

Summary
Wir stellen eine Methodik vor, um die Bestäubungsanforderungen vonAprikosensorten (Prunus armeniaca L.) zu ermitteln, die die Bestimmung der Selbstverträglichkeit durch Fluoreszenzmikroskopie mit der Identifizierung des S-Genotyps durch PCR-Analyse kombinieren.
Abstract
Die Selbstinkompatibilität in Rosaceae wird durch ein Gametophytic Self-Incompatibility System (GSI) bestimmt, das hauptsächlich vom multiallelic locus S gesteuert wird. In Der Aprikosen-Aprikosen-Beziehung wird die Bestimmung von Selbst- und Inter-(In-)Kompatibilitätsverhältnissen immer wichtiger, da die Freisetzung einer bedeutenden Anzahl neuer Sorten zu einer Zunahme von Sorten mit unbekannten Bestäubungsanforderungen geführt hat. Hier beschreiben wir eine Methodik, die die Bestimmung der Selbstverträglichkeit durch Handbestäubungen und Mikroskopie mit der Identifizierung des S-Genotypsdurch PCR-Analyse kombiniert. Zur Selbst-(In-)Kompatibilitätsbestimmung wurden im Ballonstadium aus jeder Sorte Blumen aus jeder Sorte gesammelt, im Labor von Hand bestäubt, fixiert und mit Anilineblau für die Beobachtung des Pollenrohrverhaltens unter der Fluoreszenzmikroskopie gefärbt. Für die Herstellung von Inkompatibilitätsbeziehungen zwischen Sorten wurde DNA aus jeder Sorte Saus jungen Blättern extrahiert und S-Allele wurden von PCR identifiziert. Dieser Ansatz ermöglicht die Einrichtung von Inkompatibilitätsgruppen und die Aufklärung von Inkompatibilitätsbeziehungen zwischen Sorten, die eine wertvolle Information liefern, um geeignete Bestäubern bei der Gestaltung neuer Obstgärten zu wählen und geeignete Eltern in Zuchtprogrammen auszuwählen.
Introduction
Selbstinkompatibilität ist eine Strategie der Blühenden Pflanzen, um Selbstbestäubung zu verhindern und Outcrossing1zu fördern. In Rosaceae wird dieser Mechanismus durch ein Gametophytic Self-Incompatibility System (GSI) bestimmt, das hauptsächlich durch den multiallelic locus S2gesteuert wird. Im Stil kodiert das RNase-Gen den S-s-Tylar-Determinanten, eine RNase3, während ein F-Box-Protein, das die S-Pollen-Determinantebestimmt, durch das SFB-Gen 4kodifiziert wird. S-s Die Selbstinkompatibilitätsinteraktion erfolgt durch die Hemmung des Pollenrohrwachstums entlang des Stils, der die Befruchtung der Ovule5,6verhindert.
In Der Aprikosen-Aprikosen-Aprikosen-Erneuerung hat in den letzten zwei Jahrzehnten weltweit eine Sortenerneuerung stattgefunden7,8. Diese Einführung einer wichtigen Anzahl neuer Sorten, aus verschiedenen öffentlichen und privaten Zuchtprogrammen, hat zu einer Zunahme von Aprikosensorten mit unbekannten Bestäubungsanforderungen geführt8.
Verschiedene Methoden wurden verwendet, um den Bestäubungsbedarf in Aprikosen zu bestimmen. Im Feld kann die Selbstverträglichkeit durch kontrollierte Bestäubungen in Käfigbäumen oder in entkalkten Blüten festgestellt werden und anschließend den Prozentsatz der Fruchtmenge9,10,11,12. Darüber hinaus wurden kontrollierte Bestäubungen im Labor durch semi-in vivo Kultur von Blumen und Analyse des Pollenrohrverhaltens unter Fluoreszenzmikroskopie8,13,14,15,16,17durchgeführt. In jüngster Zeit haben molekulare Techniken wie PCR-Analyse und Sequenzierung die Charakterisierung von Inkompatibilitätsbeziehungen ermöglicht, die auf der Untersuchung der RNase- und SFB-Gene 18,19basieren. In Aprikosen wurden 33 S-Allele gemeldet (S1 bis S20, S22 bis S30, S52, S53, Sv, Sx), einschließlich eines Alleels im Zusammenhang mit Selbstkompatibilität (Sc)12,18,20,21,22,23,24. Bisher wurden 26 Inkompatibilitätsgruppen in dieser Art nach dem S-Genotyp8, S9,,17,25,26,27festgestellt. Sorten mit den gleichen SS-Allelen sind miteinander kompatibel, während Sorten mit mindestens einem anderen S-Allel, die in verschiedenen inkompatiblen Gruppen zugeordnet sind, miteinander kompatibel sind. S
Um die Bestäubungsanforderungen von Aprikosensorten zu definieren, beschreiben wir eine Methode, die die Bestimmung der Selbstverträglichkeit durch S-Fluoreszenzmikroskopie mit der Identifizierung des S-Genotyps durch PCR-Analyse in Aprikosensorten kombiniert. Dieser Ansatz ermöglicht die Etablierung von Inkompatibilitätsgruppen und die Aufklärung von Inkompatibilitätsbeziehungen zwischen Sorten.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Selbst-(In-)Kompatibilitätsbestimmung
- Probieren Sie die Blumen auf dem Feld. Es ist notwendig, die Blumen in ballonPhase zu sammeln (Abbildung 1A), entsprechend Stufe 58 auf der BBCH-Skala für Aprikosen28, um unerwünschte vorherige Bestäubung zu vermeiden.
