

Summary
Un protocole pour évaluer l'évolution temporelle de l'entraînement neuronal aux stimuli répétitifs externes est présenté. Les enregistrements à état stable de la même condition expérimentale sont acquis et moyens dans le domaine temporel. La dynamique de l'état stable est analysée en traçant l'amplitude de la réponse en fonction du temps.
Abstract
L'entraînement neuronal se réfère à la synchronisation de l'activité neuronale à la périodicité des stimuli sensoriels. Cette synchronisation définit la génération de réponses évoquées à l'état stable (c.-à-d. oscillations dans l'électroencéphalogramme enphase verrouillée aux stimuli de conduite). L'interprétation classique de l'amplitude des réponses évoquées à l'état stable suppose une réponse neuronale stéréotypée invariante et des fluctuations aléatoires de fond, de sorte que la moyenne sur les présentations répétées du stimulus récupère le réponse stéréotypée. Cette approche ignore la dynamique de l'état stable, comme dans le cas de l'adaptation suscitée par des expositions prolongées au stimulus. Pour analyser la dynamique des réponses à l'état stable, on peut supposer que l'évolution temporelle de l'amplitude de réponse est la même dans différentes courses de stimulation séparées par des pauses suffisamment longues. Sur la base de cette hypothèse, une méthode pour caractériser l'évolution temporelle des réponses à l'état stable est présentée. Un nombre suffisamment élevé d'enregistrements sont acquis en réponse à la même condition expérimentale. Les exécutions expérimentales (enregistrements) sont moyennes en colonne (c.-à-d. que les exécutions sont moyennes, mais l'époque dans les enregistrements ne sont pas moyennes avec les segments précédents). La moyenne de colonne-sage permet l'analyse des réponses à l'état régulier dans les enregistrements avec des rapports signal-bruit remarquablement élevés. Par conséquent, le signal moyen fournit une représentation précise de l'évolution temporelle de la réponse à l'état stable, qui peut être analysée dans les domaines du temps et de la fréquence. Dans cette étude, une description détaillée de la méthode est fournie, en utilisant des potentiels visuellement évoqués à l'état stable comme exemple de réponse. Les avantages et les mises en garde sont évalués sur la base d'une comparaison avec des méthodes à essai unique conçues pour analyser l'entraînement neuronal.
Introduction
Lorsqu'il est enregistré à partir du cuir chevelu, l'activité électrique du cerveau est observée comme des changements continus et réguliers dans les tensions au fil du temps. Cette activité électrique est appelée électroencéphalogramme (EEG) et a été décrite pour la première fois par Hans Berger à la fin des années vingt du siècle dernier1. Des études séminales ultérieures ont décrit l'EEG comme une série chronologique composée, dans laquelle différents modèles rythmiques ou répétitifs peuvent être observés2,3,4. De nos jours, l'EEG est généralement divisé en cinq bandes de fréquence bien établies, delta, theta, alpha, bêta et gamma, qui sont associées aux différents processus sensoriels et cognitifs.
Pendant des années, l'étude des oscillations cérébrales à l'aide de l'EEG s'est limitée à l'analyse du spectre dans l'activité continue ou aux changements de l'activité oscillatoire suscités par des événements sensoriels non périodiques. Au cours des dernières décennies, différentes méthodologies ont été mises en œuvre pour moduler les oscillations d'EEG en cours et explorer les effets de ces modulations sur les processus perceptuels et cognitifs, y compris la présentation de la stimulation sensorielle rythmique pour induisant l'entraînement neuronal. Le terme entraînement neuronal se réfère à la synchronisation de l'activité neuronale avec les propriétés périodiques des stimuli sensoriels. Ce processus conduit à la génération de potentiels évoqués à l'état stable (c.-à-d., oscillations EEG verrouillées aux propriétés périodiques des stimuli de conduite). Les potentiels évoqués à l'état stable sont le plus souvent obtenus par stimulation visuelle, auditive et vibrotactile, en utilisant des stimuli transitoires présentés à un rythme constant ou une stimulation continue modulée en amplitude à la fréquence des intérêts. Considérant que les potentiels évoqués à l'état stable somatosensoriel (SSSEPs) sont enregistrés en réponse à la stimulation tactile répétitive5,6, les potentiels évoqués visuellement à l'état stable (SSVEP) sont généralement obtenus par les potentiels périodiques présentation de scintillements de luminance, des images, et des visages7,8. Les réponses auditifs à l'état stable (ASSR) sont généralement générées par des trains de stimuli acoustiques transitoires ou par la présentation continue de tons modulés par amplitude9,10.
L'extraction de potentiels évoqués à l'état stable à partir de l'EEG mesuré repose essentiellement sur la moyenne des époques d'EEG acquises par la suite à l'époque du stimulus11. En raison de la périodicité des réponses, elles peuvent être analysées dans les domaines du temps et de la fréquence. Après la transformation de la fréquence-domaine, la réponse sensorielle est observée comme des pics d'amplitude au taux de présentation ou de la fréquence de modulation des stimuli externes, et de leurs harmoniques correspondantes. Ces procédures (moyenne du domaine temporel et transformation subséquente du domaine de fréquence) ont été essentielles pour développer un test auditif basé sur la détection des méthodes ASSR à des fins cliniques12,13,14 ,15,16.
En outre, la moyenne classique de domaine temporel des époques d'EEG a été extrêmement utile pour analyser des processus physiologiques tels que la génération et l'extinction de SSVEP17,18. Présentant des trains consécutifs de feux clignotants et une moyenne d'époques subséquentes dans un enregistrement, Wacker et coll.19 ont observé que l'indice de verrouillage de phase du SSVEP a rapidement augmenté au cours des 400 premiers m s de stimulation et est demeuré élevé par la suite. . Ils ont également signalé que l'entraînement visuel robuste a été établi entre 700-1 100 ms après le début de stimulus. Un certain degré d'entraînement est resté efficace après le décalage du train de stimulation, qui a duré environ trois périodes de la réponse oscillatoire17,19. Ces comportements ont été interprétés comme l'effet engageant/désinvoltant des oscillations observées, qui est une conséquence du traitement de l'information non linéaire dans le système visuel humain17. Alternativement, il est connu que dans certaines conditions expérimentales, la stimulation clignotante peut susciter des réactions au début, et hors-réponse à la fin des trains de stimulation au lieu de l'entraînement neuronal18.
L'hypothèse principale à la moyenne consécutivement acquis époques EEG est que le signal EEG représente une combinaison linéaire de la réponse sensorielle et le bruit de fond20. En outre, l'amplitude, la fréquence et la phase de la réponse oscillatoire sont supposées être stationnaires, tandis que le bruit de fond est considéré comme une activité aléatoire. Cependant, dans les cas où cette hypothèse n'est pas satisfaite, l'amplitude de réponse calculée après plusieurs époques ne correspond pas nécessairement à l'amplitude instantanée du potentiel évoqué.
