

Summary
En protokoll for å vurdere tiden utviklingen av nevrale entrainment til eksterne repeterende stimuli presenteres. En fast-State opptak av samme eksperimentelle tilstand er ervervet og gjennomsnitt i tids domenet. Den steady-state dynamikk analyseres ved å plotte responsen amplitude som en funksjon av tid.
Abstract
Neural entrainment refererer til synkronisering av nevrale aktivitet til periodisitet av sensoriske stimuli. Denne synkroniseringen definerer generering av steady-state fremkalt respons (dvs. svingninger i Elektroencefalogram fase-låst til kjøringen stimuli). Den klassiske tolkningen av amplituden til steady-state fremkalt respons forutsetter en stereotype tid-invariant neural respons pluss tilfeldig bakgrunn svingninger, slik at snitt over gjentatte presentasjoner av stimulans gjenoppretter stereotype respons. Denne tilnærmingen ignorerer dynamikken i steady-state, som i tilfelle av tilpasningen elicited ved langvarig eksponering for stimulans. For å analysere dynamikken i steady-state svar, kan det antas at tiden utviklingen av responsen amplitude er den samme i ulike stimulering går atskilt med tilstrekkelig lange pauser. Basert på denne antakelsen, en metode for å karakterisere tiden utviklingen av steady-state responser er presentert. Et tilstrekkelig stort antall innspillinger er ervervet som svar på samme eksperimentelle tilstand. Eksperimentelle runs (innspillinger) er kolonne-messig gjennomsnitt (dvs. kjører er gjennomsnitt, men epoken innenfor opptakene er ikke gjennomsnitt med foregående segmenter). Den kolonne-klok gjennomsnitt tillater analyse av steady-state svar i innspillinger med bemerkelsesverdig høy signal-til-støy prosenter. Derfor, gjennomsnittlig signal gir en nøyaktig representasjon av tiden utviklingen av steady-state respons, som kan analyseres i både tid og frekvens domener. I denne studien, en detaljert beskrivelse av metoden er gitt, ved hjelp av steady-state visuelt fremkalt potensial som et eksempel på et svar. Fordeler og advarsler er evaluert basert på en sammenligning med én prøve metoder utviklet for å analysere nevrale entrainment.
Introduction
Når den er innspilt fra hodebunnen, er hjernen elektrisk aktivitet observert som kontinuerlige og regelmessige endringer i spenninger over tid. Denne elektriske aktiviteten kalles Elektroencefalogram (EEG) og ble først beskrevet av hans Berger i slutten av tyve årene av forrige århundre1. Påfølgende banebrytende studier beskrev EEG som en sammensatt tidsserie, der ulike rytmiske eller repeterende mønstre kan observeres2,3,4. I dag er EEG vanligvis delt inn i fem veletablerte frekvensbånd, delta, theta, Alpha, beta og gamma, som er forbundet med ulike sensoriske og kognitive prosessen.
For år, studiet av hjernen svingninger bruker EEG var begrenset til enten analyse av spekteret i den pågående aktiviteten eller endringer i oscillasjon aktivitet elicited av ikke-periodiske sensoriske hendelser. I de siste ti årene har ulike metoder blitt implementert for modulerende pågående EEG-svingninger og utforsking av virkningene av slike modulasjoner på de kognitive og kognitiv prosessene, inkludert presentasjon av rytmisk sensorisk stimulering for inducing neural entrainment. Begrepet neural entrainment refererer til synkronisering av nevrale aktivitet med periodiske egenskaper sensoriske stimuli. Denne prosessen fører til generering av steady-state fremkalt potensialer (dvs. EEG svingninger låst til de periodiske egenskapene til kjøringen stimuli). Steady-state vakte potensialer er oftest elicited av visuell, hørbar og vibrotactile stimulering, ved hjelp av enten forbigående stimuli presentert på en konstant hastighet eller kontinuerlig stimulering modulert i amplitude på hyppigheten av interesse. Mens somatosensory steady-state vakte potensialer (SSSEPs) registreres som svar på repeterende taktil stimulering5,6, steady-state visuelt fremkalt respons (SSVEPs) er generelt elicited av den periodiske presentasjon av lysstyrke flimrer, bilder og ansikter7,8. Hørbar steady-state responser (ASSRs) er vanligvis generert av tog av forbigående akustiske stimuli eller ved kontinuerlig presentasjon av amplitude-modulert toner9,10.
Utvinning av steady-state fremkalt potensialer fra den målte EEG hovedsak er avhengig av snitt senere ervervet EEG epoker tid-låst til stimulans11. På grunn av periodisitet av svarene, kan de bli analysert i både tid og frekvens domener. Etter at frekvens-domene transformasjon, er sensorisk respons observert som topper amplitude på presentasjons hastighet eller modulering frekvens av eksterne stimuli, og deres tilsvarende harmoniske. Disse prosedyrene (tid-domene snitt og påfølgende frekvens-domene transformasjon) har vært avgjørende for å utvikle en hørselstest basert på påvisning av ASSR metoder med kliniske formål12,13,14 ,15,16.
