerking: gewogen EEG-tijdperken worden verkregen door elk voltage monster te delen door de amplitude variantie van het tijdperk waartoe ze behoren, zodat variantie wordt gebruikt als een maat voor amplitude variabiliteit en weegfactor40. - Kolom-Wise gemiddelde van de gegevensset. Om dit te doen, gemiddelde time-domein de tijdperken die overeenkomen met hetzelfde tijdvenster in de verschillende opnames.
Opmerking: deze stap laat de berekening van de steady-state amplitude toe in opnames met een opmerkelijk hoge signaal-ruis verhouding (SRV). - Exporteer de tijdreeksen die voortvloeien uit het gemiddelde voor verdere analyse van de tijdsevolutie van de meevoering in externe software.
- Bereken de amplitude van de steady-state respons in elk tijdperk als gevolg van de kolom-verstandig middeling, met behulp van de snelle Fourier Transform (FFT).
Opmerking: de lengte van de FFT moet overeenkomen met de lengte van één tijdperk. De implementatie van een windowing techniek is niet verplicht. De amplitude van de steady-state respons wordt gedefinieerd als de spectrale amplitude die is verkregen bij de frequentie van de amplitudemodulatie van de sensorische stimuli.
- Bereken de amplitude van de steady-state respons in elk tijdperk als gevolg van de kolom-verstandig middeling, met behulp van de snelle Fourier Transform (FFT).
- Vector gemiddelde de amplitude van een ad-hoc aantal FFT-bakken aan elke kant van de frequentie van de respons om het rest geluidsniveau te berekenen (RNL). Het aantal FFT-bakken moet overeenkomen met een frequentieband van ongeveer 3 Hz, aan elke zijde van de frequentie van de respons.
Opmerking: de hoge frequentie-specificiteit van de steady-state responsen maakt de respons amplitude onafhankelijk van die achtergrond oscillaties met vergelijkbare frequenties, die op zijn beurt gelijkmatig verdeelt in een relatief smalle frequentieband41 , 42 , 43. - Plot de amplitude van de steady-state respons en de RNL als een functie van kolomindex (d.w.z. het nummer van het verworven tijdperk) om de evolutie van de steady-state respons te verkennen tijdens het stimulatie interval.
Representative Results
SSVEP werd opgewekt door continue visuele stimuli van 40 s in lengte, waarbij de lichtintensiteit werd gemoduleerd door een sinusvormige Golf van 10 Hz (Modulatiediepte van 90%). Stimuli werden geleverd door vier lichtgevende diodes (leds) gelegen in het midden van een 50 cm x 50 cm zwart scherm, als hoekpunten van een 5 cm x 5 cm vierkant. Wanneer de deelnemer zat 70 cm van het scherm, de oppervlakte van het plein van de LEDs subneigt een visuele hoek van ongeveer 4 °. Het LED-scherm is ontworpen met behulp van een USB-gebaseerde microcontroller ontwikkelsysteem en vier super helder witte Led's van 10 mm diameter. De Pulsbreedtemodulatie (PWM)-techniek werd gebruikt om de aan de Led's geleverde voeding te regelen. Deze techniek beheerste de LEDs intensiteiten bij een bepaalde frequentie en het genereren van de uiteindelijke sinusvormige envelop. Een PWM-frequentie van 40 kHz werd gebruikt om een waarneembaar effect te voorkomen.
Er werden dertig opnames verkregen, die werden gesegmenteerd in tijdperken van 4 sec. Daarom is een gegevensset die bestaat uit 10 kolommen (aantal EEG-tijdperken binnen opnames) en 30 rijen (aantal opnamen, aantal experimentele runs) verkregen.
De neurale oscillatie tijd-vergrendeld aan de stimulatie werd duidelijk als de kolom-Wise Middelings werd uitgevoerd (Figuur 2). Significant, het interval waarop de SSVEP wordt gegenereerd kan worden waargenomen in traces die overeenkomen met kolom 1. In die kolom, 0,2 s van pre-stimulus basislijn worden uitgezet naast de eerste 0,8 s van neurale entrainment. Daarom kan de hier beschreven procedure karakteriseren van 1) de dynamiek van de oscillatoire respons zodra neurale meevoering al is vastgesteld en 2) de betrokkenheid van neurale oscillaties. Een of meer tijdperken die na het einde van de stimulatie zijn opgenomen, kunnen ook in de gegevensmatrix worden opgenomen om uitsterven van de steady-state respons te bestuderen na verschuiving van de stimulus.
