

Summary
אנו מציגים מתודולוגיה להקמת הצרכים של האבקה של משמש (שזיף armeniaca L.) המשלבים את התאימות העצמית (in) של מיקרוסקופ הקרינה הפלואורסצנטית עם זיהוי של ה-S-גנוטיפ על ידי ניתוח PCR.
Abstract
אי-סבילות עצמית ב ורדיים נקבעת על ידי מערכת התאמה עצמית של Gamet Tic (GSI) כי הוא נשלט בעיקר על ידי multiallelic לוקוס S. בשמש, הקביעה של יחסי התאימות העצמית והבין (in) היא חשובה יותר ויותר, שכן שחרורו של מספר חשוב של זנים חדשים הביא לגידול זנים עם דרישות האבקה לא ידועות. כאן, אנו מתארים מתודולוגיה המשלבת את ההגדרה של התאמה עצמית (בתוך) באמצעות המאביקים ומיקרוסקופ עם זיהוי של ה-S-גנוטיפ על ידי ניתוח PCR. בקביעת תאימות עצמית (in), הפרחים הנמצאים בשלב בלון מכל זנים נאספו בשדה, מואבקים בתוך המעבדה, קבועים ומוכתמים בכחול אנילין להתבוננות בהתנהגות שפופרת האבקה תחת המיקרוסקופיה הפלואורסצנטית. להקמת יחסי התאמה בין זנים, דנ א מכל זנים הופק מעלים צעירים ו-S-alleles זוהו על ידי ה-PCR. גישה זו מאפשרת להקים קבוצות אי-תאימות ולהבהיר קשרי אי-תאימות בין זנים, המספקים מידע רב ערך לבחירת המאביקים המתאימים בעיצוב מטעים חדשים ולבחירת הורים מתאימים בתוכניות רבייה.
Introduction
אי-סבילות עצמית היא אסטרטגיה של צמחים פורחים כדי למנוע האבקה עצמית ולקדם את המעבר1. ב ורדיים, מנגנון זה נקבע על ידי מערכת התאמה עצמית של Gamet Tic (GSI) כי הוא נשלט בעיקר על ידי multiallelic לוקוס S2. בסגנון, הגן rnase מקודד את דטרמיננטה s-sטיילר, rnase3, בעוד חלבון בקופסה F, אשר קובע את הדטרמיננטה של אבקה, הוא מסווג על ידי גן sfb 4. האינטראקציה עצמית אי-תאימות מתרחשת דרך עיכוב של צמיחת שפופרת אבקה לאורך הסגנון מניעת הפריה של הביצית5,6.
בשמש, התקיימה התחדשות בין-שנים ברחבי העולם בשני העשורים האחרונים7,8. הקדמה זו של מספר גדול של זנים חדשים, מתוכניות הרבייה הציבוריות והפרטיות השונות, הביאה לגידול זנים משמשים בעלי דרישות האבקה לא ידועות8.
שיטות שונות שימשו לקביעת דרישות ההאבקה בשמש. בתחום, ניתן ליצור תאימות עצמית (in) על-ידי המאביקים מבוקרים בעצי בכלוב או בפרחים מסורס ולאחר מכן מקליטים את אחוז הפרי שהוגדר9,10,11,12. בנוסף, המאביקים מבוקרים בוצעו במעבדה על ידי vivo למחצה ב תרבות של פרחים וניתוח של התנהגות שפופרת אבקה תחת מיקרוסקופ הזריחה8,13,14,15,16,17. לאחרונה, טכניקות מולקולריות, כגון ניתוח PCR ורצף, הרשו לאפיון של קשרי אי-תאימות המבוססים על חקר הגנים של rnase ו- sfb 18,19. בשמש, 33 S-alleles דווחו (s1 עד S20, s22 אל s30, s52, s53, sv, sx), כולל alleles אחד הקשורים בתאימות עצמית (Sc)12,18,20,21,22,23,24. עד עכשיו, 26 קבוצות אי-סבילות הפכו לאורווה במין זה על פי ה- S-גנוטיפ8,9,17,25,26,27. זנים עם אותו ס-אללים הם בלתי-תואמים, בעוד שהזנים עם שינוי s-alleles אחד לפחות, וכתוצאה מכך, מוקצים בקבוצות שונות שאינן תואמות, הן תואמות.
כדי להגדיר את דרישות ההאבקה של זנים משמש, אנו מתארים מתודולוגיה המשלבת את התאימות העצמית (בתוך) של מיקרוסקופ הקרינה הפלואורסצנטית עם זיהוי של ה-S-גנוטיפ על ידי ניתוח PCR בזנים משמש. גישה זו מאפשרת להקים קבוצות אי-תאימות ולהבהיר קשרי אי-תאימות בין זנים.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. קביעת תאימות עצמית (in)
- . לדגום את הפרחים בשדה יש צורך לאסוף את הפרחים בשלב הבלון (איור 1א), המתאים לשלב 58 בסולם bbch עבור משמש28, כדי למנוע האבקה קודמת לא רצויה.
-
המאביקים במעבדה
- הסירו את האנונים של הפרחים על במת בלון ומניחים אותם על פיסת נייר להתייבש בטמפרטורת המעבדה.
- לאחר 24 שעות, מסננת את גרגרי האבקה באמצעות רשת משובחת (0.26 מ"מ) (איור 1ב).
