

Summary
PCR 분석에 의한 S-유전자형의 식별과 형광 현미경 검사법에 의한 자기(in)호환성을 결합한살구(Prunus armeniaca L.) 품종의 수분 요건을 확립하는 방법론을 제시한다.
Abstract
Rosaceae의 자기 비호환성은 주로 다골 궤적 S에 의해 제어되는 게임토피틱 자기 비호환성 시스템(GSI)에 의해 결정됩니다. 살구에서, 새로운 품종의 중요한 수의 방출이 알려지지 않은 수분 요구 사항을 가진 품종의 증가를 초래했기 때문에 자기 및 인터 -(in) 호환성 관계의 결정은 점점 더 중요해지고 있습니다. 여기서는 PCR 분석에 의한 S-유전자형식별과 손-오염 및 현미경 검사법에 의한 자기(in)호환성의 측정을 결합한 방법론을 설명한다. S- 자기(in)호환성 측정을 위해 각 품종의 풍선 스테이지에서의 꽃은 현장에서 수집되고, 실험실에서 수분, 고정 및 형광 현미경 검사하에 꽃가루 튜브 거동을 관찰하기 위해 애니라인 블루로 염색하였다. 품종 간의 비호환성 관계의 설립을 위해, 각 품종에서 DNA는 젊은 잎에서 추출되었고 S-alleles는 PCR에 의해 확인되었다. S 이 접근 법은 비호환성 그룹을 수립하고 품종 사이의 비호환성 관계를 해명 할 수 있으며, 이는 새로운 과수원 설계에 적합한 수분제를 선택하고 번식 프로그램에서 적절한 부모를 선택하는 귀중한 정보를 제공합니다.
Introduction
자기 비호환성은 식물을 꽃피는 전략으로 자가 수분을 방지하고 횡단 을 촉진하는1을촉진합니다. Rosaceae에서, 이 기계장치는 주로 다발성 궤적 S2에의해 제어되는 게임토피틱 자기 비호환성 시스템 (GSI)에 의해 결정됩니다. 스타일에서, RNase 유전자는 S-s타이라 결정제, RNase3을인코딩하고, S-꽃가루결정제를 결정하는 F 박스 단백질은 SFB 유전자4에의해 명문화된다. 자가비호환성 상호작용은 배란5,,6의수정을 방지하는 스타일을 따라 꽃가루 관 성장의 억제를 통해 일어난다.
살구에서, 품종 갱신은 지난 2 년 동안 전 세계적으로 일어났다7,,8. 이러한 새로운 품종의 중요한 수의 도입, 다른 공공 및 민간 사육 프로그램에서, 알 수없는 수분 요구 사항8살구 품종의 증가를 초래했다.
살구의 수분 요구 사항을 결정하는 데 다른 방법론이 사용되었습니다. 현장에서, 자체(in)호환성은 케이지 나무 또는 에뮬레이트된 꽃에서 제어된 오염에 의해 확립되고, 이후에 과일 세트9,,10,,11,12의백분율을기록할 수 있다. 또한, 대조된 오염은 꽃의 반 생체 배양에 의해 실험실에서 수행되었으며 형광 현미경검사법 8,13,,,14, 15,,16,,17하에서꽃가루 튜브 거동을 분석한다.15, 최근에는 PCR 분석 및 시퀀싱과 같은 분자 기법은 RNase 및 SFB 유전자18,,19의연구에 기초하여 비호환성 관계의 특성화를 허용하고 있다. 살구에서, 33 S-alleles(S1 ~ S20, S22 ~ S30,S52, S53,Sv, S S Sx)자가 호환성(SSc)12,18,,20,,,21,21,,22,,23,,24. 지금까지, 26 비호환성 그룹은 S-유전자형8,9,,17,25,,,,26,,27에따라 이 종에서 찌르고 있다. 동일한 S-대립구를 S가진 품종은 상호 호환되지 않는 반면, 적어도 하나의 다른 S-대립구를 가진 품종및 결과적으로, 다른 호환되지 않는 그룹에 할당, 상호 호환됩니다. S
살구 품종의 수분 요구 사항을 정의하기 위해, 우리는 살구 품종에서 PCR 분석에 의한 S-유전자형의식별과 형광 현미경 검사법에 의한 자기(in) 호환성의 측정을 결합하는 방법론을 설명합니다. 이 방법을 사용하면 비호환성 그룹을 설정하고 품종 간의 비호환성 관계를 설명할 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 자체(in) 호환성 결정
- 필드에 있는 꽃을 샘플링합니다. 원치 않는 이전 수분을 피하기 위해, 살구28에대한 BBCH 규모에 스테이지 58에 해당하는 풍선 단계(그림 1A)에서꽃을 수집 할 필요가있다.
-
실험실에서 자체 및 교차 pollinations
- 풍선 단계에서 꽃의 anthers를 제거하고 실험실 온도에서 건조 종이 조각에 배치합니다.
- 24시간 후, 미세 메쉬(0.26 mm)를 사용하여 꽃가루 곡물을 체질(도1B).
