

Summary
ここでは、高精度かつ分類分解能で水生捕食真核生物の放牧率を定量化する単細胞、エピ蛍光顕微鏡ベースの技術のためのプロトコルです。
Abstract
捕食やその効果などの栄養相互作用を解明することは、生態学の多くの研究者にとって頻繁な作業です。微生物群相の研究には多くの制限があり、捕食者、獲物、捕食速度の決定はしばしば困難です。ここでは、蛍光標識された獲物をトレーサーとして添加することに基づいて最適化された方法であり、水生捕食真核生物における放牧率の信頼性の高い定量化と、より高い栄養レベルへの栄養移動の推定を可能にする。
Introduction
ヘテロ栄養原核生物は、水生系における重要な生物学的成分であり、プランクトンバイオマス1、2、3のかなりの部分を占める。その豊富さ、多様性、および活動を制御する要因は、生物地球化学的サイクリング(すなわち、有機炭素およびその他の栄養素の運命と原核生物から高い栄養レベルへのエネルギーの流れ)におけるその役割を理解するために重要です。原生動物の放牧は、これらの重要な要因の一つです。異種栄養性ナノフラグラテスおよび毛様体の細菌学は、原核生物の豊富さ、コミュニティ機能、構造、多様性、さらには特定の細菌群の細胞形態および増殖速度に対して強力なトップダウン制御を課す4、 5,6.いくつかのシステムでは、プロティストは細菌死亡率6、7の主な原因として機能します。
現在しばらく使用されている原生菌を評価するために使用される標準的なアプローチは、蛍光標識細菌(FLB)を獲物類似体および上蛍光顕微鏡として使用することを含む。細胞特異的取り込み速度は、選択した時間経過8にわたってプロテチスタン食品真空中の標識された獲物粒子の数を定量化することによって決定することができる。この方法にはいくつかの利点があります。トレーサーは、自然の捕食者と獲物の組み立てと自然なサンプルに追加されます。インキュベーションの前に最小サンプル操作、追加されたFLBトレーサーによる最小サンプル改変、およびインキュベーション時間は、その中で近い条件下で得られる音の結果を確実にするために短いです。あるいは、細菌性プロティストや動物プランクトンの数が少ない環境(例えば、沖合の海洋システム)では、低量(2%-3%トレーサー)でサンプルに添加されたFLBの消失率は、長期(12-24時間)のフローサイトメトリーを介して検出することができる。インキュベーション実験。次に、始点と終点におけるFLBの数(すべての細菌の影響を統合する)は、フローサイトメトリーによって定量される(詳細については、前の出版物9を参照)。しかし、このようなパラメータは、特定のプロチスタンおよび動物プランクトン放牧者群または種に直接起因することができない総集約細菌学率のみを表す。
全体的に、水生環境におけるプロテスタン種または形態型特異的細菌死亡率を正確かつ生態学的意味で定量化することは困難であり得る。一部のプロティストは選択的な放牧者であり、追加されたFLBトレーサーの大きさおよび細胞形状は、獲物摂取の自然な速度を歪める可能性がある10、11。さらに、プロティスタン活性と代謝は非常に温度感受性12である。したがって、追加されたFLBトレーサーの量は、個々のサンプルタイプごとに慎重に操作する必要があります(細菌の自然な豊富さ、大きさ、形態、細菌の種類だけでなく、温度にも基づいて)。ほとんどの研究は、バルクプロテスタン放牧活性に焦点を当てています。しかしながら、特定のプロテスタン種の細菌学は、多くの場合、はるかに高い情報値を保持し、好ましい場合があります。この場合、サンプル中に存在するプロチスト種の分類知識と、その挙動を理解する必要がある。したがって、特定のプロテスタン属または種に起因する細菌類の細菌学的割合に関する健全な結果を得るためには、かなりの時間と労力が必要である。
これらの困難にもかかわらず、このアプローチは、自然な設定で前腺細菌を評価するために現在利用可能な最も適切なツールのままです。ここでは、水生微生物生態学研究のトレーサーとしてFLBを使用するための包括的で簡単に従う方法です。アプローチの前述の問題のある側面のすべてを説明し、改善されたワークフローが説明され、対照的な環境からの2つの実験だけでなく、対照的なシリアート種を例として説明する。
最初のケーススタディは、チェコ共和国のメソトロフィック・ジモフ水溜所からエピリムネティック環境で行われ、ほとんどの表面淡水体に匹敵する放牧者と細菌の豊富さを示した(cf.5,7)。第2のケーススタディは、両方の放牧ミキシック性毛様の非常に高い数を有する水生肉食植物ウトリピア反射のトラップ内の非常に特異的な環境で行われた(テトラヒメナutriculariae)と細菌細胞。両方のサンプルタイプの細胞特異的放牧率と細菌スタンディングストックの計算が示されています。その後、結果の生態学的解釈の範囲が議論され、可能なフォローアップ研究の例が最終的に提案されます。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. サンプルコレクション
-
貯水池水サンプルの採取:最初のケーススタディ(Exp I;その地の捕食者と獲物の豊富なシステムにおける低い自然)
- 適切な深さで所望の場所から水のサンプルを収集します。実験室への輸送中に、サンプルを温度制御されたクーラーに入れて保管してください(温度ショックを避け、プロティストの取り込み速度は温度に依存することに留意してください)。
注:私たちのサンプリングは、メソ・エトロフィック・キャニオン形のシモフ貯水池(南ボヘミア、体積34.5 x 106 m3、最大深さ43m、平均保持時間100日、小数点以下)で行われました。サンプリングサイトは、ダムに近い30メートルの深さに位置していました。深さ0.5mの混合サンプルを2lフリードンガーサンプラーで採取した。 - できるだけ早くセクション 1.2 に進みます。