- מחליש קבוצה של 30 פרחים באותו שלב בלונים לכל האבקה עצמית והאבקה ומניחים את הpistils על קצף פרחים במים בטמפרטורת המעבדה (איור 1ג).
- יד המאביק את הpistils בעזרת מכחול עם אבקה מפרחים של זנים זהה 24 שעות אחרי אה. בנוסף, המאביק קבוצה אחרת של pistils של כל זנים עם אבקה מפרחים של מאביק תואם כפקד (איור 1ד).
- לאחר 72 h, לתקן את pistils בתמיסה קבע של אתנול/חומצה אצטית (3:1) לפחות 24 h ב 4 ° c29. ואז למחוק את הקבע ולהוסיף 75% אתנול להבטיח כי הדגימות הן לגמרי שקוע בפתרון. ניתן לשימור דוגמאות בתמיסה זו ב-4 ° c עד לשימוש ב-8,17,30,31,32.
-
הערכת הכדאיות של אבקה דרך נביטה של אבקה מחוץ לגופית
- כדי להכין את המדיום נביטה, משקל 25 גרם של סוכרוז, 0.075 g של חומצה בורית (H3בו3) ו 0.075 g של סידן חנקתי (Ca (NO3)2)33.
- הוסיפו את רכיבי המדיום ב-250 mL של מים מזוקקים והתמוססות לחלוטין.
- לחזק את המדיום הוספת 2 גרם של agarose ולערבב על ידי מתערבל.
- בדוק את ה-pH של המדיום באמצעות מד pH ולהתאים את הערך 7.0 עם פתרון NaOH או HCl.
- לחיטוי התערובת כדי לחטא את המדיום.
- לאחר הניקוי האוטומטי, הצנן את המדיום והפץ אותו לתוך מנות פטרי במכסה זרימה סטרילי.
- פזר את גרגרי האבקה של אותם זנים ששימשו להמאביקים הנשלטים במדיום נביטה של האבקה המתמצקת ולהתבונן בהם תחת המיקרוסקופ לאחר 24 שעות6.
הערה: כדי לעקר את כיסוי הזרימה המבינארי, נקה את המשטח עם 70% אתנול והפעל את מנורת ה-UV במהלך 10 דקות. - יש לאחסן את מנות פטרי במקרר ב-4 ° צ' עד השימוש.
-
תצפיות מיקרוסקופיה
- רוחצים את הpistils שלוש פעמים עבור 1 h עם מים מזוקקים ולהשאיר אותם ב 5% נתרן גופרית ב 4 ° c. לאחר 24 שעות, האוטובים אותם ב 1 ק"ג/cm2 במהלך 10 דקות ב נתרן גופרית כדי לרכך את הרקמות34.
- המקום pistils בלוק על שקופית זכוכית, בעזרת אזמל, להסיר את trichomes סביב השחלה כדי לקבל ויזואליזציה טובה יותר של שפופרות אבקה. ואז, למחוץ את הpistils עם זכוכית כיסוי.
- הכינו 0.1% (v/v) הכתם כחול אנילין: לערבב 0.1 mL של כחול אנילין ב 100 mL של 0.1 N אשלגן פוספט tribasic (K3הפו4). החל טיפה של כחול אנילין על פני ההכנות להכתים כתמים התצהירים במהלך צמיחת שפופרת אבקה.
- להתבונן את הצינורות אבקה לאורך הסגנון על ידי מיקרוסקופ עם האפיפלואורסצנטית UV באמצעות 340-380 bandpass ו 425 longpass מסננים.
2. הפקת דנ א
- לדוגמה 2-3 עלים בשדה באביב. מומלץ לדגום את העלים בשלבים צעירים מאז ה-DNA המתקבל הוא באיכות גבוהה יותר רמות נמוכות של תרכובות פנופילית לעומת עלים ישנים.
- לחלץ דנ א גנומית בעקבות השלבים המתוארים בערכה מסחרית זמין (ראה טבלת חומרים).
- לנתח את הכמות והאיכות של ריכוזי DNA באמצעות ספקטרוסקופיה UV-vis (260 nm).
3. S-allele זיהוי
-
הקמת תגובות ה-PCR
- הכינו דילול 50 ng/μL במים מזוקקים של כל דגימת ה-DNA.
- להפשיר את החומרים הPCR. לאט ולהשאיר אותם על הקרח תשאיר את הדנ א פולימראז. במקפיא עד שיהיה צורך
- הכינו את תגובות ההגברה באמצעות צירופים שונים של התחל. צור את המיקס של תגובת ה-PCR על-ידי שילוב הרכיבים בטבלה 1. וורטקס התגובה של ה-PCR מערבבים היטב ומפיצים את אמצעי האחסון המצוין עבור השילובים השונים של התחל לכל היותר בצלחת ה-PCR. לאחר מכן, הוסף 1 μL של דילול ה-DNA בכל באר.
- הציבו את צלחת ה-PCR בציקלוניר והפעל את תוכנית ה-PCR המתאימה בטבלה 1.
- לנתח את השברים מוגבר. ישנן בעיקר שתי דרכים שונות לנתח את החלקיקים המגבתים של ה-PCR: אלקטרופורזה קפינגית (לסה נ) עם התחל המסומנת בצבעי פלורסנט או כמגביר הדמיה של אלקטרופורזה בג ג'ל בצבעי היסוד שאינם מתויג.