Videre har den klassiske tid-domene gjennomsnitt av EEG epoker vært svært nyttig for å analysere fysiologiske prosesser som generasjon og utryddelse av SSVEP17,18. Wacker et al.19 , som presenterer fortløpende tog med flimmer og gjennomsnittlig påfølgende epoker i et opptak, observerte at den fase lås ende INDEKSEN til SSVEP raskt økte i løpet av den første 400-MS av stimulering og forble høy etterpå . De rapporterte også at robust visuell entrainment ble etablert mellom 700-1 100 MS etter stimulans utbruddet. En viss grad av entrainment forble effektiv etter forskyvningen av stimulering toget, som varte i omtrent tre perioder av oscillasjon respons17,19. De atferd har blitt tolket som engasjerende/frigjøre effekten av den observerte svingninger, som er en konsekvens av den ikke-lineære informasjonsbehandling i det menneskelige visuelle systemet17. Alternativt er det kjent at under visse eksperimentelle forhold, flimmer stimulering kan lokke fram på svar i begynnelsen, og off-respons på slutten av stimulering tog i stedet for nevrale entrainment18.
Den viktigste forutsetningen til gjennomsnittlig etter hverandre ervervet EEG epoker er at EEG signalet representerer en lineær kombinasjon av sensorisk respons og bakgrunnsstøy20. Videre er amplituden, frekvensen og fasen av oscillasjon respons antatt å være stasjonær, mens bakgrunnsstøyen betraktes som en tilfeldig aktivitet. Men i tilfeller der denne forutsetningen ikke er oppfylt, er responsen amplitude beregnet etter flere epoker ikke nødvendigvis tilsvarer den umiddelbare amplitude av fremkalt potensiale.
Det har nylig blitt rapportert at ASSR generert i hjernestammen av rotter tilpasser seg den kontinuerlige presentasjonen av amplitude-modulert toner (dvs. responsen amplitude reduseres eksponentielt over tid)21,22. Tilpasning har blitt tolket som en nerve mekanisme som reflekterer tapet av nyheten om en monotont repeterende sensorisk stimulans, øke følsomheten til relevante svingninger i det akustiske miljøet23,24. I hørsels veien kan tilpasningen forbedre taleforståelsen i støyende omgivelser. Videre kan denne prosessen være en del av eksisterende mekanismer for å overvåke hørsels tilbakemeldingen til ens egen stemme for å kontrollere tale produksjonen.
Analysere tiden utviklingen av 40 Hz ASSR i mennesker, Van Eeckhoutte et al.25 observert en betydelig, men liten nedgang i respons amplitude over tid (rundt-0,0002 μV/s basert på gruppe analyse, når forutsatt en lineær nedgang over tid). Følgelig konkluderte disse forfatterne med at 40 Hz ASSR hos mennesker ikke tilpasser seg stimulering. Hos mennesker, ikke-stasjonær atferd har blitt observert når analysere stabiliteten i SSVEP26. Disse forfatterne observert at amplituden til grunnleggende frekvens og den andre harmoniske av SSVEP var stasjonære i bare 30% og 66,7% av fagene de testet, henholdsvis. Fasene av både SSVEP frekvens komponenter, men relativt stabil over tid, utstilt små driver26.
Derfor, selv om den klassiske tid-domenet i snitt av senere ervervet epoker tillater utforsking av stasjonære egenskaper av Neural entrainment, denne metodikken må revideres når langsiktige dynamikken i entrainment er fokus for forskning, eller når gjennomsnittet av kortsiktige dynamikk er ødelagt av forekomsten av langsiktige dynamikk. For å karakterisere ikke-stasjonær atferd av steady-state svar, det fremkalt respons beregnet på et gitt tidspunkt vindu bør ikke bli kompromittert av de som er beregnet i de foregående EEG segmentene. Med andre ord, det vakte potensialet bør trekkes ut fra bakgrunnsstøyen uten epoker være tid-domene gjennomsnitt med de foregående EEG segmenter.
I denne studien, en metode for å vurdere dynamikken i nevrale entrainment presenteres. Faste-State svar er full av gjentagelser registrert som svar på samme stimulering, der påfølgende innspillinger er sammenflettet av et hvileintervall på tre ganger lengden på eksperimentell kjøre. Tatt i betraktning at hvis tiden utviklingen av fysiologiske responsen er den samme i ulike uavhengige eksperimentelle går (uavhengig innspillinger), innspillinger er kolonne-klok gjennomsnitt. Med andre ord, epoker tilsvarende til samme sted i de ulike opptakene er gjennomsnitt, uten snitt epoker innenfor et opptak. Følgelig vil respons amplituden som beregnes ved et hvilket som helst stimulering intervall tilsvare den umiddelbare amplituden til det fremkalt potensialet. Den sensoriske reaksjoner kan enten analyseres i tid-domene eller forvandlet til frekvens-domene, avhengig av målet for eksperimentet. I alle fall kan amplituder tegnes som en funksjon av tid til å analysere tid utviklingen av steady-state respons. Generasjon og utryddelse av steady-state fremkalt potensial kan vurderes ved å begrense analysen til første og siste epoker av opptakene.
Dynamikken i neural entrainment kan analyseres ved hjelp av andre tilnærminger, for eksempel Smalbånd filtrering enkelt-rettssaken målinger rundt hyppigheten av interesse og databehandling konvolutten av strømsignalet ved hjelp av low-pass filtrering25 og Hilbert transformasjon27. Sammenlignet med disse metodene, den kolonne-klok gjennomsnitt av epoker tillater databehandling steady-state parametre basert på signaler med høyere signal-til-støy-forhold (SNR). Nylig har Kalman filtrering dukket opp som en lovende teknikk for estimering av 40-Hz ASSR amplituder28,29,30. Implementering av Kalman filtrering kan forbedre påvisning av steady-state responser nærmere elektrofysiologisk terskelen og redusere tiden for hørselstest29. Videre stasjonære tiltak er ikke nødvendig å bli antatt når en Kalman filtrering tilnærming brukes til å anslå ASSR amplitude30. Likevel har bare én studie analysert tiden utviklingen av ASSRs bruker Kalman filtrering25. Konklusjonen av studien er at 40-Hz ASSR amplitude er stabil over stimulering intervallet. Kalman filtrering må derfor testes i forhold der ASRR ikke er i ro.