- 適切な深さで所望の場所から水のサンプルを収集します。実験室への輸送中に、サンプルを温度制御されたクーラーに入れて保管してください(温度ショックを避け、プロティストの取り込み速度は温度に依存することに留意してください)。
-
肉食性ウトリピア反射植物のトラップからの流体のコレクション:第2のケーススタディ(Exp II;高い捕食者と獲物の豊富さ、非常に小さなサンプル量を持つシステム)
- 水中で植物をゆっくりと振り、栽培容器から取り出し、吸収性物質の上に置いて余分な水分を吸収します。植物の堅牢性とトラップのサイズに応じて、8-10植物の個体を選択します。
- トラップ器官を持つ葉のノードを数えることによって、各シュートをほぼ等しい部分に分割します。各シュートセグメントは、若い、中年、および古いトラップを表す混合サンプルを収集するのに役立ちます。
- 薄いガラスキャピラリーとエッペンドルフバイアルを取り付け、サンプル収集用に蠕動ポンプに取り付けます。トラップ開口部に毛細血管の先端を挿入する手順に進みます。真空ポンプを使用して、各トラップからすべての流体を吸い出し、900 ±100 μLのトラップ流体が各トラップ年齢カテゴリに回収されるまで採取します。
- プロチスタン放牧実験には、プールされたトラップ液の200μLのトリプリケートサブサンプルを使用します。セクション 3 で詳しく説明したように、これらを直ちに処理します。以下に詳述するように、流体中に生息する微生物成分の他のすべての分析のために、サンプルの残りの±300 μLを保持します(セクション2)。
- 直ちにセクション 2 に進みます。
2. 収集したサンプルの固定
- Exp IおよびII:細菌列挙のための水/トラップ流体サブサンプル(セクション4、それぞれ約20 mLおよび0.3 mL)をホルムアルデヒドで少なくとも1時間固定し、各サンプルの2%の最終容積:体積濃度を得る。
注:ホルムフードの中でホルムアルデヒドを排他的に取り扱い、サンプルを操作しながら常に手袋を着用してください。
3. サンプルろ過
- サンプル希釈(Exp I):貯水池の水サンプルに希釈は必要ありません。(Exp II):トラップ液サンプルを粒子フリーMQ水で10x-100x希釈し、エピ蛍光顕微鏡でカウントする前にフィルター表面上の標的微生物の適切な分布を達成します。
- 濾過漏器(直径25mm)を用いた濾過器を用いた黒い0.2 μmの細孔サイズフィルターに対する細菌用トラップ液のサブサンプルの貯留水の1-2 mL(Exp II)または10-30 μL(Exp II)をフィルターします。
- DAPI(4',6-diamidino-2'-フェニリンドール二塩酸塩,0.2%最終濃度)で4分間フィルターを汚します。
注:皮膚や作業面の汚染を避け、手袋を着用してください。 - 濃縮された微生物を含むフィルターを、顕微鏡スライド上の浸漬油(蛍光顕微鏡検査用)の上に置きます。フィルターの中央に別のオイルドロップを置き、カバースリップでカバーし、オイルが均等に分配されていることを確認します。
- この時点で、好ましくは直ちに試料を処理し、または代わりに、さらに分析するまで数週間から数ヶ月間冷凍庫(-20°C)に保存する。
4. フィルター上の細菌数の列挙
- スライドをエピ蛍光顕微鏡の下に置きます(フルオロクロムDAPIに対応するフィルターセット付き)。10 x 10 のカウント グリッドを眼の 1 つに配置します。スライドをランダムな位置に移動します。
- カウントグリッドの面積(倍率1000倍下)で細菌細胞(青色蛍光)を定量する。カウントには、棚卸グリッドの左端と上端を横切るセルを含め、右端と下端に位置するセルを除外します。
- 別のランダムな位置に移動し、合計でカウントされる 500 個のセルに相当する少なくとも 10 ~ 15 個のカウント グリッドで列挙を繰り返します。
- フィルタの総有効濾過面積に対する1グリッドの面積の比率に関する知識に基づいて、所定の顕微鏡および倍率の変換係数を確立する。次に、カウントされたセルの合計数をカウントしたグリッド数で除算し、グリッドあたりの平均細菌数を生成します。
- 後者のパラメータに設定された変換係数を乗算し、サンプルのmL当たりの結果数を正規化し(濾過されたサンプルの量に応じて)、mL当たりの総細菌量を得ます。
5. プロティスタンの豊富さの決定
- グルタルアルデヒド(1%最終濃度1%、固定後数日から数週間処理される粒子を含むクロロフィルの存在を有するサンプルに適した)またはトラップ流体(Exp II)サブサンプルを固定するフォルモルチオ硫酸塩脱色技術は、以下に示す。
注:両方の保存技術は、プロティスト8の食物空洞からの摂取材料のエジェストを防ぎます。 - フォルモルチオ硫酸塩脱色技術に関しては、水/トラップ流体サブサンプル(Exp I/Exp II)の20mL/200 μLにLugolの溶液の100 μL/1 μLを追加します。
- 直ちに0.5mL/50 μLのボレート緩衝ホルマリンを加え、次いで3%チオスルフェートナトリウムの20 μL/2 μL(それぞれExp I/Exp II)を加えて従ってください。
注:チオ硫酸ナトリウムは、上蛍光顕微鏡8の下で細胞の観察を可能にするために、ルゴルの黄色の色を脱色します。 - 既知のサンプル量(標的前駆体の数に応じて)を1 μmの細孔サイズの黒いポリカーボネートフィルターに濾過します。
- 前駆体種の数を推定する場合は、カウントグリッドを用いて倍率600倍以下の少なくとも200細胞を数えることによってである(上記参照)。
- ろ過漏斗内の真空下の試料の体積を低真空で約2mLに減らします。次いで、アンダープレッシャーを放出し、DAPIフルオロクロム(4',6-ジアミド-2-フェニリンドール二塩酸塩13,0.2%最終濃度)を2分間添加する。
6. プランクトンサンプルにおける毛様体のコミュニティ構造の決定