Il a été récemment rapporté que l'ASSR généré dans le tronc cérébral des rats s'adapte à la présentation continue des tons amplitudes-modulés (c.-à-d., l'amplitude de réponse diminue exponentiellement au fil du temps)21,22. L'adaptation a été interprétée comme un mécanisme neuronal qui reflète la perte de nouveauté d'un stimulus sensoriel monotone répétitif, augmentant la sensibilité aux fluctuations pertinentes dans l'environnement acoustique23,24. Dans la voie auditive, l'adaptation peut améliorer la compréhension de la parole dans les environnements bruyants. En outre, ce processus peut faire partie des mécanismes existants pour surveiller la rétroaction auditive de sa propre voix pour contrôler la production de la parole.
En analysant l'évolution temporelle de l'ASSR 40 Hz chez l'homme, Van Eeckhoutte et coll.25 ont observé une diminution significative mais faible de l'amplitude de réponse au fil du temps (autour de -0,0002 V/s sur la base de l'analyse de groupe, en supposant une diminution linéaire au fil du temps). Par conséquent, ces auteurs ont conclu que l'ASSR 40 Hz chez l'homme ne s'adapte pas à la stimulation. Chez l'homme, des comportements non stationnaires ont été observés lors de l'analyse de la stabilité du SSVEP26. Ces auteurs ont observé que l'amplitude de la fréquence fondamentale et la deuxième harmonique du SSVEP étaient stationnaires dans seulement 30% et 66.7% des sujets qu'ils ont examinés, respectivement. Les phases des deux composants de fréquence SSVEP, bien que relativement stables au fil du temps, présentaient de petites dérives26.
Par conséquent, bien que la moyenne classique du domaine temporel des époques acquises ultérieurement permetd explore les propriétés stationnaires de l'entraînement neuronal, cette méthodologie doit être révisée lorsque la dynamique à long terme de l'entraînement est au centre de la recherche, ou lorsque la moyenne de la dynamique à court terme est corrompue par l'apparition d'une dynamique à long terme. Pour caractériser les comportements non stationnaires des réponses à état stable, la réponse évoquée calculée à une fenêtre de temps donnée ne devrait pas être compromise par ceux calculés dans les segments EEG précédents. En d'autres termes, le potentiel évoqué doit être extrait du bruit de fond sans que les époques soient moyennes temporelles avec les segments EEG précédents.
Dans cette étude, une méthode pour évaluer la dynamique de l'entraînement neuronal est présentée. Les réponses à l'état régulier sont enregistrées de façon répétitive en réponse à la même stimulation, où les enregistrements consécutifs sont entrelacés par un intervalle de repos de trois fois la durée de la course expérimentale. Considérant que si l'évolution temporelle de la réponse physiologique est la même dans différentes séries expérimentales indépendantes (enregistrements indépendants), les enregistrements sont en moyenne en fonction des colonnes. En d'autres termes, les époques correspondant au même endroit dans les différents enregistrements sont moyennes, sans moyenne d'époques dans un enregistrement. Par conséquent, l'amplitude de réponse calculée à n'importe quel intervalle de stimulation correspondra à l'amplitude instantanée du potentiel évoqué. Les réponses sensorielles peuvent être analysées dans le domaine temporel ou transformées en domaine de fréquence, selon le but de l'expérience. Dans tous les cas, les amplitudes peuvent être tracées en fonction du temps pour analyser l'évolution du temps de la réponse à l'état stable. La génération et l'extinction des potentiels évoqués à l'état stable peuvent être évaluées en limitant l'analyse à la première et à la dernière époque des enregistrements.
La dynamique de l'entraînement neuronal peut être analysée à l'aide d'autres approches, telles que le filtrage à bande étroite des mesures à un seul essai autour de la fréquence d'intérêt et le calcul de l'enveloppe du signal de puissance à l'aide du filtrage à faible passage25 et le Hilbert transformation27. Par rapport à ces méthodologies, la moyenne des époques en fonction de la colonne permet de calculer des paramètres à l'état stable basés sur des signaux avec le rapport signal-bruit plus élevé (SNR). Récemment, le filtrage Kalman a émergé comme une technique prometteuse pour l'estimation des amplitudes ASSR 40-Hz28,29,30. La mise en œuvre du filtrage Kalman peut améliorer la détection des réponses à l'état stable plus près du seuil électrophysiologique et réduire le temps de l'essai auditif29. De plus, il n'est pas nécessaire de supposer des réponses fixes lorsqu'une approche de filtrage Kalman est utilisée pour estimer l'amplitude ASSR30. Néanmoins, une seule étude a analysé l'évolution du temps des ASSR à l'aide de Kalman filtrant25. La conclusion de l'étude est que l'amplitude ASSR de 40-Hz est stable au cours de l'intervalle de stimulation. Par conséquent, le filtrage Kalman doit être testé dans des conditions dans lesquelles l'ASRR n'est pas stationnaire.
Bien que longue, la méthode de moyenne de colonne-sage est modèle-libre et n'a pas besoin des valeurs d'initialisation et/ou des définitions de priori du comportement de bruit. En outre, puisqu'il n'implique pas de temps de convergence, la moyenne de colonne-sage peut fournir une représentation plus fiable du début de l'entraînement neuronal. Par conséquent, les résultats obtenus avec la méthode de moyenne de colonne-sage peut être considéré comme la vérité de sol pour analyser la dynamique de l'entraînement neuronal utilisant le filtrage de Kalman.
Cette description du protocole est basée sur un exemple de SSVEP. Cependant, il est important de noter que la méthode présentée ici est indépendante de la modalité, de sorte qu'elle peut également être utilisée pour analyser l'évolution temporelle du SSSEP et de l'ASSR.
Protocol
La présente étude a été réalisée sous l'approbation du Comité de recherche et d'éthique de l'Universidad de Valparaso, Chili (code d'évaluation CEC170-18), confirmé aux lignes directrices nationales pour la recherche sur des sujets humains.
1. Préparation
- Bienvenue sur le sujet.
- Expliquer les objectifs et la pertinence de l'étude. Fournir une description des détails techniques pertinents. Répondez à toutes les questions à fond.
- Mentionnez explicitement qu'il peut interrompre la session expérimentale à tout moment si désiré.
- Demandez au bénévole de lire le consentement éclairé au sujet et de signer le formulaire correspondant. Interrompre la séance expérimentale si le consentement éclairé n'est pas obtenu.
2. Préparation du sujet
- Demandez au sujet de s'asseoir dans une chaise de laboratoire dans une position confortable.
- Nettoyer le cuir chevelu avec de l'éthanol (une solution à 95%) pour enlever la couche de cellules mortes de la peau et le sébum qui la recouvrent. Cette étape est importante pour réduire l'entrave entre les électrodes et le cuir chevelu.
- Mesurer la circonférence de la tête à l'utilisation d'un ruban à mesurer pour définir la taille du bouchon d'électrode.
- Demandez au sujet de porter le bouchon d'électrode. Fournir les instructions pour un positionnement confortable mais correct du bouchon.
- Mesurer la distance entre la nasion (Nz, le point médian de la suture nasofrontal, qui peut être identifiée par la dépression entre les yeux et le haut du nez) et l'inion (Iz, la proéminence de l'os occipital) à l'aide d'un ruban à mesurer.
- Mesurez la distance entre les points pré-auriculaires gauche et droit (identifié comme la dépression juste avant l'oreillette des oreilles) à l'aide d'un ruban à mesurer.
- Corriger la position du bouchon d'électrode, de sorte que l'intersection entre les lignes imaginaires définies dans les étapes précédentes correspondent au sommet de la tête. Assurez-vous que le sujet est confortable après les ajustements.
