

Summary
Viene presentato un protocollo per valutare l'evoluzione temporale dell'ingresso neurale verso stimoli ripetitivi esterni. Le registrazioni di stato fisso della stessa condizione sperimentale vengono acquisite e mediate nel dominio temporale. Le dinamiche dello stato fisso vengono analizzate tracciando l'ampiezza della risposta in funzione del tempo.
Abstract
L'intrappolamento neurale si riferisce alla sincronizzazione dell'attività neurale con la periodicità degli stimoli sensoriali. Questa sincronizzazione definisce la generazione di risposte evocate allo stato costante (cioè oscillazioni nell'elettroencefalogramma che si bloccano in fase agli stimoli di guida). L'interpretazione classica dell'ampiezza delle risposte allo stato costante presuppone una risposta neurale stereotipata invariante nel tempo più fluttuazioni di fondo casuali, in modo tale che la media su ripetute presentazioni dello stimolo recuperi il risposta stereotipata. Questo approccio ignora la dinamica dello stato stazionario, come nel caso dell'adattamento suscitato da esposizioni prolungate allo stimolo. Per analizzare la dinamica delle risposte allo stato stazionario, si può presumere che l'evoluzione temporale dell'ampiezza di risposta sia la stessa in diversi cicli di stimolazione separati da pause sufficientemente lunghe. Sulla base di questo presupposto, viene presentato un metodo per caratterizzare l'evoluzione temporale delle risposte in stato stazionario. Un numero sufficientemente elevato di registrazioni viene acquisito in risposta alla stessa condizione sperimentale. Le esecuzioni sperimentali (registrazioni) sono mediate a colonne (ad esempio, le esecuzioni sono mediate, ma l'epoca all'interno delle registrazioni non viene mediata con i segmenti precedenti). La media a colonne consente l'analisi delle risposte allo stato fisso nelle registrazioni con rapporti segnale-rumore notevolmente elevati. Pertanto, il segnale mediato fornisce una rappresentazione accurata dell'evoluzione temporale della risposta allo stato costante, che può essere analizzata sia nei domini di tempo che in quelli di frequenza. In questo studio, viene fornita una descrizione dettagliata del metodo, utilizzando potenziali evocati visivamente allo stato costante come esempio di risposta. Vantaggi e avvertenze vengono valutati sulla base di un confronto con metodi a prova singola progettati per analizzare l'ingresso neurale.
Introduction
Quando viene registrata dal cuoio capelluto, l'attività elettrica del cervello è osservata come cambiamenti continui e regolari nelle tensioni nel tempo. Questa attività elettrica è chiamata elettroencefalogramma (EEG) ed è stata descritta per la prima volta da Hans Berger alla fine degli anni venti del secolo scorso1. Successivi studi seminali hanno descritto l'EEG come una serie temporale composta, in cui si possono osservare diversi modelli ritmici o ripetitivi2,3,4. Al giorno d'oggi, l'EEG è tipicamente diviso in cinque bande di frequenza ben consolidate, delta, theta, alfa, beta e gamma, che sono associate al diverso processo sensoriale e cognitivo.
Per anni, lo studio delle oscillazioni cerebrali utilizzando EEG è stato limitato a due analisi dello spettro nell'attività in corso o ai cambiamenti nell'attività oscillatoria suscitati da eventi sensoriali non periodici. Negli ultimi decenni sono state implementate diverse metodologie per modulare le oscillazioni EEG in corso ed esplorare gli effetti di tali modulazioni sui processi percettivi e cognitivi, compresa la presentazione della stimolazione sensoriale ritmica per indurre l'intrappolamento neurale. Il termine rientro neurale si riferisce alla sincronizzazione dell'attività neurale con le proprietà periodiche degli stimoli sensoriali. Questo processo porta alla generazione di potenziali evocati a stato costante (cioè oscillazioni EEG bloccate alle proprietà periodiche degli stimoli di guida). I potenziali evocati da stato costante sono più comunemente provocati dalla stimolazione visiva, uditiva e vibrotilativa, usando sia stimoli transitori presentati a un ritmo costante o stimolazione continua modulata in ampiezza alla frequenza di interesse. Mentre i potenziali somatosensoriali con rimorsi costanti (SSSEP) sono registrati in risposta alla stimolazione tattile ripetitiva5,6, i potenziali sottoposti a residuo visivamente evocati (SSVEP) sono generalmente suscitati dal periodico presentazione di sfarfallii di luminanza, immagini e volti7,8. Le risposte uditiche a stato stabile (ASSR) sono solitamente generate da treni di stimoli acustici transitori o dalla presentazione continua di toni modulati in ampiezza9,10.
L'estrazione di potenziali evocati dallo stato stazionario EEG si basa essenzialmente sulla media di ePochs EEG successivamente acquisita, bloccate nel tempo allo stimolo11. A causa della periodicità delle risposte, possono essere analizzate sia nei domini di tempo che in quelli di frequenza. Dopo la trasformazione frequenza-dominio, la risposta sensoriale viene osservata come picchi di ampiezza alla velocità di presentazione o alla frequenza di modulazione degli stimoli esterni e le armoniche corrispondenti. Queste procedure (media tra domini temporali e successiva trasformazione frequenza-dominio) sono state essenziali per lo sviluppo di un test dell'udito basato sul rilevamento di metodi ASSR con scopi clinici12,13,14 ,15,16.