Tijdens de kolom-Wise middeling van tijdperken, daalde de gemiddelde amplitude van de SSVEP (spectrale amplitude bij 10 Hz, berekend door toepassing van de FFT) tijdens het gemiddelde van de eerste tijdperken van de kolommen en de neiging om daarna te stabiliseren (Figuur 3a). Dit resultaat stemt overeen met eerdere studies die de evolutie van ASSR analyseren tijdens het gemiddelde van opeenvolgend verworven tijdperken21,22,40,43,44. Het gedrag van de amplitude van de respons tijdens het gemiddelde wordt meestal verklaard door de relatief hoge bijdrage van ongemiddeld geluid aan de respons amplitude berekend in de eerste tijdperken, die wordt verzwakt naarmate het gemiddelde wordt uitgevoerd13, 44 , 45 , 46 , 47. opmerkelijk, de SSVEP amplitude variabiliteit daalde significant naarmate het gemiddelde vorderde.
We analyseerden ook de RNL van de metingen tijdens de kolom-Wise middeling van de tijdperken (Figuur 3b). De RNL werd berekend in een smalle frequentieband (3 Hz) aan beide zijden van de frequentie van de SSVEP. Hoewel deze procedure niet gebruikelijk is wanneer ssvep wordt geanalyseerd, is vector-middelaging een gegeven aantal frequentie bakken rond die van de neurale meevoering de standaard voor het inschatten van de RNL in ASSR-metingen41,42, 43. zoals verwacht nam de RNL geleidelijk af naarmate het aantal gemiddelde tijdperken toenam en het asymptotische niveau bereikte nadat er ongeveer 20 tijdperken werden verwerkt. In tegenstelling tot wat werd waargenomen toen de SSVEP-amplitude werd geanalyseerd, bleef de standaarddeviatie van de RNL relatief constant, omdat het aantal gemiddelde tijdperken toenam, wat suggereert dat de opnamecondities stabiel waren tijdens de experimentele sessie.
De hierboven gepresenteerde resultaten bepaalden de veranderingen in de piek signaal-ruis verhouding (pSNR) van metingen tijdens de kolom-Wise middeling van het tijdperk (Figuur 3C). Deze term wordt hier gedefinieerd als de ratio (in dB) tussen de vierkante amplitude van de respons (SSVEP) en de vierkante amplitude van de RNL. Naarmate het gemiddelde vorderde, steeg de psnr naarmate het aantal gemiddelde tijdperken steeg tot 18, ongeveer. Verdere verhogingen in het aantal gemiddelde tijdperken hadden geen significant effect op de kwaliteit van het signaal. De variabiliteit van de pSNR daalde naarmate meer tijdperken werden gemiddeld.
Ten slotte worden de dynamiek van de SSVEP-amplitude en de RNL weergegeven in Figuur 4. Deze tijd evoluties werden verkregen door het uitzetten van de reactie parameters berekend aan het einde van de kolom-verstandig middeling van tijdperken als een functie van het aantal kolommen (als functie van de tijd). Zoals aangetoond door Labecki et al.26, kan de dynamiek van SSVEP aanzienlijk variëren tussen onderwerpen. Aangezien de in Figuur 4 gepresenteerde resultaten overeenkomen met één individu, kunnen generalisaties niet worden gemaakt. In dit onderwerp, de amplitude van de SSVEP weergegeven een relatief complex gedrag (figuur 4a). De respons amplitude nam geleidelijk toe tijdens de eerste 12 seconden na het begin van de stimulus (tijd die overeenkomt met de lengte van 3 Epochs). Naarmate de stimulus bleef, daalde de SSVEP consequent gedurende de volgende 12 seconden en bleef daarna relatief constant. Deze resultaten kunnen niet worden verklaard door het gedrag van de RNL, omdat deze parameter relatief constant was tijdens het stimulatie interval (figuur 4b). De toename van de SSVEP-amplitude na het begin van de stimulus blijkt uit de sporen in Figuur 2 en kan worden verklaard door integratieprocessen, wat resulteert in stabilisatie van de neurale entrainment. De daaropvolgende afname van de amplitude suggereert de aanpassing van SSVEP aan de aanhoudende stimulatie. Niettemin moeten deze hypotheses worden getest in gecontroleerde experimenten met toegewezen steekproefgrootten.
