

Summary
Ett protokoll för att bedöma tidsutvecklingen av den neurala övningsprovet till yttre repetitiva stimuli presenteras. Steady-state-inspelningar av samma experimentella tillstånd förvärvas och i genomsnitt i tidsdomänen. Steady-state-dynamiken analyseras genom att plotta responsamplituden som en funktion av tiden.
Abstract
Neural övningsprovet hänvisar till synkroniseringen av neurala aktivitet till periodicitet sensoriska stimuli. Denna synkronisering definierar generering av steady-state framkallade svar (dvs svängningar i elektroencefalogram fas-låst till drivande stimuli). Den klassiska tolkningen av amplituden av steady-state framkallat svar förutsätter en stereotypa tid-invariant neurala svar plus slumpmässiga bakgrund fluktuationer, så att medelvärdes över upprepade presentationer av stimulansen återvinner stereotypa respons. Detta tillvägagångssätt ignorerar dynamiken i steady-state, som i fallet med den anpassning som framkallas av långvarig exponering för stimulans. För att analysera dynamiken i steady-state-svar, kan man anta att tidsutvecklingen för responsamplituden är densamma i olika stimuleringskörningar åtskilda av tillräckligt långa raster. Baserat på detta antagande, en metod för att karakterisera tidsutvecklingen av steady-state svar presenteras. Ett tillräckligt stort antal inspelningar förvärvas som svar på samma experimentella tillstånd. Experimentella körningar (inspelningar) är i genomsnitt kolumn mässigt (dvs. löpningar är i genomsnitt, men epoken inom inspelningarna är inte medelvärdet för de föregående segmenten). Kolonnen-Wise genomsnitt möjliggör analys av steady-state svar i inspelningar med anmärkningsvärt hög signal-brus förhållanden. Därför ger den genomsnittliga signalen en korrekt representation av tidsutvecklingen av svaret vid steady-state, som kan analyseras i både tid och frekvens domäner. I denna studie, en detaljerad beskrivning av metoden ges, med hjälp av steady-state visuellt framkallade potentialer som ett exempel på ett svar. Fördelar och förbehåll utvärderas baserat på en jämförelse med en-rättegång metoder för att analysera neural övningsprovet.
Introduction
När den registreras från hårbotten, hjärnan elektrisk aktivitet observeras som kontinuerliga och regelbundna förändringar i spänningar över tiden. Denna elektriska aktivitet kallas elektroencefalogram (EEG) och beskrevs första gången av hans Berger i slutet av tjugotalet av förra århundradet1. Efterföljande nyskapande studier beskrev EEG som en sammansatt tidsserie, där olika rytmiska eller repetitiva mönster kan observeras2,3,4. Numera är EEG typiskt uppdelad i fem väletablerade frekvensband, delta, theta, alfa, beta, och gamma, som är förknippade med de olika sensoriska och kognitiva processen.
I åratal har studiet av hjärn svängningar med EEG begränsats till antingen analys av spektrumet i den pågående aktiviteten eller förändringar i oscillerande aktivitet som framkallas av icke-periodiska sensoriska händelser. Under de senaste decennierna har olika metoder införts för att modulera pågående EEG-svängningar och undersöka effekterna av sådana modulationer på perceptuella och kognitiva processer, inklusive presentationen av rytmisk sensorisk stimulering för inducera neurala övningsprovet. Termen neurala övningsprovet hänvisar till synkroniseringen av neurala aktivitet med de periodiska egenskaperna hos sensoriska stimuli. Denna process leder till generering av steady-state framkallade potentialer (dvs., EEG-svängningar låsta till de periodiska egenskaperna hos de drivande stimuli). Steady-state framkallade potentialer är oftast framkallas av visuell, auditiv, och vibrotaktil stimulering, med antingen övergående stimuli presenteras vid en konstant hastighet eller kontinuerlig stimulering modulerat i amplitud på frekvensen av intresse. Somatosensorisk steady-state framkallade potentialer (ssseps) registreras som svar på upprepad taktil stimulering5,6, steady-state visuellt framkallade potentialer (ssveps) i allmänhet framkallas av den periodiska presentation av luminans flirar, bilder och ansikten7,8. Auditiva steady-state svar (assrs) genereras vanligtvis av tåg av övergående akustiska stimuli eller genom kontinuerlig presentation av amplitud-modulerade toner9,10.
Utvinning av steady-state framkallade potentialer från den uppmätta EEG i huvudsak förlitar sig på medelvärdes senare förvärvade EEG epoker tid-låst till stimulus11. På grund av den periodicitet av svaren, kan de analyseras i både tid och frekvens domäner. Efter frekvens-domänomvandling, den sensoriska reaktionen observeras som toppar av amplitud vid presentationen hastighet eller modulering frekvens av yttre stimuli, och deras motsvarande övertoner. Dessa förfaranden (tids domänens medelvärdes förändring och den påföljande frekvensen-domänomvandling) har varit avgörande för att utveckla ett hörseltest baserat på detektion av ASSR-metoder med kliniska syften12,13,14 ,15,16.