-
אלקטרופורזה קפילר
- כדי להכין את מאגר הטעינה, לערבב 35 μL של הטופסשינוי עם 0.45 μL של התווית שינוי גודל סטנדרטי. מערבולת הכימית לערבב היטב, ולאחר מכן לוותר 35.5 μL לתוך הבאר של לוחית הקורא.
- הוסף 1 μL של מוצר ה-PCR לתוך הבאר. בנוסף, להוסיף טיפה של שמן מינרלי כדי למנוע אידוי מים.
- הכן את לוחית ההפרדה הוספת מאגר הפרדה.
- השתמש בתוכנה המסחרית הכלולה במנתח הגנים (ראה טבלת חומרים). צור צלחת מדגם חדש ולשמור את שמות לדוגמה עבור כל הבארות על הצלחת.
- בחר את שיטת הניתוח. במקרה זה, לשלב את הדגימות ב 90 ° c עבור 120 s, להזריק ב 2.0 kV עבור 30 s, ונפרד ב 6.0 kV עבור 35 min.
- הכנס את שני הצלחות. לתוך מנתח הגנים ממלאים את מערך נימי עם מים מזוקקים.
- העמיסו את ג'ל פוליאקרילמיד (LPA) הפטנט ליניארי. לבסוף, לחץ על הפעל.
-
ג'ל אלקטרופורזה
- הכן 1% agarose ג'ל הוספת 1.5 g של ביולוגיה מולקולרית כיתה agarose ב 150 מ ל של 1 x טאה (טריס-אצטט-EDTA) אלקטרופורזה במאגר (40 mM Tris, 20 מ"מ חומצה אצטית, ו 1 מ"מ EDTA ב-pH 8.0). מתמוסס את הצמח על ידי חימום מיקרוגל עבור 2-3 דקות.
- כדי להמחיש את ה-DNA, להוסיף 4 μL של כתם חומצה גרעין (ראה לוח חומרים) ולערבב בעדינות.
- הוסיפו מסרק ג'ל, עם בארות מספיקות לסולמות, שולטת ודוגמיות, למגש ג'ל. ואז, לשפוך לאט את התערובת לתוך באמצע מגש ג'ל ולהימנע בועות.
- תנו לג להתקרר למשך 30-45 דקות בטמפרטורת החדר עד שהג התחזק לחלוטין. להציג את הג בחדר האלקטרופורזה, להסיר את המסרק ג'ל ולמלא את התא עם מספיק 1x מאגר טה כדי לכסות את ג'ל.
הערה: בדוק את מיקום הג. יש להציב את הבארות קרוב לקוטב השלילי מאז שהוא מחויב לעבר הקתודה. - הוסף 5 μL של מאגר טעינה (0.1% (v/v) ברומאופנול כחול) למוצרי ה-PCR וערבבו היטב.
- כדי להעריך את גודל הלהקות, טען 5 μL של סולם משקל מולקולרי DNA (ראה טבלת חומרים).
- העמיסו את הדגימות לתוך בארות נוספות של ג'ל.
- לאחר כל הדגימות ואת הסולם דנ א מולקולרית משקל טעונים, להפעיל את הג ב 90 V עבור 1-1.5 h, עד קו הצבע הכחול הוא כ-75% אורך של ג'ל.
- דמיינו את הלהקות בתוך ממאיר לחומצות גרעין.
-
אלקטרופורזה קפילר
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
לימודי האבקה בשמש דורשים שימוש בפרחים בשלב הבלון המאוחר יום לפני התזה (איור 1א). שלב זה נחשב לחיובי ביותר עבור האוסף האבקה והפיסטיל, שכן מבנים פרחוניים הם כמעט בוגרים, אבל הדחיניות עדיין לא התרחשה. זה מונע הפרעה של אבקה בלתי רצויה, לא רק של אבקה מאותו פרח אלא גם מפרחים אחרים, מאז הכותרת הסגורה לעכב את הגעתו של חרקים נושאת אבקה חיצונית. גרגרי האבקה מלאים בקלות באמצעות רשת משובחת (איור 1ב) מביתההארחה, שהונח בעבר על פיסת נייר בטמפרטורה של 24 שעות בטמפרטורת החדר או בחום קל במיוחד. כמו כן, מתקבלים הpistils מפרחים בבמת בלונים לאחר הסרת עלי כותרת, עלי גביע ואבקנים בעזרת מלקחיים או ציפורניים (איור 1ג). Pistils יכול להיות self-מואבקים עם מברשת יפה (איור 1ד).
לפרחי המאדיסטיים של משמש יש חמישה עלי גביע אדומים כהים, חמישה עלי כותרת לבנים (איור 1א), פיסטיל אחד (איור 2א) ו-25-30 אבקנים. לפיסטיל יש שלושה מבנים עיקריים: סטיגמה, סגנון ושחלה. השחלה מקבלת שני מובנים, וההפריה של לפחות אחת מהן נדרשת לקביעת פרי. במהלך האבקה, חרקים, בעיקר דבורים, העברת גרגרי אבקה לסטיגמה (איור 1א), שם הם לנבוט (איור 2ב) תוך 24 שעות אחרי האבקה. שפופרת אבקה מופק כל גרגר אבקה הנבטי, אשר גדל באמצעות מבני פיסטיל להגיע השחלה לאחר 3-4 ימים ולהפרות אחד של שני שניהם לאחר סביב 7 ימים. בזנים שאינם בעלי התאמה עצמית שבה ה- אלל של גרגר האבקה זהה לאחד משני הpistils, צינור האבקה מפסיק לצמוח בסגנון העליון, ומונע הפריה (איור 2ג). עם זאת, צינורות אבקה של זנים תואמים, עם allele S שונים, יכול לגדול דרך הסגנון (איור 2ד), להגיע לשחלה (איור 2E) ולהפרות אחד של שני הבאלידו.