- 각 자가 수분 및 교차 수분에 대해 동일한 풍선 단계에서 30개의 꽃그룹을 방출하고 실험실 온도에서 꽃집 폼에 피스틸을 배치합니다(도1C).
- 손으로 는 방출 후 같은 품종 24 h의 꽃가루로 페인트 브러시의 도움으로 피슬을 수분합니다. 또한, 대조군으로 호환되는 꽃가루에서 꽃가루로 각 품종의 또 다른 피스틸 세트를 수분(도1D).
- 72h 후, 4°C29에서적어도 24시간 동안 에탄올/아세트산(3:1)의 고정용액으로 미스틸을 수정한다. 그런 다음 고정을 폐기하고 75% 에탄올을 추가하여 샘플이 용액에 완전히 침수되도록 합니다. ,샘플은8,17,,30,31, 32를사용할 때까지 4 °C에서이 용액에서 보존 될 수있습니다.,,
-
체외 꽃가루 발아를 통해 꽃가루 생존 가능성 평가
- 발아 배지를 준비하려면 자당 25g, 붕산 0.075 g(H3BO3)및 질산 칼슘 0.075 g(Ca(NO3)2)33을준비한다.
- 250mL의 증류수에 매체의 성분을 넣고 완전히 녹입니다.
- 아가로즈 2 g를 첨가하여 배지를 고형화하고 소용돌이에 섞습니다.
- pH 미터를 사용하여 매체의 pH를 확인하고 NaOH 또는 HCl 용액을 사용하여 값을 7.0으로 조정합니다.
- 배지를 살균하기 위해 혼합물을 자동 클로 처리합니다.
- 오토클레이브 후, 매체를 식히고 멸균 라미나르 플로우 후드에 페트리 접시에 분배합니다.
- 고화화 꽃가루 발아 배지에서 제어 된 꽃가루에 사용되는 동일한 품종의 꽃가루 곡물을 분산하고 24 h6후 현미경으로 관찰한다.
참고: 라미나르 플로우 후드를 살균하려면 표면에 70% 에탄올로 청소하고 10분 동안 UV 램프를 켭분으로 전환합니다. - 페트리 요리를 4°C의 냉장고에 보관하십시오.
-
현미경 관찰
- 증류수로 1시간 동안 3회 소염제를 세척하고 4°C에서 5% 설파이트 나트륨에 넣습니다. 24시간 후, 설파이트 나트륨에서 10분 동안 1kg/cm2로 오토클레이브하여 조직을 부드럽게한다(34).
- 유리 슬라이드 위에 오토클레이브 피슬을 놓고 메스의 도움으로 난소 주변의 삼각집을 제거하여 꽃가루 튜브의 더 나은 시각화를 얻습니다. 그런 다음 피슬을 커버 유리로 분쇄합니다.
- 준비 0.1% (v/v) 애니라인 블루 스테인: 0.1 mL의 애니라인 블루를 0.1mL의 100mL에서 0.1N 칼륨 인산염 삼각염(K3PO4)으로혼합한다. 꽃가루 튜브 성장 중 칼로스 증세를 얼룩지게하기 위해 준비 에일린 블루 한 방울을 적용하십시오.
- 340-380 밴드패스와 425개의 롱패스 필터를 사용하여 UV 에피플루오렌스를 사용하여 현미경으로 스타일을 따라 꽃가루 튜브를 관찰하십시오.
2. DNA 추출
- 봄에 필드에 2-3 잎을 샘플링합니다. DNA가 얻은 것은 오래된 잎에 비해 페놀 화합물의 높은 품질과 낮은 수준이기 때문에 젊은 단계에서 잎을 샘플링하는 것이 좋습니다.
- 시판되는 키트에 기재된 단계를 따라 유전체 DNA를 추출합니다(재료표참조).
- UV-비스 분광광미터(260nm)를 사용하여 DNA 농도의 양과 품질을 분석합니다.
3. S-알레알렐 식별
-
PCR 반응 설정
- 각 DNA 추출 샘플의 증류수에 50 ng/μL 희석을 준비합니다.
- PCR 시약을 천천히 해동하고 얼음 위에 보관하십시오. 필요할 때까지 DNA 폴리머라아제를 냉동실에 둡니다.
- 프라이머의 다른 조합을 사용하여 증폭 반응을 준비합니다. 표 1의구성 요소를 결합하여 PCR 반응 믹스를 만듭니다. PCR 반응이 잘 섞여 서 프라이머의 상이한 조합에 대해 표시된 부피를 PCR 플레이트의 각 웰에 분배한다. 그런 다음 각 우물에 DNA 희석의 1 μL을 추가합니다.
- PCR 플레이트를 열순환기에 놓고 표 1에표시된 해당 PCR 프로그램을 실행합니다.
- 증폭된 조각을 분석합니다. PCR 증폭 된 단편을 분석하는 두 가지 방법이 주로 있습니다 : 형광 라벨 프라이머를 가진 모세관 전기 포근 (CE) 또는 표기되지 않은 프라이머와 아가로즈 젤 전기 포의 앰프슨을 시각화.