-
Selbst- und Kreuzbestäubungen im Labor
- Entfernen Sie die Anther der Blumen im Ballonstadium und legen Sie sie auf ein Stück Papier, um bei Labortemperatur zu trocknen.
- Nach 24 h die Pollenkörner mit einem feinen Maschen (0,26 mm)(Abbildung 1B) absieben
- Entleeren Sie eine Gruppe von 30 Blüten in der gleichen Ballonstufe für jede Selbstbestäubung und Kreuzbestäubung und legen Sie die Stempel auf Floristschaum in Wasser bei Labortemperatur (Abbildung 1C).
- Hand bestäuben die Stempel mit Hilfe eines Pinsels mit Pollen aus Blüten der gleichen Sorte 24 h nach der Entstung. Darüber hinaus bestäuben Sie einen weiteren Satz von Stempeln jeder Sorte mit Pollen aus Blüten eines kompatiblen Bestäubern als Kontrolle (Abbildung 1D).
- Nach 72 h die Stempel in einer fixativen Lösung von Ethanol/Essigsäure (3:1) für mindestens 24 h bei 4 °C29fixieren. Dann entsorgen Sie das Fixativ und fügen Sie 75% Ethanol hinzu, um sicherzustellen, dass die Proben vollständig in die Lösung eingetaucht sind. Proben können in dieser Lösung bei 4 °C bis zum Gebrauchvon 8,17,30,31,32konserviert werden.
-
Bewertung der Tragfähigkeit von Pollen durch In-vitro-Pollenkeimung
- Zur Herstellung des Keimmediums Gewicht 25 g Saccharose, 0,075 g Borsäure (H3BO3) und 0,075 g Calciumnitrat (Ca(NO3)2) 33.
- Fügen Sie die Komponenten des Mediums in 250 ml destilliertem Wasser hinzu und lösen Sie sie vollständig auf.
- Erstarren Sie das Medium, das 2 g Agarose hinzufügt, und mischen Sie es durch Wirbeln.
- Überprüfen Sie den pH-Wert des Mediums mit einem pH-Messgerät und stellen Sie den Wert mit NaOH- oder HCl-Lösung auf 7,0 ein.
- Autoclave die Mischung, um das Medium zu sterilisieren.
- Nach dem Autoklavieren das Medium abkühlen und in einer sterilen laminaren Fließhaube in Petrischalen verteilen.
- Streuen Sie die Pollenkörner der gleichen Sorten, die für die kontrollierten Bestäubungen im erstarrten Pollenkeimungsmedium verwendet werden, und beobachten Sie sie nach 24 h6unter dem Mikroskop.
HINWEIS: Um die laminare Durchflusshaube zu sterilisieren, reinigen Sie die Oberfläche mit 70% Ethanol und schalten Sie die UV-Lampe während 10 min ein. - Die Petri-Gerichte bis zum Gebrauch im Kühlschrank bei 4 °C aufbewahren.
-
Mikroskopische Beobachtungen
- Die Stempel dreimal 1 h mit destilliertem Wasser waschen und bei 4 °C in 5% Natriumsulfit lassen. Nach 24 h, autoklavieren Sie sie bei 1 kg/cm2 während 10 min in Natriumsulfit, um das Gewebe zu erweichen34.
- Legen Sie die autoklavierten Stempel über eine Glasrutsche und entfernen Sie mit Hilfe eines Skalpells die Trichome um den Eierstock, um eine bessere Visualisierung der Pollenröhren zu erhalten. Dann die Stempel mit einem Deckglas zerquetschen.
- 0,1% (v/v) anilineblauer Fleck vorbereiten: 0,1 ml Aniinblau in 100 ml 0,1 N Kaliumphosphat-Tribasis (K3PO4) mischen Tragen Sie einen Tropfen Anilineblau über die Präparate auf, um Callose-Ablagerungen während des Pollenrohrwachstums zu färben.
- Beobachten Sie die Pollenröhren entlang des Stils durch ein Mikroskop mit UV-Epifluoreszenz mit 340-380 Bandpass und 425 Langpassfiltern.
2. DNA-Extraktion
- Probe 2-3 Blätter im Feld im Frühjahr. Es wird empfohlen, die Blätter in jungen Stadien zu beproben, da die erhaltene DNA von höherer Qualität und niedrigeren Konzentrationen von Phenolverbindungen im Vergleich zu alten Blättern ist.
- Extrahieren Sie Genomische DNA nach den schritten, die in einem handelsüblichen Kit beschrieben sind (siehe Tabelle der Materialien).
- Analysieren Sie die Quantität und Qualität der DNA-Konzentrationen mit dem UV-Vis-Spektrophotometer (260 nm).
3. SS-Allel-Identifikation
-
Einrichten der PCR-Reaktionen
- Bereiten Sie eine Verdünnung von 50 ng/L in destilliertem Wasser jeder DNA-Extraktionsprobe vor.
- Die PCR-Reagenzien langsam auftauen und auf Eis halten. Lassen Sie die DNA-Polymerase im Gefrierschrank, bis sie benötigt wird.