Dessutom har den klassiska tidsdomänen medelvärdes av EEG-epoker varit mycket användbart för att analysera fysiologiska processer såsom generering och utrotning av ssvep17,18. Genom att presentera på varandra följande tåg med flimmerljus och medelvärdes efterföljande epoker inom en inspelning, konstaterade Wacker et al.19 att fas låsnings indexet för ssvep snabbt ökade under de första 400 MS av stimulering och förblev högt efteråt . De rapporterade också att robust visuell övningsprovet etablerades mellan 700-1 100 MS efter stimulus debut. En bestämd grad av övningsprovet återstod effektiv efter förskjutningen av stimuleringdrevet, som varade ungefärligt tre perioder av det oscillatoriska svaret17,19. Dessa beteenden har tolkats som den engagerande/Disengaging effekten av de observerade svängningarna, vilket är en följd av den ickelinjära informationsbehandlingen i det mänskliga visuella systemet17. Alternativt, det är känt att under vissa experimentella förhållanden, flimmerstimulering kan framkalla on-svar i början, och off-svar i slutet av stimulering tåg i stället för neurala övningsprovet18.
Det huvudsakliga antagandet att genomsnittliga i följd förvärvade EEG-epoker är att EEG-signalen representerar en linjär kombination av sensorisk respons och bakgrundsbrus20. Dessutom antas amplituden, frekvensen och fasen av oscillatoriska responsen vara stillastående, medan bakgrundsbullret betraktas som en slumpmässig aktivitet. Men i fall där detta antagande inte uppfylls, responsamplitud beräknas efter flera epoker inte nödvändigtvis motsvarar den momentana amplitud av framkallat potential.
Det har nyligen rapporterats att ASSR genereras i hjärnstammen av råttor anpassar sig till den kontinuerliga presentationen av amplitud-modulerade toner (dvs., responsamplituden minskar exponentiellt över tiden)21,22. Anpassning har tolkats som en neural mekanism som återspeglar förlusten av nyheten om en monotont repetitiva sensorisk stimulans, öka känsligheten för relevanta svängningar i den akustiska miljön23,24. I hörselgången kan anpassningen förbättra talförståelsen i bullriga miljöer. Dessutom kan denna process vara en del av befintliga mekanismer för att övervaka den auditiva återkopplingen av sin egen röst för att kontrollera talproduktionen.
Analysera tidsutvecklingen av 40 Hz ASSR hos människor, van Eeckhoutte et al.25 observerade en betydande men liten minskning av responsamplituden över tiden (runt-0,0002 μV/s baserat på grupp analys, när man antar en linjär minskning över tiden). Följaktligen, dessa författare drog slutsatsen att 40 Hz ASSR hos människor inte anpassar sig till stimulering. Hos människor, icke-stationärt beteenden har observerats vid analys av stabiliteten i SSVEP26. Dessa författare konstaterade att amplituden av den grundläggande frekvensen och den andra harmonisk av SSVEP var stillastående i endast 30% och 66,7% av de ämnen som de testade, respektive. Faserna av båda SSVEP frekvens komponenterna, även om relativt stabila över tid, uppvisade små drivor26.
Därför, även om den klassiska tiden-domän genomsnitt av senare förvärvade epoker möjliggör utforskning av stationära egenskaper av neurala övningsprovet, denna metod måste revideras när långsiktig dynamik i övningsprovet är i fokus för forskning, eller när medelvärdet av kortsiktig dynamik är skadad av förekomsten av långsiktig dynamik. För att karakterisera icke-stationära beteenden av steady-state-responsen bör det framkallade svaret som beräknats vid ett givet tidsintervall inte äventyras av de som beräknats i föregående EEG-segment. Med andra ord bör den framkallade potentialen extraheras från bakgrundsbullret utan att epokerna är tidsdomänerna i genomsnitt med de föregående EEG-segmenten.
I denna studie, en metod för att bedöma dynamiken i neurala övningsprovet presenteras. Steady-state-respons registreras upprepade gånger som svar på samma stimulering, där konsekutiva inspelningar är Interleaved med ett vilointervall på tre ggr längden på försöks körningen. Med tanke på att om tidsutvecklingen av den fysiologiska responsen är densamma i olika oberoende experimentella körningar (oberoende inspelningar), är inspelningar kolumnvis genomsnitt. Med andra ord är epoker motsvarande samma plats i de olika inspelningarna i genomsnitt, utan medelvärdes epoker inom en inspelning. Följaktligen kommer responsamplituden beräknad vid något stimuleringsintervall att motsvara den momentana amplituden för den framkallade potentialen. De sensoriska Svaren kan antingen analyseras i tids-domän eller omvandlas till frekvens-domän, beroende på syftet med experimentet. I vilket fall som helst kan amplituderna ritas som en funktion av tid för att analysera tidsutvecklingen av steady-state-svaret. Generering och utrotning av steady-state framkallade potentialer kan bedömas genom att begränsa analysen till den första och sista epokerna av inspelningarna.
Dynamiken i den neurala övningsprovet kan analyseras med hjälp av andra metoder, såsom smalbandsfiltrering enda rättegång mätningar runt frekvensen av intresse och beräkning av kuvertet av ström signalen med låg pass filtrering25 och Hilbert omformning27. Jämfört med dessa metoder tillåter kolonnwise-medelvärdet av epokerna beräkning av steady-state-parametrar baserade på signaler med det högre signal-brus-förhållandet (SNR). Nyligen har Kalman filtrering vuxit fram som en lovande teknik för uppskattning av 40-Hz ASSR amplituder28,29,30. Implementering av Kalman filtrering kan förbättra detektion av steady-state svar närmare den elektrofysiologiska tröskeln och minska tiden för förhandlingen test29. Dessutom behövs inte stationära svar för att antas när en Kalman filtrering metod används för att uppskatta ASSR amplitud30. Ändå har endast en studie analyserat tidsutvecklingen av ASSRs med Kalman filtrering25. Slutsatsen av studien är att 40-Hz-ASSR-amplituden är stabil över stimuleringsintervallet. Därför måste Kalman filtrering testas under förhållanden under vilka ASRR inte är stillastående.