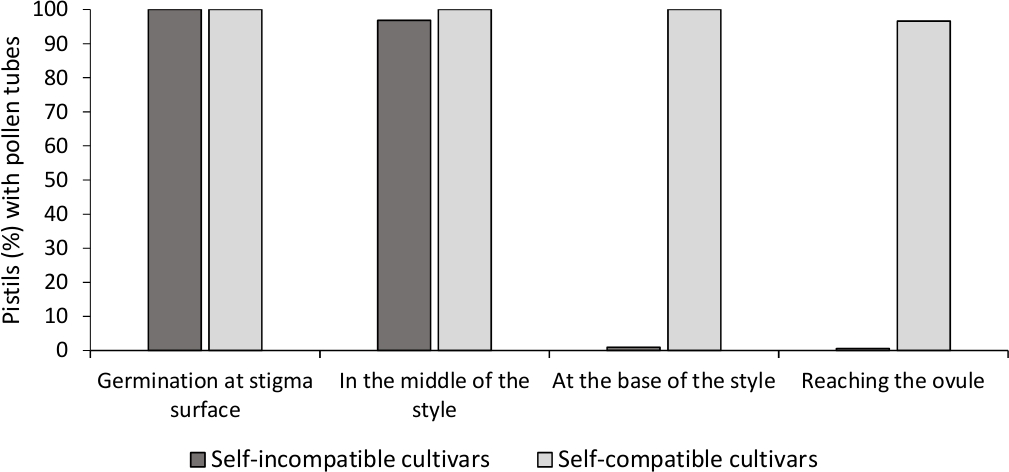
הניתוח של נביטת האבקה מחוץ לגוף הראה את הכדאיות הטובה של אבקה בכל הזנים שנותחו כאן, מאחר שצינורות האבקה של רוב הזמן היו ארוכים יותר מאורכם של גרגר אבקה לאחר 24 שעות במדיום התרבותי. גרגרי אבקה מונבטים נצפו במשטח הסטיגמה (איור 2ב) ב pistils מכל המאביקים, המציין האבקה נאותה (איור 3).
כדי לקבוע את התאימות העצמית (בתוך) עבור כל זנים, התנהגות צינור אבקה בתוך העצמי, ו-המאביקים נעשה בתנאים מבוקרת מעבדה נצפתה תחת מיקרוסקופ הזריחה. הצמיחה שפופרת אבקה נרשמה לאורך הסגנון בכל pistils בדק. זנים נחשבו בלתי תואמים עצמית כאשר הצמיחה שפופרת אבקה נעצרה לאורך הסגנון ברוב pistils מואבקים (איור 2C, איור 3) ותואם עצמית כאשר לפחות צינור אבקה אחד הגיע לבסיס הסגנון ברוב הpistils בדק (איור 2E, איור 3).
המחקר של ה -s-לוקוס ידי ניתוח PCR מותר לאפיון s-גנוטיפ של כל זנים. ראשית, s-alleles זוהו על ידי הגברה של הראשון s-rnase אינטרון באמצעות התחל SRc-F/src-R (שולחן 2). הגודל של שברי מוגבר נותחו על ידי אלקטרופורזה קפילר (איור 4א) ושימש לסווג את ה-גנוטיפים שנותחו בקבוצת אי התאימות המקבילה שלהם (I.G.) (שולחן 3).
זוגות של alleles כגון s1 ו -s7 או s6 ו -s9, הראו גדלי קטעים דומים עבור intron הראשון. כך, הבידול של אללים אלה נעשה על ידי הגברה על אזור של אינטרון השני של rnase עם התחל pru-C2/PruC4R, SHLM1/SHLM2 ו SHLM3/SHLM4 (שולחן 2). השילוב PruC2/PruC4R פריימר שימש להבחנה בין s6 ו -s9. עבור S6, קטע של 1300 bp היה מוגבר ואילו קטע של סביב 700 bp נצפתה עבור S9 אלל (איור 4b, שולחן 3). SHLM1 התחל הספציפי/SHLM2 ו SHLM3/SHLM4 הוגדל קטע של כ 650 bp ב s1 אלל ו 413 bp ב s7 אלל (איור 4ג, שולחן 3).
AprFBC8 התחל-(F/R) המגבירים את האזורים V2 ו-hvb משתנה של הגן sfb שימשו כדי להבחין sc ו -s8 אללים מאז שני האללים להציג רצף rnase זהה. אלל S8 הראו קטע PCR של כ 150 bp ואילו משבר bp 500 התכתב עםאלל שלc (איור 4D, שולחן 3). לאחר שנקבע את ה-s-גנוטיפ עבור כל הזנים, הזנים שאינם בעלי התאמה עצמית הוקצו לקבוצות התאימות המקבילות שלהם בהתבסס על ה-s-alleles שלהם (שולחן 3).