Inoltre, la media classica del dominio temporale delle epoche EEG è stata estremamente utile per analizzare i processi fisiologici come la generazione e l'estinzione di SSVEP17,18. Presentando treni consecutivi di luci sfarfallio e media di epoche successive all'interno di una registrazione, Wacker et al.19 ha osservato che l'indice di blocco di fase del SSVEP è aumentato rapidamente durante i primi 400 ms di stimolazione ed è rimasto alto in seguito . Hanno anche riferito che è stato stabilito un robusto intrappolamento visivo tra 700-1 100 ms dopo l'insorgenza di stimoli. Un certo grado di intrappolamento è rimasto efficace dopo l'offset del treno di stimolazione, che è durato circa tre periodi della risposta oscillatoria17,19. Tali comportamenti sono stati interpretati come l'effetto coinvolgente/disinnesto delle oscillazioni osservate, che è una conseguenza dell'elaborazione delle informazioni non lineari nel sistema visivo umano17. In alternativa, è noto che in determinate condizioni sperimentali, la stimolazione dello sfarfallio può suscitare risposte all'inizio e off-risposta alla fine dei treni di stimolazione invece di intrappolamento neurale18.
L'ipotesi principale di epoche EEG acquisite consecutivamente nella media è che il segnale EEG rappresenta una combinazione lineare della risposta sensoriale e del rumore di fondo20. Inoltre, si presume che l'ampiezza, la frequenza e la fase della risposta oscillatoria siano stazionarie, mentre il rumore di fondo è considerato un'attività casuale. Tuttavia, nei casi in cui questa ipotesi non è soddisfatta, l'ampiezza di risposta calcolata dopo diverse epoche non corrisponde necessariamente all'ampiezza istantanea del potenziale evocato.
È stato recentemente riportato che l'ASSR generato nel tronco encefalico dei ratti si adatta alla continua presentazione di toni modulati con ampiezza (cioè, l'ampiezza di risposta esponenzialmente nel tempo)21,22. L'adattamento è stato interpretato come un meccanismo neurale che riflette la perdita di novità di uno stimolo sensoriale monotonamente ripetitivo, aumentando la sensibilità alle fluttuazioni rilevanti nell'ambiente acustico23,24. Nel percorso uditivo, l'adattamento può migliorare la comprensione del linguaggio in ambienti rumorosi. Inoltre, questo processo può essere una parte dei meccanismi esistenti per monitorare il feedback uditivo della propria voce per controllare la produzione vocale.
Analizzando l'evoluzione temporale dell'ASSR a 40 Hz negli esseri umani, Van Eeckhoutte et al.25 ha osservato una significativa ma piccola diminuzione dell'ampiezza di risposta nel tempo (circa -0.0002 V/s basata sull'analisi di gruppo, quando assumendo una diminuzione lineare nel tempo). Di conseguenza, questi autori hanno concluso che l'ASSR a 40 Hz nell'uomo non si adatta alla stimolazione. Nell'uomo, sono stati osservati comportamenti non stazionari durante l'analisi della stabilità dell'SSVEP26. Questi autori hanno osservato che l'ampiezza della frequenza fondamentale e la seconda armonica dell'SSVEP erano stazionarie solo nel 30% e nel 66,7% dei soggetti testati, rispettivamente. Le fasi di entrambi i componenti di frequenza SSVEP, anche se relativamente stabili nel tempo, hanno mostrato piccole derive26.
Pertanto, anche se la media classica del dominio temporale delle epoche successivamente acquisite consente di esplorare le proprietà stazionarie dell'interioramento neurale, questa metodologia deve essere rivista quando la dinamica a lungo termine dell'intrappolamento è al centro del ricerca, o quando la media delle dinamiche a breve termine è corrotta dall'insorgenza di dinamiche a lungo termine. Per caratterizzare i comportamenti non stazionari delle risposte allo stato stazionario, la risposta evocata calcolata in un determinato intervallo di tempo non deve essere compromessa da quelle calcolate nei segmenti EEG precedenti. In altre parole, il potenziale evocato dovrebbe essere estratto dal rumore di fondo senza che le epoche siano mediate nel dominio temporale con i segmenti EEG precedenti.
In questo studio, viene presentato un metodo per valutare la dinamica dell'inserimento neurale. Le risposte dello stato fisso vengono registrate ripetutamente in risposta alla stessa stimolazione, in cui le registrazioni consecutive vengono interfogliate da un intervallo di riposo di tre volte la lunghezza della corsa sperimentale. Considerando che se l'evoluzione temporale della risposta fisiologica è la stessa in diverse esecuzioni sperimentali indipendenti (registrazioni indipendenti), le registrazioni sono in media a colonne. In altre parole, vengono mediate le epoche corrispondenti alla stessa posizione nelle diverse registrazioni, senza una media delle epoche all'interno di una registrazione. Di conseguenza, l'ampiezza di risposta calcolata a qualsiasi intervallo di stimolazione corrisponderà all'ampiezza istantanea del potenziale evocato. Le risposte sensoriali possono essere analizzate nel periodo temporale o trasformate nel dominio di frequenza, a seconda dello scopo dell'esperimento. In ogni caso, le ampiezza possono essere tracciate in funzione del tempo per analizzare l'evoluzione del tempo della risposta allo stato costante. Generazione ed estinzione dei potenziali evocati allo stato costante possono essere valutati limitando l'analisi alla prima e all'ultima epoche delle registrazioni.
La dinamica dell'ingresso neurale può essere analizzata utilizzando altri approcci, come il filtraggio a banda stretta di misurazioni a singola prova intorno alla frequenza di interesse e il calcolo dell'inviluppo del segnale di alimentazione utilizzando il filtraggio a passa basso25 e il Trasformazione di Hilbert27. Rispetto a queste metodologie, la media a colonne delle epoche consente di calcolare i parametri a stato costante in base ai segnali con il più alto rapporto segnale-rumore (SNR). Recentemente, il filtraggio Kalman è emerso come una tecnica promettente per la stima di ampiezza ASSR 40-Hz28,29,30. L'implementazione del filtraggio Kalman può migliorare il rilevamento di risposte allo stato costante più vicino alla soglia elettrofisiologica e ridurre il tempo del test dell'udito29. Inoltre, non è necessario assumere risposte fisse quando si utilizza un approccio di filtraggio Kalman per stimare l'ampiezza dell'ASSR30. Tuttavia, solo uno studio ha analizzato l'evoluzione temporale degli ASSR utilizzando Kalman filtering25. La conclusione dello studio è che l'ampiezza ASSR a 40 Hz è stabile nell'intervallo di stimolazione. Pertanto, il filtraggio Kalman deve essere testato in condizioni in cui l'ASRR non è stazionario.
