

Summary
本文提出了一种方法,将荧光显微镜的自(in)相容性与通过PCR分析识别S-基因型相结合的方法,确定杏(Prunus亚美尼亚卡L.)的授粉要求。
Abstract
罗萨塞的自不相容性是由一种主要由多环位点S控制的Game-phy-s不兼容系统(GSI)决定的。在杏中,确定自和(在)兼容性关系变得越来越重要,因为大量新的培养素的释放导致具有未知授粉要求的培养品种的增加。在这里,我们描述了一种方法,将通过手工授粉和显微镜确定自(in)相容性与通过PCR分析识别S-基因型的方法相结合。为了进行自我(in)相容性测定,从每个品种的气球阶段收集花在野外,在实验室手工授粉,固定,并沾上含艾琳蓝,以观察荧光显微镜下的花粉管行为。为了建立品种之间的不相容关系,从幼叶中提取了每个品种的DNA,通过PCR鉴定了S-等位物。这种方法允许建立不相容组,阐明品种之间的不相容关系,这为在设计新果园时选择合适的授粉器和在育种计划中选择合适的父母提供了有价值的信息。
Introduction
自我不相容是开花植物的一种策略,以防止自授,促进跨越1。在罗萨塞,这种机制是由一个游戏学自不相容系统(GSI)决定的,该系统主要由多环位点S22控制。在风格中,RNase基因编码S-stylar决定因素,RNase3,而决定S-花粉决定因素的F盒蛋白由SFB基因4编纂。自我不相容相互作用通过抑制花粉管生长的方式,防止卵管55,66的受精。
在过去二十年中,全世界都进行了品种更新, 7,,8.从不同的公共和私人育种项目引进了大量新的品种,导致杏培养品种增加,对未知授粉要求8。
使用不同的方法来确定杏的授粉要求。在现场,通过在笼子里的树木或被授精的花丛中进行对照授粉,然后记录水果集99、10、11、1210,11的百分比,12可以建立自(内)相容性。此外,在实验室中,通过半体内花卉培养和荧光显微镜下花粉管行为分析88、13、14、15、16、17,进行了控制授粉。,13,14,15,16,17最近,分子技术,如PCR分析和测序,允许在研究RNase和SFB基因18,19,19的基础上,对不相容关系进行表征。在杏中,有33个S-allee被报告(S1到S20,S S22到S30,S S52,S S53,S Sv,S Sx),包括一个与自我S兼容(Scc)12,18,20,21,22,23,24相关的等位基因。12,18,20,21,22,23,24截至目前,该物种已根据S-基因型,,8、9、17、25、26、279,17组,对该物种进行了25,26次不相容组。826具有相同S-等位基因的培养体是互不兼容的,而具有至少一个不同 S-alleleS的培养器,因此,在不同的不兼容组中分配,是相互兼容的。
为了确定杏培养的授粉要求,我们描述了一种方法,将荧光显微镜的自(in)相容性测定与杏仁培养体中PCR分析的S-基因型的识别相结合的方法。这种方法允许建立不兼容组,并阐明品种之间的不相容关系。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 自(内)兼容性确定
- 在田里品尝花。有必要在气球阶段采集花(图1A),与杏子28的BBCH刻度第58阶段相对应,以避免不必要的先前授粉。
-
实验室的自检和交叉授粉
- 在气球阶段取出花的麻醉剂,放在一张纸上,在实验室温度下干燥。
- 24小时后,使用细网(0.26毫米)筛分花粉颗粒(图1B)。