- Mettez le gel conducteur dans les supports d'électrode, selon les emplacements considérés pour l'expérience.
- Utilisez 64 emplacements de cuir chevelu suivant le système international 10-2031 pour utiliser les résultats du protocole pour effectuer l'analyse de localisation des sources. Le nombre plus élevé d'électrodes (128) sur le cuir chevelu peut être utilisé si nécessaire.
- Mettre en œuvre des paramètres cliniques ou ambulatoires (avec seulement quelques électrodes) si l'analyse de localisation des sources n'est pas planifiée. Utilisez des emplacements occipitaux pour enregistrer SSVEP, des emplacements temporels pour acquérir asSR, et des emplacements pariétals pour enregistrer SSSEP.
- Poussez les électrodes dans les supports d'électrodes. Assurez-vous que l'étiquette de l'électrode correspond à l'étiquette de l'emplacement dans le bouchon.
- Accompagner le volontaire à la salle expérimentale (de préférence, une chambre blindée et atténuée par le son). Demandez au sujet de s'asseoir dans une chaise à l'intérieur de la pièce, dans une position confortable.
- Placez des électrodes externes sur le nez et les lobes d'oreilles si une référence physique (différente des électrodes du cuir chevelu) sera utilisée pour ré-référencer l'enregistrement de l'EEG (à l'étape 3.8.1).
- Placer les électrodes externes dans des endroits périoculaires.
- Placez des électrodes sur la joue et la région frontale de la tête, à environ 1 cm au-dessus du sourcil, pour enregistrer le clignotement (à l'étape 2.6.1).
- Placez des électrodes sur le canthus externe des yeux, à environ 1 cm au-dessus/au-dessous de la ligne médiane, pour enregistrer les mouvements oculaires (à l'étape 2.6.1).
REMARQUE : L'électrooculogramme (EOG) sera utilisé à l'étape 3.8.5 pour enlever les artefacts EGG induits par les clignotements et les mouvements oculaires.
- Activez le système d'acquisition d'EEG et vérifiez l'entrave à l'électrode si un système de faible impedance est utilisé pour l'enregistrement de l'EEG. Corriger l'impédance, au besoin, selon les directives du fabricant. L'impédance doit être maintenue en dessous de 10k'32.
- Demandez au sujet de cligner des yeux et déplacez les yeux dans différentes directions pour vous assurer que l'EOG est correctement enregistré.
- Pour analyser la dynamique de SSVEP, ajustez l'emplacement de l'écran dans la direction verticale, pour correspondre à l'angle de vue du sujet. Dim les lumières de la salle jusqu'à ce qu'un niveau confortable est atteint. Ajustez le niveau de luminance de l'écran à la limite supérieure du niveau de confort du participant.
- Pour analyser la dynamique de l'ASSR, insérez les écouteurs à l'aide des inserts de mousse corrects, de sorte que les écouteurs s'adaptent au conduit auditif. Vérifier que les sons sont livrés à l'intensité désirée (p. ex., un niveau psychophysique confortable33).
3. Acquisition et prétraitement de l'EEG
- Définir les paramètres de stimulus définis dans la conception expérimentale. Consultez le manuel d'utilisation fourni par le fabricant du système de stimulation pour plus de détails sur le logiciel.
REMARQUE : Pour des explications complètes du stimulus utilisé pour la génération de SSVEP et asSR, voir Norcia et coll.8 et Rance34, respectivement. - Instruisez le sujet à prêter attention à la stimulation, dans le cas où l'entraînement visuel est le sujet de l'expérience.
- Présentez un film sous-titré avec le son éteint lorsque l'entraînement auditif est le sujet de l'expérience.
REMARQUE: La présentation d'un film muet permet de détourner l'attention de la stimulation acoustique tout en maintenant le niveau d'excitation25.
- Présentez un film sous-titré avec le son éteint lorsque l'entraînement auditif est le sujet de l'expérience.
- Stimuli actuels de plus de 90 s, comme cela a été fait pour étudier l'évolution du temps des SSVEPs et asSR chez les humains et les modèles animaux21,22,25,26.
REMARQUE : Présentez des stimuli de plus courte durée si une étude pilote a été effectuée. - Pause de la stimulation pendant 2 min si une seule condition expérimentale est testée. Interagissez avec le sujet pour vérifier la sensibilisation.
REMARQUE : La durée de la pause dépend de la durée de la stimulation. Pauses 3x plus longtemps que les intervalles de stimulation s'assurera qu'une réponse obtenue par un stimulus n'est pas affectée par la stimulation précédente. Des pauses plus longues sont autorisées si le sujet le demande.- Pause de la stimulation pour au moins 10 s lorsque différentes conditions expérimentales sont testées depuis la stimulation en alternance avec des pauses de 10 s a été proposé pour diminuer les effets d'adaptation supplémentaires et de réduire la longueur de l'expérience25.
- Répétez les étapes de présentation (étapes 3.3-3.4) au moins 30x pour assurer le SNR élevé des mesures après la moyenne des époques (étape 4.4).
- Enregistrez l'EEG en utilisant les procédures standard35. Créez un fichier EEG séparé pour chaque exécution expérimentale.
REMARQUE : Consultez le manuel d'utilisation du système d'acquisition pour plus de détails sur le logiciel. - Surveillez l'enregistrement de l'EEG pour détecter les périodes de sommeil en fonction du niveau d'activité alpha et de la fréquence à laquelle apparaissent les artefacts clignotants. Pause de l'expérience lorsque des niveaux alpha accrus accompagnés de fréquences clignotantes diminuées sont détectés, ce qui est indicatif de la somnolence. Rejetez la course expérimentale d'une analyse plus approfondie lorsque des périodes de sommeil sont détectées.
- Calculez l'amplitude de la réponse à l'état stable à la fin de chaque exécution expérimentale, suivant l'instruction fournie dans le manuel d'utilisation du logiciel d'acquisition utilisé dans l'expérience.
- Surveillez le niveau d'attention du sujet en comparant l'amplitude des réponses à l'état stable obtenues à la fin de chaque course expérimentale. Définir l'amplitude à l'état stable obtenue dans les premières courses expérimentales comme amplitude de référence.
- Fixer un seuil de rejet (diminution de l'amplitude de réponse de 5 % en ce qui concerne l'amplitude de référence). Rejetez les pistes expérimentales dans lesquelles l'amplitude de la réponse à l'état stable rencontre le critérium de rejet.
- Terminez la session expérimentale après l'acquisition du nombre de pistes définies dans la conception expérimentale.
- Prétraiter les données EEG hors ligne à l'aide des procédures EEG standard35 décrites dans les prochaines étapes selon les directives du fabricant.
- Re-re-référence l'enregistrement à l'aide d'une référence moyenne (moyenne de toutes les électrodes d'enregistrement) ou de la moyenne d'un sous-ensemble d'électrodes. Vous pouvez également utiliser une référence physique (p. ex., électrodes externes placées sur le nez et les lobes d'oreilles décrites à l'étape 2.4).
- Convertir les coordonnées d'électrodes au système international 10/20 si le système de coordonnées radiales a été utilisé lors des acquisitions d'EEG. Consultez le manuel du fabricant pour plus de détails sur la conversion.