Anche se richiede molto tempo, il metodo di media per colonna è privo di modelli e non richiede valori di inizializzazione e/o definizioni a priori del comportamento del rumore. Inoltre, poiché non comporta tempi di convergenza, la media a colonne può fornire una rappresentazione più affidabile dell'insorgenza dell'insorgenza dell'insorgenza del rientro neurale. Pertanto, i risultati ottenuti con il metodo di media a colonne può essere considerato come la verità di base per analizzare la dinamica dell'intrappolamento neurale utilizzando il filtraggio Kalman.
Questa descrizione del protocollo si basa su un esempio di SSVEP. Tuttavia, è importante notare che il metodo qui presentato è indipendente dalla modalità, in modo che possa essere utilizzato anche per analizzare l'evoluzione temporale di SSSEP e ASSR.
Protocol
Il presente studio è stato condotto sotto l'approvazione del Comitato di Ricerca ed Etica dell'Universidad de Valparaso, Cile (codice di valutazione CEC170-18), confermato alle linee guida nazionali per la ricerca con soggetti umani.
1. Preparazione
- Accogliete il soggetto.
- Spiegare gli obiettivi e la pertinenza dello studio. Fornire una descrizione dei dettagli tecnici pertinenti. Rispondi a tutte le domande a fondo.
- Menzionato esplicitamente che lei/lui può interrompere la sessione sperimentale in qualsiasi momento, se lo si desidera.
- Chiedi al volontario di leggere il Consenso Informato soggetto e firmare il modulo corrispondente. Interrompere la sessione sperimentale se il consenso informato non viene ottenuto.
2. Preparazione del soggetto
- Chiedere al soggetto di sedersi su una sedia da laboratorio in una posizione comoda.
- Pulire il cuoio capelluto con etanolo (una soluzione al 95%) per rimuovere lo strato di cellule morte della pelle e sebo che lo coprono. Questo passaggio è importante per ridurre l'impedibile tra gli elettrodi e il cuoio capelluto.
- Misurare la circonferenza della testa con un metro a nastro per definire le dimensioni del tappo dell'elettrodo da utilizzare.
- Chiedere al soggetto di indossare il cappuccio dell'elettrodo. Fornire le istruzioni per un corretto ma corretto posizionamento del cappuccio.
- Misurare la distanza tra la nasione (Nz, il punto centrale della sutura nasofrontale, che può essere identificata dalla depressione tra gli occhi e la parte superiore del naso) e l'inione (Iz, la prominenza dell'osso occipitale) utilizzando un metro a nastro.
- Misurare la distanza tra i punti pre-auricolari sinistro e destro (identificato come la depressione appena prima del auricolare delle orecchie) utilizzando un nastro di misurazione.
- Correggere la posizione del tappo dell'elettrodo, in modo che l'intersezione tra le linee immaginarie definite nei passaggi precedenti corrisponda al vertice della testa. Assicurarsi che il soggetto sia confortevole dopo le regolazioni.
- Mettere gel conduttivo nei supporti dell'elettrodo, in base alle posizioni considerate per l'esperimento.
- Utilizzare 64 posizioni del cuoio capelluto dopo il sistema Internazionale 10-2031 per utilizzare il risultato del protocollo per eseguire l'analisi della localizzazione di origine. Il maggior numero di posizioni dell'elettrodo (128) sul cuoio capelluto può essere utilizzato se necessario.
- Implementare impostazioni cliniche o ambulatoriali (con pochi elettrodi) se l'analisi della localizzazione di origine non è pianificata. Utilizzare posizioni occipitali per registrare SSVEP, posizioni temporali per acquisire ASSR e posizioni parietali per registrare SSSEP.
- Spingere gli elettrodi nei supporti dell'elettrodo. Assicurarsi che l'etichetta dell'elettrodo corrisponda all'etichetta di posizione nel cappuccio.
- Accompagnare il volontario nella sala sperimentale (preferibilmente, una camera schermata e attenuata dal suono). Chiedere al soggetto di sedersi su una sedia all'interno della stanza, in una posizione comoda.
- Posizionare elettrodi esterni sul naso e sui lobi delle eari se verrà utilizzato un riferimento fisico (diverso dagli elettrodi del cuoio capelluto) per rifare riferimento alla registrazione EEG (al punto 3.8.1).
- Posizionare gli elettrodi esterni in posizioni perioculari.
- Posizionare gli elettrodi sulla guancia e sulla regione frontale della testa, circa 1 cm sopra il sopracciglio, per registrare lampeggiante (nel punto 2.6.1).
- Posizionare gli elettrodi sul canto esterno degli occhi, circa 1 cm sopra/sotto la linea mediana, per registrare i movimenti degli occhi (nel punto 2.6.1).
NOTA: L'elettrooculogramma (EOG) verrà utilizzato nel passaggio 3.8.5 per rimuovere gli artefatti EGG indotti da lampeggiamento e movimenti oculari.
- Accendere il sistema di acquisizione EEG e controllare l'impedezione dell'elettrodo se viene utilizzato un sistema a basso consumo per la registrazione dell'EEG. Correggere l'impedimento, se necessario, in base alle indicazioni del produttore. Impedance deve essere mantenuto al di sotto di 10 k.
- Chiedere al soggetto di lampeggiare e spostare gli occhi in direzioni diverse per garantire che EOG venga registrato correttamente.