- 在同一气球阶段为每一个自授粉和交叉授粉而对30朵花进行浸化,并在实验室温度下将花花泡沫上的皮皮放在水中(图1C)。
- 在消瘦后24小时,手用画笔用来自同一品种24小时的花朵的花粉对皮丝进行授粉。此外,将每种品种的另一组皮质与来自兼容授粉器花的花粉进行控制(图1D)。
- 72小时后,在4°C29下将皮酸固定在乙醇/醋酸(3:1)的固定溶液中至少24小时。然后丢弃固定剂,加入75%乙醇,确保样品完全浸入溶液中。样品可在4°C下保存在4°C下,直到使用88、17、30、31、32。,17,30,31,32
-
通过体外花粉发芽评估花粉生存能力
- 制备发芽介质时,重量为25克蔗糖、0.075克硼酸(H3BO3)和0.075克硝酸钙(Ca(NO33)2)33。233
- 将介质的成分加入250 mL蒸馏水中,然后完全溶解。
- 凝固介质加入2克琼脂,并通过旋转混合。
- 使用 pH 仪表检查介质的 pH 值,并使用 NaOH 或 HCl 解决方案将值调整为 7.0。
- 高压灭菌混合物以消毒介质。
- 高压后,冷却介质并将其分发到无菌层流罩中的培养皿中。
- 在凝固的花粉发芽介质中分散用于受控授粉的相同品种的花粉颗粒,并在24小时6后在显微镜下观察它们。
注:要消毒层流罩,请使用 70% 乙醇清洁表面,并在 10 分钟内打开紫外线灯。 - 将培养皿存放在冰箱中,温度为 4°C,直到使用。
-
显微镜观察
- 用蒸馏水洗三次1小时,在4°C下将其留在5%硫酸钠中。24小时后,在硫酸钠10分钟内以1公斤/厘米2高压灭菌,以软化组织34。
- 将自切除的皮片放在玻璃幻灯片上,在手术刀的帮助下,取出卵巢周围的三角,以更好地可视化花粉管。然后,用盖玻璃挤压皮片。
- 制备 0.1% (v/v) 艾林蓝色污渍:将 0.1 mL 的艾林蓝色混合在 100 mL 的 0.1 N 磷酸钾三基 (K3PO4)。在准备上涂上一滴青联蓝,在花粉管生长过程中染色钙化物沉积。
- 使用 340-380 带通和 425 长通滤波器,通过具有紫外线外皮的显微镜观察沿样式的花粉管。
2. 脱氧核糖核酸提取
- 在春季的田间中取样2-3片。建议在幼年阶段对叶子进行取样,因为获得的DNA与旧叶相比,酚类化合物的质量更高,水平更低。
- 按照商用套件中描述的步骤提取基因组DNA(参见材料表)。
- 使用 UV-vis 分光光光光度计 (260 nm) 分析 DNA 浓度的数量和质量。
3. S-等位基因识别
-
设置 PCR 反应
- 在每个DNA提取样品的蒸馏水中制备50纳克/μL稀释剂。
- 慢慢解冻PCR试剂,并把它们保存在冰上。将DNA聚合酶留在冰箱中,直到需要为止。
- 使用引素的不同组合准备扩增反应。通过组合表 1中的组件创建 PCR 反应组合。涡旋PCR反应混合良好,并将PCR板每个孔中用于不同底漆组合的体积分配。然后,在每个井中加入1μL的DNA稀释。
- 将 PCR 板放在热循环器中,并运行表 1所示的相应 PCR 程序。
- 分析放大的片段。分析PCR放大片段主要有两种不同的方法:毛细管电泳(CE)与荧光标记的底漆或作为无标签底漆的琼脂凝胶电泳的可视化放大器。
-
毛细血管电泳
- 要准备装载缓冲液,请将 35 μL 的去离子化形式酰胺与 0.45 μL 的标签尺寸标准混合。将试剂搅拌好,然后将35.5μL分配到读卡板的孔中。