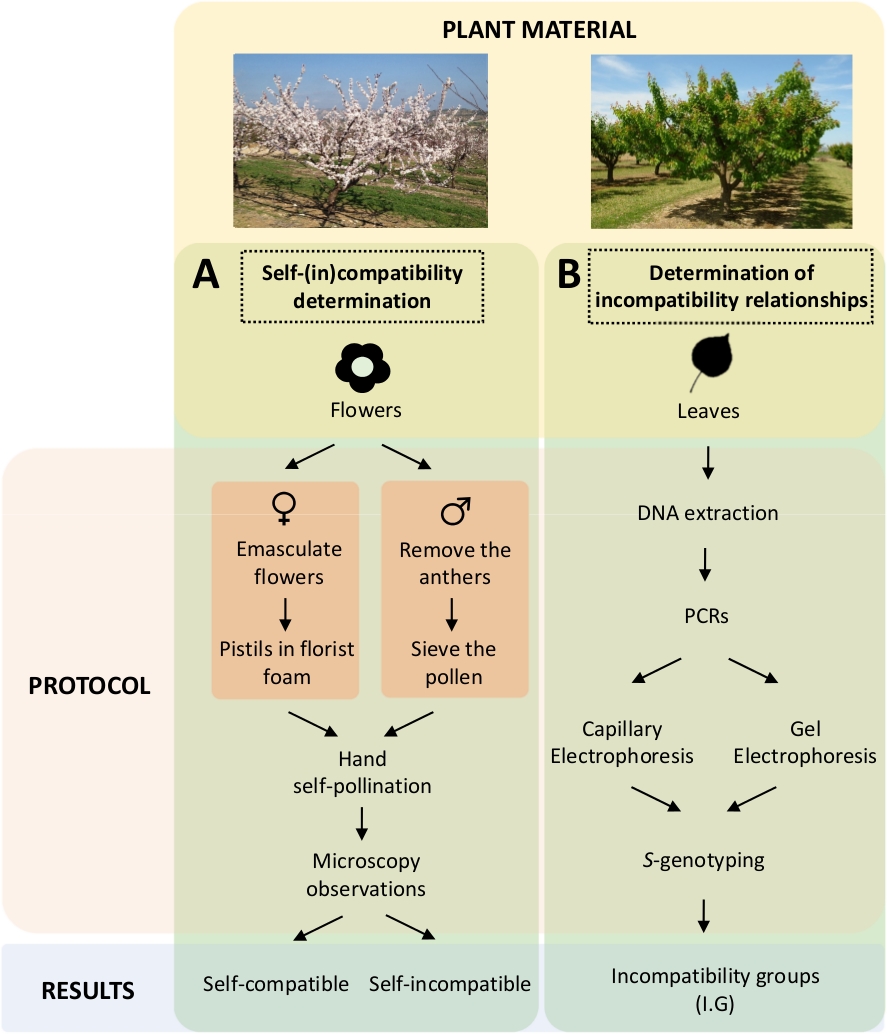
גישה זו מחייבת קביעת התאימות העצמית (בתוך) של כל אחד מהזנים על-ידי שליטה עצמית והצלבה במעבדה (איור 5א) במקביל לאפיון ה-S-גנוטיפ באמצעות ניתוח גנטי (איור 5ב'). כתוצאה מכך, ניתן לקבוע את דרישות ההאבקה של כל אחד מהזנים ואת היחסים בין אי-התאמה בין זנים משמש.

איור 1. ניסיוני מוגדר לקביעת תאימות עצמית (in) בשמש.
(א) פרחים בשלבי בלונים (חיצים שחורים) בשדה. (ב) הנפה של גרגרי אבקה בעזרת רשת משובחת. (ג) Pistils ממוקם על קצף פרחים במים. (ד) יד-האבקה של הpistils בעזרת מברשת צבע. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

. איור 2 ייצוג מערכת יחסים של אי-תאימות gamettic בפרחי משמש.
(א) בחוסר התאמה עצמית (gsi), גרגרי אבקה מתאימים ובלתי תואמים לנבוט על הסטיגמה. גרגר אבקה נושא אחד משניהצדדים של גנוטיפ המקורי, במקרה זה או1 או s2. אם S-allele של הדגן אבקה מתאים אחד של שני S-allele של pistil, במקרה זה s1S3, הצמיחה צינור אבקה מעוכבים בשליש העליון של הסגנון. (ב) נביטה של גרגרי אבקה על משטח הסטיגמה. (ג) שפופרת אבקה נעצרה בסגנון המציין התנהגות בלתי תואמת. (ד) שפופרות של אבקה הגדלים לאורך הסגנון. (ה) שפופרות אבקה בבסיס הסגנון המציין התנהגות תואמת. סרגלי קנה מידה, 100 μm. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

. איור 3 תוצאות מייצגות של נביטה של אבקה וצמיחת שפופרת אבקה דרך הסגנון לזנים בעלי תאימות עצמית ובלתי תואמת עצמית לאחר המאביקים עצמית.
אחוז הpistils עם גרגרי אבקה מונטים במשטח הסטיגמה, עם צינורות אבקה בחצי הדרך, בבסיס הסגנון, ומגיע לביצית. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

איור 4. הגברה של החלק האנטי-PCR באמצעות שילובי זוגות מפריימר לזיהוי ה-S-alleles.
(א) מנתחגנים התפוקה עבור SRc-(F/R) המראה הראשוני מראה את הגודל של שני שברי מוגבר של אזור אינטרון הראשון rnase המתאימים ל- S-alleles. (ב) PCR הגברה באמצעות הPruC2 התחל/PruC4R לזיהוי ה-s6 ו- s9 אלס. (ג) PCR מוצרים שהתקבלו באמצעות SHLM1 התחל הספציפי ו SHLM2 עבור הבידול של s1 אלל ו SHLM3 ו SHLM4 כדי להבחין s7 אלל. (ד) PCR הגברה עם AprFBC8-(F/R) התחל לזהות את ה-sc ו- s8 אלס. MI: סולם DNA 1 kb. MII: 100 הסולם DNA bp. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

. איור 5 תכנית התכנון הניסיוני להבהיר את יחסי התאימות העצמיים והבין-(in) בזנים של משמש.