- Per analizzare la dinamica di SSVEP, regolare la posizione dello schermo nella direzione verticale, in modo che corrisponda all'angolo di visualizzazione del soggetto. Smorta le luci della stanza fino a raggiungere un livello confortevole. Regolare il livello di luminanza dello schermo al limite superiore del livello di comfort del partecipante.
- Per analizzare la dinamica dell'ASSR, inserire gli auricolari utilizzando gli inserti di schiuma corretti, in modo che gli auricolari si adattino al condotto uditivi. Verificare che i suoni siano esivi all'intensità desiderata (ad esempio, un livello psicofisico confortevole33).
3. Acquisizione e pre-elaborazione EEG
- Impostare i parametri di stimolo definiti nella progettazione sperimentale. Per informazioni dettagliate sul software, fare riferimento al manuale dell'utente fornito dal produttore del sistema di stimolazione.
NOTA: per spiegazioni complete dello stimolo utilizzato per la generazione di SSVEP e ASSR, vedere Norcia et al.8 e Rance34, rispettivamente. - Istruire il soggetto a prestare attenzione alla stimolazione, nel caso in cui l'intrappolamento visivo sia l'argomento dell'esperimento.
- Presentare un filmato sottotitolato con il suono spento quando l'intrappolamento uditivo è l'argomento dell'esperimento.
NOTA: la presentazione di un filmmuto consente di deviare l'attenzione dalla stimolazione acustica mantenendo il livello di eccitazione25.
- Presentare un filmato sottotitolato con il suono spento quando l'intrappolamento uditivo è l'argomento dell'esperimento.
- Stimoli attuali più lunghi di 90 s, come sono stati fatti per studiare l'evoluzione del tempo di SSVEP e ASSR in entrambi gli esseri umani e modelli animali21,22,25,26.
NOTA: Presentare gli stimoli di durata più breve se è stato eseguito uno studio pilota. - Mettere in pausa la stimolazione per 2 min se si sta testando una sola condizione sperimentale. Interagire con il soggetto per verificare la consapevolezza.
NOTA: la durata della pausa dipende dalla durata della stimolazione. Le pause 3 volte più lunghe degli intervalli di stimolazione garantiranno che una risposta suscitata da uno stimolo non sia influenzata dalla stimolazione precedente. Sono consentite pause più lunghe se l'oggetto lo richiede.- Mettere in pausa la stimolazione per almeno 10 s quando vengono testate diverse condizioni sperimentali poiché è stata proposta la stimolazione alternata con pause di 10 s per diminuire gli effetti di adattamento extra e ridurre la lunghezza dell'esperimento25.
- Ripetere i passaggi di presentazione (passaggi 3.3-3.4) almeno 30x per garantire l'elevato SNR delle misurazioni dopo la media delle epoche (passaggio 4.4).
- Registrare l'EEG utilizzando le procedure standard35. Creare un file EEG separato per ogni esecuzione sperimentale.
NOTA: Fare riferimento al manuale dell'utente del sistema di acquisizione per i dettagli sul software. - Monitorare la registrazione EEG per rilevare i periodi di sospensione in base al livello di attività alfa e alla frequenza con cui appaiono gli artefatti lampeggianti. Metti in pausa l'esperimento quando vengono rilevati livelli alfa aumentati accompagnati da frequenze lampeggianti in diminuzione, il che è indicativo della sonnolenza. Rifiutare l'esecuzione sperimentale da un'ulteriore analisi quando vengono rilevati periodi di sospensione.
- Calcolare l'ampiezza della risposta allo stato fisso alla fine di ogni esecuzione sperimentale, seguendo le istruzioni fornite nel manuale dell'utente del software di acquisizione utilizzato nell'esperimento.
- Monitorare il livello di attenzione del soggetto confrontando l'ampiezza delle risposte allo stato costante ottenute alla fine di ogni corsa sperimentale. Impostare l'ampiezza dello stato stazionario ottenuta nelle prime esecuzioni sperimentali come ampiezza di riferimento.
- Impostare una soglia di rifiuto (una diminuzione dell'ampiezza di risposta del 5% per quanto riguarda l'ampiezza di riferimento). Rifiutare le esecuzioni sperimentali in cui l'ampiezza della risposta allo stato stazionario soddisfa il criterio di rifiuto.
- Terminare la sessione sperimentale dopo aver acquisito il numero di corse definito nella progettazione sperimentale.
- Pre-elaborare i dati EEG offline utilizzando le procedure EEG standard35 descritte nei passaggi successivi per le indicazioni del produttore.
- Rifare riferimento alla registrazione utilizzando un riferimento medio (media di tutti gli elettrodi di registrazione) o la media di un sottoinsieme di elettrodi. In alternativa, utilizzare un riferimento fisico (ad esempio, elettrodi esterni posizionati sul naso e lobi delle eateri descritti al punto 2.4).
- Convertire le coordinate dell'elettrodo nel sistema internazionale 10/20 se il sistema di coordinate radiali è stato utilizzato durante le acquisizioni EEG. Consultare il manuale del produttore per informazioni dettagliate sulla conversione.
- Filtro passa-banda il segnale EEG tra 0,5-300 Hz. Impostare un filtro tacca (centrato a 50 Hz o 60 Hz) se necessario.
- Campione verso il basso del segnale EEG per ridurre il tempo di esecuzione dell'algoritmo selezionato per la rimozione degli artefatti oculari (passaggio 3.8.5).
NOTA: Una frequenza di campionamento di 512 Hz è sufficiente per analizzare le oscillazioni cerebrali della frequenza al di sotto di 40 Hz35. - Rimuovere gli artefatti oculari.
NOTA: A tal fine, è possibile utilizzare diverse tecniche (vedere Urig-en e Garcia-Apirain35 per un'ampia revisione degli algoritmi di rimozione degli artefatti). Tra questi, l'analisi dei componenti indipendenti è una delle metodologie più estese ed è implementata sia nei software di analisi commerciali che liberi37,38,39. - Segmentare i dati EEG in epoche bloccate nel tempo per la stimolazione. Selezionare la lunghezza delle epoche in base allo scopo dell'esperimento.