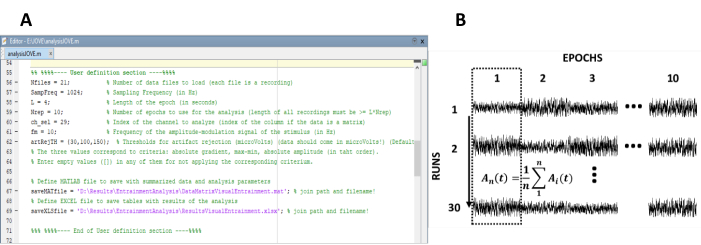
Figuur 1 : Kritieke stappen voor het extraheren van de tijdsevolutie van de amplitude van steady-state responsen. (A) Screenshot van de verwerkings code, waarbij analyseparameters zijn gedefinieerd. B) representatief diagram dat de organisatie van de DataSet illustreert. Een data matrix bestaande uit 30 opnames van 10 tijdperken is vertegenwoordigd. De kolom-Wise middeling van tijdperken wordt in de eerste kolom gemarkeerd. De verticale lijn geeft de richting van het gemiddelde aan. Klik hier om een grotere versie van dit cijfer te bekijken.

Figuur 2 : Veranderingen in de golfvorm van steady-state visueel Evoked mogelijkheden (ssvep) tijdens de kolom-Wise middeling van tijdperken. De respons werd opgewekt door de continue presentatie van licht gemoduleerd in amplitude bij 10 Hz. De rijen tonen de golfvormen verkregen na het gemiddelde van alle voorgaande opnames (dat wil zeggen, rij 1 is de eerste opname, rij 5 is de golfvorm verkregen na het gemiddelde van de eerste vijf opnames, en de laatste rij is het gemiddeld van alle opnames). Meer betrouwbare golfvormen van SSVEP werden waargenomen in elke kolom als het aantal Middelings runs verhoogd. Om duidelijkheid te verschaffen (om de oscillaties van de SSVEP zichtbaar te maken), wordt alleen de eerste seconde van de tijdperken weergegeven. De uitzonderingen zijn sporen in de eerste kolom van de gegevensset, waarvoor 0,2 seconden van pre-stimulus basislijn worden weergegeven. Klik hier om een grotere versie van dit cijfer te bekijken.

Figuur 3 : Veranderingen in de respons-en opnameparameters tijdens de kolom-Wise middeling van tijdperken. A) evolutie van de amplitude van de SSVEP. B) gedrag van de RNL. (C) wijzigingen in de psnr. Zwarte lijnen vertegenwoordigen de gemiddelde waarden die voor elke kolom zijn verkregen (n = 10) en de grijze schaduw vertegenwoordigt het gebied dat wordt bedekt door een standaarddeviatie van ± één. Klik hier om een grotere versie van dit cijfer te bekijken.

Figuur 4 : Tijd evolutie van de SSVEP opgewekt door de presentatie van continue visuele stimulatie, gemoduleerd in amplitude bij 10 Hz. (A) tijdsverloop van de amplitude van de SSVEP. B) tijdsverloop van de RNL. Klik hier om een grotere versie van dit cijfer te bekijken.
Discussion
Dit werk beschrijft een experimentele procedure voor het analyseren van de dynamiek van oscillerende hersen responsen. Een dergelijke methodologie bestaat uit het verwerven van een voldoende aantal onafhankelijke experimentele runs van dezelfde experimentele toestand, en tijd-domein Middelings tijdvakken overeenkomend met hetzelfde tijdvenster in de verschillende opnames (kolommen-Wise middeling in Figuur 1B). De amplitude berekend in de gemiddelde gegevens vertegenwoordigen de momentane amplitude van de oscillatoire respons. Het uitzetten van deze amplituden als functie van de tijd (of het aantal kolommen in de dataset) maakt het mogelijk om de tijdsevolutie van de oscillatoire responstijd te analyseren-vergrendeld aan de stimulatie. Deze methodologie is een wijziging van die voorgesteld door Ritter et al.23 voor het analyseren van de aanpassing van voorbijgaande corticale evoked potentials. De methode is gebruikt voor het analyseren van de dynamiek van auditieve Evoked mogelijkheden in beide mensen24 en dierlijke modellen20,21.