-
모세관 전기 포레시스
- 로딩 버퍼를 준비하려면 35 μL의 탈이온화된 포름아미드를 라벨이 붙은 크기 조정 표준의 0.45 μL을 혼합합니다. 시약이 잘 섞인 다음 35.5 μL을 판독판의 우물에 분배합니다.
- PCR 제품의 1 μL을 우물에 넣습니다. 또한, 물 증발을 방지하기 위해 미네랄 오일 한 방울을 추가합니다.
- 분리 버퍼를 추가하는 분리 플레이트를 준비합니다.
- 유전자 분석기와 함께 포함된 상용 소프트웨어를 사용합니다(재료표참조). 새 샘플 플레이트를 만들고 플레이트의 모든 우물에 대한 샘플 이름을 저장합니다.
- 분석 방법을 선택합니다. 이 경우, 120s에 대해 90°C에서 샘플을 분해하고, 30초 동안 2.0kV에서 주입하고, 35분 동안 6.0kV로 분리한다.
- 유전자 분석기에 두 개의 플레이트를 삽입합니다. 모세관 배열을 증류수로 채웁니다.
- 특허받은 선형 폴리아크라이아미드(LPA) 젤을 적재한다. 마지막으로 실행을 클릭합니다.
-
젤 전기전증
- 1.5g의 분자생물학급 아가로즈를 1xTAE(Tris-아세테이트-EDTA) 전기포고리(40m Tris, 20mM 아세트산, 1mM EDTA pH 8.0)로 첨가하여 1%의 아가로즈 젤을 준비한다. 2-3 분 동안 전자 레인지 가열에 의해 아가로즈를 녹입니다.
- DNA를 시각화하려면 핵산 얼룩의 4 μL을 추가하고(재료 표 참조) 부드럽게 섞습니다.
- 사다리, 컨트롤 및 샘플을 위한 충분한 우물이 있는 젤 빗을 젤 트레이에 넣습니다. 그런 다음 젤 트레이 의 중간에 천천히 믹스를 붓고 거품을 피하십시오.
- 젤이 완전히 고화될 때까지 실온에서 30-45분 동안 젤을 식힙니다. 전기 전광 챔버에 젤을 소개하고, 젤 빗을 제거하고 젤을 덮을 충분한 1x TAE 버퍼로 챔버를 채웁니다.
참고: 젤의 배치를 확인합니다. 우물은 음극쪽으로 부정적인 충전 된 DNA가 이동하기 때문에 부정적인 극에 가깝게 배치해야합니다. - PCR 제품에 로딩 버퍼 5μL(0.1% (v/v) 브로모페놀 블루를 추가하고 잘 섞는다.
- 대역의 크기를 추정하려면 DNA 분자량 사다리의 5 μL을 적재합니다(재료 표참조).
- 샘플을 젤의 추가 우물에 적재합니다.
- 모든 샘플과 DNA 분자량 사다리가 로드되면, 블루 염료 라인이 젤의 길이약 75%가 될 때까지 90 V에서 1-1.5h로 젤을 실행합니다.
- 핵산에 대한 일루미에이터에서 밴드를 시각화합니다.
-
모세관 전기 포레시스
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
살구의 수분 연구는마취(그림 1A)가발생하기 하루 전에 늦은 풍선 단계에서 꽃을 사용해야 합니다. 이 단계는 꽃가루와 피스티트 컬렉션 모두에 가장 유리한 것으로 간주됩니다, 꽃 무늬 구조는 거의 성숙하기 때문에, 하지만 anther dehiscence는 아직 발생하지 않았습니다. 이는 폐쇄된 꽃잎이 외부 꽃가루를 운반하는 곤충의 도착을 방해하기 때문에 같은 꽃가루뿐만 아니라 다른 꽃가루의 간섭을 방지합니다. 꽃가루 곡물은 이전에 실온에서 24 시간 동안 종이에 놓인 데시드 마취제에서 미세 메쉬(그림 1B)를통해 쉽게 체질되거나 약간의 여분의 열을 가합니다. 마찬가지로, 피스틸은 핀셋이나 손톱의 도움으로 꽃잎, 세팔 및 stamens를 제거 한 후 풍선 단계에서 꽃에서 얻을 수 있습니다(그림 1C). 피스틸은 미세 한 브러시(도 1D)로자체 및 교차 수분 될 수 있습니다.