- Bereiten Sie die Amplifikationsreaktionen mit den verschiedenen Kombinationen von Primern vor. Erstellen Sie den PCR-Reaktionsmix, indem Sie die Komponenten in Tabelle 1kombinieren. Vortex die PCR-Reaktion gut mischen und verteilen Sie das Volumen für die verschiedenen Kombinationen von Primern auf jeden Brunnen der PCR-Platte angegeben. Fügen Sie dann in jedem Brunnen 1 L der DNA-Verdünnung hinzu.
- Legen Sie die PCR-Platte in den Thermocycler und führen Sie das entsprechende PCR-Programm aus, das in Tabelle 1dargestellt ist.
- Analysieren Sie die verstärkten Fragmente. Es gibt hauptsächlich zwei verschiedene Möglichkeiten, die PCR-verstärkten Fragmente zu analysieren: Kapillarelektrophorese (CE) mit fluoreszierend gekennzeichneten Primern oder als visualisierende Amplicons der Agarose-Gel-Elektrophorese mit nicht gekennzeichneten Primern.
-
Kapillarelektrophorese
- Zur Vorbereitung des Ladepuffers 35 l entionisiertes Formamid mit 0,45 l beschrifteten Größenstandard mischen. Das Reagenz gut zu mischen, und geben Sie dann 35,5 l in den Brunnen der Leseplatte.
- Fügen Sie 1 L des PCR-Produkts in den Brunnen ein. Darüber hinaus fügen Sie einen Tropfen Mineralöl hinzu, um eine Verdunstung von Wasser zu verhindern.
- Bereiten Sie die Trennplatte vor, die einen Trennpuffer hinzufügt.
- Verwenden Sie die kommerzielle Software, die im Genanalysator enthalten ist (siehe Tabelle der Materialien). Erstellen Sie eine neue Probenplatte und speichern Sie die Probennamen für alle Brunnen auf der Platte.
- Wählen Sie die Analysemethode aus. In diesem Fall denaturieren Sie die Proben bei 90 °C für 120 s, injizieren Bei 2,0 kV für 30 s und trennen Sie bei 6,0 kV für 35 min.
- Setzen Sie die beiden Platten in den Genanalysator ein. Füllen Sie das Kapillararray mit destilliertem Wasser.
- Laden Sie das patentierte lineare Polyacrylamid (LPA) Gel. Klicken Sie schließlich auf Ausführen.
-
Gelelektrophorese
- Bereiten Sie ein 1% Agarose-Gel zu, das 1,5 g molekularbiologische Sanarose in 150 ml 1x TAE (Tris-Acetat-EDTA) Elektrophorese-Laufpuffer (40 mM Tris, 20 mM Essigsäure und 1 mM EDTA bei pH 8,0) hinzufügt. Lösen Sie die Agarose durch Mikrowellenheizung für 2-3 min.
- Um die DNA zu visualisieren, fügen Sie 4 L eines Nukleinsäureflecks hinzu (siehe Tabelle der Materialien) und mischen Sie sanft.
- Fügen Sie einen Gelkamm mit ausreichenden Brunnen für Leitern, Steuerungen und Proben in ein Geltablett ein. Dann gießen Sie langsam die Mischung in die Mitte des Geltabletts und vermeiden Sie Blasen.
- Lassen Sie das Gel 30-45 min bei Raumtemperatur abkühlen, bis das Gel vollständig erstarrt ist. Führen Sie das Gel in die Elektrophoresekammer ein, entfernen Sie den Gelkamm und füllen Sie die Kammer mit genügend 1x TAE Puffer, um das Gel zu bedecken.
HINWEIS: Überprüfen Sie die Platzierung des Gels. Die Brunnen sollten in der Nähe des negativen Pols platziert werden, da negativ geladene DNA zur Kathode wandert. - Den PCR-Produkten 5 l Ladepuffer (0,1% (v/v) Bromophenolblau) hinzufügen und gut mischen.
- Um die Größe der Bänder zu schätzen, laden Sie 5 l DNA-Molekulargewichtsleiter (siehe Tabelle der Materialien).
- Die Proben in die zusätzlichen Brunnen des Gels laden.
- Sobald alle Proben und die DNA-Molekulargewichtsleiter geladen sind, führen Sie das Gel bei 90 V für 1-1,5 h, bis die blaue Farbstofflinie etwa 75% der Länge des Gels beträgt.
- Visualisieren Sie die Bänder in einem Transilluminator für Nukleinsäuren.
-
Kapillarelektrophorese
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Bestäubungsstudien an Aprikosen erfordern die Verwendung von Blumen im späten Ballonstadium einen Tag vor der Anthese (Abbildung 1A). Diese Phase gilt als die günstigste für Pollen und Stempel Sammlung, da florale Strukturen sind fast ausgereift, aber Anther Dehiscence hat noch nicht stattgefunden. Dies verhindert die Interferenz unerwünschter Pollen, nicht nur von Pollen aus der gleichen Blüte, sondern auch von anderen Blüten, da die geschlossenen Blütenblätter die Ankunft von Insekten behindern, die externe Pollen tragen. Die Pollenkörner lassen sich leicht durch ein feines Netz (Abbildung 1B) aus entlüften Anthern, die zuvor auf einem Blatt Papier für 24 h bei Raumtemperatur oder mit leichter Zusatzwärme platziert wurden, versieben werden. Ebenso werden Stempel aus Blumen im Ballonstadium nach dem Entfernen von Blütenblättern, Kelchblättern und Stamens mit Hilfe von Pinzetten oder Fingernägeln gewonnen (Abbildung 1C). Pistile können selbstbefruchtet und mit einem feinen Pinsel befruchtet werden (Abbildung 1D).