Vanuit methodologisch oogpunt is de combinatie van parameters die worden gebruikt om de steady-state respons te wekken en die geïmplementeerd om de neurale respons van achtergrondruis te extraheren, essentieel om de tijd evolutie van steady-state Evoked mogelijkheden te analyseren 22. de stimulus lengte die in het hier gepresenteerde experiment werd gebruikt (40 s) werd geselecteerd op basis van resultaten die in een pilot studie werden behaald. Deze stimulus-lengte was voldoende voor het analyseren van de aanpassing van ASSR gegenereerd in de rat hersenstam21,22. Bovendien moet de stimulus-lengte de tijd overschrijden waarop de asymptotische momentane band kracht van SSVEPs wordt bereikt (Figuur 1 in Labecki et al.26). Niettemin, de asymptotische momentane band macht van SSVEPs kan worden bereikt dan 60 in sommige gevallen (Figuur 2 in Labecki et al.26). Daarom wordt het uitvoeren van een pilot-studie met een klein monster aanbevolen om de stimulus lengte van de stimulatie te definiëren. Anders wordt een stimulerende lengte langer dan 90 s aanbevolen om een volledige weergave van de tijdsevolutie van de respons te bereiken. Het gebruik van voldoende lange pauzes tussen opeenvolgende opnames impliceert het overwegen van opeenvolgende experimentele runs om statistisch onafhankelijk te zijn (d.w.z. verschillende, onafhankelijke maatregelen van dezelfde variabele). Naar het beste van onze kennis zijn er geen experimenten uitgevoerd om de optimale pauze tussen runs te analyseren (minimale pauze vereist om runs onafhankelijk van elkaar te maken). Het criterium van het gebruik van pauzes ten minste 3x langer dan de stimulus-lengte is conservatief genoeg om ervoor te zorgen dat de steady-state respons opgenomen in een gegeven run niet wordt beïnvloed door de vorige stimulatie.
Onlangs zijn alternerende stimuli (experimentele omstandigheden) voorgesteld als een keuze om de pauze tussen experimentele runs te verminderen, waardoor extra adaptatie effect25wordt vermeden. Evenzo, het aantal experimentele runs (30) geïmplementeerd in dit experimentele protocol is conservatief, omdat de asymptotische RNL en pSNR meestal worden bereikt na gemiddeld 20 experimentele runs, ongeveer. Wanneer prikkels binnen het midden-bovenste gebied van het dynamische bereik van de respons vallen (hoge sensatie niveaus), zijn lagere aantallen runs waarschijnlijk nodig om de dynamiek van de Evoked Response te analyseren. Niettemin is, in gevallen waarin verschillende experimentele omstandigheden worden getest, het hebben van hetzelfde aantal experimentele runs cruciaal voor het maken van vergelijkingen onder omstandigheden (d.w.z. verschillende sensatie niveaus).
Naast de kolom-Wise middeling van tijdperken, is de dynamiek van oscillerende Evoked mogelijkheden geanalyseerd door het filteren van de enkelvoudige metingen in een smalle frequentieband rond de frequentie van belangstelling en het berekenen van de envelop van het vermogen signaal met behulp van low-pass filtering26. Evenzo is er één proef analyse geïmplementeerd om de overgangsperiode te karakteriseren die voorafgaat aan de stabiele regio van SSVEP48, en de veranderingen in amplitude en fase van de ssvep tijdens de stabiele regio van de respons49. Hoewel enkelvoudige proef analyses discriminatie van relatief snelle fluctuaties in respons amplitude mogelijk maken, kunnen experimentele ontwerpen om de gemiddelde respons te analyseren in blokken die gescheiden zijn met een bepaald interval tussen de blokkeringen alleen rekening worden gehouden met lange termijn variaties in de amplitude van de opgeroepen potentiaal50,51. De kolom-Wise middeling van tijdperken staat tussen deze twee opties. Converteren van het gemiddelde signaal naar de frequentie-domein met behulp van de FFT impliceert het analyseren van de dynamiek van de respons met een resolutie die gelijk is aan de lengte van het tijdperk. In het voorbeeld hier gepresenteerd, de SSVEP werd gerapporteerd elke 4 s. Hoewel 4 s van resolutie is voldoende om te beschrijven van de dynamiek die zich voordoet met intervallen van tijd te overtreffen tientallen seconden, zoals die van de SSVEP26, gedeeltelijk overlappende tijdperken in de originele opnames beschrijven de tijdsevolutie van de Steady-State respons op een meer verfijnde manier25.