살구의 헤르마포디틱 꽃은 5개의 어두운 붉은 세팔, 5개의 흰 꽃잎(그림1A),단일 피스틸(그림2A)및 25-30개의 스타멘을 가지고 있습니다.Figure 2 오명, 스타일 및 난소의 세 가지 주요 구조가 있습니다. 난소에는 두 개의 배란이 있으며 과일 설정에는 적어도 하나의 변속이 필요합니다. 수분 중, 곤충, 주로 꿀벌, 꽃가루 곡물을 오명으로 옮기는 동안(도 1A),그들은 발아(그림 2B)24 시간 수분 다음. 꽃가루 튜브는 3-4 일 후에 난소에 도달하고 약 7 일 후에 2개의 난소 중 하나를 기름지게 하는 피질 구조물을 통해 성장하는 각 발아 꽃가루 곡물에서 생성됩니다. 꽃가루 곡물의 S 골자가 두 가지 양슬 중 하나와 동일한 자체 호환 품종에서 꽃가루 튜브는 상부 스타일로 성장하여 수정을 방지합니다(도 2C). 그러나, 호환되는 품종으로부터의 꽃가루 튜브는 다른 S 알레를 가진, 난소(도 2E)에도달하고 두 배란 중 하나를 비옥하게 하는 스타일(도Figure 22D)을통해 성장할 수 있다.Figure 2
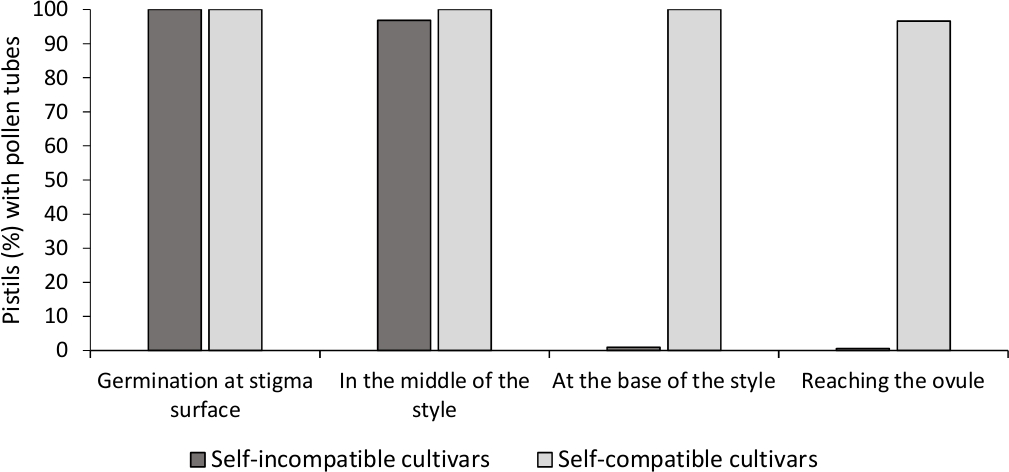
체외 꽃가루 발아의 분석은 대부분의 꽃가루 튜브가 배양 배지에서 24 시간 후 꽃가루 곡물의 길이보다 길기 때문에 여기에서 분석 된 모든 품종에서 좋은 꽃가루 생존력을 보였다. 발아 꽃가루 곡물은 모든 꽃가루로부터 의약표면(도 2B)에서관찰되었으며, 이는 적절한 수분(도3)을나타낸다.
각 품종에 대한 자체(in) 호환성을 결정하기 위해, 실험실 대조 조건에서 수행된 자체 및 교차 오염물질에서 꽃가루 튜브 동작이 형광 현미경 검사법에 따라 관찰되었다. 꽃가루 튜브 성장은 검사 된 모든 피스틸의 스타일을 따라 기록되었습니다. 품종은 꽃가루 튜브 성장이 대부분의 자가 수분 된 피스티틸(도 2C, 도 3)에서스타일에 따라 체포되었을 때 자체 호환되지 않는 것으로 간주되었으며, 적어도 하나의 꽃가루 튜브가 대부분의 피스티틸에서 스타일의 베이스에 도달했을 때(그림 2E, 도 3).
PCR 분석에 의한 S-궤적의연구는 각 품종의 S-유전자형을특성화할 수 있었습니다. 먼저, S-알레일은 프라이머 SRc-F/SRc-R(표2)을이용하여 제1 S S-RNase 인트론의 증폭에 의해 확인되었다. 증폭된 단편의 크기는 모세관 전기포고증(도Figure 44A)에의해 분석되었으며 해당 비호환성 군(I.G.)에서 분석된 유전자형을 분류하는 데 사용되었습니다. (표3).