Die hermaphroditischen Blüten von Aprikosen haben fünf dunkelrote Kelchblätter, fünf weiße Blütenblätter (Abbildung 1A), einen einzelnen Stempel (Abbildung 2A) und 25-30 Stamens. Der Stempel hat drei Hauptstrukturen: Stigma, Stil und Eierstock. Der Eierstock hat zwei Eizellen, und die Befruchtung von mindestens einem von ihnen ist für die Fruchteinstellung erforderlich. Während der Bestäubung übertragen Insekten, hauptsächlich Bienen, Pollenkörner in das Stigma (Abbildung 1A), wo sie innerhalb von 24 h nach der Bestäubung keimen (Abbildung 2B). Aus jedem keimenden Pollenkorn wird ein Pollenrohr hergestellt, das durch die Stempelstrukturen wächst, um nach 3-4 Tagen den Eierstock zu erreichen und nach etwa 7 Tagen eine der beiden Eizellen zu düngen. Bei selbstinkompatiblen Sorten, in denen das S-Allel des Pollenkorns mit einem der beiden Stempel identisch ist, hört das Pollenrohr auf, im oberen Stil zu wachsen und verhindert die Befruchtung ( Abbildung2C). Die Pollenröhren einer kompatiblen Sorte mit einem anderen S-Allel können jedoch durch den Stil wachsen (Abbildung 2D), den Eierstock erreichen (Abbildung 2E) und eine der beiden Eizellen befruchten. S
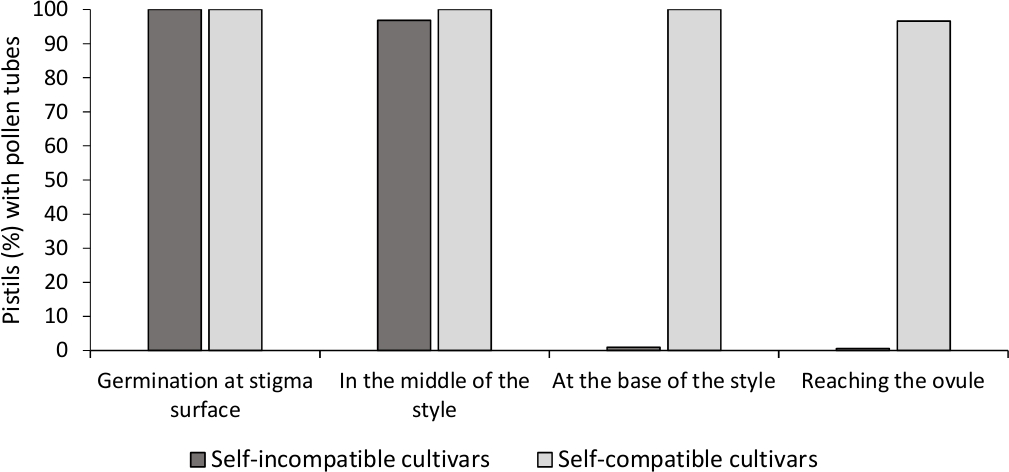
Die Analyse der In-vitro-Pollenkeimung zeigte eine gute Pollenlebensfähigkeit in allen hier analysierten Sorten, da die meisten Pollenröhren länger waren als die Länge des Pollenkorns nach 24 h im Kulturmedium. Gekeimte Pollenkörner wurden an der Stigmatisierungsoberfläche beobachtet (Abbildung 2B) in Stempeln aller Bestäubungen, was auf eine ausreichende Bestäubung hindeutet (Abbildung 3).
Um die Selbstverträglichkeit für jede Sorte zu bestimmen, wurde das Verhalten von Pollenröhren bei Selbst- und Kreuzbestäubungen unter laborgesteuerten Bedingungen unter Fluoreszenzmikroskopie beobachtet. Pollenrohrwachstum wurde entlang des Stils in allen untersuchten Stempeln aufgezeichnet. Kultivaren wurden als selbstkompatibel angesehen, wenn Pollenrohrwachstum entlang des Stils in den meisten selbstbefruchteten Stempeln (Abbildung 2C, Abbildung 3) und selbstkompatibel, wenn mindestens ein Pollenrohr die Basis des Stils in den meisten der untersuchten Stempel erreicht wurde ( Abbildung2E, Abbildung 3).
Die Untersuchung des S-Lokusdurch PCR-Analyse ermöglichte die Charakterisierung des S-Genotypsjeder Sorte. Zunächst wurden die S-Alleledurch die Verstärkung des ersten S-RNase-Introns mit den Primern SRc-F/SRc-R ( Tabelle2) identifiziert. Die Größe der verstärkten Fragmente wurde durch Kapillarelektrophorese analysiert (Abbildung 4A) und wurde verwendet, um die analysierten Genotypen in ihrer entsprechenden Inkompatibilitätsgruppe (I.G.) zu klassifizieren. (Tabelle 3).