(א) זרימת עבודה של קביעת תאימות עצמית (in) באמצעות המאביקים מבוקרים במעבדה. (ב) זרימת עבודה של זיהוי S-אלל באמצעות גישות מולקולריות. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
| PCR מיקס מאסטר | תנאי הציקלטרנר | ||||||
| רכיבים | ריכוז סופי | 15 תגובת μL | מחזור שלב | טמפרטורה | זמן | מחזורי | |
| 10x NH4 מאגר התגובות | 10x | 1.5 מיקרומטר | דנטורציה ראשונית | 94 ° c | 3 דקות | 1 | |
| 50 מ"מ פתרון MgCl2 | 25 ממ ' | 1.2 מיקרומטר | דנטור, הרפתקאות | 94 ° c | 1 דקות | 35 | |
| 100 מ"מ ממדי dNTP | 2.5 ממ ' | 0.6 מיקרומטר | ריפוי | 55 ° c | 1 דקות | ||
| פריימר SRc-F | 10 μM | 0.6 מיקרומטר | סיומת | 72 ° c | 3 דקות | ||
| פריימר SRc-R | 10 μM | 0.6 מיקרומטר | הארכה סופית | 72 ° c | 5 דקות | 1 | |
| 500 יו-אן-איי פלאז | 0.5 יו | 0.2 מיקרומטר | 4 מעלות צלזיוס | החזיק | |||
| H2O | 8.3 מיקרומטר | ||||||
| רכיבים | ריכוז סופי | התגובה ל -25 μL | מחזור שלב | טמפרטורה | זמן | מחזורי | |
| מאגר ה-PCR 10x | 10x | 2.5 מיקרומטר | דנטורציה ראשונית | 94 ° c | 2 דקות | 1 | |
| 5x Q-פתרון | 5x | 5 מיקרומטר | דנטור, הרפתקאות | 94 ° c | עשרה מנות | 10 | |
| 100 מ"מ ממדי dNTP | 2.5 ממ ' | 0.5 מיקרומטר | ריפוי | 55 ° c | 2 דקות | ||
| פריימר PruC2 | 10 μM | 0.2 מיקרומטר | סיומת | 68 ° c | 2 דקות | ||
| פריימר C4R | 10 μM | 0.2 מיקרומטר | דנטור, הרפתקאות | 94 ° c | עשרה מנות | 25 | |
| 250 יו-אן-איי פלאז | 10 יו | 0.13 מיקרומטר | ריפוי | 58 ° c | 2 דקות | ||
| H2O | 15.5 מיקרומטר | סיומת | 68 ° c | 2 דקות | |||
| הארכה סופית | 72 ° c | 5 דקות | 1 | ||||
| 4 מעלות צלזיוס | החזיק | ||||||
| * עם 10 s הוסיף כל מחזור לשלב 68% C הארכה. | |||||||
| רכיבים | ריכוז סופי | התגובה ל -25 μL | מחזור שלב | טמפרטורה | זמן | מחזורי | |
| מאגר ה-PCR 10x | 10x | 2.5 מיקרומטר | דנטורציה ראשונית | 94 ° c | 2 דקות | 1 | |
| 5x Q-פתרון | 5x | 5 מיקרומטר | דנטור, הרפתקאות | 94 ° c | שלושים בנות | 35 | |
| 100 מ"מ ממדי dNTP | 2.5 ממ ' | 0.5 מיקרומטר | ריפוי | 62 ° c | 1.5 דקות | ||
| פריימר SHLM1 | 10 μM | 0.2 מיקרומטר | סיומת | 72 ° c | 2 דקות | ||
| פריימר SHLM2 | 10 μM | 0.2 מיקרומטר | הארכה סופית | 72 ° c | 5 דקות | 1 | |
| 250 יו-אן-איי פלאז | 10 יו | 0.13 מיקרומטר | 4 מעלות צלזיוס | החזיק | |||
| H2O | 15.5 מיקרומטר | ||||||
| רכיבים | ריכוז סופי | 20 התגובה μL | מחזור שלב | טמפרטורה | זמן | מחזורי | |
| מאגר ה-PCR 5x | 5x | 4 מיקרומטר | דנטורציה ראשונית | 98 ° c | שלושים בנות | 1 | |
| מיכל הדntp | 2.5 ממ ' | 1.6 מיקרומטר | דנטור, הרפתקאות | 98 ° c | עשרה מנות | 35 | |
| פריימר SHLM3 | 10 μM | 1 ליטר | ריפוי | 51 ° c | שלושים בנות | ||
| פריימר SHLM4 | 10 μM | 1 ליטר | סיומת | 72 ° c | 1 דקות | ||
| 100 ה-DNA פולימראז | 5 ה, שמרו | 0.2 מיקרומטר | הארכה סופית | 72 ° c | 5 דקות | 1 | |
| H2O | 12.4 מיקרומטר | 4 מעלות צלזיוס | החזיק | ||||
| רכיבים | ריכוז סופי | התגובה ל -25 μL | מחזור שלב | טמפרטורה | זמן | מחזורי | |
| מאגר ה-PCR 10x | 10x | 2.5 מיקרומטר | דנטורציה ראשונית | 94 ° c | 2 דקות | 1 | |
| 100 מ"מ ממדי dNTP | 2.5 ממ ' | 2 מיקרומטר | דנטור, הרפתקאות | 94 ° c | שלושים בנות | 35 | |
| פריימר FBC8-F | 10 μM | 1 ליטר | ריפוי | 55 ° c | 1.5 דקות | ||
| פריימר FBC8-R | 10 μM | 1 ליטר | סיומת | 72 ° c | 2 דקות | ||
| 250 יו-אן-איי פלאז | 10 יו | 0.125 מיקרומטר | הארכה סופית | 72 ° c | 5 דקות | 1 | |
| H2O | 17.4 מיקרומטר | 4 מעלות צלזיוס | החזיק | ||||
. שולחן 1 תנאי תגובה ורכיבה על אופניים לשילובים שונים בשימוש בפרוטוקול זה.