- Filtre de bande-pass le signal d'EEG entre 0.5-300 Hz. Définir un encoche-filtre (centré à 50 Hz ou 60 Hz) si nécessaire.
- Échantillonversez le signal EEG pour réduire le temps d'exécution de l'algorithme sélectionné pour supprimer les artefacts oculaires (étape 3.8.5).
REMARQUE : Une fréquence d'échantillonnage de 512 Hz est suffisante pour analyser les oscillations cérébrales de la fréquence inférieure à 40 Hz35. - Retirez les artefacts oculaires.
REMARQUE : À cette fin, différentes techniques peuvent être utilisées (voir Urigen et Garcia-Zapirain35 pour un examen approfondi des algorithmes de suppression d'artefacts). Parmi eux, l'analyse indépendante des composants est l'une des méthodologies les plus étendues et est mise en œuvre dans les logiciels d'analyse commerciale et libre37,38,39. - Segmentez les données de l'EEG dans les époques verrouillées dans le temps jusqu'à la stimulation. Sélectionnez la longueur des époques en fonction du but de l'expérience.
REMARQUE : Les époques doivent être suffisamment longues pour permettre l'analyse de la réponse à l'état stable dans le domaine de fréquence avec une résolution spectrale adéquate. - N'exécutez pas d'algorithmes de rejet d'artefacts à ce stade pour détecter et supprimer les époques contenant des artefacts.
REMARQUE : La suppression de l'époque à ce stade induit des erreurs lorsque le jeu de données est organisé pour exécuter la moyenne des époques (étapes 4.2 et 4.4). Les algorithmes de rejet sont implémentés à une étape de traitement ultérieure (étape 4.1.4). - Exécutez la fonction DC-detrend pour calculer les tendances DC dans les différentes époques d'EEG et les corriger.
- Exécutez la fonction de correction de base pour corriger la ligne de base de l'enregistrement. Sélectionnez les intervalles de temps pré-stimulus de plus de 200 ms.
REMARQUE : La correction de base consiste à faire la moyenne des données dans l'intervalle de temps sélectionné. La moyenne est calculée pour chaque canal et soustraite de chaque point de données à chaque époque.
4. Calcul des amplitudes de réponse
- Entrez les paramètres nécessaires au calcul des réponses à l'état stable (figure 1A).
REMARQUE : Le code interne utilisé pour le traitement des données est disponible gratuitement à l'adresse suivante : https://figshare.com/projects/Steady-state-visually-evoked-potentials-SSVEP-elicited-in-humans-by-continuos-light-modulated-in-amplitude-at-10Hz/62573.. Consultez le texte d'aide à l'intérieur du code pour d'autres instructions. De même, un sous-ensemble des données utilisées dans cette étude est disponible.- Entrez le nombre d'enregistrements (pistes expérimentales) de l'expérience.
- Entrez la longueur des époques pour segmenter les enregistrements individuels.
- Entrez la fréquence d'échantillonnage de l'expérience.
- Sélectionnez des algorithmes de rejet d'artefacts pour détecter et supprimer les époques contenant des artefacts. Les critères de sélection disponibles sont 1) gradient (différence absolue entre deux échantillons consécutifs), 2) max-min (la différence entre l'amplitude maximale et l'amplitude minimale à l'époque) et 3) l'amplitude (amplitudes maximales et minimales absolues).
- Exécutez le code de traitement.
REMARQUE : Les étapes 4.2-4.7 sont automatiquement exécutées lorsque cette option est sélectionnée. Exécutez les étapes manuellement, le cas échéant.
- Réarrangez les époques en une matrice de données de n rows et m colonnes, dans laquelle n représente le nombre d'enregistrements (pistes expérimentales) et m le nombre d'époques (Figure 1B).
- Poids de l'époque pour atténuer l'effet du mouvement et des artefacts musculaires.
REMARQUE : Les époques pondérées d'EEG sont obtenues en divisant chaque échantillon de tension par la variance d'amplitude de l'époque à laquelle elles appartiennent, de sorte que la variance est utilisée comme mesure de la variabilité de l'amplitude et du facteur de pondération40. - Moyenne de colonnes de l'ensemble de données. À cette fin, la moyenne de domaine temporel correspond aux époques correspondant à la même fenêtre de temps dans les différents enregistrements.
REMARQUE : Cette étape permet le calcul de l'amplitude à l'état stable dans les enregistrements avec un rapport signal-bruit remarquablement élevé (SNR). - Exporter les séries chronologiques résultant de la moyenne pour une analyse plus approfondie de l'évolution du temps de l'entraînement dans les logiciels externes.
- Calculez l'amplitude de la réponse à l'état stable à chaque époque résultant de la moyenne de colonne-sage, en utilisant la transformation rapide de Fourier (FFT).
REMARQUE : La longueur de la FFT doit correspondre à la longueur d'une époque. La mise en œuvre d'une technique de fenémie n'est pas obligatoire. L'amplitude de la réponse à l'état stable est définie comme l'amplitude spectrale obtenue à la fréquence de la modulation de l'amplitude des stimuli sensoriels.
- Calculez l'amplitude de la réponse à l'état stable à chaque époque résultant de la moyenne de colonne-sage, en utilisant la transformation rapide de Fourier (FFT).
- La moyenne vectorielle de l'amplitude d'un nombre ad hoc de bacs FFT de chaque côté de la fréquence de la réponse pour calculer le niveau de bruit résiduel (RNL). Le nombre de bacs FFT doit correspondre à une bande de fréquence d'environ 3 Hz, de chaque côté de la fréquence de la réponse.
REMARQUE : La spécificité de haute fréquence des réponses à l'état stable rend l'amplitude de réponse indépendante de ces oscillations de fond avec des fréquences similaires, qui à son tour distribue uniformément dans une bande de fréquence relativement étroite41 , 42 Ans, états-unis ( , 43. - Tracer l'amplitude de la réponse à l'état stable et le RNL en fonction de l'index de colonne (c.-à-d., le nombre de l'époque acquise) pour explorer l'évolution de la réponse à l'état stable pendant l'intervalle de stimulation.
Representative Results
SSVEP a été obtenu par des stimuli visuels continus de 40 s de longueur, où l'intensité lumineuse a été modulée par une onde sinusoïdale de 10 Hz (profondeur de modulation de 90%). Les stimuli ont été délivrés par quatre diodes électroluminescentes (DEL) situées au centre d'un écran noir de 50 cm x 50 cm, sous forme de vertéxes d'un carré de 5 cm x 5 cm. Lorsque le participant s'est assis à 70 cm de l'écran, la zone du carré de LED sous-tend un angle visuel d'environ 4 degrés. L'écran LED a été conçu à l'aide d'un système de développement de microcontrôleur usb et de quatre LED blanches super brillantes de 10 mm de diamètre. La technique de modulation de la largeur d'impulsion (PWM) a été utilisée pour contrôler la puissance fournie aux LED. Cette technique a contrôlé les intensités de LED à une fréquence donnée et a généré l'enveloppe sinusoïdale finale. Une fréquence PWM de 40 kHz a été employée pour éviter un effet de scintillement perceptible.