Einige Allelepaare, wie S1 und S7 oder S6 und S9,zeigten ähnliche Fragmentgrößen für das erste Intron. So erfolgte die Differenzierung dieser Allele durch Verstärkung eines Bereichs des zweiten Introns der RNase mit den Primern Pru-C2/PruC4R, SHLM1/SHLM2 und SHLM3/SHLM4 (Tabelle 2). Die PruC2/PruC4R Primer-Kombination wurde verwendet, um zwischen S6 und S9zu unterscheiden. Für S6wurde ein Fragment von 1300 bp verstärkt, während für das S9-Allel ein Fragment von etwa 700 bp beobachtet wurde ( Abbildung4B, Tabelle 3). Die spezifischen Primer SHLM1/SHLM2 und SHLM3/SHLM4 verstärkten ein Fragment von etwa 650 bp im S1-Allel und 413 bp im S7-Allel ( Abbildung4C, Tabelle 3).
Die Primer AprFBC8-(F/R), die die V2- und HVb-Variablenbereiche des SFB-Gens verstärken, wurden verwendet, um Sc- und S8-Allele zu unterscheiden,da beide Allele identische RNase-Sequenz aufweisen. Das S 8-Allel8 zeigte ein PCR-Fragment von etwa 150 bp, während ein 500 bp-Fragment demS-C-Allel entsprach ( Abbildung4D, Tabelle 3). S Sobald der S-Genotyp für alle Sorten bestimmt wurde, wurden selbstinkompatible Sorten ihren entsprechenden Inkompatibilitätsgruppen auf der Grundlage ihrer S-Allelezugeordnet (Tabelle 3). S
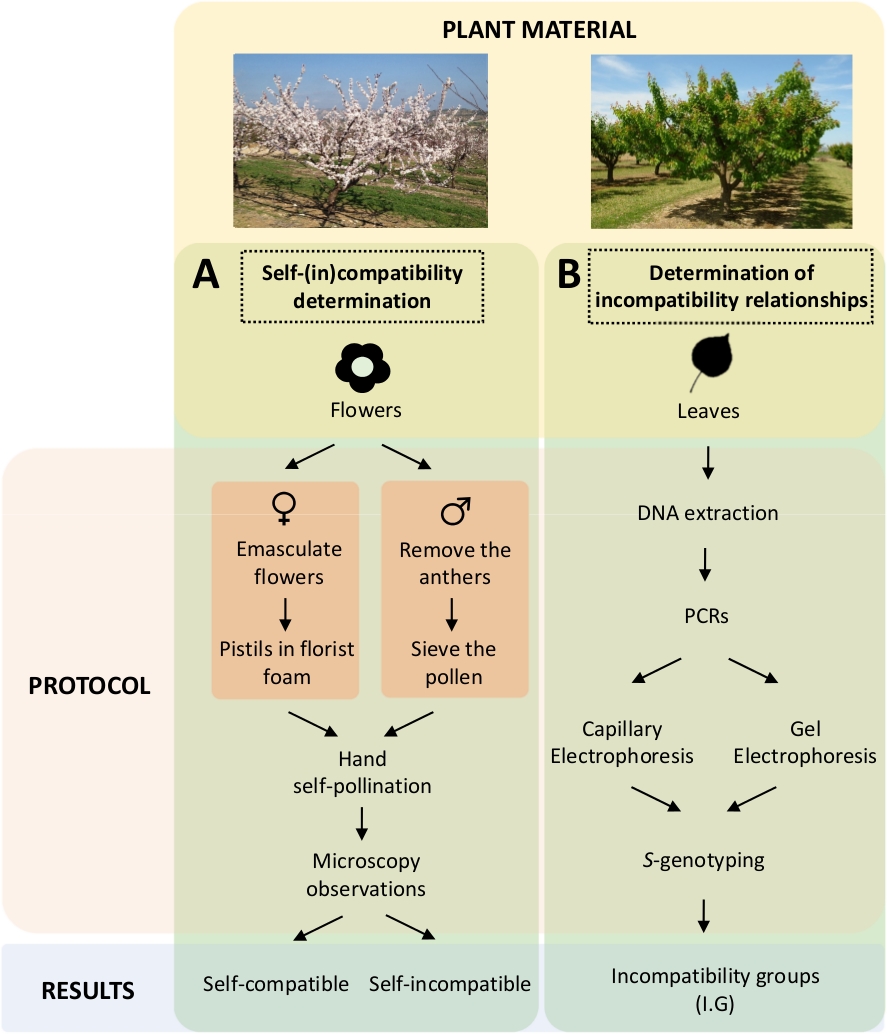
Dieser Ansatz erfordert die Bestimmung der Selbstverträglichkeit jeder Sorte durch kontrollierte Selbst- und Kreuzbestäubungen im Labor (Abbildung 5A) gleichzeitig mit der Charakterisierung des S-Genotyps durch genetische Analyse (Abbildung 5B). S Als Ergebnis können die Bestäubungsanforderungen jeder Sorte und die Inkompatibilitätsbeziehungen zwischen Aprikosensorten bestimmt werden.

Abbildung 1. Versuchsaufbau zur Bestimmung der Selbstverträglichkeit in Aprikosen.
(A) Blumen in der Ballonstufe (schwarze Pfeile) im Feld. (B) Sieb von Pollenkörnern mit einem feinen Maschen. (C) Pistils auf Floristenschaum in Wasser gelegt. (D) Handbestäubung der Stempel mit Hilfe eines Pinsels. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.

Abbildung 2. Schematische Darstellung von gametophytischen Inkompatibilitätsbeziehungen in Aprikosenblüten.