Även tidskrävande är kolumnvis medelvärdes metoden modell fri och behöver inte initierings värden och/eller a priori-definitioner av brus beteendet. Dessutom, eftersom det inte innebär konvergens tider, kolonnen-Wise medelvärdes kan ge en mer tillförlitlig representation av uppkomsten av neurala övningsprovet. Därför, de resultat som erhålls med kolonnen-Wise medelvärdes metoden kan betraktas som marken sanningen för att analysera dynamiken i neurala övningsprovet med Kalman filtrering.
Den här beskrivningen av protokollet baseras på ett exempel på SSVEP. Det är dock viktigt att notera att den metod som presenteras här är modalitet-oberoende, så att den också kan användas för att analysera tidsutvecklingen av SSSEP och ASSR.
Protocol
Den aktuella studien utfördes under godkännande av forsknings-och etikkommittén vid Universidad de Valparaíso, Chile (bedömnings kod CEC170-18), bekräftad till de nationella riktlinjerna för forskning med försökspersoner.
1. förberedelser
- Välkomna ämnet.
- Förklara studiens syfte och relevans. Ge en beskrivning av relevanta tekniska detaljer. Besvara alla frågor grundligt.
- Uttryckligen nämna att hon/han kan avbryta den experimentella sessionen när som helst om så önskas.
- Be volontären att läsa ämnet informerat samtycke och underteckna motsvarande blankett. Avbryta den experimentella sessionen om informerat samtycke inte erhålls.
2. förberedelse av ämne
- Be motivet att sitta i en laboratorie stol i en bekväm position.
- Rengör hårbotten med etanol (en lösning på 95%) för att ta bort lagret av döda hudceller och talg som täcker den. Detta steg är viktigt för att reducera impedansen mellan elektroderna och hårbotten.
- Mät huvudets omkrets med ett måttband för att definiera storleken på elektrod locket som ska användas.
- Be motivet att bära elektrod locket. Ge instruktionerna för bekväm men korrekt positionering av locket.
- Mät avståndet mellan nasion (NZ, den mellersta punkten av nasofrontal suturen, som kan identifieras genom depression mellan ögonen och toppen av näsan) och Inion (iz, framträdande av occipital benet) med hjälp av en måttband.
- Mät avståndet mellan vänster och höger pre-auricular punkter (identifierad som depression strax före öron örat) med hjälp av ett måttband.
- Korrigera positionen för elektrod locket, så skärningspunkten mellan de imaginära linjer som definierats i föregående steg motsvarar huvud hörnet av huvudet. Se till att motivet är bekvämt efter justeringarna.
- Sätt ledande gel i elektrodhållare, enligt de platser som beaktas för experimentet.
- Använd 64 Scalp platser efter International 10-20 system31 att använda resultatet av protokollet för att utföra käll lokalisering analys. Det högre antalet elektrod platser (128) på hårbotten kan användas vid behov.
- Implementera kliniska eller ambulatoriska inställningar (med endast ett fåtal elektroder) om käll lokaliseringsanalys inte planeras. Använd occipital platser för att registrera SSVEP, temporala platser att förvärva ASSR och parietala platser att registrera SSSEP.
- Tryck elektroderna i elektrod hållarna. Kontrollera att elektrodens etikett överensstämmer med etikett platsen i locket.
- Åtfölja volontären till experiment rummet (företrädesvis en avskärmad, ljudförsvagad kammare). Be motivet att sitta i en stol inne i rummet, i en bekväm position.
- Placera externa elektroder på näsan och earloberna om en fysisk referens (skiljer sig från hårbotten elektroder) kommer att användas för att åter referera till EEG-inspelning (i steg 3.8.1).
- Placera externa elektroder i periokulära platser.
- Placera elektroder på kinden och den främre regionen av huvudet, cirka 1 cm ovanför ögonbrynet, för att spela in blinkande (i steg 2.6.1).
- Placera elektroder på den yttre Cantus av synar, ungefärligt 1 cm ovanför/nedanfört mittlinjen, för att anteckna synar förehavanden (i steg 2.6.1).
Obs: den elektrooculogram (EOG) kommer att användas i steg 3.8.5 för att ta bort ägg artefakter induceras av blinkande och ögonrörelser.
- Vrid på EEG-anskaffningssystemet och kontrollera elektroimpedansen om ett system med låg impedans används för att registrera EEG. Korrigera impedansen, efter behov, enligt tillverkarens anvisningar. Impedansen ska hållas under 10 kΩ32.
- Be motivet blinka och flytta ögonen i olika riktningar för att säkerställa att EOG registreras korrekt.
- För att analysera dynamiken i SSVEP, justera placeringen av skärmen i vertikal riktning, för att matcha vyvinkeln för motivet. Dämpa belysningen i rummet tills en bekväm nivå uppnås. Justera skärmens luminansnivå till den övre gränsen för deltagarens komfort nivå.