S1 및 S7 또는 S6 및 S9와같은 일부 항자 쌍은 첫 번째 인트론에 대해 유사한 단편 크기를 보였다. 따라서, 이들 알레일의 분화는 프라이머 프루-C2/PruC4R, SHLM1/SHLM2 및 SHLM3/SHLM4(표2)와함께 RNase의 제2 인트론영역을 증폭시킴으로써 이루어졌다. PruC2/PruC4R 프라이머 조합은 S6과 S9를구별하는 데 사용되었습니다. S6의6경우, 1300 bp의 단편이 증폭된 반면 약 700bp의 단편은 S9 알렐(도4B, 표 3)에대해 관찰되었다. 특정 프라이머 SHLM1/SHLM2 및 SHLM3/SHLM4는S7 알레일에서 약650bp, S77 알렐레에서 413bp의 단편을 증폭시켰다(도4C, 표 3).Figure 4
SFB 유전자의 V2 및 HVb 가변 영역을 증폭시키는 프라이머 AprFBC8-(F/R)는 두 유전자가 동일한 RNase 서열을 나타내기 때문에 Sc 및 S8 알렐을 구별하는 데 사용되었습니다. S88 알렐은 약 150bp의 PCR-단편을 보였고, 500bp 단편은 Sc 알레(도4D, 표 3)에대응하였다.Figure 4 S-유전자형이 모든 품종에 대해 결정되면, 자체 호환되지 않는 품종은 S-alleles(표3)에기초하여 해당 비호환성 그룹에 할당되었다. S
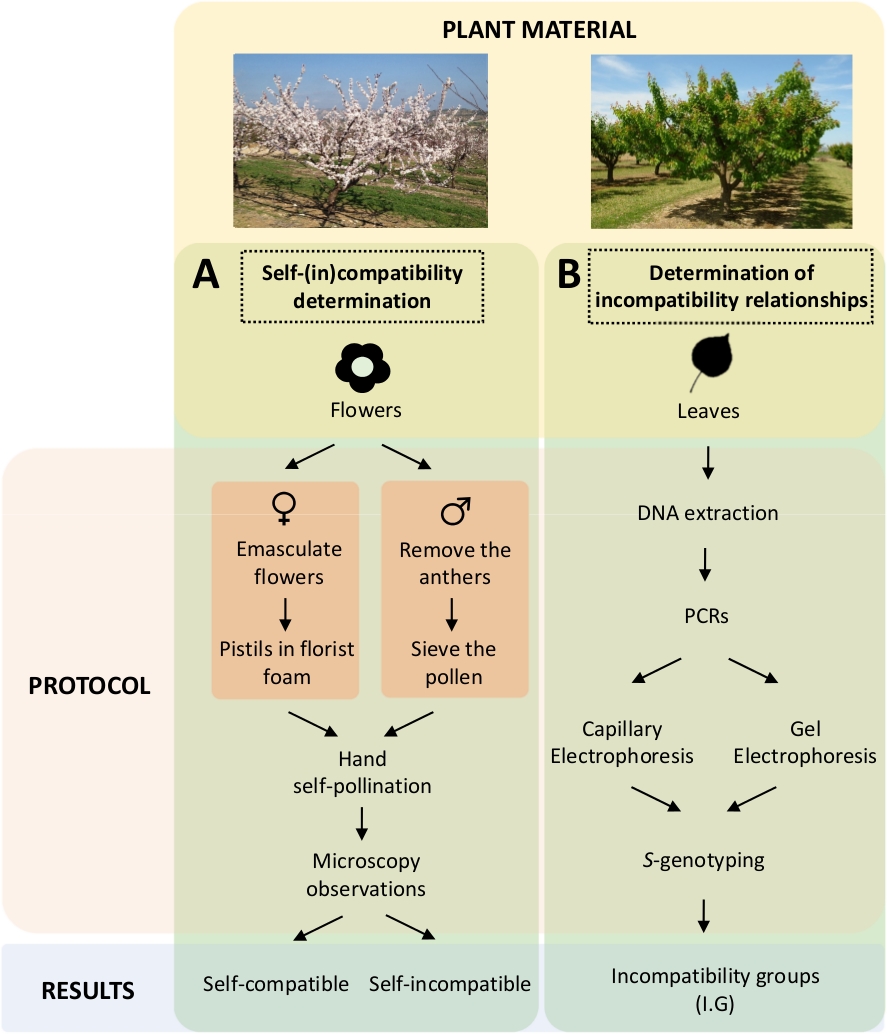
이러한 접근법은 실험실에서 제어된 자기 및 교차 오염에 의한 각 품종의 자체(in) 호환성을 유전자 분석에 의한Figure 5AS-유전자형의 특성화와 수반되는 것으로 판단해야 한다(도5B). S 그 결과, 각 품종의 수분 요구 사항과 살구 품종 간의 비호환성 관계를 결정할 수 있다.

그림 1. 살구에서 자체(in) 호환성의 결정을 위해 실험 설정.
(A)필드의 풍선 스테이지(검은 화살)의 꽃. (B)미세 메쉬를 사용하여 꽃가루 곡물의 체. (C)물에 꽃집 거품에 배치 피스틸. (D)페인트 브러시의 도움으로 피스틸의 손 수분. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 2. 살구 꽃에서 게임 토피성 비호환성 관계의 다이어그램 표현.
(A)게임토피틱 자가비호환성(GSI)에서는 호환되지 않는 꽃가루 곡물이 오명을 발아합니다. 꽃가루 곡물은 원래 유전자형의 두 개의 S-alleles 중 하나를 운반하며,이 경우 S1 또는 S2. S 꽃가루 곡물의 S-allele이 피슬틸의 두 가지 S-alleles중 하나와 일치하는 경우,이 경우 S1S3,꽃가루 튜브 의 성장이 스타일의 상위 1/3에서 억제된다. S (B)낙인 표면에 꽃가루 곡물의 발아. (C)꽃가루 튜브는 호환되지 않는 행동을 나타내는 스타일로 체포. (D)꽃가루 튜브는 스타일을 따라 성장. (E)호환 동작을 나타내는 스타일의 기저에 꽃가루 튜브. 스케일 바, 100 μm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 3. 자체 오염 후 자체 호환 및 자체 호환 되지 않는 품종에 대 한 스타일을 통해 꽃가루 발 아 게 와 꽃가루 튜브 성장의 대표적인 결과.