Selv om tidkrevende, kolonnen-klok gjennomsnitt metoden er modell-fri og trenger ikke initialisering verdier og/eller a priori definisjoner av støy atferd. Videre, siden det ikke involverer konvergens ganger, kolonnen-klok snitt kan gi en mer pålitelig representasjon av utbruddet av nevrale entrainment. Derfor, resultatene oppnådd med kolonne-klok gjennomsnitt metoden kan betraktes som bakken sannheten for å analysere dynamikken i nevrale entrainment bruker Kalman filtrering.
Denne beskrivelsen av protokollen er basert på et eksempel på SSVEP. Det er imidlertid viktig å merke seg at metoden som presenteres her er modalitet-uavhengige, slik at den kan også brukes til å analysere tiden utviklingen av SSSEP og ASSR.
Protocol
Den nåværende studien ble utført under godkjennelse av forsknings-og etikk komitéen av Universidad de Valparaíso, Chile (vurderings Oppgjørs kode CEC170-18), bekreftet til de nasjonale retningslinjer for forskning med menneskelige.
1. forberedelse
- Velkommen emnet.
- Forklar hvilke mål og relevans studien har. Gi en beskrivelse av relevante tekniske detaljer. Svar på alle spørsmålene grundig.
- Eksplisitt nevne at hun/han kan avbryte den eksperimentelle økten når som helst hvis ønskelig.
- Be den frivillige om å lese emnet informert samtykke og signere tilsvarende skjema. Avbryte eksperimentell økt hvis informert samtykke ikke er innhentet.
2. forberedelse
- Be faget om å sitte i en laboratorie stol i en komfortabel stilling.
- Rengjør hodebunnen med etanol (en løsning på 95%) å fjerne laget av døde hudceller og talg som dekker det. Dette trinnet er viktig å redusere impedans mellom elektrodene og hodebunnen.
- Mål hodet omkretsen med et målebånd for å definere størrelsen på elektrode lokket som skal brukes.
- Be motivet bruke elektrode lokket. Gi instruksjoner for komfortabel, men riktig plassering av hetten.
- Mål avstanden mellom nasion (NZ, midten punktet av nasofrontal Sutur, som kan identifiseres ved depresjonen mellom øynene og toppen av nesen) og inion (iz, prominence av occipital bein) ved hjelp av en målebånd.
- Mål avstanden mellom venstre og høyre pre-auricular poeng (identifisert som depresjonen like før auricle av ørene) ved hjelp av en målebånd.
- Korriger plasseringen av elektrode lokket, slik at skjæringspunktet mellom de imaginære linjene som er definert i de forrige trinnene, tilsvarer toppunktet på hodet. Sørg for at motivet er behagelig etter justeringene.
- Sett ledende gel i elektroden holdere, i henhold til de stedene som vurderes for eksperimentet.
- Bruk 64 hodebunnen steder etter International 10-20 system31 til å bruke utfallet av protokollen til å utføre kilde lokalisering analyse. Det høyeste antallet elektrode plasseringer (128) på hodebunnen kan brukes ved behov.
- Implementer kliniske eller Oppgående innstillinger (med bare noen få elektroder) hvis kilde lokaliserings analyse ikke er planlagt. Bruk occipital lokasjoner til å registrere SSVEP, timelige lokasjoner for å anskaffe ASSR og parietal plasseringer for å registrere SSSEP.
- Skyv elektrodene i elektrode beholderne. Kontroller at etiketten på elektroden samsvarer med etiketten til posisjonen i lokket.
- Følge den frivillige til det eksperimentelle rommet (fortrinnsvis en skjermet, lyd-dempes kammer). Be faget om å sitte i en stol inne i rommet, i en komfortabel posisjon.
- Plasser eksterne elektroder på nesen og øreflippene hvis en fysisk referanse (forskjellig fra hodebunnen elektroder) vil bli brukt for å re-referere til EEG innspillingen (i trinn 3.8.1).
- Plasser eksterne elektroder på periokulære steder.
- Plasser elektroder på kinnet og frontal regionen av hodet, ca 1 cm over øyenbryn, for å spille inn blinkende (i trinn 2.6.1).
- Plasser elektroder på den ytre canthus i øynene, ca 1 cm over/under midtlinjen, for å registrere øyebevegelser (i trinn 2.6.1).
Merk: electrooculogram (EOG) vil bli brukt i trinn 3.8.5 for fjerning av EGG-artefakter indusert av blinkende og øyebevegelser.
- Slå EEG oppkjøpet systemet på og kontrollere elektroden impedans hvis en lav-impedans system brukes for innspilling av EEG. Korriger impedans, etter behov, i henhold til produsentens anvisninger. Impedans bør holdes under 10 kΩ32.
- Be motivet om å blinke og bevege øynene i forskjellige retninger for å sikre at EOG blir riktig registrert.
- For å analysere dynamikken i SSVEP, justere plasseringen av skjermen i vertikal retning, for å matche visningsvinkelen for motivet. Demp lysene på rommet til et behagelig nivå er oppnådd. Juster lysstyrken på skjermen til den øvre grensen på deltakerens komfort-nivå.