Trente enregistrements ont été obtenus, qui ont été segmentés dans des époques de 4 s. Par conséquent, un jeu de données composé de 10 colonnes (nombre d'époques EEG dans les enregistrements) et 30 lignes (nombre d'enregistrements, nombre de pistes expérimentales) a été obtenu.
L'oscillation neuronale verrouillée par le temps de stimulation est devenue évidente au fur et à mesure que la moyenne de colonnes a été réalisée (Figure 2). De manière significative, l'intervalle auquel le SSVEP est généré peut être observé dans les traces correspondant à la colonne 1. Dans cette colonne, 0,2 s de ligne de base pré-stimulus sont tracés en plus des 0,8 s premiers de l'entraînement neuronal. Par conséquent, la procédure décrite ici permet la caractérisation de 1) la dynamique de la réponse oscillatoire une fois que l'entraînement neuronal est déjà établi et 2) l'engagement des oscillations neuronales. Une ou plusieurs époques enregistrées après la fin de la stimulation peuvent également être incluses dans la matrice de données pour étudier l'extinction de la réponse à l'état stable après le décalage de stimulus.
Au cours de la moyenne des époques, l'amplitude moyenne du SSVEP (amplitude spectrale à 10 Hz, calculée par l'application de la FFT) a diminué au cours de la moyenne des premières époques des colonnes et a eu tendance à se stabiliser par la suite (figure 3A). Ce résultat est d'accord avec les études précédentes analysant l'évolution de l'ASSR au cours de la moyenne des époques acquises séquentiellement21,22,40,43,44. Le comportement de l'amplitude de réponse pendant la moyenne s'explique habituellement par la contribution relativement élevée du bruit non moyenné à l'amplitude de réponse calculer dans les premières époques, qui est atténuée pendant que la moyenne est exécutée13, 44 Ans, en est à qui , 45 Annonces , 46 Annonces , 47. Il convient de noter que la variabilité de l'amplitude du SSVEP a considérablement diminué à mesure que la moyenne progressait.
Nous avons également analysé le RNL des mesures au cours de la moyenne des périodes (Figure 3B). Le RNL a été calculé dans une bande de fréquence étroite (3 Hz) des deux côtés de la fréquence du SSVEP. Bien que cette procédure ne soit pas commune lors de l'analyse du SSVEP, la moyenne vectorielle d'un nombre donné de bacs de fréquence autour de celui de l'entraînement neuronal est la norme pour estimer le RNL dans les mesures ASSR41,42, 43. Comme prévu, le RNL a progressivement diminué à mesure que le nombre d'époques moyennes augmentait et atteignait le niveau asymptotique après le traitement d'une vingtaine d'époques. Contrairement à celle observée lors de l'analyse de l'amplitude SSVEP, l'écart type du RNL est demeuré relativement constant à mesure que le nombre d'époques moyennes augmentait, ce qui suggère que les conditions d'enregistrement étaient stables tout au long de la session expérimentale.
Les résultats présentés ci-dessus ont déterminé les changements dans le rapport signal-bruit maximal (pSNR) des mesures au cours de la moyenne de l'époque (figure3C). Ce terme est défini ici comme le rapport (en dB) entre l'amplitude carrée de la réponse (SSVEP) et l'amplitude carrée de la RNL. Au fur et à mesure que la moyenne progressait, le pSNR augmentait à mesure que le nombre d'époques moyennes augmentait jusqu'à 18, environ. D'autres augmentations du nombre d'époques moyennes n'ont pas eu d'incidence significative sur la qualité du signal. La variabilité du pSNR a diminué à mesure que l'on assurait la moyenne d'un plus grand nombre d'époques.
Enfin, la dynamique de l'amplitude SSVEP et du RNL est représentée à la figure 4. Ces évolutions temporelles ont été obtenues en traçant les paramètres de réponse calculés à la fin de la moyenne des époques en fonction du nombre de colonnes (en fonction du temps). Comme l'ont démontré Labecki et coll.26, la dynamique du SSVEP peut varier considérablement d'un sujet à l'autre. Étant donné que les résultats présentés à la figure 4 correspondent à une seule personne, il est impossible de généraliser. Dans ce domaine, l'amplitude du SSVEP présentait un comportement relativement complexe (figure 4A). L'amplitude de réponse a graduellement augmenté pendant les 12 premières secondes suivant le début de stimulus (temps qui correspond à la longueur de 3 époques). Pendant que le stimulus persistait, le SSVEP a constamment diminué pendant les 12 secondes suivantes, et est resté relativement constant par la suite. Ces résultats ne peuvent pas être expliqués par le comportement de la RNL, puisque ce paramètre était relativement constant pendant l'intervalle de stimulation (Figure 4B). L'augmentation de l'amplitude SSVEP après l'initialisation du stimulus est évidente dans les traces présentées à la figure 2 et peut s'expliquer par des processus d'intégration, qui se traduisent par la stabilisation de l'entraînement neuronal. La diminution subséquente de l'amplitude suggère l'adaptation de SSVEP à la stimulation soutenue. Néanmoins, ces hypothèses doivent être testées dans le cas d'expériences contrôlées avec des tailles d'échantillons appropriées.
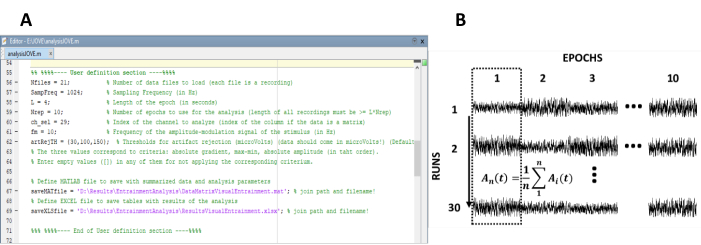
Figure 1 : Étapes critiques pour extraire l'évolution temporelle de l'amplitude des réponses à l'état stable. (A) Capture d'écran du code de traitement, où les paramètres d'analyse sont définis. (B) Diagramme représentatif illustrant l'organisation de l'ensemble de données. Une matrice de données composée de 30 enregistrements de 10 époques est représentée. La moyenne des époques dans le sens des colonnes est mise en évidence dans la première colonne. La ligne verticale représente la direction de la moyenne. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 2 : Changements dans la forme d'onde des potentiels visuellement évoqués à l'état stable (SSVEP) au cours de la moyenne des périodes. Les réponses ont été obtenues par la présentation continue de la lumière modulée dans l'amplitude à 10 Hz. Les lignes montrent les formes d'ondes obtenues après la moyenne de tous les enregistrements précédents (c.-à-d., la rangée 1 est le premier enregistrement, la rangée 5 est la forme d'onde obtenue après la moyenne des cinq premiers enregistrements, et la dernière rangée est la moyenne de tous les enregistrements). Des formes d'ondes plus fiables de SSVEP ont été observées dans chaque colonne à mesure que le nombre de courses moyennes augmentait. Pour apporter de la clarté (pour rendre visibles les oscillations du SSVEP), seule la première seconde des époques est représentée. Les exceptions sont des traces dans la première colonne de l'ensemble de données, pour lesquelles 0,2 seconde de base pré-stimulus sont affichées. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 3 : Modifications des paramètres de réponse et d'enregistrement au cours de la moyenne des époques. (A) Evolution de l'amplitude SSVEP. (B) Comportement de la RNL. (C) Changements dans le pSNR. Les lignes noires représentent les valeurs moyennes obtenues pour chaque colonne (n ' 10) et l'ombre grise représente la zone couverte par un écart type. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.