꽃가루 곡물이 오명 표면에서 발아하고 꽃가루 튜브가 스타일 의 중간, 스타일 의 기지에서, 그리고 난란에 도달하는 피스티의 백분율. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 4. S-알레일의 식별을 위해 5개의 프라이머 S쌍 조합을 사용하여 PCR 단편 증폭.
(A)SRc-(F/R) 프라이머에 대한 유전자 분석기 출력은 S-알레에대응하는 RNase 제1 인트론 영역의 2개의 증폭 된 단편의 크기를 나타내는 프라이머. (B) S6 및 S9 알레일의 식별을 위해 프라이머 PruC2/PruC4R을 사용하여 PCR 증폭. (C) SS1 알렐및 SHLM3 및 SHLM4의 분화를 위해 특정 프라이머 SHLM1 및 SHLM2를 사용하여 얻은 PCR 제품은 S7 골자를 구별한다. (D) Sc 및 S8 알렐을 식별하기 위한 AprFBC8-(F/R) 프라이머를 가진 PCR 증폭. MI: 1 kb DNA 사다리. MII: 100 bp DNA 사다리. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 5. 살구 품종에서 자체 및 인터-(in) 호환성 관계를 해명하는 실험 설계의 계획.
(A)실험실에서 제어된 오염에 의한 자체(in) 호환성 결정의 워크플로우. (B)분자 접근법에 의한 S-알레식별의 워크플로우. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
| PCR 마스터 믹스 | 열순환기 조건 | ||||||
| 구성 요소 | 최종 농도 | 15 μL 반응 | 사이클 단계 | 온도 | 시간 | 사이클 | |
| 10x NH4 반응 버퍼 | 10x | 1.5 μL | 초기 변성 | 94°C | 3분 | 1 | |
| 50 mM MgCl2 솔루션 | 25 mM | 1.2 μL | 부인 | 94°C | 1분 | 35 | |
| 100mM dNTP | 2.5 mM | 0.6 μL | 어 닐 링 | 55 °C | 1분 | ||
| 프라이머 SRc-F | 10 μM | 0.6 μL | 확장 | 72 °C | 3분 | ||
| 프라이머 SRc-R | 10 μM | 0.6 μL | 최종 확장 | 72 °C | 5분 | 1 | |
| 500 U 타크 DNA 폴리머라제 | 0.5 U | 0.2 μL | 4°C | 개최 | |||
| H2O | 8.3 μL | ||||||
| 구성 요소 | 최종 농도 | 25 μL 반응 | 사이클 단계 | 온도 | 시간 | 사이클 | |
| 10x PCR 버퍼 | 10x | 2.5 μL | 초기 변성 | 94°C | 2분 | 1 | |
| 5x Q 솔루션 | 5배 | 5 μL | 부인 | 94°C | 10 s | 10 | |
| 100mM dNTP | 2.5 mM | 0.5 μL | 어 닐 링 | 55 °C | 2분 | ||
| 프라이머 프루C2 | 10 μM | 0.2 μL | 확장 | 68 °C | 2분 | ||
| 프라이머 C4R | 10 μM | 0.2 μL | 부인 | 94°C | 10 s | 25 | |
| 250 U 타크 DNA 폴리머라제 | 10 U | 0.13 μL | 어 닐 링 | 58 °C | 2분 | ||
| H2O | 15.5 μL | 확장* | 68 °C | 2분 | |||
| 최종 확장 | 72 °C | 5분 | 1 | ||||
| 4°C | 개최 | ||||||
| * 68 %C 확장 단계에 각 주기를 10 s추가. | |||||||
| 구성 요소 | 최종 농도 | 25 μL 반응 | 사이클 단계 | 온도 | 시간 | 사이클 | |
| 10x PCR 버퍼 | 10x | 2.5 μL | 초기 변성 | 94°C | 2분 | 1 | |
| 5x Q 솔루션 | 5배 | 5 μL | 부인 | 94°C | 30 s | 35 | |
| 100mM dNTP | 2.5 mM | 0.5 μL | 어 닐 링 | 62 °C | 1.5분 | ||
| 프라이머 SHLM1 | 10 μM | 0.2 μL | 확장 | 72 °C | 2분 | ||
| 프라이머 SHLM2 | 10 μM | 0.2 μL | 최종 확장 | 72 °C | 5분 | 1 | |
| 250 U 타크 DNA 폴리머라제 | 10 U | 0.13 μL | 4°C | 개최 | |||
| H2O | 15.5 μL | ||||||
| 구성 요소 | 최종 농도 | 20 μL 반응 | 사이클 단계 | 온도 | 시간 | 사이클 | |
| 5x PCR 버퍼 | 5배 | 4 μL | 초기 변성 | 98 °C | 30 s | 1 | |
| dNTP | 2.5 mM | 1.6 μL | 부인 | 98 °C | 10 s | 35 | |
| 프라이머 SHLM3 | 10 μM | 1 μL | 어 닐 링 | 51 °C | 30 s | ||
| 프라이머 SHLM4 | 10 μM | 1 μL | 확장 | 72 °C | 1분 | ||
| 100 U DNA 폴리머라제 | 5 U | 0.2 μL | 최종 확장 | 72 °C | 5분 | 1 | |
| H2O | 12.4 μL | 4°C | 개최 | ||||
| 구성 요소 | 최종 농도 | 25 μL 반응 | 사이클 단계 | 온도 | 시간 | 사이클 | |
| 10x PCR 버퍼 | 10x | 2.5 μL | 초기 변성 | 94°C | 2분 | 1 | |
| 100mM dNTP | 2.5 mM | 2 μL | 부인 | 94°C | 30 s | 35 | |
| 프라이머 FBC8-F | 10 μM | 1 μL | 어 닐 링 | 55 °C | 1.5분 | ||
| 프라이머 FBC8-R | 10 μM | 1 μL | 확장 | 72 °C | 2분 | ||
| 250 U 타크 DNA 폴리머라제 | 10 U | 0.125 μL | 최종 확장 | 72 °C | 5분 | 1 | |
| H2O | 17.4 μL | 4°C | 개최 | ||||
표 1. 이 프로토콜에 사용되는 다양한 프라이머 조합에 대한 반응 및 사이클링 조건.