(A) In Gametophytic Self-Incompatibility (GSI) keimen sowohl kompatible als auch inkompatible Pollenkörner auf dem Stigma. Das Pollenkorn trägt eines von zwei S-Allelendes ursprünglichen Genotyps, in diesem Fall entweder S1 oder S2. Entspricht das S-Alleldes Pollenkorns einem der beiden S-Alleledes Stempels, in diesem Fall S1S3, wird das Pollenrohrwachstum im oberen Drittel des Stils gehemmt. (B) Keimung von Pollenkörnern auf der Stigmatisierungsoberfläche. (C) Pollenrohr in dem Stil verhaftet, die ein inkompatibles Verhalten. (D) Pollenröhren, die entlang des Stils wachsen. (E) Pollenröhren an der Basis des Stils, die auf ein kompatibles Verhalten hinweisen. Schuppenstäbe, 100 m. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.

Abbildung 3. Repräsentative Ergebnisse der Pollenkeimung und Pollenrohrwachstum durch den Stil für selbstkompatible und selbstinkompatible Sorten nach Selbstbestäubung.
Prozentsatz der Stempel mit Pollenkörnern, die an der Stigmatisierungsoberfläche keimen, mit Pollenröhren in der Hälfte des Stils, an der Basis des Stils, und das Erreichen der Eizelle. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.

Abbildung 4. PCR-Fragmentverstärkung mit fünf Primerpaarkombinationen zur SIdentifizierung von S-Allelen.
(A) Gene-Analyzer-Ausgabe für die SRc-(F/R)-Primer, die die Größe der beiden verstärkten Fragmente der rNase ersten Intron-Region zeigen, die den S-Allelenentspricht. (B) PCR-Verstärkung mit den Primern PruC2/PruC4R zur Identifizierung derS6- und S9-Allele.9 (C) PCR-Produkte, die mit den spezifischen Primern SHLM1 und SHLM2 zur Differenzierung des S1-Alleels und SHLM3 und SHLM4 zur Unterscheidung des S7-Alleels hergestellt wurden. (D) PCR-Verstärkung mit den AprFBC8-(F/R)-Primern zur Identifizierung von Sc- und S8-Allelen. MI: 1 kb DNA Leiter. MII: 100 bp DNA-Leiter. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.

Abbildung 5. Schema des experimentellen Designs zur Aufklärung der Selbst- und Inter-(in)-Kompatibilitätsbeziehungen in Aprikosensorten.
(A) Workflow der Selbst-(In-)Kompatibilitätsbestimmung durch kontrollierte Bestäubungen im Labor. (B) Workflow der S-Allel-Identifikationdurch molekulare Ansätze. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
| PCR Master Mix | Thermocycler-Bedingungen | ||||||
| Komponenten | Endgültige Konzentration | 15 L-Reaktion | Zyklusschritt | Temperatur | Zeit | Zyklen | |
| 10x NH4 Reaktionspuffer | 10x | 1,5 l | Anfängliche Denaturierung | 94 °C | 3 Min. | 1 | |
| 50 mM MgCl2 Lösung | 25 mM | 1,2 l | Denaturierung | 94 °C | 1 Min. | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 l | Glühen | 55 °C | 1 Min. | ||
| Primer SRc-F | 10 m | 0,6 l | Erweiterung | 72 °C | 3 Min. | ||
| Primer SRc-R | 10 m | 0,6 l | Endgültige Verlängerung | 72 °C | 5 Min. | 1 | |
| 500 U Taq DNA Polymerase | 0,5 U | 0,2 l | 4 °C | Halten | |||
| H2O | 8,3 l | ||||||
| Komponenten | Endgültige Konzentration | 25-L-Reaktion | Zyklusschritt | Temperatur | Zeit | Zyklen | |
| 10x PCR-Puffer | 10x | 2,5 l | Anfängliche Denaturierung | 94 °C | 2 Min. | 1 | |
| 5x Q-Lösung | 5-fach | 5 l | Denaturierung | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 l | Glühen | 55 °C | 2 Min. | ||
| Primer PruC2 | 10 m | 0,2 l | Erweiterung | 68 °C | 2 Min. | ||
| Primer C4R | 10 m | 0,2 l | Denaturierung | 94 °C | 10 s | 25 | |
| 250 U Taq DNA Polymerase | 10 U | 0,13 l | Glühen | 58 °C | 2 Min. | ||
| H2O | 15,5 l | Erweiterung* | 68 °C | 2 Min. | |||
| Endgültige Verlängerung | 72 °C | 5 Min. | 1 | ||||
| 4 °C | Halten | ||||||
| * mit 10 s, die jeden Zyklus zum 68 %C-Erweiterungsschritt hinzugefügt haben. | |||||||
| Komponenten | Endgültige Konzentration | 25-L-Reaktion | Zyklusschritt | Temperatur | Zeit | Zyklen | |
| 10x PCR-Puffer | 10x | 2,5 l | Anfängliche Denaturierung | 94 °C | 2 Min. | 1 | |
| 5x Q-Lösung | 5-fach | 5 l | Denaturierung | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 l | Glühen | 62 °C | 1.5 min | ||
| Primer SHLM1 | 10 m | 0,2 l | Erweiterung | 72 °C | 2 Min. | ||
| Primer SHLM2 | 10 m | 0,2 l | Endgültige Verlängerung | 72 °C | 5 Min. | 1 | |
| 250 U Taq DNA Polymerase | 10 U | 0,13 l | 4 °C | Halten | |||