- For å analysere dynamikken i ASSR, sett inn øretelefonene med riktig skuminnlegg, slik at øretelefonene passer øre kanalen. Kontroller at lydene er levert med ønsket intensitet (f.eks. en psykofysiske behagelig nivå33).
3. EEG oppkjøp og pre-prosessering
- Angi stimulans parametere definert i eksperimentell design. Se i brukerhåndboken som er gitt av produsenten av stimulering systemet for detaljer om programvaren.
Merk: for omfattende forklaringer av stimulans som brukes for generering av SSVEP og ASSR, se henholdsvis Norcia et al.8 og Rance34. - Instruere faget til å ta hensyn til stimulering, i tilfelle at visuelle entrainment er temaet for eksperimentet.
- Presenter en film med teksting med lyden når hørbar entrainment er tema for eksperimentet.
Merk: presentasjon av en stumfilm som gjør det mulig å følge oppmerksomheten fra den akustiske stimulering og samtidig opprettholde opphisselse nivå25.
- Presenter en film med teksting med lyden når hørbar entrainment er tema for eksperimentet.
- Presentere stimuli lengre enn 90 s, som har blitt gjort for å undersøke tiden utviklingen av SSVEPs og ASSR i både mennesker og dyremodeller21,22,25,26.
Merk: Presenter stimuli kortere i varighet hvis en pilotstudie er utført. - Pause stimulering i 2 minutter hvis bare en eksperimentell tilstand blir testet. Samhandle med motivet for å sjekke bevisstheten.
Merk: varigheten av pausen avhenger av varigheten av stimulering. Pauser 3x lengre enn stimulering intervallene vil sikre at et svar elicited av en stimulans ikke er påvirket av den forrige stimulering. Lengre pauser er tillatt hvis motivet så forespørsler.- Pause stimulering i minst 10 s når ulike eksperimentelle forhold er testet siden vekslende stimulering med pauser på 10 s har blitt foreslått å redusere ekstra tilpasning effekter og redusere lengden på eksperimentet25.
- Gjenta presentasjonen trinnene (trinn 3.3-3.4) minst 30x for å sikre høy SNR av målingene etter gjennomsnitt av epoker (trinn 4,4).
- Record EEG bruke standard prosedyrer35. Opprett en egen EEG-fil for hver eksperimentelle kjøring.
Merk: du finner mer informasjon om programvaren i brukerhåndboken for anskaffelses systemet. - Overvåk EEG-opptaket for å oppdage søvn perioder basert på nivået av Alfa aktivitet og hvor ofte blinkende artefakter vises. Pause eksperimentet når økt Alpha nivåer ledsaget av redusert blinkende frekvenser oppdages, noe som indikerer søvnighet. Avvis den eksperimentelle kjøringen fra videre analyse når søvn perioder oppdages.
- Beregn amplituden til fast-State-responsen på slutten av hver eksperimentelle kjøring, og følg instruksjonene i brukerhåndboken for anskaffelses programvaren som brukes i eksperimentet.
- Overvåk oppmerksomhets nivået av faget ved å sammenligne amplitude av steady-state responser innhentet på slutten av hver eksperimentell kjøre. Angi fast-State amplitude innhentet i den første eksperimentelle kjører som en referanse amplitude.
- Angi en avvisnings terskel (en reduksjon i respons amplituden på 5% i forhold til referanse amplituden). Avvis den eksperimentelle går der amplituden til steady-state respons møter avvisning criterium.
- Finish den eksperimentelle økten etter å anskaffe antall runs definert i eksperimentell design.
- Pre-Process EEG data frakoblet ved hjelp av standard EEG prosedyrer35 beskrevet i de neste trinnene i henhold til produsentens anvisninger.
- Re-referanse innspillingen med en gjennomsnittlig referanse (gjennomsnitt av alle opptaks elektroder) eller gjennomsnittet av et delsett av elektroder. Du kan også bruke en fysisk referanse (f.eks. eksterne elektroder plassert på nesen og øreflippene beskrevet i trinn 2,4).
- Konverter elektrode koordinatene til det internasjonale 10/20-systemet hvis radial koordinatsystemet ble brukt under EEG-oppkjøp. Se produsentens bruksanvisning for detaljer om konvertering.
- Band-Pass filtrere EEG signalet mellom 0,5-300 Hz. Still inn et hakk-filter (sentrert på 50 Hz eller 60 Hz) om nødvendig.
- Ned-sample EEG-signalet for å redusere utførelsestiden for algoritmen som er valgt for fjerning av øye artefakter (trinn 3.8.5).
Merk: en samplingsfrekvens på 512 Hz er tilstrekkelig til å analysere hjernens svingninger frekvens under 40 Hz35. - Fjern øye gjenstandene.
Merk: for dette formål, kan ulike teknikker brukes (se Urigüen og Garcia-Zapirain35 for en omfattende gjennomgang av gjenstand fjerning algoritmer). Blant dem, uavhengig komponent analyse er en av de mest utvidede metoder og er implementert i både kommersielle og gratis analyseprogramvare37,38,39. - Segmentere EEG data i epoker tid-låst til stimulering. Velg epoker lengde i henhold til formålet med eksperimentet.
Merk: epoker bør være tilstrekkelig lang til å tillate analyse av steady-state respons i frekvens-domene med en tilstrekkelig Spectral oppløsning. - Ikke Kjør algoritmer for å avvise artefakter på dette stadiet for å oppdage og fjerne epoker som inneholder artefakter.