- 将 PCR 产品的 1 μL 添加到井中。此外,添加一滴矿物油,以防止水蒸发。
- 准备添加分离缓冲液的分离板。
- 使用基因分析仪附带的商业软件(参见材料表)。创建新的样品板,并保存板上所有孔的样品名称。
- 选择分析方法。在这种情况下,在90°C下对样品进行120 s变性,在2.0 kV下注射30秒,在6.0 kV下分离35分钟。
- 将两个板插入基因分析仪。用蒸馏水填充毛细管阵列。
- 加载专利的线性聚丙烯酰胺 (LPA) 凝胶。最后,单击"运行"。
-
凝胶电泳
- 制备1%的琼脂胶凝胶,在150 mL的1x TAE(三酸-醋酸-EDTA)电泳跑步缓冲液(40 mM Tris,20 mM醋酸,和1 mM EDTA在pH 8.0)中加入1.5克分子生物学级阿加松。通过微波加热溶解琼脂,2-3 分钟。
- 要可视化DNA,请加入4μL的核酸染色(参见材料表),然后轻轻混合。
- 将凝胶梳子与足够的孔用于梯子、控制和样品,放入凝胶托盘中。然后,将混合物缓慢倒入凝胶托盘中间,避免气泡。
- 让凝胶在室温下冷却 30-45 分钟,直到凝胶完全凝固。将凝胶引入电泳室,取出凝胶梳子,在腔室中充满足够的 1x TAE 缓冲液来盖住凝胶。
注:检查凝胶的位置。由于负电荷的DNA向阴极迁移,油井应放置在负极附近。 - 在 PCR 产品中加入 5 μL 的加载缓冲液(0.1%(v/v)溴酚蓝),并混合好。
- 要估计波段的大小,加载5μL的DNA分子量阶梯(参见材料表)。
- 将样品装入凝胶的附加井中。
- 一旦所有样品和DNA分子量阶梯被加载,在90V下运行凝胶1-1.5小时,直到蓝色染料线大约是凝胶长度的75%。
- 在转射器中可视化核酸的频带。
-
毛细血管电泳
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
杏的授粉研究要求在麻醉前一天在晚期气球阶段使用花(图1A)。这个阶段被认为是最有利的花粉和皮酸收集,因为花卉结构几乎成熟,但蚂蚁脱发尚未发生。这可以防止不想要的花粉的干扰,不仅来自同一朵花的花粉,而且来自其他花,因为封闭的花瓣会阻碍携带外部花粉的昆虫的到来。花粉颗粒很容易通过细网(图1B)从以前在室温下或稍加热量的纸张上放置24小时的脱黄醚中筛出。同样,在花瓣、花瓣和雄鹿的去除后,在钳子或指甲的帮助下,从气球阶段的花中获取皮球(图1C)。Pistils 可以使用细刷进行自发和交叉授粉(图 1D)。
杏花有五朵深红色的花瓣,五个白色的花瓣(图1A),一个皮瓣(图2A)和25-30个花瓣。皮斯蒂尔有三个主要结构:耻辱,风格和卵巢。卵巢有两个卵管,其中至少一个用于水果的受精。在授粉过程中,昆虫,主要是蜜蜂,将花粉颗粒转移到污名(图1A),在授粉后24小时内发芽(图2B)。花粉管由每个发芽的花粉颗粒产生,花粉颗粒通过皮质结构生长,在3-4天后到达卵巢,并在大约7天后受精两个卵子之一。在自不相容的品种中,花粉颗粒的S等位基因与两种皮层之一相同,花粉管停止在上部生长,防止受精(图2C)。然而,来自兼容的品种的花粉管,具有不同的S等位基因,可以通过这种风格(图2D)生长,到达卵巢(图2E),并受精两个卵子之一。
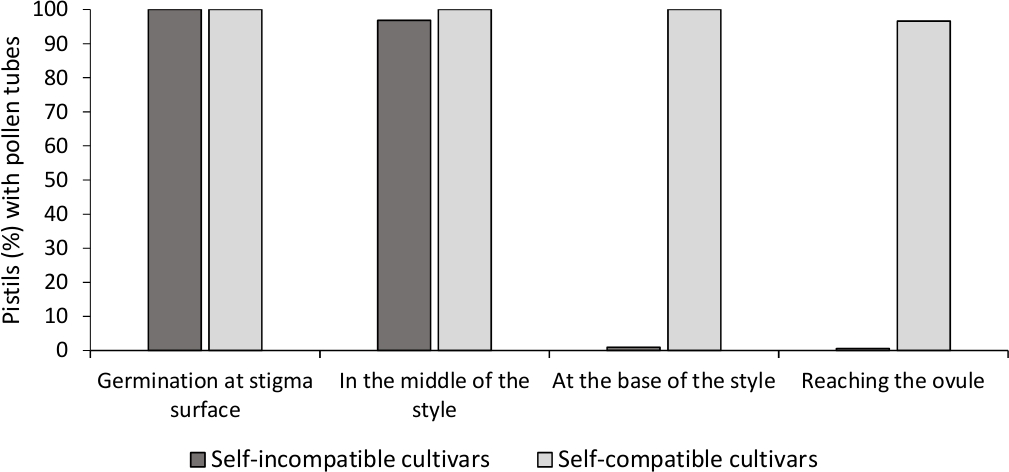