| H2O | 15,5 l | ||||||
| Komponenten | Endgültige Konzentration | 20-L-Reaktion | Zyklusschritt | Temperatur | Zeit | Zyklen | |
| 5x PCR-Puffer | 5-fach | 4 l | Anfängliche Denaturierung | 98 °C | 30 s | 1 | |
| dNTP | 2,5 mM | 1,6 l | Denaturierung | 98 °C | 10 s | 35 | |
| Primer SHLM3 | 10 m | 1 L | Glühen | 51 °C | 30 s | ||
| Primer SHLM4 | 10 m | 1 L | Erweiterung | 72 °C | 1 Min. | ||
| 100 U DNA Polymerase | 5 U | 0,2 l | Endgültige Verlängerung | 72 °C | 5 Min. | 1 | |
| H2O | 12,4 l | 4 °C | Halten | ||||
| Komponenten | Endgültige Konzentration | 25-L-Reaktion | Zyklusschritt | Temperatur | Zeit | Zyklen | |
| 10x PCR-Puffer | 10x | 2,5 l | Anfängliche Denaturierung | 94 °C | 2 Min. | 1 | |
| 100 mM dNTP | 2,5 mM | 2 l | Denaturierung | 94 °C | 30 s | 35 | |
| Primer FBC8-F | 10 m | 1 L | Glühen | 55 °C | 1.5 min | ||
| Primer FBC8-R | 10 m | 1 L | Erweiterung | 72 °C | 2 Min. | ||
| 250 U Taq DNA Polymerase | 10 U | 0,125 l | Endgültige Verlängerung | 72 °C | 5 Min. | 1 | |
| H2O | 17,4 l | 4 °C | Halten | ||||
Tabelle 1. Reaktions- und Zyklusbedingungen für verschiedene Primerkombinationen, die in diesem Protokoll verwendet werden.
| Zündkapseln | Sequenz | Verweis |
| SRc-F | 5'-CTCTCTTTCCTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTCTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCTCTCTTAG-3' | 26 |
Tabelle 2. Primer, die in diesem Protokoll, dieser Sequenz und Referenz für die S-Genotypisierungin Prunus armeniacaverwendet werden.
| Sorte | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotyp | Inkompatibilitätsgruppe (I.G) |
| Wonder Kinderbett8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Magic Cot8 | 334, 420 | 749 | S2S9 | Xx | |||
| Goldschlag8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ⅰ | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| Cooper Kinderbett8 | 274, 408 | 650 | S1S3 | Xviii | |||
| Apriqueen | 358, 358 | 500 | ScSc | - | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | - | |||
| Frühling erröten8 | 274, 358 | 150 | S3S8 | Xxi |
Tabelle 3. S-genotypisierung von Aprikosensorten mit fünf Primerpaaren, die in diesem Protokoll verwendet werden, und Inkompatibilitätsgruppenzuweisung. Die verschiedenen Polymerase-Kettenreaktionsproduktgrößen von S-Allelen, verstärkt mit SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 und AprFBC8-(F/R)-Primern, sind in der Tabelle dargestellt. S
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Traditionell waren die meisten kommerziellen Aprikosen-Sorten in Europa selbstkompatibel36. Nichtsdestotrotz hat die Verwendung von nordamerikanischen selbstinkompatiblen Sorten als Eltern in Zuchtprogrammen in den letzten Jahrzehnten zur Freisetzung einer wachsenden Anzahl neuer selbstinkompatibler Sorten mit unbekannten Bestäubungsanforderungen7,8,37geführt. So wird die Bestimmung von Selbst- und Inter-(in)Kompatibilitätsverhältnissen in Aprikosensorten immer wichtiger. Dies wird in den Bereichen, in denen die Winterkälte abnimmt, noch verstärkt, da Schwankungen in der Blütezeit von Jahr zu Jahr in vielen Fällen den Zufall bei der Blüte von Sorten und deren Pollenisierern verhindern, insbesondere in Sorten mit hohem Kühlbedarf38. Die hier beschriebene Methode, die Handbestäubung, Mikroskopie und genetische Analysen kombiniert, war sehr nützlich, um die Selbstverträglichkeit jeder Sorte zu bestimmen und ihre potentiellen Bestäuizersorten zu ermitteln.
Die Bestäubungsanforderungen können durch Feldkontrollexperimente unter Obstplantagen11,39bestimmt werden. Die Exposition gegenüber externen Faktoren, einschließlich meteorologischer Widderbedingungen, kann jedoch zu Bestäubungsstörungen10führen, was zu falschen Diagnosen der Selbstinkompatibilität führen kann. Die hier beschriebene Methode ermöglicht es, die Selbstverträglichkeit durch Mikroskopiebeobachtungen von handbefruchteten Blüten unter laborgesteuerten Bedingungen genauer zu bewerten und dabei Umwelteinflüsse zu vermeiden. Darüber hinaus ermöglicht dieser Ansatz die Analyse einer höheren Anzahl von Sorten pro Jahr, da nur eine kleine Anzahl von Blumen anstelle von mehreren erwachsenen Bäumen für jede Sorte benötigt wird, die in Feldversuchen40benötigt werden.