| 뇌관 | 시퀀스 | 참조 |
| SRc-F | 5'-CTCGCTTTCCTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| 프루 C2 | 5'-CTTTGGCCAAGTAATTCAAACC-3' | 35 |
| 프루-C4R | 5'-GGGTGGTAC가가아가그CG-3' | 35 |
| SHLM1-F | 5'-GGGAGGGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGAGAGAGAGGTAG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTCTCTCTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATG-3' | 17 |
| 4월BC8-F | 5'-CATGGAAAAGCTGACTTATGG-3' | 26 |
| 4월BC8-R | 5'-GCCTAATGTCATCTCTCTCTCTTAG-3' | 26 |
표 2. 이 프로토콜에 사용되는 프라이머, Prunus armeniaca에서 S-유전자형특성에 대한 서열 및 참조.
| 품종 | SRc-(F/R) (bp) | PruC2/PruC4R(bp) | SHLM1/SHLM2(bp) | SHLM3/SHLM4(bp) | 4월BC8-(F/R) (bp) | S-유전자형 | 비호환성 그룹(I.G) |
| 원더 코트8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| 매직 코트8 | 334, 420 | 749 | S2S9 | Xx | |||
| 골드스트라이크8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ⅰ | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| 쿠퍼 코트8 | 274, 408 | 650 | S1S3 | Xviii | |||
| 아프리퀸 | 358, 358 | 500 | ScSc | - | |||
| 베르그코트8 | 334, 358 | 500 | S2Sc | - | |||
| 봄 블러시8 | 274, 358 | 150 | S3S8 | Xxi |
표 3. 이프로토콜 및 비호환성 그룹 할당에 사용되는 5개의 프라이머 쌍을 가진 살구 품종의 S-genotyping. SRc-(F/R), PruC2/PruC4R, SHLM1/SHLM2, SHLM3/SHLM4 및 AprFBC8-(F/R) 프라이머를 사용하여 증폭된 S-Alleles의 상이한 폴리머라제 연쇄 반응 제품 크기가 표에 표시됩니다. S
Subscription Required. Please recommend JoVE to your librarian.
Discussion
전통적으로, 대부분의 상업 살구 유럽 품종은 자기 호환36이었다. 그럼에도 불구하고, 지난 수십 년 동안 번식 프로그램에서 부모로서 북미 자가 호환되지 않는 품종의 사용은 알 수없는 수분 요구 사항7,,8,,37새로운자기 호환 품종의 증가의 출시 결과. 따라서 살구 품종에서 의외의 호환성 관계의 결정은 점점 더 중요해지고 있다. 이것은 겨울 냉각이 감소하는 지역에서 강조된다, 꽃의 시간에 높은 년마다 변화는 많은 경우에 품종과 꽃가루의 꽃에 우연의 일치를 방지하기 때문에, 특히 높은 냉각 요구 사항과 품종38. 본 원에 기술된 방법론, 수분, 현미경 및 유전 분석을 결합하여 각 품종의 자체(in) 호환성을 결정하고 잠재적인 수분화 품종을 확립하는 데 매우 유용하였다.
수분 요구 사항은 과수원조건(11,,39)에서현장 제어 실험을 통해 결정될 수 있다. 그러나, 기상 불리한 조건을 포함하여 외부 요인에 대한 박람회는 수분 실패를 일으킬 수 있습니다(10)자기 비호환성의 잘못된 진단을 초래할 수 있습니다. 본 원에 설명된 방법론은 실험실 에서 수분이 있는 꽃의 현미경 관찰에 의해 자기(in) 호환성을 보다 정확하게 평가하여 환경적 영향을 피할 수 있게 한다. 더욱이, 이러한 접근법은 필드실험(40)에필요한 각 품종에 대해 여러 성인용 나무 대신 소수의 꽃만 필요하기 때문에 연간 더 많은 품종을 분석할 수 있다.