对体外花粉发芽的分析表明,这里分析的所有品种中花粉活力良好,因为大多数花粉管在培养基中比24小时后花粉粒的长度要长。在所有授粉的皮质中,在污名表面观察到发芽的花粉颗粒(图2B),表明授粉充足(图3)。
为了确定每个品种的自(in)相容性,在实验室控制条件下观察到在实验室控制条件下进行的自发和交叉授粉中的花粉管行为。花粉管生长记录沿风格在所有研究的皮斯蒂。当花粉管生长在大多数自授的皮层(图2C,图3)中沿样式被检查时,培养素被认为是自我不兼容的,当至少一个花粉管到达大多数研究的皮层中,一个花粉管达到样式的基础时,则自我兼容(图E,E图3)。
通过PCR分析对S-locus的研究允许对每种品种的S-基因型进行特征化。首先,S-alleles是通过使用引漆SRc-F/SRc-R(表2)放大第一个S-RNase内特罗来识别的。通过毛细血管电泳分析放大片段的大小(图4A),用于对其相应的不兼容组(I.G.)中分析的基因型进行分类。(表3)。
一些等位对,如S S1和S77或S66和S9,显示类似的片段大小为第一个intron。 S因此,这些等位元的分化是通过用引漆普鲁-C2/普鲁C4R、SHLM1/SHLM2和SHLM3/SHLM4放大RNase第二个内锥的一个区域来实现的(表2)。普鲁C2/PruC4R底漆组合用于区分S6和S9。对于S6,1300bp的碎片被放大,而6S9等位基因则观察到约700bpS的碎片(图4B,表3)。特定的引源SHLM1/SHLM2和SHLM3/SHLM4放大了S1等位距中约650bp的碎片和S7等位基因中的413bp碎片(图4C,C表3)。
用于放大SFB基因V2和HVb变量区域的引底器AprFBC8-(F/R)用于区分Sc和S88等位基因,因为两个等位基因都显示相同的RNase序列。S8等位基因显示PCR片段约150bp,而500bp片段对应于Sc等位元(图4D,表3)。一旦S-基因型被确定为所有品种,自我不兼容的培养体被分配到其相应的不相容组基于他们的S-等位基因(表3)。
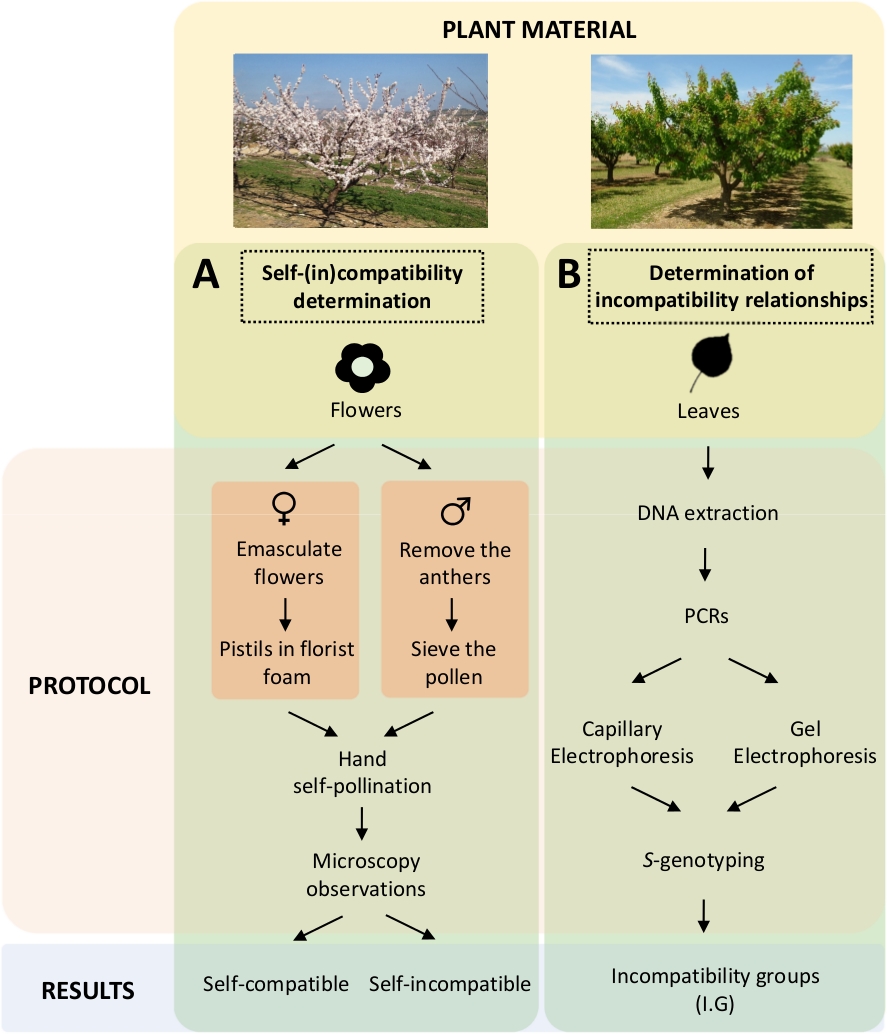
这种方法要求通过实验室中的受控自发和交叉授粉来确定每个品种的自(in)相容性(图5A),同时通过基因分析对S-基因型进行表征(图5B)。因此,可以确定每个品种的授粉要求和杏培养的不相容关系。

图 1.用于测定杏的自(in)兼容性的实验设置。
(A) 气球级的花(黑色箭头)在田野里。(B) 使用细网筛花粉粒。(C) 放在花花泡沫上在水中的皮斯蒂尔。(D) 在画笔的帮助下,对皮笔进行手授。请点击此处查看此图形的较大版本。

图 2.杏花中博弈性不相容关系的图表示。
(A) 在游戏性自我不相容 (GSI) 中, 相容和不兼容的花粉颗粒在污名上发芽.花粉颗粒携带原始基因型的两个S-等位基因之一,在这种情况下,S1或S2。 S如果花粉颗粒的S-等位基因与皮丝的两个SS-allee之一匹配,在这种情况下S1S3,花粉管的生长在样式的上三分之一中被抑制。(B) 花粉颗粒在污名表面的发芽。(C) 花粉管在风格上被捕,表明行为不相容。(D) 花粉管沿样式生长.(E) 在样式底部的花粉管指示兼容的行为。刻度柱,100 μm。请点击此处查看此图形的较大版本。

图 3.具有代表性的结果花粉发芽和花粉管生长通过风格自我兼容和自我不相容的培养体后自我授粉。
花粉颗粒在污名表面发芽的皮晶百分比,花粉管在半边形,在风格底部,并到达卵子。请点击此处查看此图形的较大版本。