Merk: å fjerne epoke på dette stadiet vil indusere feil når datasettet er organisert for å kjøre kolonne-klok gjennomsnitt av epoker (trinn 4,2 og 4,4). Algoritmer for avvisning implementeres ved et senere behandlingstrinn (trinn 4.1.4). - Kjør DC-detrend-funksjonen for å beregne DC-trender i individuelle EEG-epoker og korrigere dem.
- Kjør funksjonen for grunnlinje korrigering for å korrigere grunnlinjen for innspillingen. Velg pre-stimulans tidsintervaller lenger enn 200 MS.
Merk: grunnlinje korrigering består av snitt dataene i det valgte tidsintervallet. Gjennomsnittet beregnes for hver kanal og trekkes fra hvert datapunkt i hver epoke.
4. beregning av responsen amplituder
- Angi parameterne som er nødvendige for beregningen av de steady-state-svarene (figur 1a).
Merk: internt kode som brukes til behandling av data er fritt tilgjengelig på < https://figshare.com/prosjekter/Steady-state_visually_evoked_potentials_SSVEP_elicited_in_humans_by_continuos_light_modulated_in_amplitude_at_10Hz/62573 >. Se hjelpeteksten i koden for ytterligere instruksjoner. På samme måte er et delsett av dataene som brukes i denne studien, tilgjengelige.- Angi antall innspillinger (eksperimentelle kjøringer) av eksperimentet.
- Angi lengden på epoker for å segmentere de enkelte innspillingene.
- Angi samplingsfrekvensen for eksperimentet.
- Velg algoritmer for å avvise artefakter for å oppdage og fjerne epoker som inneholder artefakter. De tilgjengelige utvalgskriteriene er 1) gradering (absolutt forskjell mellom to påfølgende prøver), 2) Max-min (forskjellen mellom maksimum og minimum amplitude i epoken), og 3) amplitude (absolutt maksimum og minimum amplituder).
- Kjør behandlingskoden.
Merk: trinn 4.2-4.7 utføres automatisk når dette alternativet er valgt. Kjør trinnene manuelt hvis det er aktuelt.
- Re-arrangere epoker i en data Matrix av n rader og m kolonner, der n representerer antall innspillinger (eksperimentelle runs) og m antall epoker (figur 1B).
- Vekt epoken å dempe effekten av bevegelse og muskuløse gjenstander.
Merk: vektet EEG epoker oppnås ved å dele hver spennings prøve av amplitude variansen av epoken de tilhører, slik at variansen brukes som et mål på amplitude variasjon og vekting faktor40. - Kolonne-messig gjennomsnitt datasettet. For dette formål er gjennomsnittet av tids domenet epoker som tilsvarer samme tidsvindu i de forskjellige innspillingene.
Merk: dette trinnet gjør det mulig å beregne steady-state amplitude i innspillinger med et bemerkelsesverdig høyt signal-til-støy-forhold (SNR). - Eksporter tidsserien som følge av gjennomsnittet for videre analyse av tids utviklingen av entrainment i ekstern programvare.
- Beregn amplituden til steady-state respons i hver epoke som følge av kolonnen-klok gjennomsnitt, ved hjelp av rask Fourier Transform (FFT).
Merk: FFT lengden skal tilsvare lengden av en epoke. Gjennomføringen av en vindaugesystemet teknikk er ikke obligatorisk. Amplituden til steady-state responsen er definert som Spectral amplitude oppnådd ved hyppigheten av amplitude moduleringshjul av sensoriske stimuli.
- Beregn amplituden til steady-state respons i hver epoke som følge av kolonnen-klok gjennomsnitt, ved hjelp av rask Fourier Transform (FFT).
- Vector gjennomsnitt amplituden av et ad-hoc-antall FFT-hyller på hver side av hyppigheten av responsen for å beregne gjenværende støynivå (RNL). Antall FFT-hyller må samsvare med et frekvensbånd på ca. 3 Hz, på hver side av frekvensen til responsen.
Merk: den høye frekvensen-spesifisitet av steady-state responser gjør responsen amplitude uavhengig av disse bakgrunns svingninger med lignende frekvenser, som igjen distribuerer jevnt i et relativt smalt frekvensbånd41 , 42 for alle , i 43. - Plot amplituden av steady-state respons og RNL som en funksjon av kolonnen indeks (dvs. antall ervervet epoken) for å utforske utviklingen av steady-state respons under stimulering intervall.
Representative Results
SSVEP ble elicited av kontinuerlige visuelle stimuli av 40 s i lengde, der lys intensiteten ble modulert av en sinusformet bølge på 10 Hz (modulerings dybde på 90%). Stimuli ble levert av fire Light-Emitting dioder (lysdioder) ligger i sentrum av en 50 cm x 50 cm svart skjerm, som ytterpunkt av en 5 cm x 5 cm kvadrat. Når deltakeren satt 70 cm fra skjermen, arealet av kvadratet av lysdioder subtends en visuell vinkel på ca 4 °. LED-skjermen ble designet ved hjelp av en USB-basert microcontroller utviklingssystem og fire super lyse hvite lysdioder på 10 mm diameter. Den pulsbreddemodulering (PWM) teknikken ble brukt til å styre strøm levert til lysdioder. Denne teknikken kontrollerte lysene intensitet på en gitt frekvens og generere den endelige sinusformet konvolutten. En PWM-frekvens på 40 kHz ble brukt for å unngå en forstås flimmer effekt.