注:淡水生息地のciliateコミュニティは非常に多様である14、15、16、18、そして彼らの顕微鏡的な決定は挑戦的です。ciliate基を機能ギルド10、14、16、17に分類すると、ペラジックバクテリボレスとして異なる毛様基のより詳細な分析が可能になります。
- 次を組み合わせて、ciliate コミュニティ構造を評価します。
- 上蛍光顕微鏡におけるDAPI染色サンプル(異なるサイズおよび形態のマクロ核および微小核の明るい蛍光を有する毛様細胞を局在化させる)と蛍光標識細菌の取り込み(FLB8;詳細については、以下を参照してください)は、細菌を養うために毛様の能力を追跡します。
- 選択した症例17,19のライブサンプル観察。ciliates を異なる分類カテゴリにグループ化するために使用される上記のアプローチと基準の詳細については、前の出版物16,17を参照してください。
注:引用された研究は、シチコトリシア(ジェネラ・ハルテリアとペラゴハルテリア)とオリゴトリシア(すなわちリモストロミジウム属)の中で最も重要なペラジック消費者であることを示しています。淡水生息地の大部分でバクテリオプランクトン10,17,18.
7. 毛様の放牧率の推定
- トレーサーの平均数の変化(すなわち、インキュベーション時間(5〜15分)に関連する毛様化当たりFLB8)とFLBのトレーサー量に基づいて細菌に対する毛様の取り込み率を計算し、全細菌の5%~15%を占める。
- 異なるプロテチスタン種間の取り込み率を比較するために、取り込み速度を1時間当たりの細菌数として正規化し、実際のインキュベーション時間とトレーサーFLBの割合に基づく計算を加えた。
注:FLB法アプリケーションの一般的なスキームは、天然サンプル中の細胞または種特異的およびバルクバクテリア率の両方を推定する図1に示されている。 - 細菌株から淡水環境へのFLBの調製8
- 調査対象の水生系におけるバクテリプランクトン/細菌細胞の典型的なサイズを効果的に模倣できるように、適切なサイズ(平均細胞体積=MCV)と細菌の形態を選択します。
注:Exp Iの場合、リムノビアン属およびポリヌクレオバクター属の研究場所から単離された株の混合物が使用された(すなわち、典型的な、湖沼における非常に豊富なバクテリオプランクトン)20。菌株の形態とサイズの詳細については、以前の出版物3、17、18、21を参照してください。 - 初期の静止期で15分間遠心分離(5,000 x g)で培養から細菌細胞を収穫し、選択した場所で細菌の典型的なMCVに対応する混合物中の細胞のMCV±SDを得る数値比でそれらを混合する。
- リン酸緩衝生理食べ物の10mLでペレットを中断する(PBS;pH=9)。
- 黄緑色蛍光色素5-(4,6-ジクロロトリアジン-2-yl)の2mgをリン酸生理食塩水バッファー内の細胞懸濁液に添加し、2時間60°Cの水浴中でインキュベートする。
- インキュベーション後、細胞を遠心分離し、DTAF溶液をデカントし、PBSで洗浄および遠心分離機3倍を洗浄する。
- 最終洗浄後、PPi-生理液バッファーの20mLで細胞を再中断する。
- FLBサスペンションとピペット1.5mLアリコートを2mLクライオバイアルに渦を入れ、使用するまでPP i-salineバッファで凍結(-20°C)を保ちます。
- 次のステップで使用するための0.2 μmポリカーボネートフィルターを通してPPi-生理食べ物バッファを事前にフィルタリングします。
- FLB濃度を決定するには、小さなアリコート(通常は20-40 μL)を粒子フリーPP i-salineバッファーの2 mLに移し、30Wで30Wで超音波処理を行い、0.2 μmのポリカーボネートブラックフィルターに濾過して、上蛍光を介して列挙します。DTAF(448 nm/520-540 nm)の光学フィルタ設定下で顕微鏡検査(1,000倍)。
- 調査対象の水生系におけるバクテリプランクトン/細菌細胞の典型的なサイズを効果的に模倣できるように、適切なサイズ(平均細胞体積=MCV)と細菌の形態を選択します。
- 毛様性細菌の推定のためのトレーサー技術
- 自然なプランクトン生息地での放牧実験では、300 mLサンプルを十分にすすりすいで1Lフラスコに分配し、15分間その中の温度でインキュベートします(プロティストがハンドリングショックから回復できるようにします)。
- FLBトレーサーを追加して、季節と水温に応じて添加した量で、全細菌の5%~15%を構成します。
注:大きさのいくつかの順序にまたがる毛様種特異的取り込み速度(すなわち、101-104細菌から-1時間あたりの-1)10、16、16に及ぶ非常に広い、季節依存性スペクトルがあります。 17,18,22,23,24. - 取り込み率の高い毛様の発生が増加した期間(通常は夏の間)、また、非常に低いFLB添加物で並列インキュベーションを実行し、トレーサーFLBによる毛様化真空の過剰な負荷を避けるために、総細菌の2%-4%のみを構成する(参照)図 2の例)。
- FLBでシリート/プランクトンサンプルを5~15分間インキュベートします。
- プロティスト8の食品バキュールから摂取した材料のエジェストを防ぐためにサンプル固定のための2つの可能性があります。1%グルタルアルデヒド(藻類などのクロロフィル含有粒子を有するサンプルに適した最終濃度)を添加してインキュベーションを終了する。あるいは、Lugolの溶液の100 μL/10 μLを水/トラップ流体サブサンプルの20 mL/200 μLに使用し、その直後にボレート緩衝ホルマリンの0.5 mL/10 μLを添加し、次いで3%チオスルフェートナトリウム(ExpI/Exp II)の200 μL/2 μLを添加する。
- 固定剤を添加した後、サンプルを4°Cの暗闇の中で少なくとも1時間休止させ、毛様細胞の完全な保存を確保します。