图 4.PCR 片段放大使用五个引底对组合来识别SS-alleles。
(A) SRc-(F/R) 引素的基因分析仪输出显示RNase第一个与 S-alle 相对体对应S的内特罗区域的两个放大片段的大小。(B) PCR 放大使用引漆 PruC2/PruC4R 来识别S6和S9等位元。(C) PCR 产品使用特定的引物 SHLM1 和 SHLM2 进行S1等位基因和 SHLM3 和 SHLM4 的区分,以区分S7等位基因。(D) PCR 放大与四月FBC8-(F/R) 引物,用于识别Sc和S8等位物。MI: 1 kb DNA 梯子。MII: 100 bp DNA 梯子.请点击此处查看此图形的较大版本。

图 5.实验设计方案,阐明杏培养剂中自和(内)的兼容性关系。
(A) 通过实验室中受控授粉进行自我(in)兼容性测定的工作流程。(B) 通过分子方法进行S-等位基因识别的工作流程。请点击此处查看此图形的较大版本。
| PCR 主混合 | 热循环器条件 | ||||||
| 组件 | 最终浓度 | 15 μL 反应 | 循环步骤 | 温度 | 时间 | 周期 | |
| 10x NH4反应缓冲器 | 10 倍 | 1.5 μL | 初始变性 | 94 °C | 3分钟 | 1 | |
| 50 mM MgCl2解决方案 | 25 mM | 1.2 μL | 变性 | 94 °C | 1 分钟 | 35 | |
| 100 mM dNTP | 2.5 mM | 0.6 μL | 退火 | 55 °C | 1 分钟 | ||
| 底漆 SRc-F | 10 μM | 0.6 μL | 扩展 | 72 °C | 3分钟 | ||
| 引漆 SRc-R | 10 μM | 0.6 μL | 最终扩展 | 72 °C | 5分钟 | 1 | |
| 500 U 塔克 DNA 聚合酶 | 0.5 U | 0.2 μL | 4 °C | 保持 | |||
| H2O | 8.3 μL | ||||||
| 组件 | 最终浓度 | 25 μL 反应 | 循环步骤 | 温度 | 时间 | 周期 | |
| 10 倍 PCR 缓冲器 | 10 倍 | 2.5 μL | 初始变性 | 94 °C | 2分钟 | 1 | |
| 5x Q 解决方案 | 5 倍 | 5 μL | 变性 | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2.5 mM | 0.5 μL | 退火 | 55 °C | 2分钟 | ||
| 引漆普鲁C2 | 10 μM | 0.2 μL | 扩展 | 68 °C | 2分钟 | ||
| 引漆 C4R | 10 μM | 0.2 μL | 变性 | 94 °C | 10 s | 25 | |
| 250 U 塔克 DNA 聚合酶 | 10 U | 0.13 μL | 退火 | 58 °C | 2分钟 | ||
| H2O | 15.5 μL | 扩展* | 68 °C | 2分钟 | |||
| 最终扩展 | 72 °C | 5分钟 | 1 | ||||
| 4 °C | 保持 | ||||||
| * 每个周期都添加 10 s 到 68 %C 扩展步骤。 | |||||||
| 组件 | 最终浓度 | 25 μL 反应 | 循环步骤 | 温度 | 时间 | 周期 | |