- För att analysera dynamiken i ASSR, sätt i hörlurarna med rätt skum skär, så att hörlurarna passar hörselgången. Kontrollera att ljud levereras med önskad intensitet (t. ex. en psykofysisk bekväm nivå33).
3. EEG-förvärv och förbehandling
- Ställ in de stimulans parametrar som definierats i experiment designen. Se den bruksanvisning som tillhandahålls av tillverkaren av stimuleringssystemet för mer information om programvaran.
Anmärkning: för omfattande förklaringar av den stimulans som används för generering av SSVEP och ASSR, se Norcia et al.8 och rance34, respektive. - Instruera ämnet att uppmärksamma stimulering, i fall att visuell övningsprovet är ämnet för experimentet.
- Presentera en textade film med ljudet av när auditiv övningsprovet är ämnet för experimentet.
Obs: presentation av en tyst film tillåter avböjning av uppmärksamheten från den akustiska stimulering samtidigt som upphetsning nivå25.
- Presentera en textade film med ljudet av när auditiv övningsprovet är ämnet för experimentet.
- Nuvarande stimuli längre än 90 s, som har gjorts för att undersöka tidsutvecklingen av ssveps och ASSR i både människor och djurmodeller21,22,25,26.
Anmärkning: nuvarande stimuli kortare varaktighet om en pilotstudie har utförts. - Pausa stimulering i 2 min om bara ett försöks tillstånd testas. Interagera med motivet för att kontrollera medvetenheten.
Obs: varaktigheten av pausen beror på varaktigheten av stimulering. Pauser 3x längre än stimuleringsintervallen kommer att säkerställa att ett svar som framkallas av en stimulans inte påverkas av den tidigare stimulansen. Längre pauser är tillåtna om motivet så begär.- Pausa stimulering i minst 10 sekunder när olika experimentella förhållanden testas sedan alternerande stimulering med pauser på 10 s har föreslagits för att minska extra anpassnings effekter och minska längden på experimentet25.
- Upprepa presentationen steg (steg 3.3-3.4) minst 30x för att säkerställa hög SNR av mätningarna efter medelvärdet av epokerna (steg 4,4).
- Registrera EEG med standardprocedurer35. Skapa en separat EEG-fil för varje experimentell körning.
Obs: Se användarhandboken för förvärvs systemet för mer information om programvaran. - Övervaka EEG-inspelning för att upptäcka sömn perioder baserat på nivån av alfaaktivitet och frekvensen där blinkande artefakter visas. Pausa experimentet när förhöjda alfa nivåer åtföljs av minskade blinkande frekvenser upptäcks, vilket är ett tecken på sömnighet. Avvisa experiment körningen från ytterligare analys när sömn perioder upptäcks.
- Beräkna amplituden för steady-state-svaret i slutet av varje försöks körning, enligt instruktionen i användarhandboken för anskaffningsprogram varan som används i experimentet.
- Övervaka den uppmärksamhet nivån av ämnet genom att jämföra amplituden av steady-state svar som erhållits i slutet av varje experimentell körning. Ställ in den steady-state amplitud som erhålls i de första experimentella körningar som en referenamplitud.
- Ange ett tröskelvärde för avvisande (en minskning av responsamplituden på 5% avseende referensamplitud). Avvisa experimentella körningar där amplituden för steady-state-responsen uppfyller avvisnings kriteriet.
- Avsluta den experimentella sessionen efter att ha förvärvat antalet körningar som definierats i experimentell design.
- Pre-process EEG data offline med hjälp av standard EEG förfaranden35 beskrivs i nästa steg enligt tillverkarens anvisningar.
- Re-Reference inspelningen med hjälp av en genomsnittlig referens (genomsnittet av alla inspelnings elektroder) eller genomsnittet av en delmängd av elektroder. Alternativt kan du använda en fysisk referens (t. ex. externa elektroder placerade på näsan och earloberna som beskrivs i steg 2,4).
- Omvandla elektrod koordinaterna till det internationella 10/20-systemet om det radiella koordinatsystemet användes under EEG-anskaffningarna. Mer information om konvertering finns i tillverkarens handbok.
- Band-pass filtrerar EEG-signalen mellan 0,5-300 Hz. Ställ in ett notch-filter (centrerat vid 50 Hz eller 60 Hz) vid behov.
- Ner-prov EEG-signalen för att minska körningstiden för den algoritm som valts för att ta bort okulära artefakter (steg 3.8.5).
Anmärkning: en provtagningsfrekvens på 512 Hz är tillräcklig för att analysera hjärnans svängningar av frekvensen under 40 Hz35. - Ta bort okulära artefakter.
Anmärkning: för detta ändamål, olika tekniker kan användas (se Urigüen och Garcia-Zapirain35 för en omfattande översyn på artefakt avlägsnande algoritmer). Bland dem är oberoende komponent analys en av de mest utökade metoder och genomförs i både kommersiella och fria analys mjukvaror37,38,39. - Segmentera EEG-data i epoker som är tidslåsta till stimulering. Välj epoker längd enligt experimentets syfte.
Anm.: epoker bör vara tillräckligt långa för att möjliggöra analys av steady-state-responsen i frekvensområdet med en adekvat spektralupplösning. - Kör inte artefakt avvisnings algoritmer i det här skedet för att identifiera och ta bort epoker som innehåller artefakter.