- 4-30 mL/10-30 μL(Exp I/Exp II、体積はそれぞれ毛様数に依存)から自然プランクトンサブサンプルを採取し、DAPI(最終濃度0.2%wt/vol;詳細については、上記のステップ3.2を参照)で染色します。
- 1 μmの黒いフィルターを通過し、上蛍光顕微鏡を介して検査して、毛様化(600倍倍)を数え、以前の出版物2、17で詳述したように、摂取したFLBトレーサーの数(主に1000倍倍)を列挙する.保存後7日以内に検査を行います。
- 原虫/種特異的放牧の合計を推定するには、すべての毛様体の平均取り込み速度、またはその中で検出された種のみを乗算します。
- シモフ水貯水池からのその場データからの細胞ごとの取り込み率の計算の例は、以下のように説明します。
- 細菌濃度が3.55 x 106細菌/mLであり、FLBが加えられたトレーサーFLBが0.25 x 106 FLB/mLであると仮定すると、総細菌粒子の合計は3.8 x 106細菌/mL(天然細菌+FLB =獲物粒子の100%)自然なサンプルの咽頭のプロティストに利用できる。
注:このように追加されたFLBトレーサーは、総細菌粒子の6.58%(0.25/0.038のプロジェクト)を表します。ハルテリアsp.当たりのFLBの平均数は、5分のインキュベーションで6.2 FLBです。 - 1 時間あたりの取り込み取り込みを正規化するには、次の計算を使用します: (6.2 x 12)/(6.58/100) =ハルテリアセル/hあたり 1131 細菌。
注:変動水温下で検出されたハルテリアsp.の個々の取り込み速度の分布のより多くの例、トレーサーFLBの量(パーセントとして)が追加され、FLBとの異なるインキュベーション時間は、図3を参照してください。
- 細菌濃度が3.55 x 106細菌/mLであり、FLBが加えられたトレーサーFLBが0.25 x 106 FLB/mLであると仮定すると、総細菌粒子の合計は3.8 x 106細菌/mL(天然細菌+FLB =獲物粒子の100%)自然なサンプルの咽頭のプロティストに利用できる。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
実験の例は、捕食者や獲物の豊富さが低い自然の自然地であるシモフ水貯水池(南ボヘミア、CZ)で実施した。代表的なデータは、ピコプランクトン(<2 μm)粒子10、16、17、18の豊富で効率的なグレイザーである全能の毛様種ハルテリア・グランジネラについて報告されています。 、22.図3は、1時間当たりの細菌取り込み率に再計算された、ハルテリアsp.のセル当たりFLB数の箱ひげプロット(図3A)を示す(図3B)。4月、5月、8月、9月に実施された4つの個別実験で検出されました。 水温の時間的な違いが主に引き起こされた、毛様化取り込み速度の変動が大きかった。
Q10パラメータは、微生物プロセスが10°C12の温度上昇により約2.5倍速く実行され、細菌の取り込み率も高くなるという事実を反映していることに留意すべきです。この生理学的ルールを念頭に置いて、FLBとインキュベーション時間のかなり異なる割合が異なる季節に使用されました(詳細については、図3Aを参照)。したがって、期待される温度効果を補正し、実験設定では、毛様細胞当たり約5〜10 FLBの取り込み率の最適化平均値と中央値値が得られた。一般に、これらの量の取り込まれたFLBは簡単にカウント可能です(図2の例、左の写真2枚を参照)、トレーサーの正確な推定値(主に1~15FLBの間にある)取り込み率を生成します。ただし、FLB トレーサの変更により(%)サンプルインキュベーションの異なる時間の絶対値(1時間あたりの細菌の放牧された数として表される)は有意に異なっていた(p < 0.01、Kruskal-Wallisテスト;ダンの多重比較試験、p<0.05;図の例を参照) 3B)を実験中にした。このデータはまた、ハルテリア・グランジネラのプランクトン集団における絶対細菌学的率における典型的な自然変動を示し、その平均値と中値に近い一致を示す(図3)。
ペリチスチ酸様などのサンプル中に非常に効率的な細菌性の毛様が存在すると、全細菌の5%~10%の典型的なトレーサー量でFLBによって重く「過剰標識」される可能性があります(図2の右側の写真を参照)。これは、摂取されたFLBの正確な定量を強く制限する可能性があります。このような場合、全細菌のわずか1.5%-3%のFLB会計の低量で追加の並列インキュベーションを実行することが示唆される。しかし、一般に、トレーサー量とインキュベーション時間の両方を操作して、セルあたりのFLB数を最適化することができます(図2)。
実験IIの例:表示されるのは、大きな捕食者と獲物の豊富なシステムからのデータで、非常に小さなサンプル量のみが、ジリエートテトラヒメナutriculariaeの細菌学的率を実験的に推定するために利用可能である25.肉食性ウトリピア反射植物26,27のトラップの中で排他的に豊富に生息する適度な細菌放牧者である。図4は、異なる実験設定(図4A,B)下でのT.ウトリキュラリアの細胞あたりのFLB数の箱とウィスカーのプロットを示し、1時間あたりの細菌取り込み率に再計算する(図4)。C,D)若い、成熟した、古いトラップで検出されます。 興味深いことに、トラップでは、毛様体T.ウトリキュラリアのクロロプラストを有する集団が検出され、T.utriculariaeの無形成集団はトラップから単離され、増殖する混合細菌懸濁液に維持された。 暗闇の中の小麦粒(詳細については、前の出版物26の図1を参照)。
クロロプラストを持つ集団は、照らされたトラップに住んでいます。したがって、葉緑体は、毛様体宿主に追加の有機炭素源および酸素を提供することができる。テストされた仮説の1つは、細菌が毛様体の暗く成長した単離された亜集団に利用可能な有機炭素の唯一の粒子源を表すので、アポクロリック・シリアート集団が細菌を有意に速く放牧したことです。