| 10 倍 PCR 缓冲器 | 10 倍 | 2.5 μL | 初始变性 | 94 °C | 2分钟 | 1 | |
| 5x Q 解决方案 | 5 倍 | 5 μL | 变性 | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2.5 mM | 0.5 μL | 退火 | 62 °C | 1.5 分钟 | ||
| 引漆 SHLM1 | 10 μM | 0.2 μL | 扩展 | 72 °C | 2分钟 | ||
| 引漆 SHLM2 | 10 μM | 0.2 μL | 最终扩展 | 72 °C | 5分钟 | 1 | |
| 250 U 塔克 DNA 聚合酶 | 10 U | 0.13 μL | 4 °C | 保持 | |||
| H2O | 15.5 μL | ||||||
| 组件 | 最终浓度 | 20 μL 反应 | 循环步骤 | 温度 | 时间 | 周期 | |
| 5x PCR 缓冲器 | 5 倍 | 4 μL | 初始变性 | 98 °C | 30 s | 1 | |
| dNTP | 2.5 mM | 1.6 μL | 变性 | 98 °C | 10 s | 35 | |
| 引漆 SHLM3 | 10 μM | 1 μL | 退火 | 51 °C | 30 s | ||
| 引漆 SHLM4 | 10 μM | 1 μL | 扩展 | 72 °C | 1 分钟 | ||
| 100 U DNA 聚合酶 | 5 U | 0.2 μL | 最终扩展 | 72 °C | 5分钟 | 1 | |
| H2O | 12.4 μL | 4 °C | 保持 | ||||
| 组件 | 最终浓度 | 25 μL 反应 | 循环步骤 | 温度 | 时间 | 周期 | |
| 10 倍 PCR 缓冲器 | 10 倍 | 2.5 μL | 初始变性 | 94 °C | 2分钟 | 1 | |
| 100 mM dNTP | 2.5 mM | 2 μL | 变性 | 94 °C | 30 s | 35 | |
| 底漆 FBC8-F | 10 μM | 1 μL | 退火 | 55 °C | 1.5 分钟 | ||
| 底漆 FBC8-R | 10 μM | 1 μL | 扩展 | 72 °C | 2分钟 | ||
| 250 U 塔克 DNA 聚合酶 | 10 U | 0.125 μL | 最终扩展 | 72 °C | 5分钟 | 1 | |
| H2O | 17.4 μL | 4 °C | 保持 | ||||
表1.该协议中使用的不同引素组合的反应和循环条件。
| 引 | 序列 | 参考 |
| SRc-F | 5'-CTCTTTTTTTTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTTTCCCCTG-3' | 18 |
| 普鲁-C2 | 5'-CTGGCCAGTATTAAACC-3' | 35 |
| 普鲁-C4R | 5'-格格特格塔加加CG-3' | 35 |
| SHLM1-F | 5'-GGGGAGGTGATATAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTCTAAGCTG-3' | 17 |