Anmärkning: ta bort epok i det här skedet kommer att orsaka fel när datauppsättningen är organiserad för att köra kolumnvis medelvärde av epoker (steg 4,2 och 4,4). Avvisnings algoritmer implementeras vid ett senare bearbetningssteg (steg 4.1.4). - Kör funktionen DC-detrend för att beräkna DC-trender i enskilda EEG-epoker och korrigera dem.
- Kör funktionen baslinjekorrigering för att korrigera baslinjen för inspelningen. Välj pre-stimulus tidsintervall längre än 200 MS.
Anmärkning: baseline korrigering består av medelvärdes data i det valda tidsintervallet. Medelvärdet beräknas för varje kanal och subtrastreras från varje datapunkt i varje epok.
4. beräkning av responsamplituderna
- Ange de parametrar som behövs för beräkningen av svaren från steady-state (figur 1a).
Obs: den egen kod som används för att behandla data är fritt tillgänglig på < https://figshare.com/projects/steady-state_visually_evoked_potentials_SSVEP_elicited_in_humans_by_continuos_light_modulated_in_amplitude_at_10Hz/62573 >. Mer information finns i hjälptexten i koden. På samma sätt är en delmängd av de data som används i denna studie tillgänglig.- Ange antalet inspelningar (experimentella körningar) av experimentet.
- Ange längden på epokerna för att segmentera de enskilda inspelningarna.
- Ange samplingsfrekvensen för experimentet.
- Välj artefakt avvisnings algoritmer för att identifiera och ta bort epoker som innehåller artefakter. De tillgängliga urvalskriterierna är 1) gradient (absolut skillnad mellan två på varandra följande prover), 2) max-min (skillnaden mellan den maximala och minsta amplituden i epoken), och 3) amplitud (absolut högsta och lägsta amplituder).
- Kör bearbetnings koden.
Anmärkning: steg 4.2-4.7 utförs automatiskt när detta alternativ är valt. Kör stegen manuellt om det behövs.
- Arrangera om epokerna in i en data matris av n ror och m kolonner, som n föreställer i numrera av inspelningar (experimentella körningar) och m numrera av epoker (figurera 1b).
- Vikt epoken för att dämpa effekten av rörelse och muskulös artefakter.
Anmärkning: viktade EEG-epoker erhålls genom att varje spännings prov divideras med amplitudvariationen för den epok de tillhör, så att varians används som ett mått på amplitudvariabilitet och viktningsfaktor40. - Kolumn mässigt genomsnitt datauppsättningen. För detta ändamål, tid-domän genomsnitt epokerna motsvarande samma tid fönstret i de olika inspelningarna.
Anmärkning: detta steg möjliggör beräkning av steady-state amplitud i inspelningar med en anmärkningsvärt hög signal-brus-förhållande (SNR). - Exportera tidsserierna till följd av medelvärdet för ytterligare analys av tidsutvecklingen av övningsprovet i extern programvara.
- Beräkna amplituden för steady-state-respons i varje epok som resulterar från kolonnwise-medelvärdet, med hjälp av fast Fourier Transform (FFT).
Anmärkning: FFT-längden bör motsvara längden på en epok. Implementering av en fönster teknik är inte obligatoriskt. Amplituden av steady-state svaret definieras som spektralamplitud erhålls vid frekvensen av amplitud modulering av sensoriska stimuli.
- Beräkna amplituden för steady-state-respons i varje epok som resulterar från kolonnwise-medelvärdet, med hjälp av fast Fourier Transform (FFT).
- Vektor genomsnitt amplituden för ett ad-hoc-antal FFT papperskorgar på vardera sidan av frekvensen av svaret för att beräkna den återstående bullernivån (RNL). Antalet FFT-bingar bör motsvara ett frekvensband på ungefär 3 Hz, på vardera sidan om responsfrekvensen.
Anmärkning: den höga frekvensen-specificitet av steady-state Svaren gör responsamplitud oberoende av de bakgrunds svängningar med liknande frekvenser, som i sin tur distribuerar enhetligt i ett relativt smalt frekvensband41 , 42 , 43. - Rita upp amplituden för steady-state-responsen och RNL som en funktion av kolumnindex (dvs. antalet förvärvade epoken) för att undersöka utvecklingen av steady-state-responsen under stimuleringsintervallet.
Representative Results
SSVEP eliciserades av kontinuerliga visuella stimuli av 40 s i längd, där ljusintensiteten modulerades av en sinusformad våg av 10 Hz (modulationsdjup på 90%). Stimuli levererades av fyra lysdioder (leds) ligger i mitten av en 50 cm x 50 cm svart skärm, som hörn av en 5 cm x 5 cm kvadrat. När deltagaren satt 70 cm från skärmen, det område av kvadraten på lysdioder subansar en visuell vinkel på ca 4 °. LED-skärmen utformades med hjälp av en USB-baserad mikrokontroller utvecklingssystem och fyra super ljusa vita lysdioder av 10 mm i diameter. Tekniken för puls breddmodulering (PWM) användes för att styra den ström som levererades till lysdioderna. Denna teknik kontrollerade lysdioder intensiteter vid en given frekvens och generera det slutliga sinusformade kuvertet. En PWM-frekvens på 40 kHz användes för att undvika en märkbar flimmereffekt.
Trettio inspelningar erhölls, som segmenterades i epoker av 4 s. Därför erhölls en datauppsättning bestående av 10 kolumner (antal EEG-epoker inom inspelningar) och 30 rader (antal inspelningar, antal experimentella körningar).