Tretti innspillinger ble innhentet, som var segmentert i epoker av 4 s. Derfor ble et datasett bestående av 10 kolonner (antall EEG-epoker i innspillinger) og 30 rader (antall innspillinger, antall eksperimentelle kjøringer) oppnådd.
Den nevrale pendling tid-låst til stimulering ble tydelig som kolonne-klok snitt ble utført (figur 2). Betydelig, det intervallet som SSVEP genereres kan observeres i spor som samsvarer med kolonne 1. I denne kolonnen, 0,2 s av pre-stimulans Baseline er plottet i tillegg til den første 0,8 s av nevrale entrainment. Derfor fremgangsmåten beskrevet her kan karakterisering av 1) dynamikken i oscillasjon respons gang nevrale entrainment er allerede etablert og 2) engasjement av nevrale svingninger. En eller flere epoker registrert etter slutten av stimulering kan også inkluderes i data Matrix å studere utryddelse av steady-state respons etter stimulans offset.
Under den kolonne-klok gjennomsnitt av epoker, gjennomsnittlig amplitude av SSVEP (Spectral amplitude ved 10 Hz, beregnet ved å bruke FFT) redusert i snitt av den første epoker av kolonnene og tendens til å stabilisere etterpå (figur 3a). Dette resultatet er enig med tidligere studier analysere utviklingen av ASSR i snitt av sekvensielt ervervet epoker21,22,40,43,44. Atferden til responsen amplitude undersnitt er vanligvis forklares med relativt høye bidrag av unaveraged støy til responsen amplitude beregnet i den første epoker, som er svekket som snitt utføres13, 44 for alle , 45 for alle , 46 for alle , 47. BEMERKELSESVERDIG, SSVEP amplitude variasjon betydelig redusert som gjennomsnitt utviklet.
Vi har også analysert RNL av målingene i kolonnen-klok gjennomsnitt av epoker (figur 3b). RNL ble beregnet i et smalt frekvensbånd (3 Hz) på begge sider av frekvensen til SSVEP. Selv om denne prosedyren er ikke vanlig når SSVEP er analysert, vektor-gjennomsnitt et gitt antall frekvens binger rundt at av Neural entrainment er standard for estimering av RNL i ASSR målinger41,42, 43. som forventet, RNL gradvis redusert som antall gjennomsnitt epoker økt og nådde asymptotiske nivå etter ca 20 epoker ble behandlet. I motsetning til det som ble observert da SSVEP-amplituden ble analysert, forble standardavviket til RNL relativt konstant ettersom antall gjennomsnittlige epoker økte, noe som tyder på at opptaksforholdene var stabile langs den eksperimentelle sesjonen.
Resultatene presentert ovenfor bestemt endringene i peak signal-til-støy-forhold (pSNR) av målinger i den kolonnen-klok gjennomsnitt av epoken (Figur 3C). Dette begrepet er definert her som forholdet (i dB) mellom den firkantede amplitude av responsen (SSVEP) og kvadrat amplitude av RNL. Som gjennomsnitt utviklet, pSNR økt som antall gjennomsnitt epoker økt opp til 18, ca. Ytterligere trinn i antall gjennomsnitt epoker ikke signifikant innvirkning på kvaliteten på signalet. Variasjonen av pSNR ble redusert etter hvert som flere epoker var i gjennomsnitt.
Til slutt, dynamikken i SSVEP amplitude og RNL er representert i Figur 4. Disse tid videreutviklinger ble innhentet ved å plotte responsen parametrene beregnet på slutten av kolonnen-klok gjennomsnitt av epoker som en funksjon av antall kolonner (som en funksjon av tid). Som demonstrert av Labecki et al.26, kan DYNAMIKKEN i SSVEP signifikant variere blant. Siden resultatene presentert i Figur 4 tilsvarer en enkelt person, kan generaliseringer ikke gjøres. I dette emnet, amplitude av SSVEP vist en relativt kompleks atferd (figur 4a). Responsen amplitude gradvis økt i løpet av de første 12 sekundene etter stimulans utbruddet (tid som tilsvarer lengden av 3 epoker). Som stimulans faste, SSVEP konsekvent redusert i løpet av følgende 12 sekunder, og forble relativt konstant etterpå. Disse resultatene kan ikke forklares med oppførselen til RNL, siden denne parameteren var relativt konstant under stimulering intervallet (figur 4b). Økningen i SSVEP amplitude etter stimulans utbruddet er tydelig i sporene presentert i figur 2 og kan forklares ved integrasjons prosesser, som resulterer i stabilisering av nevrale entrainment. Den påfølgende reduksjonen i amplitude antyder tilpasningen av SSVEP til den vedvarende stimulering. Likevel, disse hypoteser må testes i kontrollerte eksperimenter med bevilget utvalgsstørrelser.