確かに、若くて成熟した古い神経衰弱に生息するシリアの細菌学的率に有意な差はなかったが(図4A,C)、T.utriculariaeのアポクロリック集団は放牧された細菌が有意に(p < 0.01,Kruskal-Wallisテスト;続いてダンの多重比較試験、p<0.05)、若く、成熟した、古いトラップに生息するクロロプラストベアリングの毛様体よりも約3倍速い(図4C,D)。繰り返しになりますが、トレーサー量とインキュベーション時間(図4A、B、上)の両方を変更して、セルあたりのFLB数(一般に1~15)を最適化し、平均値と中央値は5FLB/ciliateの周りに行った。これらの数字は、毛様化食品の真空で区別され、正確なトレーサーカウントが可能です。しかし、1時間当たりの放食菌数では、クロロプラストベアリングとアポクロリック集団はそれぞれ約350と1,000個の細菌を1時間あたりに放牧した。この実験的なセットアップは、著しく異なる環境制約の下で生きている同じ毛様種の2つの異なる亜集団の代謝および生理学的形質に新しい洞察をもたらした25,26, 27.
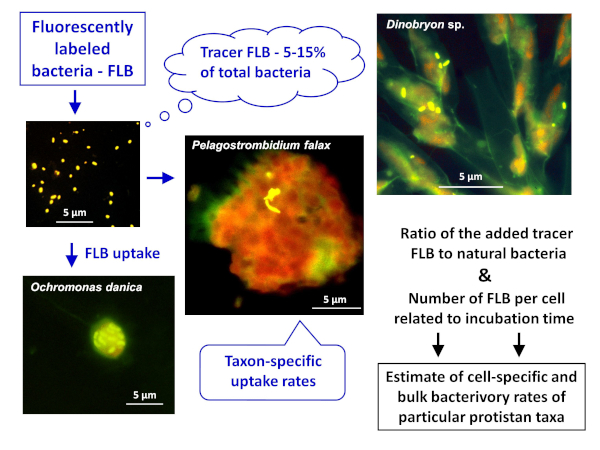
図 1: 蛍光標識細菌(FLB)を用いて、試料中の天然細菌の総数に対する摂取トレーサーFLBの比から細胞および種特異的放牧率を推定するワークフロー。詳細については、プロトコルのセクション 7 を参照してください。この図のより大きなバージョンを表示するには、ここをクリックしてください。

図 2:富栄養魚池のプランクトンからの毛様細胞の例。例は、毛様細胞中のカウント可能なFLBを持つ池から示されている(一般に細胞当たり1〜10トレーサーFLB、左2マイクロフォトグロープ)、腹膜シリアベオロボルチエラナタン(右側のマイクロ写真)と比較して。短い、5分の潜伏期間の間に、それはトレーサーFLBによって「過剰標識」となり、摂取されたFLBの定量を不正確またはほとんど不可能にした。この場合、トレーサー量を全菌の1.5%〜3%に減少することが示唆される。しかしながら、一般に、トレーサー量とインキュベーション時間の両方を操作して、細胞当たりのFLB摂取数を最適化することができる。詳細については、プロトコルのセクション 7 を参照してください。この図のより大きなバージョンを表示するには、ここをクリックしてください。

図 3: ハルテリアsp.の細胞当たりのFLB数のボックスウィスカープロット(Exp I)(A)から、1時間当たりの細菌取り込み率(B)に再計算した。データは、4月から9月までの4つの例で表される、異なる季節設定で検出されました。パネルAの上部には、水温、追加された異なるFLBトレーサー(%)、およびサンプルインキュベーションの異なる時間に関する情報が表示されます。後者の2つのパラメータは、セルあたりのFLB数を最適化するために変更することができ、平均(全線)と中央値(破線)値は、1つの毛様セルあたり約5〜10FLB(A)の間で行うことができることに留意されたい。バーは、すべてのデータの25番目と75パーセンティル(50-180細胞検査)を示し、ウィスカーは1番目と99パーセンシーを表します。(B)異なる小文字は、研究期間中のハルテリアsp.の細胞特異的細菌学率に有意な差を示す。この図のより大きなバージョンを表示するには、ここをクリックしてください。

図 4: セルあたりの FLB 数のボックス とウィスカープロット。プロットは、若い、成熟した、およびウトリキュラリア反射(Exp II)(A)の三量的処置からクロロプラストベアリングテトラヒメナutriculariaeを示し、1時間当たりの細菌取り込み率(C)に再計算される。データは、トラップから分離されたT.utriculariae(B,D)の重複する無声集団の細菌取り込み率と比較されたが、暗闇の中で小麦粒に成長する混合細菌懸濁液に維持された。パネル A と B の上部に、異なる FLB トレーサーが追加されました (%)サンプルインキュベーションの異なる時間が示されています。後者の2つのパラメータは、セルあたりのFLB数を最適化するために変更され、平均(全線)と中央値(破線)値は、1つの毛様セルあたり約5〜10FLB(A,B)の間で行われたことに留意された。バーは、すべてのデータの5番目と95パーセンシー(50-100細胞検査)を示し、ウィスカーは1番目と99パーセンシーを表します。この図のより大きなバージョンを表示するには、ここをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
水生系における栄養相互作用の解読は、特にプロティストとその獲物である細菌を含むナノプランクトンスケールにおいて、常に28に挑戦している。栄養摂取経路と定量に関しては、生物学的相互作用の複雑さが高いため、より高い栄養レベルで正常に使用される方法の適用は不可能です。これらには、例えば、安定した同位板標識アプローチが含まれる。このプロトコルは、炭素流を追跡し、半定量化/推定するためのトレーサーとして上蛍光顕微鏡と蛍光標識細菌を使用する利点を示しています(細菌の獲物:ミキソートロフィックグレイザー29を含む様々なプロチスタングレイザー)微生物の食物網の基盤を通る経路。そのような利点の1つは、単一細胞アプローチの高精度であり、もう一つは、グレイザーコミュニティ構造に関する前例のない解像度であり、異なる機能ギルド、種(Exp I)、さらには同じ亜集団を区別することです。種(エクスプII)。
プロトコルの重要な手順
プロトコルにはいくつかの重要な手順があり、方法論の利点を最大限に活用できます。第一に、実験開始前の研究環境の基本的な理解は、常に有益である。これには、水柱内の潜在的な放牧者の多様性と豊富さ、細菌の獲物サイズ、および獲物分布の両方の顕微鏡スクリーニングが含まれます(例えば、エピリムニオンから低体温に至る垂直プロファイル)と2の場合ダム流入トランセクト上の渓谷形の貯水池。第二に、収集されたサンプルとの注意深い操作は代表的な結果を保障する。温度は、原生体放牧速度を含むほとんどの微生物プロセス12に影響を与える非常に重要な要因である(図3)。