Dynamiek van de steady-state reacties verkregen na kolom-Wise middeling van tijdperken vertegenwoordigen voornamelijk de evolutie van de oscillerende activiteit die wordt gesynchroniseerd tussen de gemiddelde EEG segmenten (die die het gemiddelde overleven). Daarom is een belangrijk probleem met betrekking tot de haalbaarheid van de methodologie is de mogelijke verzwakking van respons amplituden als gevolg van variaties in de fase van neurale oscillaties van een onafhankelijke experimentele run naar de andere (dat wil zeggen, tussen opnames). Dit onderwerp moet experimenteel worden aangepakt. Echter, bewijs geeft aan dat de fase van de hersenen oscillerende reacties is minder variabel dan verwacht. In feite, verschillende studies hebben gemeld een regelmatigheid in de verwachte fase van de menselijke 80 Hz ASSR47,48,49. Wanneer latenties worden geschat op basis van de fase van de oscillerende activiteit, is het voorspelbare effect van de intensiteit en de draagfrequentie van de akoestische stimuli op de latentie van de auditieve responsen waargenomen (d.w.z. de latentie afname als de verhoging van de intensiteit en de draagfrequentie)52,53,54. Bovendien zijn typische maturatie veranderingen in amplitude en de links-naar-rechts asymmetrie in het gehoor niveau ook waargenomen wanneer latenties worden geschat uit de fase van de ASSR47,55,56 , 57 , 58. bij het beschrijven van de tijd evolutie van SSVEP met behulp van enkelvoudige analyse, merkte labecki et al.26 op dat hoewel de variabiliteit van de respons amplitudes binnen hetzelfde onderwerp aanzienlijk hoog was, de variabiliteit van de fase significant minder uitgesproken.
Op basis van hun waarnemingen suggereerde Labecki et al.26 dat een minimum van 50 proeven moet worden gemiddeld om een betrouwbare schatting te verkrijgen van de gemiddelde vermogens enveloppe van de respons. Deze resultaten geven aan dat, zelfs wanneer de amplitude van de respons wordt berekend in één proef, het gemiddelde (van enveloppen in dat geval) nodig is om betrouwbare resultaten te rapporteren. Bovendien suggereert de intertrial variabiliteit in de amplitude van SSVEP gerapporteerd door Labecki et al.26 suggereren dat de berekening van deze parameter in enkelvoudige proeven sterk kan worden beïnvloed door achtergrondruis. Gezien de evolutie van de signaal-ruis verhouding zoals weergegeven in Figuur 2, vermindert de berekening van de respons in het gemiddelde signaal in plaats van enkelvoudige proeven aanzienlijk het aantal EEG-segmenten dat moet worden verwerkt voor het verkrijgen van betrouwbare Metingen. Bovendien, de lage variabiliteit in fase verkregen door labecki et al.26 steunt het idee dat de kolom-Wise middeling van de tijdperken gepresenteerd hier is een geldige procedure voor het berekenen van de dynamiek van oscillerende evoked potentials.
Het gemiddelde van de gegevens op verschillende niveaus leidt tot een verschillende interpretatie van de resultaten. Met betrekking tot oscillerende evoked potentials, het berekenen van de respons amplitude na de tijd-domein middeling van onafhankelijke runs impliceert het analyseren van alleen tijdvergrendelde oscillaties (dat wil zeggen, degenen die het gemiddelde overleven). Deze procedure kan relevante informatie met betrekking tot de dynamiek van de respons in individuele proeven filteren. Het staat echter garant voor een voldoende hoge signaal-ruis verhouding van de metingen. Dit aspect kan van belang zijn wanneer de responsen dicht bij de elektrofysiologische drempel liggen, een aandoening waarbij de detectie van het meevoering kan worden aangetast door een lage signaal-ruis verhouding van de meting.
Disclosures
De auteurs hebben niets te onthullen.
Acknowledgments
De auteurs erkennen Lucía Zepeda, Grace A. Whitaker en Nicolas Nieto voor hun bijdragen aan videoproductie. Dit werk werd deels ondersteund door CONICYT Programma's BASAL FB0008, MEC 80170124 en PhD beurs 21171741, evenals het Nationaal Instituut voor doofheid en andere communicatiestoornissen van de National Institutes of Health onder het awardnummer P50DC015446. De inhoud is uitsluitend de verantwoordelijkheid van de auteurs en vertegenwoordigt niet noodzakelijkerwijs de officiële standpunten van de National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
References
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G. Jr, Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. An introduction to the event-related potential technique. Second Edition. , MIT Press. 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J. Jr, Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