Den neurala svängning tid-låst till stimulering blev uppenbar som kolonnen-Wise medelvärdes utfördes (figur 2). I hög grad kan det intervall vid vilket SSVEP genereras observeras i spår som motsvarar kolumn 1. I denna kolumn, 0,2 s av pre-stimulus baseline ritas utöver de första 0,8 s av neurala övningsprovet. Därför, det förfarande som beskrivs här tillåter karakterisering av 1) dynamiken i oscillatoriska svar gång neural övningsprovet är redan etablerad och 2) engagemang av neurala svängningar. En eller flera epoker som spelats in efter stimuleringen kan också inkluderas i data matrisen för att studera utrotning av steady-state-responsen efter stimulus offset.
Under kolonnwise-medelvärdet av epokerna minskade medelamplituden för SSVEP (spektralamplitud vid 10 Hz, beräknad genom tillämpning av FFT) under medelvärdet av de första epokerna i kolonnerna och tenderade att stabiliseras efteråt (figur 3a). Detta resultat överensstämmer med tidigare studier analysera utvecklingen av ASSR under medelvärdet av sekventiellt förvärvade epoker21,22,40,43,44. Beteendet hos responsamplituden under medelvärdet förklaras vanligtvis av det relativt höga bidraget från ogenomsnittligt brus till responsamplituden som beräknats i de första epokerna, som dämpas som medelvärdesberäkning utförs13, 44 , 45 , 46 , 47. anmärkningsvärt, minskade ssvep amplitudvariationen signifikant som genomsnitt fortskred.
Vi analyserade också RNL av mätningarna under kolonnen-Wise medelvärdes av epokerna (figur 3b). RNL beräknades i ett smalt frekvensband (3 Hz) på båda sidor om frekvensen av SSVEP. Även om detta förfarande är inte vanligt när ssvep analyseras, vektor-genomsnitt ett visst antal frekvens behållare runt den i neurala övningsprovet är standarden för att uppskatta RNL i ASSR mätningar41,42, 43som väntat minskade RNL successivt när antalet genomsnittliga epoker ökade och nådde den asymptotiska nivån efter cirka 20 epoker behandlades. Till skillnad från vad som observerades när SSVEP-amplituden analyserades förblev standardavvikelsen för RNL relativt konstant då antalet genomsnittliga epokerna ökade, vilket tyder på att inspelningsförhållandena var stabila längs försöks sessionen.
De resultat som presenteras ovan fastställt förändringarna i Peak signal-brus-förhållande (pSNR) av mätningar under kolonnen-Wise medelvärdes epoken (figur 3C). Denna term definieras här som förhållandet (i dB) mellan kvadratamplituden av svaret (SSVEP) och kvadratamplituden av RNL. Som genomsnitt fortskred, ökade pSNR som antalet genomsnittliga epokerna ökade upp till 18, ungefär. Ytterligare ökningar i antalet genomsnittliga epoker påverkade inte signifikant kvaliteten på signalen. Variationen i pSNR minskade då fler epoker var i genomsnitt.
Slutligen är dynamiken i SSVEP-amplituden och RNL representerade i figur 4. Dessa tids utvecklingar erhölls genom plottning av svars parametrarna som beräknats i slutet av kolonnwise-medelvärdet av epokerna som en funktion av antalet kolonner (som en funktion av tiden). Som framgår av Labecki et al.26, dynamiken i SSVEP kan avsevärt variera mellan ämnen. Eftersom de resultat som presenteras i figur 4 motsvarar en enskild individ, kan generaliseringar inte göras. I detta ämne, amplituden av SSVEP visade ett relativt komplext beteende (figur 4a). Responsamplituden ökade gradvis under de första 12 sekunderna efter den stimulans debut (tid som motsvarar längden på 3 epoker). När stimulansen framhärdade minskade SSVEP genomgående under de följande 12 sekunderna och förblev relativt konstant efteråt. Dessa resultat kan inte förklaras av beteendet hos RNL, eftersom denna parameter var relativt konstant under stimuleringsintervallet (figur 4b). Ökningen av SSVEP amplituden efter stimulus debut är uppenbar i spåren presenteras i figur 2 och kan förklaras av integrationsprocesser, vilket resulterar i stabilisering av neurala övningsprovet. Den efterföljande minskningen av amplitud tyder på anpassning av SSVEP till ihållande stimulering. Dessa hypoteser måste dock testas i kontrollerade experiment med tillägade provstorlekar.
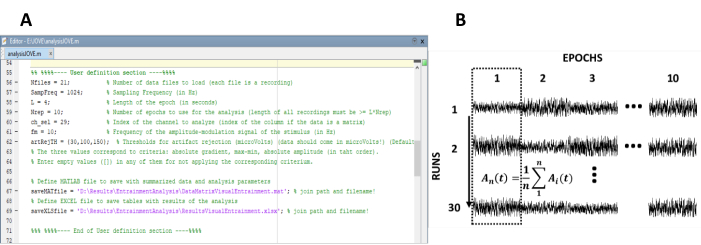
Figur 1 : Kritiska steg för att extrahera tid utvecklingen av amplituden av steady-state svar. (A) skärmbild av bearbetnings koden, där analys parametrarna definieras. (B) representativt diagram som illustrerar datamängdens organisation. En data matris som komponeras av 30 inspelningar av 10 epoker, föreställs. I den första kolumnen framhävs kolumn mässigt medelvärdet av epokerna. Den vertikala linjen representerar riktningen av medelvärdes gränsen. Vänligen klicka här för att se en större version av denna siffra.