第3に、サンプル中の細菌細胞またはグレイザーの種類の定量に基づいて添加されたトレーサーの量を操作することで、過剰標識(図2)の問題が解消されることを保証する。毛様種特異的取り込み率には非常に広いスペクトルがあることに留意すべきです(詳細については、ステップ7.2を参照)。したがって、プロトコルを適切に適用するためには、その時間経過率を有する主要な毛様種の事前知識が不可欠である。可能なシリートアンダーラベリング(シリートセル当たり取り上げられ、統計的に不健全なデータを生成するFLBが少なすぎる)または過剰ラベリング(FLBの数が多いように見える)を避けるために、異なるトレーサー量で予備実験を行うことを強くお勧めします。トレーサーによって詰められた毛様体食品の真空中に「凝縮FLB雲」または群れを形成し、その正確な定量を著しく制限する。図 2の右上の例を参照してください。また、FLBのインキュベーション時間は一般的に30分より短く、毛様化によるピコプランクトンの平均消化時間は約1.5時間であり、消化が始まる(摂取したピコプランクトン細胞は、その典型的な形状と色を失う)。45-60分後30.同様に、顕微鏡表示の前にフィルター上のサンプルの最適な希釈と分布は、正確な結果を得るために達成される必要があります。
変更とトラブルシューティング
この手法の主な手順、可能な変更、およびトラブルシューティングの変更を図 1および図 2 に示します。さらに、高濃度の有害粒子、植物プランクトン細胞、またはプランクトン中のコロニーの場合、そのようなサンプル1)は、個々のグレイザー細胞が可能な段階を達成するために対応して希釈されるべきであることに留意すべきである。フィルター表面と2)食品真空中身の定量化を行う。
制限
この方法の正常な適用のための主な制限は、エピ蛍光の下で明確なサンプルの表示を妨げる量の付着した細菌または凝集体を有する様々な有機的な不毛または豊富な無機/有機粒子の存在にある顕微鏡とトレーサー量の正確な推定を追加しました。提示されたトレーサー技術は、主に粒子に付着していない自由(すなわち、懸濁)細菌で動作することに留意すべきである。しかし、私たち自身の経験と文献の参考文献に基づいて(以前の出版物2、4、8、10、16、18、21を参照してください)、26)、提示された方法論は、ほとんどの水生環境に適している。栄養状態、有害含有量、およびグレイザーの多様性と数が異なる2つの自然な対照的なシステムの例が提供されています(図3および図4)。
既存の方法に対するアプローチの意義
重要なのは、バクテリアの分類/分類の豊富さとその種特異的な細菌学率の知識から、プロテスタン分類(または総毛様体組み立て)のバルクバクテリア率を計算することができる。このアプローチが自然なプランクトン環境に同時に適用される場合(バクテリオプランクトン2、6、7の主要な放牧者を表す)、プロチスタン所定の環境における細菌集団の放牧誘発ターンオーバー時間は、16、17、18、22と推定することができる。このようなデータは、微生物食品ウェブにおける炭素流動力の推定にとって基本的な重要性を持っています。
今後のアプリケーション
このメソッドは、いくつかの変更を加えて、正常に使用できる他の特定の環境があります。これらには、活性汚泥システム、ルーメン生態系、水生堆積物、および肥大性魚池17が含まれる。しかし、これらの栄養と微生物が豊富な環境でのアプリケーションは、典型的なサイズ分布やその他の特性を模倣することができる適切なサイズ、形態、およびトレーサーFLBの数に関するプロトコルを最適化するための予備テストを必要とします。環境に固有の獲物細菌の。
現在、このアプローチを触媒レポーター堆積蛍光と組み合わせることに関心が高まっており、グレイザー細胞の同一性(例えば、ヘテロ栄養性旗石)が特定の方法で検出される場ハイブリダイゼーション(CARD-FISH)において、FISHプローブおよび取り込み速度は、同じ顕微鏡スライド31上のフラグレート細胞の食品真空中のFLB含有量に基づいている。二重ハイブリダイゼーション32と呼ばれる洗練された新しいアプローチは、捕食者細胞および獲物細菌のレベルにおけるFISHプローブの組み合わせである(これはまた、系統的株、細菌系統特異的FISHプローブによって特異的に標識される)。このアプローチはエレガントですが、時間もかかるもので、特定のスキルと経験を必要とします31,32, 様々なFLB取り込みアプローチの変更の適用は、実験室での日常的な使用のためにより簡単に採用することができます.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者は何も開示していない。
Acknowledgments
この研究は、K.Sh.とD.S.に授与された研究助成金13-00243Sと19-16554Sの下でチェコ科学財団によって支援されました。この記事はまた、欧州地域開発基金が出資する「ダム貯水池の水質改善ツールとしてのバイオマニピュレーション」(CZ.02.1.01/0.0.0.0/06_025/0007417)プロジェクトの支援を受けました。そして教育。
Materials
| Name | Company | Catalog Number | Comments |
| 0.2-µm pore-size filters | SPI supplies, https://www.2spi.com/ | B0225-MB | Black, polycarbonate track etch membrane filters, diameter approprite for filtering apparatus used |