Inkompatibilitätsbeziehungen können hergestellt werden, indem Handbestäubungen und Mikroskopie14kombiniert werden. Bestäubungen können jedoch nur für einen kurzen Zeitraum während der Blütezeit im Frühjahr durchgeführt werden, und erwachsene Bäume in der Nähe des Labors werden benötigt, da die Lebensdauer der gesammelten Blumen sehr kurz ist. Somit ist die Anzahl der Inkompatibilitätsbeziehungen, die durch kontrollierte Handbestäubungen in jeder Saison analysiert werden können, sehr gering. Die Charakterisierung der vom S-locuskodierten Gene hat die Entwicklung pcR-basierter Methoden für S-AlleleGenotypisierung18,41ermöglicht. Dieser Ansatz Sbeschleunigt die S-Allel-Identifikation, da es keine Blüten erfordert, und die Experimente können mit jedem vegetativen Gewebe42durchgeführt werden. Dies verlängert den Zeitraum, in dem Pflanzenmaterial, in der Regel junge Blätter, gesammelt werden kann43. Darüber hinaus können die Blätter lyophilisiert oder gefroren werden, so dass die Analyse zu jeder Zeit des Jahres durchgeführt werden kann, im Gegensatz zu Bestäubungen, die nur auf frischen Blüten während der Blütezeit44durchgeführt werden können. Ein weiterer Vorteil ist, dass Blätter von jungen Bäumen gesammelt werden können, noch bevor sie in das Blütealter eintreten, was die Entnahme von Proben und die frühe Erzielung von Ergebnissenerleichtert 45.
Die genetische Analyse ermöglicht eine bessere Differenzierung von Selbstinkompatibilitätsallelen, da sie präzise Ergebnisse der verstärkten Fragmentgrößen21,46liefert. Bis heute wurden 33 S-Allelein Aprikosen12,18,20,21,22,23,24, identifiziert, die erlaubt haben, 36 Inkompatibilitätsgruppen basierend auf S-Genotyp8,9,,17,25,26,27zu etablieren. Ein Nachteil dieser Methode besteht dagegen darin, dass verschiedene Allele in der gleichen Bereichsgröße oder Mutationen fälschlicherweise als dasselbe Allel identifiziert werden können. Somit sind Sc- und S 8-Allele8 für die RNase-Sequenz identisch, aber eine 358-bp-Einfügung findet sich im SFB-Gen von Sc19. Ebenso ist der erste Intron-Bereich der Allele S1 und S7 identisch und mit den Primern SRc-F/SRc-R nicht zu unterscheiden. Darüber hinaus wurden mehrere Homologien, wie S6 und S528 oder S20 und S55sowie S7, S13 (EF062341) und S4617, gefunden, weil einige dieser Allele teilweise sequenziert wurden oder durch Ausfälle während der PCR-Verstärkung, und folglich ist weitere Arbeit erforderlich, um sie richtig zu unterscheiden.
PCR-Analyse und S-RNase-Sequenzierung sind ausreichend, um Inkompatibilitätsbeziehungen durch die Identifizierung von S-Allelenund die Zuordnung von Sorten in ihrer entsprechenden Inkompatibilitätsgruppe8,17,26,27herzustellen. Diese Methode hat jedoch die Einschränkung, die Bestimmung der Selbstverträglichkeit für bestimmte Aprikosensorten zu verhindern. Selbstkompatibilität (SC) wurde mit bestimmten S-Allelenin anderen Prunus-Arten 47assoziiert, als Mandel (Sf)48,49 oder Süßkirsche (S4')50,51. Bei Aprikosen kann jedoch das Sc-Allel, das sc21zugeordnet wurde, fälschlicherweise als S8identifiziert werden, ein selbstinkompatibles Allel19,22und mögliche Mutationen, die nicht mit dem S-Ort verbunden sind, wie der M-Locus12,52, SC-Verleihende identifiziert wurden. Sc Kürzlich wurde Mder M-Locus mit SSR-Markern12genotypisiert. Daher bedarf die genetische Identifizierung von SC für Aprikosengenotypen weiterer Forschung, und S um Fehler aufgrund von Faktoren zu vermeiden, die nicht mit dem S-Lokus in Verbindung stehen, wurde in dieser Arbeit die Charakterisierung der Selbstverträglichkeit auch durch Phänotypisierung des Verhaltens der Pollenröhren durch den Stempel selbstbefruchtender Blüten bestimmt.
Die hier beschriebene Methode, die die Bestimmung der Selbstverträglichkeit durch Handbestäubungen unter Laborbedingungen mit der anschließenden Beobachtung des Verhaltens von Pollenröhren im Stempel kontrollierter Selbstbestäubungen unter der Fluoreszenzmikroskopie und der Identifizierung des S-Genotyps durch PCR-Analyse kombiniert, ermöglicht die Ermittlung des Bestäubungsbedarfs von Aprikosensorten. S Dies bietet eine wertvolle Information für Züchter und Züchter, da es ermöglicht, die Inkompatibilitätsbeziehungen zwischen Sorten herzustellen, um geeignete Bestäubern bei der Gestaltung neuer Obstgärten zu wählen sowie geeignete Eltern auszuwählen, um neue Kreuze in Aprikosenzuchtprogrammen zu entwerfen.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Die Autoren haben nichts zu verraten.
Acknowledgments
Diese Forschung wurde von Ministerio de Ciencia, Innovacién y Universidades-European Regional Development Fund, European Union (AGL2016-77267-R und AGL2015-74071-JIN) finanziert. Instituto Nacional de Investigacion y Tecnologa Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-00); Gobierno de Aragén-Europäischer Sozialfonds, Europäische Union (Grupo Consolidado A12_17R), Fundacion Biodiversidad und Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}