Figur 2 : Förändringar i vågform av steady-state visuellt framkallade potentialer (SSVEP) under kolonnen-Wise medelvärdes av epoker. Svaren uppnåddes genom kontinuerlig presentation av ljus modulerad i amplitud vid 10 Hz. Raderna visar vågformerna som erhålls efter medelvärdes alla tidigare inspelningar (dvs. rad 1 är den första inspelningen, rad 5 är vågformen som erhålls efter att ha i genomsnitt de första fem inspelningarna, och den sista raden är genomsnittet av alla inspelningar). Mer pålitliga vågformer av SSVEP observerades i varje kolumn som antalet medelvärdes körningar ökade. För att ge klarhet (för att göra svängningarna av SSVEPEN synligt), endast föreställs den första understödja av epokerna. Undantagen är spår i den första kolumnen i datauppsättningen, som 0,2 sekunder av före stimulus baseline visas. Vänligen klicka här för att se en större version av denna siffra.

Figur 3 : Förändringar i parametrarna för respons och registrering under kolumn mässigt medelvärde av epoker. A) utvecklingen av SSVEP-amplituden. (B) RNL-beteendet. C) ändringar i psnr. Svarta linjer representerar medelvärden som erhålls för varje kolumn (n = 10) och den gråa skuggan representerar det område som täcks av ± en standardavvikelse. Vänligen klicka här för att se en större version av denna siffra.

Figur 4 : Tidsutvecklingen av SSVEP framkallade genom presentationen av kontinuerlig Visuell stimulering, modulerad i amplitud vid 10 Hz. A) tidsförlopp för SSVEP-amplituden. (B) tidsförlopp för RNL. Vänligen klicka här för att se en större version av denna siffra.
Discussion
Detta arbete beskriver ett experiment förfarande för att analysera dynamiken i oscillatoriska hjärnan svar. En sådan metod består i att förvärva ett tillräckligt antal oberoende experimentella körningar av samma experimentella tillstånd, och tids domänerna medelvärdes epoker motsvarande samma tidsfönster i de olika inspelningarna (kolonner-Wise medelvärdes Figur 1B). Den amplitud som beräknats i de genomsnittliga data representerar den momentana amplituden för oscillerande respons. Plottning dessa amplituder som en funktion av tid (eller antalet kolumner i datauppsättningen) gör det möjligt att analysera tidsutvecklingen av oscillerande responstid-låst till stimulering. Denna metod är en modifiering av den som föreslagits av Ritter et al.23 för att analysera anpassningen av övergående kortikala framkallade potentialer. Metoden har använts för att analysera dynamiken i auditiva framkallade potentialer i både människor24 och djurmodeller20,21.
Ur en metodologisk synvinkel är kombinationen av parametrar som används för att framkalla steady-state-respons och de som genomförs för att extrahera neurala svar från bakgrundsbrus är avgörande för att analysera tidsutvecklingen av steady-state framkallade potentialer 22. den stimulans längd som används i experimentet som presenteras här (40 s) valdes baserat på resultat som erhållits i en pilotstudie. Denna stimulans längd var tillräcklig för att analysera anpassningen av ASSR genereras i råtta hjärnstammen21,22. Dessutom bör stimulans längden överstiga den tid då den asymptotiska momentana band kraften i SSVEPs uppnås (figur 1 i Labecki et al.26). Icke desto mindre, den asymptotiska momentant band makt SSVEPs kan nås bortom 60-talet i vissa fall (figur 2 i Labecki et al.26). Därför, kör en liten prov pilotstudie rekommenderas att definiera stimulans längden av stimulering. Annars, en stimulans längd längre än 90 s rekommenderas att uppnå fullständig representation av tiden utvecklingen av svaret. Att använda tillräckligt långa pauser mellan på varandra följande inspelningar förutsätter att försöks körningar i rad är statistiskt oberoende (dvs. olika, oberoende åtgärder av samma variabel). Till det bästa av vår kunskap, inga experiment har utförts för att analysera den optimala paus mellan körningar (minsta paus krävs för att göra körs oberoende av varandra). Kriteriet för att använda pauser minst 3x längre än stimulus längd är konservativ nog för att säkerställa att steady-state svar registreras i en given körning inte påverkas av den tidigare stimulering.
Nyligen har omväxlande stimuli (experimentella förhållanden) föreslagits som ett val för att minska pausen mellan experimentella körningar, undvika extra anpassnings effekt25. Likaså, antalet experimentella körningar (30) genomförs i detta experimentella protokollet är konservativ, eftersom asymptotiska RNL och pSNR är vanligtvis nås efter genomsnitt 20 experimentella körningar, cirka. När stimuli faller inom den mellersta övre regionen av det dynamiska omfånget av svaret (hög sensation nivåer), lägre antal körningar är sannolikt behövs för att analysera dynamiken i det framkallade svaret. I fall där olika försöksbetingelser testas är det dock avgörande att ha samma antal experimentella körningar för att göra jämförelser mellan olika förhållanden (dvs. andra förnimmelser).