| 5-(4,6-dichlorotriazin-2-yl) aminofluorescein (DTAF) | Any brand | ||
| Automatic pipettes with adjustable volumes | Any brand, various sizes | ||
| Centrifuge | 22 000 x g | ||
| Cryovials | Any brand, 2 mL size | ||
| DAPI (4´,6-Diamidino-2´-phenylindole dihydrochloride) | Any brand | 1 mg ml-1 | |
| Epiflorescence microscope | Magnification from 400 x up to 1000 x | ||
| Filters appropriate for viewing in the DAPI and DTAF range | |||
| Counting grid in one of the oculars | |||
| Filtering apparatus | Usually with a diameter of 25 mm | ||
| Formaldehyde | A brand for microscopy | ||
| Glutaraldehyde | A brand for microscopy | ||
| Immersion oil for microscopy | Specific oil with low fluorescence | ||
| Lugol´s solution | Any brand or see comment | Make an alkaline Lugol' solution as follows: Solution 1 - dissolve 10 g of potassium iodide in 20 ml in MQ water, then add 5 g of iodine. Solution 2 - add 5 g of sodium acetate to 50 ml of MQ water. Add the solution 2 to the solution 1 and thoroughly mix | |
| Methanol stabilized formalin | Any brand available for microscopy purposes | ||
| Microscope slides and cover slips | Any brand produced for microscopy purposes | ||
| MQ water for diluting samples | Any brand |
||
| Phosphate-buffered saline (PBS; pH = 9) | Any brand | 0.05 M Na2HPO4-NaCl solution, adjusted to pH 9 | |
| PPi-saline buffer | Any brand | 0.02 M Na4P2O7-NaCl solution. Add 0.53 g Na4P2O7 to 100 ml of MQ water plus 0.85 g NaCl | |
| Sampling device | Appropriate for obtaining representative sample | e.g. Friedinger sampler for lake plankton | |
| Sodium thiosulfate solution | Any brand | 3% solution is used in the protocol | |
| Sonicator | Any brand | 30 W | |
| Vortex | Any brand allowing thorough mixing of the solutes and samples | ||
| Water bath | Any brand allowing temperature to be maintained at 60 °C |
References
- Azam, F., et al. The ecological role of water-column microbes in the sea. Marine Ecology Progress Series. 10, 257-263 (1983).
- Šimek, K., et al. A finely tuned symphony of factors modulates the microbial food web of a freshwater reservoir in spring. Limnology & Oceanography. 59, 1477-1492 (2014).
- Šimek, K., et al. Bacterial prey food characteristics modulate community growth response of freshwater bacterivorous flagellates. Limnology & Oceanography. 63, 484-502 (2018).
- Šimek, K., et al. Changes in bacterial community composition, dynamics and viral mortality rates associated with enhanced flagellate grazing in a meso-eutrophic reservoir. Applied & Environmental Microbiology. 67, 2723-2733 (2001).