Figure 4 : Évolution temporelle du SSVEP obtenue par la présentation d'une stimulation visuelle continue, modulée en amplitude à 10 Hz. (A) Cours de temps de l'amplitude SSVEP. (B) Cours de temps de la RNL. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
Discussion
Ce travail décrit une procédure expérimentale pour analyser la dynamique des réponses cérébrales oscillatoires. Une telle méthodologie consiste à acquérir un nombre suffisant de séries expérimentales indépendantes de la même condition expérimentale, et des époques de moyenne de domaine temporel correspondant à la même fenêtre de temps dans les différents enregistrements (colonnes-sage moyenne dans Figure 1B). L'amplitude calculée dans les données moyennes représente l'amplitude instantanée de la réponse oscillatoire. Tracer ces amplitudes en fonction du temps (ou du nombre de colonnes dans l'ensemble de données) permet d'analyser l'évolution temporelle de la réponse oscillatoire en temps de jeu à la stimulation. Cette méthodologie est une modification de celle proposée par Ritter et coll.23 pour analyser l'adaptation des potentiels corticaux transitoires évoqués. La méthode a été utilisée pour analyser la dynamique des potentiels auditifs évoqués chez les humains24 et les modèles animaux20,21.
D'un point de vue méthodologique, la combinaison des paramètres utilisés pour obtenir la réponse à l'état stable et ceux mis en œuvre pour extraire la réponse neuronale du bruit de fond est essentiel pour analyser l'évolution temporelle des potentiels évoqués à l'état stable 22. La longueur de stimulation utilisée dans l'expérience présentée ici (40 s) a été choisie sur la base des résultats obtenus dans une étude pilote. Cette longueur de stimulus était suffisante pour analyser l'adaptation de l'ASSR générée dans le tronc cérébral de rat21,22. De plus, la longueur de stimulation devrait dépasser le moment où la puissance de bande instantanée asymptotique des SSVEPs est atteinte (figure1 dans Labecki et al.,26). Néanmoins, la puissance de bande instantanée asymptotique des SSVEPs peut être atteinte au-delà de la soixantaine dans certains cas (figure2 dans Labecki et al.,26). Par conséquent, il est recommandé d'exécuter une étude pilote à petit échantillon pour définir la longueur de stimulation de la stimulation. Dans le cas contraire, une longueur de stimulus supérieure à 90 s est recommandée pour obtenir une représentation complète de l'évolution temporelle de la réponse. L'utilisation de pauses suffisamment longues entre les enregistrements consécutifs implique d'envisager des exécutions expérimentales consécutives comme étant statistiquement indépendantes (c.-à-d., des mesures différentes et indépendantes d'une même variable). Au meilleur de notre connaissance, aucune expérience n'a été effectuée pour analyser la pause optimale entre les courses (pause minimale requise pour rendre les courses indépendantes les unes des autres). Le critérium d'utilisation des pauses au moins 3fois de plus que la longueur de stimulus est assez conservateur pour s'assurer que la réponse à l'état régulier enregistrée dans une course donnée n'est pas affectée par la stimulation précédente.
Récemment, des stimuli alternatifs (conditions expérimentales) ont été proposés comme un choix pour réduire la pause entre les pistes expérimentales, en évitant l'effet d'adaptation supplémentaire25. De même, le nombre de pistes expérimentales (30) mises en œuvre dans ce protocole expérimental est prudent, puisque le RNL asymptotique et le pSNR sont généralement atteints après une moyenne de 20 pistes expérimentales, environ. Lorsque les stimuli se situent dans la région moyenne-haute de la plage dynamique de la réponse (niveaux de sensation élevés), un nombre plus faible de pistes est probablement nécessaire pour analyser la dynamique de la réponse évoquée. Néanmoins, dans les cas où différentes conditions expérimentales sont testées, avoir le même nombre de pistes expérimentales est crucial pour faire des comparaisons entre les conditions (c.-à-d., différents niveaux de sensation).
En plus de la moyenne des époques, la dynamique des potentiels oscillatoires évoqués a été analysée en filtrant les mesures à essai unique dans une bande de fréquence étroite autour de la fréquence d'intérêt et en calculant l'enveloppe de la puissance signal à l'aide de filtrage à faible passage26. De même, une analyse d'essai unique a été mise en œuvre pour caractériser la période de transition qui précède la région stable du SSVEP48, et les changements dans l'amplitude et la phase du SSVEP au cours de la région stable de la réponse49. Bien que les analyses d'essais uniques permettent de dissocler les fluctuations relativement rapides de l'amplitude de réponse, les conceptions expérimentales pour analyser la réponse moyenne dans les blocs séparés avec un intervalle interbloc donné ne tiennent compte que des variations à long terme de la amplitude du potentiel évoqué50,51. La moyenne des époques se situe entre ces deux options. Convertir le signal moyen au domaine de fréquence à l'aide de la FFT implique d'analyser la dynamique de la réponse avec une résolution égale à la longueur de l'époque. Dans l'exemple présenté ici, le SSVEP a été signalé tous les 4 s. Bien que 4 s de résolution soit suffisante pour décrire la dynamique se produisant à intervalles de temps dépassant des dizaines de secondes, comme celle du SSVEP26, les époques partiellement qui se chevauchent dans les enregistrements originaux permettent de décrire l'évolution temporelle de la réponse à l'état stable d'une manière plus raffinée25.
La dynamique des réponses à l'état stable obtenues après une moyenne des époques selon les colonnes représente principalement l'évolution de l'activité oscillatoire synchronisée entre les segments d'EEG moyens (ceux qui survivent à la moyenne). Par conséquent, un problème majeur concernant la faisabilité de la méthodologie est l'atténuation possible des amplitudes de réponse en raison des variations dans la phase des oscillations neuronales d'une course expérimentale indépendante à l'autre (c.-à-d., parmi les enregistrements). Ce sujet doit être abordé expérimentalement. Cependant, les preuves indiquent que la phase des réponses oscillatoires de cerveau est moins variable que prévu. En fait, plusieurs études ont rapporté une régularité dans la phase prévue de l'homme 80 Hz ASSR47,48,49. Lorsque les latences sont estimées en fonction de la phase de l'activité oscillatoire, l'effet prévisible de l'intensité et de la fréquence des stimuli acoustiques sur la latence des réponses auditives a été observé (c.-à-d. la diminution de la latence à mesure que le augmentation de l'intensité et de la fréquence du transporteur)52,53,54. En outre, des changements de maturation typiques dans l'amplitude et l'asymétrie de gauche à droite dans les niveaux auditifs ont également été observés lorsque des latences sont estimées à partir de la phase de l'ASSR47,55,56 , 57 Annonces , 58. En décrivant l'évolution temporelle du SSVEP à l'aide de l'analyse à essai unique, Labecki et coll.26 ont observé que, bien que la variabilité inter-essais des amplitudes de réponse au sein d'un même sujet ait été considérablement élevée, la variabilité de la phase a été significativement moins prononcée.