| SHLM3-F | 5'-TATCTCTCT-3' | 17 |
| SHLM4-R | 5'-仙人掌GTGTGTG-3' | 17 |
| 四月FBC8-F | 5'-CATGGAAAGCTTTGG-3' | 26 |
| 四月FBC8-R | 5'-GCCTCTCTCTCTCTCT-3' | 26 |
表2.本议定书中使用的引物,序列和参考S-基因型表征在普鲁努斯亚美尼亚卡。
| 品种 | SRc-(F/R)(bp) | 普鲁C2/普鲁C4R(bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | 4月FBC8-(F/R)(bp) | S-基因型 | 不兼容组 (I.G) |
| 奇迹婴儿床8 | 420, 420 | 749, 1386 | S6S9 | Ⅷ | |||
| 魔法婴儿床8 | 334, 420 | 749 | S2S9 | Xx | |||
| 黄金罢工8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ⅰ | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Ⅳ | |||
| 库珀·科特8 | 274, 408 | 650 | S1S3 | 十八 | |||
| 阿普里昆 | 358, 358 | 500 | Sc Sc | - | |||
| 贝尔科特8 | 334, 358 | 500 | S2Sc | - | |||
| 春天模糊8 | 274, 358 | 150 | S3S8 | 二十一 |
表3.S-基因化杏培养素与五个引素对在该协议和不相容组分配。表中显示了使用SRc-(F/R)、普鲁C2/普鲁C4R、SHLM1/SHLM2、SHLM3/SHLM4和四月FBC8-(F/R)底漆放大S-对位物的不同聚合酶链反应产物尺寸。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
传统上,大多数商业杏欧洲品种是自我兼容的36。然而,在过去几十年中,在繁殖项目中,使用北美自我不兼容的培养品种作为父母,导致越来越多的新的自我不兼容的培养品种的释放,这些培养动物的授粉要求为77、8、37。8,37因此,确定杏培养素中的自兼容关系和相互兼容关系变得越来越重要。这在冬寒减少的地区尤为突出,因为开花时间的年复一年的变化在许多情况下阻止了品种及其花粉的开花的巧合,特别是在具有高寒寒要求的品种中38。本文所述的方法,结合手授粉,显微镜和基因分析,对于确定每个品种的自我(在)兼容性,并确定其潜在的授粉品种非常有用。
授粉要求可通过果园条件田间控制实验确定11、39。11,然而,对包括气象不良条件在内的外部因素的阐述可能导致授粉失败10,这可能导致自我不相容的错误诊断。本文所述的方法允许通过显微镜观察实验室控制条件下的手授粉花的微检,从而更准确地评估自(in)相容性,避免环境影响。此外,这种方法允许分析数量较多的品种每年,因为只需要少量的花,而不是几个成年树,每个品种需要在实地实验40。