NOTA: le epoche devono essere sufficientemente lunghe per consentire l'analisi della risposta allo stato fisso nel dominio di frequenza con una risoluzione spettrale adeguata. - Non eseguire algoritmi di rifiuto degli artefatti in questa fase per rilevare e rimuovere le epoche contenenti elementi.
NOTA: la rimozione dell'epoca in questa fase indurrà errori quando il set di dati è organizzato per eseguire la media per colonna delle epoche (passaggi 4.2 e 4.4). Gli algoritmi di rifiuto vengono implementati in una fase di elaborazione successiva (passaggio 4.1.4). - Eseguire la funzione DC-detrend per calcolare le tendenze DC nelle singole epoche EEG e correggerle.
- Eseguire la funzione di correzione della linea di base per correggere la linea di base della registrazione. Selezionare intervalli di tempo pre-stimolo più lunghi di 200 ms.
NOTA: la correzione prevista consiste nell'esivare in media i dati nell'intervallo di tempo selezionato. La media viene calcolata per ogni canale e sottratta da ogni punto dati in ogni epoca.
4. Calcolo delle ampiezza di risposta
- Immettere i parametri necessari per il calcolo delle risposte allo stato fisso (Figura 1A).
NOTA: il codice interno utilizzato per l'elaborazione dei dati è disponibile gratuitamente all'indirizzo- Immettere il numero di registrazioni (esecuzioni sperimentali) dell'esperimento.
- Immettere la lunghezza delle epoche per segmentare le singole registrazioni.
- Immettere la frequenza di campionamento dell'esperimento.
- Selezionare algoritmi di rifiuto degli artefatti per rilevare e rimuovere le epoche contenenti elementi. I criteri di selezione disponibili sono 1) gradiente (differenza assoluta tra due campioni consecutivi), 2) max-min (la differenza tra l'ampiezza massima e minima nell'epoca) e 3) ampiezza (massima assoluta e ampiezza minima).
- Eseguire il codice di elaborazione.
NOTA: i passaggi 4.2-4.7 vengono eseguiti automaticamente quando questa opzione è selezionata. Eseguire i passaggi manualmente, se appropriato.
- Ridisporre le epoche in una matrice di dati di n righe e m colonne, in cui n rappresenta il numero di registrazioni (esecuzioni sperimentali) e m il numero di epoche (Figura 1B).
- Pesare l'epoca per attenuare l'effetto del movimento e artefatti muscolari.
NOTA: le epoch EEG ponderate sono ottenute dividendo ogni campione di tensione per la varianza dell'epoca a cui appartengono, in modo che la varianza venga utilizzata come misura della variabilità dell'ampiezza e del fattore di ponderazione40. - Media per colonne del set di dati. A tal fine, la media del dominio temporale delle epoche corrispondenti alla stessa finestra temporale nelle diverse registrazioni.
NOTA: questo passaggio consente il calcolo dell'ampiezza dello stato costante nelle registrazioni con un rapporto segnale-rumore (SNR) notevolmente elevato. - Esportare le serie temporali risultanti dalla media per un'ulteriore analisi dell'evoluzione temporale dell'intrappolamento in software esterno.
- Calcolare l'ampiezza della risposta allo stato stazionario in ogni epoca risultante dalla media a colonne, utilizzando la trasformazione veloce di Fourier (FFT).
NOTA: la lunghezza FFT deve corrispondere alla lunghezza di un'epoca. L'implementazione di una tecnica di windowing non è obbligatoria. L'ampiezza della risposta allo stato stazionario è definita come l'ampiezza spettrale ottenuta alla frequenza della modulazione di ampiezza degli stimoli sensoriali.
- Calcolare l'ampiezza della risposta allo stato stazionario in ogni epoca risultante dalla media a colonne, utilizzando la trasformazione veloce di Fourier (FFT).
- Media vettoriale l'ampiezza di un numero ad hoc di contenitori FFT su ciascun lato della frequenza della risposta per calcolare il livello di rumore residuo (RNL). Il numero di bidoni FFT deve corrispondere a una banda di frequenza di circa 3 Hz, su ciascun lato della frequenza della risposta.
NOTA: L'elevata specificità delle risposte allo stato fisso rende l'ampiezza di risposta indipendente da tali oscillazioni di fondo con frequenze simili, che a sua volta si distribuisce uniformemente in una banda di frequenza relativamente stretta41 , 42 o più , 43. - Tracciare l'ampiezza della risposta allo stato costante e della RNL in funzione dell'indice di colonna (cioè il numero dell'epoca acquisita) per esplorare l'evoluzione della risposta allo stato costante durante l'intervallo di stimolazione.
Representative Results
SSVEP è stato suscitato da stimoli visivi continui di 40 s di lunghezza, dove l'intensità della luce è stata modulata da un'onda sinusoidale di 10 Hz (profondità di modulazione del 90%). Gli stimoli sono stati consegnati da quattro diodi a emissione di luce (LED) situati al centro di uno schermo nero 50 cm x 50 cm, come vertici di un quadrato di 5 cm x 5 cm. Quando il partecipante si sedeva a 70 cm dallo schermo, l'area del quadrato dei LED subisce un angolo visivo di circa 4 gradi. Lo schermo a LED è stato progettato utilizzando un sistema di sviluppo microcontrollore basato su USB e quattro LED bianchi super luminosi di 10 mm di diametro. La tecnica di modulazione della larghezza dell'impulso (PWM) è stata utilizzata per controllare l'alimentazione fornita ai LED. Questa tecnica controllava l'intensità dei LED ad una determinata frequenza e generava l'involucro sinusoidale finale. Una frequenza PWM di 40 kHz è stata utilizzata per evitare un effetto di sfarfallio percepibile.