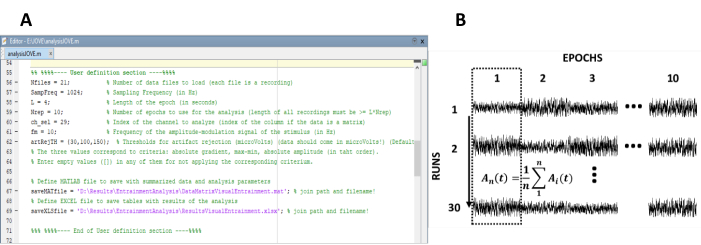
Figur 1 : Kritiske skritt for å trekke ut tiden utviklingen av amplituden til steady-state responser. (A) skjermbilde av behandlingen koden, der analyse parametere er definert. (B) representative diagram som illustrerer organiseringen av datasettet. En data matrise bestående av 30 innspillinger av 10 epoker representeres. Kolonne-klok snitt av epoker er uthevet i den første kolonnen. Den vertikale linjen representerer retningen av snitt. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 2 : Endringer i bølgeform av steady-state visuelt fremkalt potensial (SSVEP) under kolonnen-klok gjennomsnitt av epoker. Responsen ble elicited av kontinuerlig presentasjon av lys modulert i amplitude ved 10 Hz. Radene viser bølgeformene oppnådd etter snitt alle tidligere innspillinger (dvs. rad 1 er den første innspillingen, rad 5 er bølgeformen innhentet etter snitt de første fem opptakene, og den siste raden er gjennomsnittet av alle innspillinger). Mer pålitelige bølgeformer av SSVEP ble observert i hver kolonne som antall snitt går økt. For å gi klarhet (for å gjøre svingninger av SSVEP synlig), er bare første sekund av epoker representert. Unntakene er spor i den første kolonnen i datasettet, som 0,2 sekunder av før stimulans grunnlinjen vises. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 3 : Endringer i respons-og opptaks parametrene under den kolonne-klok snitt av epoker. (A) utviklingen av SSVEP amplitude. (B) oppførselen til RNL. (C) endringer i pSNR. Svarte linjer representerer middelverdiene som oppnås for hver kolonne (n = 10), og den grå skyggen representerer området som dekkes av ± ett standardavvik. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 4 : Tid utviklingen av SSVEP elicited ved presentasjon av kontinuerlig visuell stimulering, modulert i amplitude ved 10 Hz. (A) tid løpet av SSVEP amplitude. (B) tid løpet av RNL. Vennligst klikk her for å se en større versjon av dette tallet.
Discussion
Dette arbeidet beskriver en eksperimentell prosedyre for å analysere dynamikken i oscillasjon hjernen reaksjoner. Slik metodikk består av å anskaffe et tilstrekkelig antall uavhengige eksperimentelle kjøringer av samme eksperimentelle tilstand, og tids-domene snitt epoker tilsvarende til samme tid vindu i de ulike opptakene (kolonner-klok gjennomsnitt i Figur 1B). Amplituden som beregnes i gjennomsnitts dataene representerer den umiddelbare amplituden til oscillasjon respons. Plotting disse amplituder som en funksjon av tid (eller antall kolonner i datasettet) kan analysere tiden utviklingen av oscillasjon responstid-låst til stimulering. Denne metodikken er en modifikasjon av det som foreslås av Ritter et al.23 for å analysere tilpasningen av forbigående kortikale fremkalt potensialer. Metoden har blitt brukt til å analysere dynamikken i hørbar fremkalt potensial i både mennesker24 og dyremodeller20,21.
Fra en metodisk synspunkt, kombinasjonen av parametere som brukes til å lokke fram steady-state respons og de iverksatt for å trekke ut neural respons fra bakgrunnsstøy er avgjørende for å analysere tiden utviklingen av steady-state fremkalt potensialer 22. stimulans lengden som brukes i eksperimentet presenteres her (40 s) ble valgt basert på resultater oppnådd i en pilotstudie. Denne stimulans lengden var tilstrekkelig til å analysere tilpasningen av ASSR generert i rotte hjernestammen21,22. Videre bør stimulans lengden overstige den tiden hvor asymptotiske momentant bandet makt SSVEPs er nådd (figur 1 i Labecki et al.26). Likevel, den asymptotiske momentant bandet makt SSVEPs kan nås utover 60s i noen tilfeller (figur 2 i Labecki et al.26). Derfor, kjører en liten prøve pilotstudie anbefales å definere stimulans lengden på stimulering. Ellers er en stimulans lengde lengre enn 90 s anbefales å oppnå fullstendig representasjon av tiden utviklingen av responsen. Bruke tilstrekkelig lange pauser mellom påfølgende innspillinger innebærer vurderer påfølgende eksperimentelle går som å være statistisk uavhengige (dvs. forskjellige, uavhengige tiltak av samme variabel). Etter beste viten, har ingen eksperimenter blitt utført for å analysere den optimale pausen mellom løyper (minimum pause kreves for å gjøre kjøringer uavhengig av hverandre). Criterium av å bruke pauser minst 3x lengre enn stimulans lengden er konservativ nok til å sikre at steady-state respons registrert i en gitt kjøre er ikke berørt av den forrige stimulering.
Nylig alternerende stimuli (eksperimentelle forhold) har blitt foreslått som et valg for å redusere pausen mellom eksperimentelle går, unngå ekstra tilpasning effekt25. Likeledes antall eksperimentelle går (30) implementert i denne eksperimentelle protokollen er konservativ, siden asymptotiske RNL og pSNR er vanligvis nådd etter snitt 20 eksperimentelle går, ca. Når stimuli faller innenfor den midterste øvre regionen av det dynamiske spekteret av responsen (høy sensasjon nivåer), lavere Antall kjøringer er sannsynligvis nødvendig for å analysere dynamikken i fremkalt respons. Likevel, i tilfeller der ulike eksperimentelle forhold er testet, har samme antall eksperimentelle går er avgjørende for å gjøre sammenligninger mellom forhold (dvs. ulike sensasjon nivåer).