Sur la base de leurs observations, Labecki et coll.26 ont suggéré qu'un minimum de 50 essais devraient être menés en moyenne pour obtenir une estimation fiable de l'enveloppe moyenne de puissance de la réponse. Ces résultats indiquent que, même lorsque l'amplitude de la réponse est computée dans des essais uniques, la moyenne (des enveloppes dans ce cas) est nécessaire pour rapporter des résultats dignes de confiance. De plus, la variabilité inter-essais dans l'amplitude du SSVEP rapportée par Labecki et coll.26 suggère que le calcul de ce paramètre dans des essais uniques peut être fortement influencé par le bruit de fond. Compte tenu de l'évolution du rapport signal-bruit présenté à la figure 2, le calcul de la réponse dans le signal moyen au lieu d'essais uniques réduit considérablement le nombre de segments d'EEG à traiter pour obtenir des Mesures. En outre, la faible variabilité de la phase obtenue par Labecki et coll.26 soutient l'idée que la moyenne des époques présentées ici est une procédure valable pour calculer la dynamique des potentiels oscillatoires évoqués.
La moyenne des données à différents niveaux conduit à une interprétation différente des résultats. En ce qui concerne les potentiels évoqués oscillatoires, calculer l'amplitude de réponse après la moyenne temporelle des exécutions indépendantes implique d'analyser uniquement les oscillations temporelles (c.-à-d., celles qui survivent à la moyenne). Cette procédure peut filtrer les informations pertinentes concernant la dynamique de la réponse dans les essais individuels. Cependant, il garantit un rapport signal-bruit suffisamment élevé des mesures. Cet aspect peut être important lorsque les réponses sont proches du seuil électrophysiologique, une condition dans laquelle la détection de l'entraînement peut être compromise en raison d'un faible rapport signal-bruit de la mesure.
Disclosures
Les auteurs n'ont rien à révéler.
Acknowledgments
Les auteurs remercient Luca Zepeda, Grace A. Whitaker et Nicolas Nieto pour leur contribution à la production vidéo. Ce travail a été soutenu en partie par les programmes CONICYT BASAL FB0008, MEC 80170124 et phD bourse 21171741, ainsi que l'Institut national sur la surdité et autres troubles de la communication des National Institutes of Health sous le numéro de prix P50DC015446. Le contenu est uniquement de la responsabilité des auteurs et ne représente pas nécessairement les vues officielles des National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Active electrodes | Biosemi | P32-1020-32ACMS (ABC) | for channels 1-32 |
| Active electrodes | Biosemi | P32-1020-32A (ABC) | for channels 33-64 |
| Active electrodes | Biosemi | 8 x TP FLAT | external electrodes |
| Active-Two adquisition system | Biosemi | version 7.0 | EEG adquisition system |
| alcohol | Salcobrand | Code: 3309011 | for cleaning the scalp |
| Electrode cap 64 channels | Biosemi | CAP MS xx yy | cap |
| Electrode cap 64 channels | Biosemi | CAP ML xx yy | cap |
| gel | Biosemi | SIGNA BOX12 | conductive gel |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for stimulation |
| Laptop | Asus | Core i7 1TB DD + 128GB SSD 8GB RAM | computer for recording |
| LED screen | in-house production | - | The screen consists of four light-emitting diodes (LEDs) situated on the center of a 50x50 cm black screen, as vertexes of a square of 5x5 cm |
| sterile gauze | Salcobrand | Code: 8730277 | for cleaning the scalp |
References
- Berger, H. Über das elektrenkephalogramm des menschen. European Archives of Psychiatry and Clinical Neuroscience. 87 (1), 527-570 (1929).
- Berger, H. Electroencephalogram of humans. Journal fur Psychologie und Neurologie. 40, 160-179 (1930).
- Walter, W. G. The location of cerebral tumours by electro-encephalography. The Lancet. 228 (5893), 305-308 (1936).
- Jasper, H. H., Andrews, H. L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Archives of Neurology & Psychiatry. 39 (1), 96-115 (1938).
- Snyder, A. Z. Steady-state vibration evoked potentials: descriptions of technique and characterization of responses. Electroencephalography and Clinical Neurophysiology. 84 (3), 257-268 (1992).
- Giabbiconi, C. M., Trujillo-Barreto, N. J., Gruber, T., Muller, M. M. Sustained spatial attention to vibration is mediated in primary somatosensory cortex. Neuroimage. 35, 255-262 (2007).
- Rossion, B., Boremanse, A. Robust sensitivity to facial identity in the right human occipito-temporal cortex as revealed by steady-state visual-evoked potentials. Journal of Vision. 11 (2), (2011).
- Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., Rossion, B. The steady-state visual evoked potential in vision research: a review. Journal of Vision. 15 (6), 4 (2015).
- Galambos, R., Makeig, S., Talmachoff, P. J. A 40-Hz auditory potential recorded from the human scalp. Proceedings of the National Academy of Sciences of the United States of America. 78 (4), 2643-2647 (1981).
- Picton, T. W., Skinner, C. R., Champagne, S. C., Kellett, A. J., Maiste, A. C. Potentials evoked by the sinusoidal modulation of the amplitude or frequency of a tone. The Journal of the Acoustical Society of America. 82 (1), 165-178 (1987).
- Dawson, G. D. A summation technique for the detection of small evoked potentials. Electroencephalography and Clinical Neurophysiology. 6 (1), 65-84 (1954).
- Savio, G., Cardenas, J., Abalo, M. P., Gonzalez, A., Valdes, J. The low and high frequency auditory steady state responses mature at different rates. Audiology and Neurotology. 6 (5), 279-287 (2001).
- Luts, H., Desloovere, C., Kumar, A., Vandermeersch, E., Wouters, J. Objective assessment of frequencyspecific hearing thresholds in babies. International Journal of Pediatric Otorhinolaryngology. 68, 915-926 (2004).
- Valdes, J. L., et al. Comparison of statistical indicators for the automatic detection of 80 Hz auditory steady state responses. Ear and Hearing. 18 (5), 420-429 (1997).
- Wilding, T., McKay, C., Baker, R., Kluk, K. Auditory steady state responses in normal-hearing and hearing-impaired adults: an analysis of between-session amplitude and latency repeatability, test time, and F ratio detection paradigms. Ear and Hearing. 33 (2), 267 (2012).
- de Resende, L. M., et al. Auditory steady-state responses in school-aged children: a pilot study. Journal of NeuroEngineering and Rehabilitation. 12 (1), 13 (2015).
- Halbleib, A., et al. Topographic analysis of engagement and disengagement of neural oscillators in photic driving: a combined electroencephalogram/magnetoencephalogram study. Journal of Clinical Neurophysiology. 29 (1), 33-41 (2012).
- Salchow, C., et al. Rod Driven Frequency Entrainment and Resonance Phenomena. Frontiers in Human Neuroscience. 10, 413 (2016).
- Wacker, M., et al. A time-variant processing approach for the analysis of alpha and gamma MEG oscillations during flicker stimulus generated entrainment. IEEE Transactions on Biomedical Engineering. 58 (11), 3069-3077 (2011).
- Glaser, E. M., Ruchkin, D. S. Principles of Neurobiological Signal Analysis. Journal of Clinical Engineering. 2 (4), 382-383 (1977).