비호환성 관계는 손-오염및 현미경검사법(14)을결합하여 확립될 수 있다. 그러나, 꽃피는 봄에 꽃 시즌 동안 짧은 기간 동안만 수행 할 수 있으며, 수집 된 꽃의 수명이 매우 짧기 때문에 실험실 근처의 성인 나무가 필요합니다. 따라서, 각 계절에 통제된 손 질투량에 의해 분석될 수 있는 비호환성 관계의 수는 매우 낮습니다. S-메뚜기에 의해 인코딩된 유전자의 특성화는 S-allelegenotyping S18,,41에대한 PCR 기반 방법의 개발을 가능하게 했다. 이 접근법은 S꽃을 필요로하지 않기 때문에 S-allele 식별을 가속화하고, 실험은 식물조직(42)으로수행될 수 있다. 이는 식물 재료, 일반적으로 젊은 잎을 수집 할 수있는 기간을연장43. 더욱이, 잎은 라이오필화 또는 냉동될 수 있으므로,꽃시즌(44)에는신선한 꽃에서만 할 수 있는 질화와 는 달리, 일년 중 언제든지 분석을 수행할 수 있다. 추가 적인 이점은 잎이 꽃에 들어가기 전에 어린 나무에서 수집 할 수 있다는 것입니다, 샘플의 수집을 용이하게하고 결과의 조기 획득(45).
유전자 분석은 증폭된 단편 크기21,,46의정확한 결과를 제공하기 때문에 자기 비호환성 적 유전자의 더 나은 분화를 허용한다. 현재까지 33개의 S-alleles는살구,12,18,,20,,21,,22,,23,,24로,확인되었으며, S-유전자형8,9,9,17,25,,26, 26,,27에기초하여 36개의 비호환성 군을 설정할 수 있다. 한편, 이러한 방법론의 단점은 동일한 범위 크기 또는 돌연변이에 있는 상이한 색유전자가 동일한 유전자로 잘못 식별될 수 있다는 것이다. 따라서, Sc 및 S8 알레일은 RNase 서열에 대해 동일하지만 358-bp 삽입은 Sc19의SFB 유전자에서 발견된다. 마찬가지로, 올레 S1 및 S7의 첫 번째 인트론 영역은 동일하며 프라이머 SRc-F/SRc-R을 사용하여 구별할 수 없다. 또한,S6 및 S52 S8 또는 S20 및 S S55,S S7, S13(EF062341) 및 S4617과같은 여러 동종학은 이러한 알레일 중 일부가 PCR 증폭 중에 부분적으로 시퀀스되거나 실패에 의해 발견되었으며, 결과적으로 이를 올바르게 구별하기 위해 추가 작업이 필요합니다.13
PCR 분석,및 S-RNase 시퀀싱은 S-alleles의식별및 해당 비호환성 그룹8,17,26,,27의품종 할당을 통해 호환성 관계를 수립하는 데 적합합니다., 그러나, 이 방법론은 특정 살구 품종에 대한 자기(in) 호환성의 판정을 방지하는 한계를 가지고 있다. 자기 호환성(SC)은 다른 프루누스 종(47)에서특정 S-alleles에,아몬드(Sf)48, 49,49 또는 달콤한 S체리(S4)’,50,51로연관되어 있다. 그러나, 살구에서, SC 알레일(SC 21)은S8,자가 호환되지 않는 S알레일(19,22)으로 잘못 식별될 수 있으며, S-메뚜기(12, M12,52)및 SC를 수여하는 SC로서,22S-메뚜기와 연결되지 않은 돌연변이가 확인되었다. S 최근에는 SSR마커(12)를사용하여 M-메뚜기가 유전자형이 사용되었습니다. 따라서, 살구 유전형에 대한 SC의 유전자 식별은 추가 연구가 필요하며, S 궤적에 연결되지 않은 요인으로 인한 실수를 피하기 위해, 이 작품에서 자가(in)호환성의 특성화는 또한 자가 수분꽃의 피질을 통해 꽃가루 관의 동작을 표현함으로써 결정되었다.
본 명세서에 기재된 방법론은 실험실 조건에서 수로 오염물질에 의한 자체 측정을 결합하여 형광 현미경검사에 따른 조절된 자가 염분포의 피스트에서 꽃가루 관의 S거동을 관찰하고 PCR 분석에 의한 S-유전자형의 식별을 결합하여 살구 품종의 수분 요구 사항을 확립할 수 있게 한다. 이것은 재배자와 육종가를위한 귀중한 정보를 제공합니다, 그것은 새로운 과수원의 디자인에 적합한 수분화기를 선택하고 살구 사육 프로그램에서 새로운 십자가를 설계하는 적절한 부모를 선택하는 품종 사이의 비호환성 관계를 수립 할 수 있기 때문에.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자는 공개 할 것이 없습니다.
Acknowledgments
이 연구는 장관 드 시엔시아에 의해 투자되었다, 혁신시온 y Universidades-유럽 지역 개발 기금, 유럽 연합 (AGL2016-77267-R, AGL2015-74071-JIN); Instituto Nacional de Investigación y Tecnología Agraria y 알리멘타리아 (RFP2015-00015-00, RTA2017-00003-00); 고비에르노 데 아라곤-유럽 사회 기금, 유럽 연합 (그루포 콘솔리다 A12_17R), 펀다시온 바이오 다이버 시다드, 그리고 아로즈구로 S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}