I tillegg til kolonne-klok gjennomsnitt av epoker, har dynamikken i oscillasjon fremkalt potensialer blitt analysert ved å filtrere enkelt-rettssaken målinger i et smalt frekvensbånd rundt frekvensen av interesse og databehandling konvolutten av strømmen signal ved hjelp av low pass-filtrering26. Likeledes, enkelt prøveanalyse er implementert for å karakterisere overgangen perioden som forut for den stabile regionen SSVEP48, og endringene i amplitude og fase av SSVEP under den stabile regionen av responsen49. Mens en enkelt prøveanalyse tillater diskriminering av relativt raske svingninger i respons amplitude, vil eksperimentell design for å analysere gjennomsnittlig respons i blokker atskilt med et gitt mellom-blokk intervall bare ta hensyn til langsiktige variasjoner i amplituden til det utløste potensialet50,51. Den kolonne-klok gjennomsnitt av epoker står mellom disse to alternativene. Konvertering av gjennomsnittlig signal til frekvens-domene ved hjelp av FFT innebærer å analysere dynamikken i responsen med en oppløsning lik lengden på epoken. I eksempelet som presenteres her, ble SSVEP rapportert hver 4 s. Selv om 4 s av oppløsning er tilstrekkelig for å beskrive dynamikken oppstår i intervaller på tid overgå titalls sekunder, slik som for SSVEP26, delvis overlappende epoker i den opprinnelige opptakene gjør det mulig å beskrive tiden utviklingen av steady-state respons på en mer raffinert måte25.
Dynamics av steady-state responser innhentet etter kolonne-klok snitt av epoker hovedsak representerer utviklingen av oscillasjon aktivitet som er synkronisert blant de gjennomsnitt EEG segmenter (de som overlever i snitt). Derfor er et stort problem om muligheten for metodikken er mulig demping av respons amplituder på grunn av variasjoner i fasen av Neural svingninger fra en uavhengig eksperimentell kjøre til en annen (dvs. blant innspillinger). Dette emnet må rettes eksperimentelt. Imidlertid tyder bevis på at fasen av hjernen oscillasjon svar er mindre variabel enn forventet. Faktisk har flere studier rapportert en regularitet i den forventede fasen av den menneskelige 80 Hz ASSR47,48,49. Når latencies er estimert basert på fasen av den oscillasjon aktiviteten, har den forutsigbare effekten av intensiteten og bære frekvensen for de akustiske stimuli på ventetiden til hørsels responsen blitt observert (dvs. forsinkelsen reduseres etter hvert som intensitet og carrier frekvens økning)52,53,54. Videre er det også observert typiske maturational endringer i amplituden og den venstre-mot-høyre-asymmetri i hørsels nivåene når latencies er estimert fra fasen av ASSR47,55,56 , 57 for alle , 58. ved beskrivelse av tids utviklingen til SSVEP ved hjelp av en analyse med én prøveperiode, observerte Labecki et al.26 at selv om variasjon i respons amplituder innenfor samme var betydelig høy, var variasjonen i fase var betydelig mindre uttalt.
Basert på deres observasjoner, Labecki et al.26 antydet at et minimum av 50 studier bør være gjennomsnitt for å få en pålitelig estimering av gjennomsnittlig strøm konvolutt av responsen. Disse resultatene tyder på at selv når amplituden til svaret er beregnet i enkle forsøk, er gjennomsnitt (av konvolutter i dette tilfellet) nødvendig for å rapportere pålitelige resultater. Videre, den Inter-rettssaken variasjon i amplitude av SSVEP rapportert av Labecki et al.26 antyder at beregningen av denne parameteren i enkelt forsøk kan være svært påvirket av bakgrunnsstøy. Tatt i betraktning utviklingen av signal-til-støy-forholdet presentert i figur 2, reduserer beregningen av responsen i gjennomsnitt signalet i stedet for enkelt forsøk betydelig antall EEG segmenter som trengs for å bli behandlet for å oppnå pålitelig Målinger. I tillegg, den lave variasjonen i fase oppnådd ved Labecki et al.26 støtter ideen om at kolonnen-klok gjennomsnitt av epoker som presenteres her er en gyldig prosedyre for databehandling dynamikken i oscillasjon fremkalt potensialer.
Gjennomsnitt dataene på ulike nivåer fører til ulik tolkning av resultatene. Når det gjelder oscillasjon vakte potensialer, databehandling responsen amplitude etter tids domenet snitt av uavhengige kjøringer innebærer å analysere bare tid-låst svingninger (dvs. de som overlever i snitt). Denne prosedyren kan filtrere relevant informasjon om dynamikken i responsen i individuelle forsøk. Det garanterer imidlertid et tilstrekkelig høyt signal-til-støy-forhold for målingene. Dette aspektet kan være av betydning når svarene er nær elektrofysiologisk terskelen, en tilstand der påvisning av entrainment kan bli kompromittert på grunn av lavt signal-til-støy-forhold av målingen.
Disclosures
Forfatterne har ingenting å avsløre.
Acknowledgments
Forfatterne takknemlig erkjenner Lucía Zepeda, Grace A. Whitaker, og Nicolas Nieto for deres bidrag til video produksjon. Dette arbeidet ble støttet delvis av CONICYT programmer BASAL FB0008, MEC 80170124 og PhD-stipend 21171741, samt National Institute on døvhet og andre kommunikasjons forstyrrelser av National Institutes of Health under tildeling nummer P50DC015446. Innholdet er utelukkende ansvaret til forfatterne og representerer ikke nødvendigvis den offisielle synspunktene til National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
References
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G. Jr, Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. An introduction to the event-related potential technique. Second Edition. , MIT Press. 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J. Jr, Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