Sono state ottenute trenta registrazioni, che sono state segmentate in epoche di 4 s. Pertanto, è stato ottenuto un set di dati composto da 10 colonne (numero di epoche EEG all'interno delle registrazioni) e 30 righe (numero di registrazioni, numero di esecuzioni sperimentali).
L'oscillazione neurale bloccata nel tempo alla stimolazione è diventata evidente quando è stata eseguita la media a colonne (Figura2). Significativamente, l'intervallo in cui viene generato SSVEP può essere osservato nelle tracce corrispondenti alla colonna 1. In quella colonna, 0,2 s di linea di base pre-stimolo vengono tracciati in aggiunta ai primi 0,8 s di rientro neurale. Pertanto, la procedura qui descritta consente la caratterizzazione di 1) la dinamica della risposta oscillatoria una volta che l'intrappolamento neurale è già stabilito e 2) l'impegno delle oscillazioni neurali. Una o più epoche registrate dopo la fine della stimolazione possono anche essere incluse nella matrice di dati per studiare l'estinzione della risposta allo stato costante dopo l'offset dello stimolo.
Durante la media a colonne delle epoche, l'ampiezza media dell'SSVEP (ampiezza spettrale a 10 Hz, calcolata applicando l'FFT) è diminuita durante la media delle prime epoche delle colonne e tendente a stabilizzarsi in seguito (Figura 3A). Questo risultato concorda con studi precedenti che analizzano l'evoluzione di ASSR durante la media di epoche acquisite in sequenza21,22,40,43,44. Il comportamento dell'ampiezza della risposta durante la media è di solito spiegato dal contributo relativamente elevato del rumore non mediato all'ampiezza di risposta calcolata nelle prime epoche, che viene attenuata quando la media viene eseguita13, 44 (di sistema) , 45 anni , 46 per quanto , 47. Degno di nota, la variabilità dell'ampiezza SSVEP è diminuita significativamente con l'avanzare della media.
Abbiamo anche analizzato la RNL delle misurazioni durante la media a colonne delle epoche (Figura 3B). La RNL è stata calcolata in una banda di frequenza stretta (3 Hz) su entrambi i lati della frequenza del SSVEP. Anche se questa procedura non è comune quando SSVEP vengono analizzati, vettore-media di un determinato numero di contenitori di frequenza intorno a quello dell'intrappolamento neurale è lo standard per la stima del RNL nelle misurazioni ASSR41,42, 43. Come previsto, la RNL è progressivamente diminuita con l'aumentare del numero di epoche medie e ha raggiunto il livello asintotico dopo circa 20 epoche. A differenza di quanto osservato quando è stata analizzata l'ampiezza SSVEP, la deviazione standard della RNL è rimasta relativamente costante con l'aumento del numero di epoche medie, il che suggerisce che le condizioni di registrazione erano stabili lungo la sessione sperimentale.
I risultati presentati in precedenza hanno determinato le variazioni nel rapporto tra segnale di picco e rumore (pSNR) delle misurazioni durante la media a colonne dell'epoca (Figura 3C). Questo termine è definito qui come il rapporto (in dB) tra l'ampiezza quadrata della risposta (SSVEP) e l'ampiezza quadrata della RNL. Con l'avanzare della media, il pSNR è aumentato con l'aumentare del numero di epoche medie fino a 18, approssimativamente. Ulteriori incrementi del numero di epoche medie non hanno influito in modo significativo sulla qualità del segnale. La variabilità del pSNR è diminuita con la media di un maggior numero di epoche.
Infine, le dinamiche dell'ampiezza SSVEP e della RNL sono rappresentate nella Figura 4. Queste evoluzioni temporali sono state ottenute tracciando i parametri di risposta calcolati alla fine della media a colonne delle epoche in funzione del numero di colonne (in funzione del tempo). Come dimostrato da Labecki et al.26, la dinamica di SSVEP può variare significativamente tra i soggetti. Poiché i risultati presentati nella Figura 4 corrispondono a un singolo individuo, non è possibile fare generalizzazioni. In questo argomento, l'ampiezza di SSVEP visualizzato un comportamento relativamente complesso (Figura 4A). L'ampiezza di risposta è gradualmente aumentata durante i primi 12 secondi dopo l'inizio dello stimolo (tempo che corrisponde alla lunghezza di 3 epoche). Man mano che lo stimolo persisteva, l'SSVEP è diminuito costantemente durante i successivi 12 secondi e successivamente è rimasto relativamente costante. Questi risultati non possono essere spiegati dal comportamento della RNL, poiché questo parametro era relativamente costante durante l'intervallo di stimolazione (Figura 4B). L'aumento dell'ampiezza SSVEP dopo l'insorgenza dello stimolo è evidente nelle tracce presentate nella Figura 2 e può essere spiegato dai processi di integrazione, che comportano la stabilizzazione dell'insorgenza neurale. La successiva diminuzione dell'ampiezza suggerisce l'adattamento di SSVEP alla stimolazione sostenuta. Tuttavia, queste ipotesi devono essere testate in esperimenti controllati con dimensioni del campione appropriate.
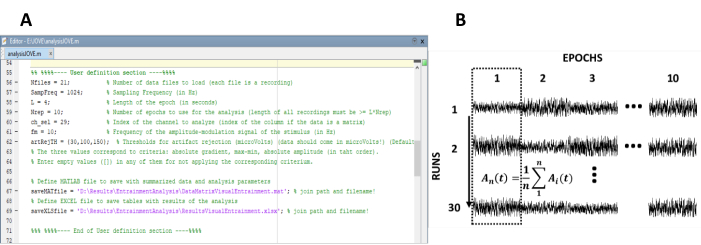
Figura 1 : passaggi critici per l'estrazione dell'evoluzione temporale dell'ampiezza delle risposte allo stato costante. (A) Screenshot del codice di elaborazione, in cui sono definiti i parametri di analisi. (B) Diagramma rappresentativo che illustra l'organizzazione del set di dati. Viene rappresentata una matrice di dati composta da 30 registrazioni di 10 epoche. La media per colonna delle epoche è evidenziata nella prima colonna. La linea verticale rappresenta la direzione della media. Fare clic qui per visualizzare una versione più grande di questa figura.