- Jürgens, K., Matz, C. Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Antonie Van Leeuwenhoek. 81, 413-434 (2002).
- Pernthaler, J. Predation on prokaryotes in the water column and its ecological implications. Nature Reviews Microbiology. 3, 537-546 (2005).
- Berninger, U. B., Finlay, J., Kuuppo-Leinikki, P. Protozoan control of bacterial abundances in freshwaters. Limnology and Oceanography. 36, 139-147 (1991).
- Sherr, E. B., Sherr, B. F. Protistan grazing rates via uptake of fluorescently labeled prey. Handbook of Methods in Aquatic Microbial Ecology. Kemp, P. F., Sherr, B. F., Sherr, E. B., Cole, J. J. , Lewis Publishers. Boca Raton, Florida. 695-701 (1993).
- Vazquez-Dominguez, E., Peters, F., Gasol, J. M., Vaqué, D. Measuring the grazing losses of picoplankton: methodological improvements in the use of fluorescently tracers combined with flow cytometry. Aquatic Microbial Ecology. 20, 119-128 (1999).
- Šimek, K., et al. Ecological role and bacterial grazing of Halteria spp.: Small oligotrichs as dominant pelagic ciliate bacterivores. Aquatic Microbial Ecology. 22, 43-56 (2000).
- Montagnes, D. J. S., et al. Selective feeding behaviour of key free-living protists: avenues for continued study. Aquatic Microbial Ecology. 53, 83-98 (2008).
- Kirchman, D. L. Processes in Microbial Ecology. 2nd Edition. , Oxford University Press. Oxford, UK. (2018).
- Porter, K. G., Feig, Y. S. The use of DAPI for identifying and counting aquatic microflora. Limnology and Oceanography. 25, 943-948 (1980).
- Foissner, W., Berger, H. A user-friendly guide to the ciliates (Protozoa, Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste waters, with notes on their ecology. Freshwater Biology. 35, 375-482 (1996).
- Foissner, W., Berger, H., Schaumburg, J. Identification and ecology of limnetic plankton ciliates. Informationsberichte des Bayer Landesamtes für Wasserwirtschaft Heft. , 3-99 (1999).
- Šimek, K., et al. Ciliate grazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum: a study at the species and community level. Limnology & Oceanography. 40, 1077-1090 (1995).
- Skibbe, O. An improved quantitative protargol stain for ciliates and other planktonic protists. Archiv für. Hydrobiolgie. 130, 339-347 (1994).
- Macek, M., et al. Growth rates of dominant planktonic ciliates in two freshwater bodies of different trophic degree. Journal of Plankton Research. 18, 463-481 (1996).
- Šimek, K., et al. Microbial food webs in hypertrophic fishponds: omnivorous ciliate taxa are major protistan bacterivores. Limnology & Oceanography. , in press (2019).
- Jezbera, J., et al. Major freshwater bacterioplankton groups: Contrasting trends in distribution of Limnohabitans and Polynucleobacter lineages along a pH gradient of 72 habitats. FEMS Microbiology Ecology. 81, 467-479 (2012).
- Kasalický, V., et al. The diversity of the Limnohabitans genus, an important group of freshwater bacterioplankton, by characterization of 35 isolated strains. PLoS One. 8, 58209 (2013).
- Stabell, T. Ciliate bacterivory in epilimnetic waters. Aquatic Microbial Ecology. 10, 265-272 (1996).
- Zingel, P., et al. Ciliates are the dominant grazers on pico- and nanoplankton in a shallow, naturally highly eutrophic lake. Microbial Ecology. 53, 134-142 (2007).
- Bickel, S. L., Tang, K. W., Grossart, H. P. Ciliate epibionts associated with crustacean zooplankton in german lakes: distribution, motility, and bacterivory. Frontiers in Microbiology. 3 (243), (2012).
- Sirová, D., et al. Hunters or gardeners? Linking community structure and function of trap-associated microbes to the nutrient acquisition strategy of a carnivorous plant. Microbiome. 6, 225 (2018).
- Šimek, K., et al. Ecological traits of a zoochlorellae-bearing Tetrahymena sp. (Ciliophora) living in traps of the carnivorous aquatic plant Utricularia reflexa. Journal of Eukaryotic Microbiology. 64, 336-348 (2017).
- Pitsch, G., et al. The green Tetrahymena utriculariae n. sp. (Ciliophora, Oligohymenophorea) with its endosymbiotic algae (Micractinium sp.), living in the feeding traps of a carnivorous aquatic plant. Journal of Eukaryotic Microbiology. 64, 322-335 (2017).
- Nielsen, J. M., Clare, E. L., Hayden, B., Brett, M. T., Kratina, P. Diet tracing in ecology: Method comparison and selection. Methods in Ecology and Evaluation. 9, 278-291 (2018).
- Beisner, B. E., Grossart, H. P., Gasol, J. M. A guide to methods for estimating phago-mixotrophy in nanophytoplankton. Journal of Plankton Research. , 1-13 (2019).
- Dolan, J. D., Šimek, K. Processing of ingested matter in Strombidium sulcatum, a marine ciliate (Oligotrichida). Limnology and Oceanography. 42, 393-397 (1997).
- Massana, R., et al. Grazing rates and functional diversity of uncultured heterotrophic flagellates. The ISME Journal. 3, 588-596 (2009).
- Grujčić, V., et al. Cryptophyta as major freshwater bacterivores in experiments with manipulated bacterial prey. The ISME Journal. 12, 1668-1681 (2018).