- Prado-Gutierrez, P., et al. Habituation of auditory steady state responses evoked by amplitude-modulated acoustic signals in rats. Audiology Research. 5 (1), (2015).
- Prado-Gutierrez, P., Martínez-Montes, E., Weinstein, A., Zañartu, M. Estimation of auditory steady-state responses based on the averaging of independent EEG epochs. PLoS ONE. 14 (1), (2019).
- Ritter, W., Vaughan, H. G. Jr, Costa, L. D. Orienting and habituation to auditory stimuli: a study of short terms changes in average evoked responses. Electroencephalography and Clinical Neurophysiology. 25 (6), 550-556 (1968).
- Malmierca, M. S., Cristaudo, S., Pérez-González, D., Covey, E. Stimulus-specific adaptation in the inferior colliculus of the anesthetized rat. Journal of Neuroscience. 29 (17), 5483-5493 (2009).
- Van Eeckhoutte, M., Luke, R., Wouters, J., Francart, T. Stability of auditory steady state responses over time. Ear and Hearing. 39 (2), 260-268 (2018).
- Labecki, M., Nowicka, M. M., Suffczynski, P. Temporal Modulation of Steady-State Visual Evoked Potentials. International Journal of Neural Systems. 29 (3), 1850050 (2019).
- Zhang, S., et al. A study on dynamic model of steady-state visual evoked potentials. Journal of Neural Engineering. 15 (4), 046010 (2018).
- Wilson, U. S., Kaf, W. A., Danesh, A. A., Lichtenhan, J. T. Assessment of low-frequency hearing with narrowband chirp evoked 40-Hz sinusoidal auditory steady state response. International Journal of Audiology. 55 (4), 239-247 (2016).
- Kaf, W. A., Mohamed, E. S., Elshafiey, H. 40-Hz Sinusoidal Auditory Steady-State Response and Tone Burst Auditory Brainstem Response Using a Kalman Filter to Determine Thresholds Pre- and Post-Myringotomy with Grommet Tube in Children With Mild, Low-Frequency Conductive Hearing Loss. American Journal of Audiology. 25 (1), 41-53 (2016).
- Luke, R., Wouters, J. Kalman filter based estimation of auditory steady state response parameters. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 25, 196-204 (2017).
- Homan, R. W. The 10-20 Electrode System and Cerebral Location. American Journal of EEG Technology. 28 (4), 269-279 (1988).
- Kappenman, E. S., Steven, J. L. The Effects of Electrode Impedance on Data Quality and Statistical Significance in ERP Recordings. Psychophysiology. 47 (5), 888-904 (2010).
- Moore, R., Gordon-Hickey, S., Jones, A. Most comfortable listening levels, background noise levels, and acceptable noise levels for children and adults with normal hearing. Journal of the American Academy of Audiology. 22 (5), 286-293 (2011).
- Rance, G. The Auditory Steady-state Response: Generation, Recording, and Clinical Application. Plural Publishing. 335, (2008).
- Luck, S. J. An introduction to the event-related potential technique. Second Edition. , MIT Press. 406 (2014).
- Urigüen, J. A., Garcia-Zapirain, B. EEG artifact removal-state-of-the-art and guidelines. Journal of Neural Engineering. 12 (3), (2015).
- Jung, T. P., et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37 (2), 163-178 (2000).
- Li, Y., Ma, Z., Lu, W., Li, Y. Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiological Measurement. 27 (4), 425 (2006).
- Mannan, M. M. N., Jeong, M. Y., Kamran, M. A. Hybrid ICA-Regression: automatic identification and removal of ocular artifacts from electroencephalographic signals. Frontiers in Human Neuroscience. 10, 193 (2016).
- John, M. S., Dimitrijevic, A., Picton, T. W. Weighted averaging of steady-state responses. Clinical Neurophysiology. 112 (3), 555-562 (2001).
- Zurek, P. M. Detectability of transient and sinusoidal otoacoustic emissions. Ear and Hearing. 13 (5), 307-310 (1992).
- Lins, O. G., Picton, P. E., Picton, T. W., Champagne, S. C., Durieux-Smith, A. Auditory steady-state responses to tones amplitude-modulated at 80-110 Hz. Journal of the Acoustical Society of America. 97 (5), 3051-3063 (1995).
- John, M. S., Picton, T. W. Human auditory steady-state responses to amplitude-modulated tones: phase and latency measurements. Hearing Research. 141 (1-2), 57-79 (2000).
- Fortuny, A. T., et al. Criterio de parada de la promediación de la respuesta auditiva de estado estable. Acta Otorrinolaringológica Española. 62 (3), 173-254 (2011).
- John, M. S., Purcell, D. W., Dimitrijevic, A., Picton, T. W. Advantages and caveats when recording steady-state responses to multiple simultaneous stimuli. Journal of the American Academy of Audiology. 13 (5), 246-259 (2002).
- Luts, H., Van Dun, B., Alaerts, J., Wouters, J. The influence of the detection paradigm in recording auditory steady-state responses. Ear and Hearing. 29 (4), 638-650 (2008).
- Choi, J. M., Purcell, D. W., John, M. S. Phase stability of auditory steady state responses in newborn infants. Ear and Hearing. 32 (5), 593-604 (2011).
- Xu, M., et al. Use of a steady-state baseline to address evoked vs. oscillation models of visual evoked potential origin. Neuroimage. 134, 204-212 (2016).
- Peachey, N. S., Demarco, P. J. Jr, Ubilluz, R., Yee, W. Short-term changes in the response characteristics of the human visual evoked potential. Vision Research. 34 (21), 2823-2831 (1994).
- Woods, D. L., Elmasian, R. The habituation of event-related potentials to speech soundsand tones. Electroencephalography and Clinical Neurophysiology. 65, 447-459 (1986).
- Ravden, D., Polich, J. Habituation of P300 from visual stimuli. International Journal of Psychophysiology. 30 (3), 359-365 (1998).
- Picton, T. W., Dimitrijevic, A., John, M. S., Van Roon, P. The use of phase in the detection of auditory steady-state responses. Clinical Neurophysiology. 112 (9), 1698-1711 (2001).
- Alaerts, J., Luts, H., Van Dun, B., Desloovere, C., Wouters, J. Latencies of auditory steady-state responses recorded in early infancy. Audiology and Neurotology. 15 (2), 116-127 (2010).
- John, M. S., Brown, D. K., Muir, P. J., Picton, T. W. Recording auditory steady-state responses in young infants. Ear and Hearing. 25 (6), 539-553 (2004).
- Purcell, D. W., John, S. M., Schneider, B. A., Picton, T. W. Human temporal auditory acuity as assessed by envelope following responses. Journal of the Acoustical Society of America. 116 (6), 3581-3593 (2004).
- Kuwada, S., et al. Sources of the scalp-recorded amplitude-modulation following response. Journal of the American Academy of Audiology. 13 (4), 188-204 (2002).
- Pauli-Magnus, D., et al. Detection and differentiation of sensorineural hearing loss in mice using auditory steady-state responses and transient auditory brainstem responses. Neuroscience. 149 (3), 673-684 (2007).
- Prado-Gutierrez, P., et al. Maturational time course of the Envelope Following Response to amplitude-modulated acoustic signals in rats. International Journal of Audiology. 51 (4), 309-316 (2012).