不兼容关系可以建立结合手授和显微镜14。然而,授粉只能在春季开花季节进行很短的时间,需要实验室附近的成年树木,因为收集的花的寿命很短。因此,每个季节可以通过受控手授分析的不相容关系数量非常低。S-locus编码的基因的表征S使得S-allele基因分型18,41,的基于SPCR的方法得以发展。41这种方法加速S-allele的识别,因为它不需要花,实验可以进行任何植物组织42。这延长了植物材料,通常是幼叶,可以收集43个。此外,叶子可以冻干或冷冻,以便分析可以在一年中的任何时间进行,不像授粉,只能在开花季节44。另一个好处是,叶子可以在进入开花年龄之前从幼树中收集,便于采集样品和提前获得结果45。
基因分析可以更好地区分自我不相容等位基因,因为它提供了放大片段大小21,46,46的精确结果。迄今为止,在杏,12、18、20、21、22、23、24,18,20,21,22,23,中已发现33个S-alleee,允许建立基于S-基因型S8、9、17、25、26、279,17的25,36,27个不相容组。248另一方面,这种方法的一个缺点是,相同范围大小或突变中的不同等位基因可以错误地识别为相同的等位基因。因此,Sc和S8等位基因对于RNase序列是相同的,但在Sc19的SFB基因中发现了358-bp的插入。同样,等位体S1和S7的第一个内高区域相同,使用引漆 SRc-F/SRc-R 无法区分。此外,还发现了一些同源性,如S6和S528或S20和S55,以及S7、S S13(EF062341)和SS4617,因为这些等位词在PCR扩增过程中部分测序或失败,因此需要进一步的工作来正确区分它们。 S
PCR分析和S-RNase测序足以通过识别S-alleles和在其相应的不相容S组88,17,26,2717,26,27中分配培养品种来建立不相容关系。然而,这种方法有防止确定特定杏培养品种的自(in)兼容性的局限性。自我相容性 (SC) 已与其他普鲁努斯物种47中的特定 S-alleles 相关,如杏仁(Sf)48、49,49或甜樱桃S(S4')450,,51。然而,在杏中,与SC21相关的Sc等位基因可以被错误地识别为S8,一种自不相容的等位基因19、22,,22以及可能与S位点无关的突变,因为M-locus12,52,,52授予SC。最近,M-locus已经使用SSR标记12基因型。因此,杏基因型SC的遗传鉴定需要进一步研究,为了避免因与S位点无关的因素而出错,在这项工作中,通过自授花的皮质对花粉管的行为进行了表征鉴定。
本文所述的方法将实验室条件下手工授粉的自(in)相容性与随后在荧光显微镜下受控自授液中花粉管行为的观察以及通过PCR分析识别S-基因型的方法相结合,可以确定杏仁培养剂的授粉要求。这为种植者和育种者提供了有价值的信息,因为它允许建立品种之间的不相容关系,以在新的果园的设计中选择合适的授粉器,以及选择合适的父母来设计杏育种计划的新十字架。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者没有什么可透露的。
Acknowledgments
这项研究由欧洲大学欧洲区域发展基金、欧洲联盟(AGL2016-77267-R)和AGL2015-74071-JIN部长资助;国家教育研究所和阿格拉里亚和阿利门塔研究所(RFP2015-00015-00,RTA2017-00003-00);戈比耶诺·德拉贡-欧洲社会基金、欧洲联盟(格鲁波·康鲁达多A12_17R)、生物潜水基金会和Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}