| Primers | רצף | פניה |
| SRc-F | 5 '-CTCGCTTTCCTTGTTCTTGC -3 ' | 18 |
| SRc-R | 5 '-הגנה מפני 3 ' | 18 |
| Pru-C2 | 5 '-הגנה מפני 3 ' | 35 |
| Pru-C4R | 5 '-GGATGTGGTACGATTGAAGCG -3 ' | 35 |
| SHLM1-F | 5 '-מרכז התיירות-3 ' | 17 |
| SHLM2-R | 5 '-כמוסות שלוש ' | 17 |
| SHLM3-F | 5 '-מיכל העצמאות 3 ' | 17 |
| SHLM4-R | 5 '-כדור שלישי-3 ' | 17 |
| AprFBC8-F | 5 '-כמוסות מצרכות-3 ' | 26 |
| AprFBC8-R | 5 '-הגנה מפני 3 ' | 26 |
. שולחן 2 השימוש התחל בפרוטוקול זה, רצף והתייחסות לאפיון S-גנוטיפ בarmeniaca.
| זנים | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | אס-גנוטיפ | קבוצת אי-תאימות (I. G) |
| וונדר קוט8 | 420, 420 | 749, 1386 | S6s9 | שמיני | |||
| מג ' יק Cot8 | 334, 420 | 749 | S2s9 | XX | |||
| גולדסטרייק8 | 334, 420 | 749 | S2s9 | - | |||
| T06917 | 334, 408 | 650 | S1s2 | אני | |||
| T12017 | 334, 408 | 650 | S1s2 | - | |||
| C-6 | 334, 408 | 413 | S2s7 | רביעי | |||
| קופר קוט8 | 274, 408 | 650 | S1s3 | XVIII | |||
| אפריל המלכה | 358, 358 | 500 | Scsc | - | |||
| ברז8 | 334, 358 | 500 | S2sc | - | |||
| האביב סומק8 | 274, 358 | 150 | S3s8 | XXI |
. שולחן 3 S-גנוהקלדה של זנים משמש עם חמישה זוגות פריימר המשמשים בפרוטוקול זה והקצאת קבוצת אי-סבילות. שרשרת פולימראז שונה מגדלים את המוצרים של S-alleles מוגבר באמצעות SRc-(f/r), PruC2/PRUC4R, SHLM1/SHLM2, SHLM3/SHLM4, ו-AprFBC8-(f/r) התחל מוצגים בטבלה.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
באופן מסורתי, זנים מסחריים משמשים באירופה היו בעלי תאימות עצמית36. למרות זאת, השימוש בזנים של צפון אמריקה בלתי-מתאימים לעצמי כהורים בתוכניות רבייה בעשורים האחרונים הביא לשחרורו של מספר גדל והולך של זנים חדשים שאינם תואמים לעצמי עם דרישות האבקה מוכרות7,8,37. לפיכך, הקביעה של יחסי התאימות העצמית והבין (in) בזנים משמש חשובה יותר ויותר. זה מודגשת באותם אזורים שבהם החורף מצמרר הוא הפחתת, מאז השנה הגבוהה וריאציות השנה בזמן של פריחה מונעים את צירוף מקרים פריחה של זנים ו pollenizers שלהם במקרים רבים, במיוחד בזנים עם דרישות מצמררת גבוהה38. המתודולוגיה המתוארת במסמך זה, שילוב האבקה, מיקרוסקופ וניתוח גנטי שימושי מאוד כדי לקבוע את התאימות העצמית (בתוך) של כל זנים ולהקים את זנים המאביקים הפוטנציאליים שלה.
ניתן לקבוע את דרישות ההאבקה באמצעות ניסויי שליטה בשטח בתנאי המטע11,39. עם זאת, התערוכה לגורמים חיצוניים כולל תנאים לוואי מטאורולוגיים יכול לגרום לכשל האבקה10, אשר עשוי לגרום לאבחנות שגויות של אי-סבילות עצמית. המתודולוגיה המתוארת בזאת מאפשרת להעריך את התאימות העצמית (in) באופן מדויק יותר על ידי מיקרוסקופיה של פרחים מואבקי ידיים בתנאים שבשליטת מעבדה, הימנעות מהשפעה סביבתית. יתר על כן, גישה זו מאפשרת לנתח מספר גבוה יותר של זנים בשנה, שכן רק מספר קטן של פרחים נדרש במקום כמה עצים למבוגרים עבור כל זנים הנדרשים ניסויים בשטח40.