Förutom kolonnen-Wise medelvärdes av epoker, dynamiken i oscillerande framkallade potentialer har analyserats genom filtrering av endosmätningar i ett smalt frekvensband runt frekvensen av intresse och beräkning av kuvertet av makten signal med låg pass filtrering26. Likaså har en enda försöks analys genomförts för att karakterisera övergångsperioden som föregår den stabila regionen SSVEP48, och förändringarna i amplitud och fas av ssvep under den stabila regionen av responsen49. Medan en enda försöks analys möjliggör diskriminering av relativt snabba svängningar i responsamplitud, kan experimentella konstruktioner för att analysera den genomsnittliga responsen i block separerade med ett givet Inter-block intervall endast redogöra för långsiktiga variationer i amplitud av den framkallat potential50,51. Den kolonn-Wise medelvärdes av epoker står mellan dessa två alternativ. Konvertera den genomsnittliga signalen till frekvens-domän med hjälp av FFT innebär att analysera dynamiken i svaret med en upplösning som är lika med längden på epoken. I det exempel som presenteras här, rapporterades SSVEP var 4: or. Även om 4 s upplösning är tillräcklig för att beskriva dynamik som inträffar vid tidsintervall som överträffar tiotals sekunder, såsom den i SSVEP26, delvis överlappande epoker i originalinspelningar gör det möjligt att beskriva tidsutvecklingen av steady-state-respons på ett mer förfinat sätt25.
Dynamiken i de steady-state-svar som erhålls efter kolumn mässigt medelvärde av epoker representerar främst utvecklingen av den oscillerande aktiviteten som synkroniseras mellan de genomsnittliga EEG-segmenten (de som överlever medelvärdes medelvärdet). Därför är en stor fråga om genomförbarheten av metoden en möjlig dämpning av responsamplituder på grund av variationer i fasen av neurala svängningar från en oberoende experimentell körning till en annan (dvs. bland inspelningar). Det här avsnittet måste åtgärdas experimentellt. Emellertid, bevis tyder på att fasen av hjärnan oscillerande svar är mindre varierande än förväntat. I själva verket, flera studier har rapporterat en regelbundenhet i den förväntade fasen av den mänskliga 80 Hz ASSR47,48,49. När latenser uppskattas baserat på fasen av oscillerande aktiviteten, den förutsägbara effekten av intensiteten och bärvågsfrekvensen av de akustiska stimuli på latensen av hörsel Svaren har observerats (dvs., latens minskning som intensitet och bärarefrekvens ökning)52,53,54. Dessutom har typiska mognande förändringar i amplitud och vänster till höger asymmetri i hörsel nivåer också observerats när latenser beräknas från fasen av ASSR47,55,56 , 57 , 58. när man beskriver tidsutvecklingen för SSVEP med en enda försöks analys, konstaterade Labecki et al.26 att även om variabiliteten mellan proven av responsamplituderna inom samma ämne var avsevärt hög, var variationen i fasen var signifikant mindre uttalad.
Baserat på deras observationer föreslog Labecki et al.26 att ett minimum av 50 prövningar skulle beräknas för att få en tillförlitlig uppskattning av det genomsnittliga effekt anslaget för responsen. Dessa resultat indikerar att, även när amplituden av svaret beräknas i enskilda prövningar, genomsnitt (av kuvert i det fallet) behövs för att rapportera pålitliga resultat. Dessutom tyder den inter-Trial variationen i amplituden av SSVEP rapporteras av Labecki et al.26 att beräkningen av denna parameter i enskilda prövningar kan vara starkt påverkad av bakgrundsljud. Med tanke på utvecklingen av signal-brus-förhållandet som presenteras i figur 2, minskar beräkningen av responsen i den genomsnittliga signalen istället för enstaka prövningar signifikant det antal EEG-segment som behövde bearbetas för att erhålla tillförlitlig Mätningar. Dessutom stöder den låga variationen i fas erhålls av Labecki et al.26 tanken att kolonnwise medelvärdes av epoker presenteras här är ett giltigt förfarande för beräkning av dynamiken i oscillerande framkallade potentialer.
Medelvärdes data på olika nivåer leder till olika tolkningar av resultaten. När det gäller oscillatoriska framkallade potentialer, Computing responsamplituden efter tidsdomänen medelvärdes av oberoende körningar innebär att analysera endast tidslåsta svängningar (dvs. de som överlever medelvärdes). Den här proceduren kan filtrera relevant information om dynamiken i svaret i enskilda prövningar. Det garanterar dock en tillräckligt hög signal-brus-förhållande av mätningarna. Denna aspekt kan vara av betydelse när Svaren ligger nära den elektrofysiologiska tröskeln, ett tillstånd där detektering av övningsprovet kan äventyras på grund av låg signal-brus-förhållande av mätningen.
Disclosures
Författarna har inget att avslöja.
Acknowledgments
Författarna erkänner tacksamt Lucía Zepeda, Grace A. Whitaker, och Nicolas Nieto för deras bidrag till videoproduktion. Detta arbete stöddes delvis av CONICYT program BASAL FB0008, MEC 80170124 och PhD stipendium 21171741, liksom det nationella institutet för dövhet och andra kommunikationsstörningar i National Institutes of Health under tilldelnings nummer P50DC015446. Innehållet är uteslutande författarnas ansvar och representerar inte nödvändigtvis de officiella åsikter som finns hos National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
References
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G. Jr, Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. An introduction to the event-related potential technique. Second Edition. , MIT Press. 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J. Jr, Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