Figura 2 : Cambiamenti nella forma d'onda di potenziali visivamente evocati dallo stato fisso (SSVEP) durante la media a colonne delle epoche. Le risposte sono state suscitate dalla continua presentazione della luce modulata in ampiezza a 10 Hz. Le righe mostrano le forme d'onda ottenute dopo aver in media tutte le registrazioni precedenti (cioè, la riga 1 è la prima registrazione, la riga 5 è la forma d'onda ottenuta dopo la media delle prime cinque registrazioni e l'ultima riga è la media di tutte le registrazioni). Sono state osservate forme d'onda più affidabili di SSVEP in ogni colonna con l'aumento del numero di corse in media. Per fornire chiarezza (per rendere visibili le oscillazioni di SSVEP), è rappresentato solo il primo secondo delle epoche. Le eccezioni sono tracce nella prima colonna del set di dati, per le quali vengono visualizzati 0,2 secondi di linea di base pre-stimolo. Fare clic qui per visualizzare una versione più grande di questa figura.

Figura 3 : modifiche nei parametri di risposta e registrazione durante la media a colonne delle epoche. (A) Evoluzione dell'ampiezza SSVEP. (B) Comportamento della RNL. (C) Modifiche nel pSNR. Le linee nere rappresentano i valori medi ottenuti per ogni colonna (n - 10) e l'ombra grigia rappresenta l'area coperta da una deviazione standard. Fare clic qui per visualizzare una versione più grande di questa figura.

Figura 4 : Evoluzione temporale dell'SSVEP suscitata dalla presentazione della stimolazione visiva continua, modulata in ampiezza a 10 Hz. (A) Corso temporale dell'ampiezza SSVEP. (B) Corso temporale della RNL. Fare clic qui per visualizzare una versione più grande di questa figura.
Discussion
Questo lavoro descrive una procedura sperimentale per analizzare la dinamica delle risposte cerebrali oscillatorie. Tale metodologia consiste nell'acquisire un numero sufficiente di esecuzioni sperimentali indipendenti della stessa condizione sperimentale e di epoche di media del settore temporale corrispondenti allo stesso intervallo temporale nelle diverse registrazioni (colonne-wise in media in Figura 1B). L'ampiezza calcolata nei dati medi rappresenta l'ampiezza istantanea della risposta oscillatoria. Tracciare queste ampiezza in funzione del tempo (o del numero di colonne nel set di dati) consente di analizzare l'evoluzione temporale della risposta oscillatoria bloccata dalla stimolazione. Questa metodologia è una modifica di quella proposta da Ritter et al.23 per analizzare l'adattamento dei potenziali corticali transitori evocati. Il metodo è stato utilizzato per analizzare la dinamica dei potenziali uditivi evocati sia in casi umani24 che in modelli animali20,21.
Da un punto di vista metodologico, la combinazione di parametri utilizzati per suscitare la risposta allo stato costante e quelli implementati per estrarre la risposta neurale dal rumore di fondo è fondamentale per analizzare l'evoluzione del tempo dei potenziali evocati allo stato costante 22. La lunghezza dello stimolo utilizzata nell'esperimento qui presentato (40 s) è stata selezionata sulla base dei risultati ottenuti in uno studio pilota. Questa lunghezza di stimolo è stata sufficiente per analizzare l'adattamento dell'ASSR generato nel tronco encefalico del ratto21,22. Inoltre, la lunghezza dello stimolo dovrebbe superare il tempo in cui viene raggiunto il potere anotoso della banda istantanea degli SSVIP (Figura 1 in Labecki et al.26). Tuttavia, il potere anodo anotico della banda istantanea degli SSVIP può essere raggiunto oltre gli anni '60 in alcuni casi (Figura 2 in Labecki et al.26). Pertanto, si raccomanda di eseguire uno studio pilota di piccoli campioni per definire la lunghezza dello stimolo della stimolazione. In caso contrario, si raccomanda una lunghezza di stimolo più lunga di 90 s per ottenere una rappresentazione completa dell'evoluzione temporale della risposta. L'utilizzo di pause adeguatamente lunghe tra registrazioni consecutive implica considerare le esecuzioni sperimentali consecutive come statisticamente indipendenti (cioè diverse misure indipendenti della stessa variabile). Per quanto ne sappiamo, non sono stati effettuati esperimenti per analizzare la pausa ottimale tra le esecuzioni (pausa minima necessaria per effettuare esecuzioni indipendenti l'una dall'altra). Il criterio dell'uso delle pause di almeno 3 volte superiore alla lunghezza dello stimolo è sufficientemente conservativo da garantire che la risposta allo stato costante registrata in una determinata esecuzione non sia influenzata dalla stimolazione precedente.
Recentemente, stimoli alternati (condizioni sperimentali) è stato proposto come una scelta per ridurre la pausa tra le esecuzioni sperimentali, evitando un effetto di adattamento supplementare25. Allo stesso modo, il numero di esecuzioni sperimentali (30) implementate in questo protocollo sperimentale è conservativo, poiché la RNL asintotica e il pSNR sono in genere raggiunti dopo una media di 20 esecuzioni sperimentali, approssimativamente. Quando gli stimoli rientrano nella regione medio-alta della gamma dinamica della risposta (alti livelli di sensazione), è probabile che siano necessari un numero inferiore di corse per analizzare le dinamiche della risposta evocata. Tuttavia, nei casi in cui vengono testate diverse condizioni sperimentali, avere lo stesso numero di esecuzioni sperimentali è fondamentale per fare confronti tra le condizioni (cioè diversi livelli di sensazione).