ניתן ליצור קשרי אי-תאימות המשלבים יד-מאביקים ומיקרוסקופ14. עם זאת, המאביקים ניתן לבצע רק תקופה קצרה במהלך עונת הפריחה באביב, ועצים למבוגרים ליד המעבדה נחוצים, מאז תוחלת החיים של הפרחים הנאספים הוא קצר מאוד. לפיכך, מספר היחסים אי-תאימות שניתן לנתח על ידי המאביקים יד מבוקרת בכל עונה הוא נמוך מאוד. אפיון הגנים המקודדים על ידי ה- S-לוקוס אפשרה פיתוח של שיטות ה-PCR ל-s-allele מבוסס18,41. גישה זו מאיצה את ההזדהות S-allele מאז הוא אינו דורש פרחים, ואת הניסויים ניתן לבצע עם כל הרקמה וגטטיבי42. זה מרחיב את התקופה שבה חומר הצמח, בדרך כלל עלים צעירים, ניתן לאסוף43. יתר על כן, העלים יכולים להיות מוקפאים או קפואים, כך הניתוח יכול להיעשות בכל עת של השנה, בניגוד המאביקים כי ניתן לעשות רק על פרחים טריים במהלך העונה פורח44. יתרון נוסף הוא כי העלים ניתן לאסוף מעצים צעירים גם לפני הכניסה גיל הפריחה, הקלה על אוסף של דגימות והשגת מוקדם של תוצאות45.
הניתוח הגנטי מאפשר הבחנה טובה יותר של אלס התאמה עצמית מאז הוא מספק תוצאות מדויקות של גודל מוגבר בגדלים21,46. עד היום, 33 S-alleles זוהו בשמש12,18,20,21,22,23,24, אשר אפשרה להקים 36 קבוצות אי תאימות מבוסס על S-גנוטיפ8,9,17,25,26,27. מצד שני, חיסרון של מתודולוגיה זו היא כי אללים שונים באותו גודל או מוטציות ניתן לזהות בטעות כמו אללים אותו. לכן, sc ו -s8 אללים זהים לרצף rnase אבל החדרת 358 bp נמצא בגנים sfb של Sc19. כמו כן, אזור אינטרון הראשון של האללים s1 ו -S7 זהים והם זהים באמצעות התחל SRc-F/SRc-R. בנוסף, מספר הומולוגיה, כגון s6 ו -s528 או s20 ו -s55, ו -s7, s13 (EF062341) ו -s4617, נמצאו מכיוון שחלק מהאללים הללו היתה משובשת חלקית או על ידי כשלים במהלך PCR הגברה, וכתוצאה מכך, עבודה נוספת נחוצה כדי להבדיל
ניתוח PCR ו -S-rnase רצף מספיקים להקמת קשרי אי-תאימות באמצעות זיהוי של s-alleles והקצאת זנים בקבוצה המקבילה שלהם אי תאימות8,17,26,27. עם זאת, למתודולוגיה זו יש הגבלה של מניעת קביעת התאימות העצמית (בתוך) של זנים משמשים מסוימים. תאימות עצמית (SC) היה קשור ל- S-alleles מינים אחרים 47, כמו שקדים (sf)48,49 או דובדבן מתוק (s4')50,51. עם זאת, משמש, sc אלל, אשר נקשר sc21, יכול להיות מזוהה בטעות כמו S8, אלל לא תואם עצמית19,22, ומוטציות אפשריות לא מקושרים לוקוס S , כמו M-לוקוס12,52, הענקת הוראות זוהו. לאחרונה, ה- M-לוקוס כבר הוקלד באמצעות סמני ssr12. לכן, הזיהוי הגנטי של SC עבור גנוטיפים משמש צריך מחקר נוסף, כדי למנוע טעויות בשל גורמים שאינם מקושרים ל -S לוקוס, בעבודה זו האפיון של עצמית (in) תאימות נקבע גם על ידי פנוטיפים התנהגות של צינורות אבקה דרך פיסטיל של פרחים עצמית מואבקים.
המתודולוגיה המתוארת בזאת משלבת את קביעת התאימות העצמית (in) באמצעות המאביקים בתנאי מעבדה עם התבוננות לאחר ההתנהגות של צינורות האבקה בפיסטיל של המאביקים העצמית הנשלטים תחת מיקרוסקופ הקרינה הפלואורסצנטית וזיהוי ה-S-גנוטיפ בניתוח PCR מאפשר לבסס את דרישות ההאבקה של זנים משמש זה מספק מידע חשוב עבור המגדלים והמגדלים, שכן הוא מאפשר לבסס את היחסים אי התאמה בין זנים כדי לבחור המאביקים מתאים בעיצוב של מטעים חדשים, כמו גם לבחור ההורים המתאימים לעצב צלבים חדשים בתוכניות הרבייה משמש.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
. למחברים אין מה לגלות
Acknowledgments
מחקר זה מומן על ידי Ministerio de Ciencia, האוניברסיטה האירופית לפיתוח האזור האירופי, האיחוד האירופי (AGL2016-77267-R, ו AGL2015-74071-ג'ין); המכון הלאומי לשימור (RFP2015-00015-00, RTA2017-00003-00); הקרן האירופית למדעי החברה, האיחוד האירופי (גריפו באסדדו A12_17R), Biodiversidad, וAgroseguro דרום אמריקה
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}