Oltre alla media a colonne delle epoche, la dinamica dei potenziali evocati oscillatori è stata analizzata filtrando le misurazioni a prova singola in una banda di frequenza stretta intorno alla frequenza di interesse e calcolando l'involucro della potenza segnale utilizzando il filtro passa-basso26. Allo stesso modo, è stata attuata un'analisi di prova singola per caratterizzare il periodo di transizione che precede la regione stabile di SSVEP48e i cambiamenti di ampiezza e fase dell'SSVEP durante la regione stabile della risposta49. Mentre le singole analisi di prova consentono la discriminazione delle fluttuazioni relativamente rapide dell'ampiezza di risposta, i progetti sperimentali per analizzare la risposta media in blocchi separati con un determinato intervallo interblocco rappresentano solo variazioni a lungo termine ampiezza del potenziale evocato50,51. La media a colonne delle epoche si distingue tra queste due opzioni. La conversione del segnale medio al dominio di frequenza utilizzando il FFT implica analizzare la dinamica della risposta con una risoluzione uguale alla lunghezza dell'epoca. Nell'esempio qui presentato, l'SSVEP è stato segnalato ogni 4 s. Sebbene 4 s di risoluzione sia sufficiente a descrivere dinamiche che si verificano a intervalli di tempo superiori a decine di secondi, come quella dell'SSVEP26, le epoche parzialmente sovrapposte nelle registrazioni originali permette di descrivere l'evoluzione temporale del risposta allo stato costante in modo più raffinato25.
Le dinamiche delle risposte allo stato stazionario ottenute dopo la media a colonne delle epoche rappresentano principalmente l'evoluzione dell'attività oscillatoria sincronizzata tra i segmenti EEG mediati (quelli che sopravvivono alla media). Pertanto, un problema importante per quanto riguarda la fattibilità della metodologia è la possibile attenuazione delle ampiezza di risposta a causa delle variazioni nella fase delle oscillazioni neurali da una corsa sperimentale indipendente a un'altra (cioè tra le registrazioni). Questo argomento deve essere affrontato sperimentalmente. Tuttavia, le prove indicano che la fase delle risposte oscillatorie cerebrali è meno variabile del previsto. Infatti, diversi studi hanno segnalato una regolarità nella fase prevista dell'ASSR umano 80 Hz47,48,49. Quando le latenze sono stimate in base alla fase dell'attività oscillatoria, è stato osservato l'effetto prevedibile dell'intensità e la frequenza portante degli stimoli acustici sulla latenza delle risposte uditive (cioè la diminuzione della latenza l'intensità e l'aumento della frequenza dei portini)52,53,54. Inoltre, sono stati osservati cambiamenti maturazionali tipici dell'ampiezza e dell'asimmetria da sinistra a destra nei livelli uditivi, quando le latenze sono stimate dalla fase dell'ASSR47,55,56 , 57 del sistema , 58. Nel descrivere l'evoluzione temporale di SSVEP mediante analisi a prova singola, Labecki et al.26 ha osservato che, sebbene la variabilità inter-prova delle ampiezza di risposta all'interno dello stesso argomento fosse considerevolmente elevata, la variabilità del fase è stata significativamente meno pronunciata.
Sulla base delle loro osservazioni, Labecki et al.26 suggerirono che si dovrebbe calcolare una media di un minimo di 50 prove per ottenere una stima affidabile dell'involucro di potenza media della risposta. Questi risultati indicano che, anche quando l'ampiezza della risposta viene calcolata in singole prove, è necessaria una media (di buste in quel caso) per riportare risultati affidabili. Inoltre, la variabilità inter-prova nell'ampiezza di SSVEP riportata da Labecki et al.26 suggerisce che il calcolo di questo parametro in singole prove può essere fortemente influenzato dal rumore di fondo. Considerando l'evoluzione del rapporto segnale-rumore presentato nella Figura 2, il calcolo della risposta nel segnale medio invece di singole prove riduce significativamente il numero di segmenti EEG necessari per ottenere affidabili Misure. Inoltre, la bassa variabilità in fase ottenuta da Labecki et al.26 sostiene l'idea che la media a colonne delle epoche qui presentata sia una valida procedura per calcolare le dinamiche dei potenziali evocati oscillatori.
La media dei dati a diversi livelli porta a un'interpretazione diversa dei risultati. Per quanto riguarda i potenziali evocati oscillatori, calcolare l'ampiezza di risposta dopo la media del dominio temporale di esecuzioni indipendenti implica l'analisi solo delle oscillazioni bloccate nel tempo (cioè quelle che sopravvivono alla media). Questa procedura può filtrare le informazioni rilevanti relative alle dinamiche della risposta nelle singole prove. Tuttavia, consente un rapporto segnale-rumore sufficientemente elevato delle misurazioni. Questo aspetto potrebbe essere significativo quando le risposte sono vicine alla soglia elettrofisiologica, una condizione in cui il rilevamento dell'ingresso può essere compromesso a causa del basso rapporto segnale-rumore della misurazione.
Disclosures
Gli autori non hanno nulla da rivelare.
Acknowledgments
Gli autori riconoscono con gratitudine Lucàa Epeda, Grace A. Whitaker e Nicolas Nieto per il loro contributo alla produzione video. Questo lavoro è stato sostenuto in parte dai programmi CONICYT BASAL FB0008, MEC 80170124 e dalla borsa di dottorato 21171741, nonché dal National Institute on Deafness and Other Communication Disorders dei National Institutes of Health con il numero di premio P50DC015446. Il contenuto è di esclusiva responsabilità degli autori e non rappresenta necessariamente le opinioni ufficiali dei National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
References
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G. Jr, Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. An introduction to the event-related potential technique. Second Edition. , MIT Press. 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J. Jr, Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
