

Summary
Vi presenterar en metod för att fastställa pollinering krav aprikos(Prunus armeniaca L.) sorter som kombinerar bestämning av self-(in)kompatibilitet genom fluorescensmikroskopi med identifiering av S-genotyp av PCR analys.
Abstract
Självinkompatibilitet i Rosaceae bestäms av ett gametophytic self-inkompatibilitetssystem (GSI) som huvudsakligen styrs av multiallelic locus S. I aprikos, beslutsamheten av själv- och inter-(in)förenlighet förhållandena är mer och mer viktig, sedan frigöraren av ett viktigt nummer av nya kultivars har resulterat i förhöjningen av sorterna med okända pollineringkrav. Här beskriver vi en metod som kombinerar bestämning av själv-(i)kompatibilitet genom handpollinationer och mikroskopi med identifiering av S-genotyp genom PCR-analys. För själv-(i)kompatibilitet bestämning, blommor på ballong skede från varje sort samlades i fältet, handpollinas i laboratoriet, fast, och färgas med aniline blå för observation av pollenröret beteende under fluorescensmikroskopi. För att upprätta oförenlighetsförhållanden mellan sorter extraherades DNA från varje sort från Sunga blad och S-alleler identifierades av PCR. Detta tillvägagångssätt gör det möjligt att upprätta inkompatibilitetsgrupper och belysa oförenlighetsrelationer mellan sorter, vilket ger en värdefull information för att välja lämpliga pollinatörer i utformningen av nya fruktträdgårdar och att välja lämpliga föräldrar i avelsprogram.
Introduction
Självinkompatibilitet är en strategi för blommande växter för att förhindra självpollinering och främja outcrossing1. I Rosaceae bestäms denna mekanism av ett gametophytic självinkompatibilitetssystem (GSI) som huvudsakligen styrs av multiallelic locus S2. I stil, RNase genen kodar S-stylar avgörande, en RNase3, medan en F-box protein, som bestämmer S-pollen beterminant, kodifieras av SFB genen 4. Den självinkompatibilitet interaktion sker genom hämning av pollen röret tillväxt längs stilen förhindra befruktning av ägglossningen5,6.
I aprikos, en sort förnyelse har ägt rum över hela världen under de senaste två decennierna7,8. Detta införande av ett stort antal nya sorter, från olika offentliga och privata avelsprogram, har resulterat i en ökning av aprikos sorter med okänd pollinering krav8.
Olika metoder har använts för att fastställa pollinering krav i aprikos. På fältet kan självkompatibilitet fastställas genom kontrollerade pollineringar i burträd eller i emasculated blommor och därefter registrera andelen frukt som9,,10,,11,12. Dessutom har kontrollerade pollineringar utförts i laboratoriet genom semi-in vivo kultur av blommor och analys av pollenröret beteende under fluorescensmikroskopi8,13,14,15,16,17. Nyligen har molekylära tekniker, såsom PCR-analys och sekvensering, gjort det möjligt att karakterisera inkompatibilitetsrelationer baserade på studien av RNase- och SFB-generna 18,19. I aprikos har trettiotre S-alleler rapporterats(S1 till S20, S22 till S30, S52, S53, Sv, Sx), inklusive en allel relaterad med självkompatibilitet(Sc)12,18,,20,,21,,22,23,24. Hittills har 26 inkompatibilitetsgrupper huggits i denna art enligt S-genotyp8,9,17,25,26,27. S Sorter med samma S-alleler är oförenliga mellan sorter, medan sorter med minst en annan S-alleloch följaktligen tilldelas i olika oförenliga grupper, är inte förenliga. S
För att definiera pollineringskraven för aprikossorter beskriver vi en metod som kombinerar bestämning av själv-(i)kompatibilitet genom fluorescensmikroskopi med identifiering av S-genotyp genom PCR-analys i aprikossorter. Detta tillvägagångssätt gör det möjligt att upprätta inkompatibilitetsgrupper och belysa inkompatibilitetsrelationer mellan sorter.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Fastställande av själv(i)förenlighet
- Prova blommorna på fältet. Det är nödvändigt att samla blommorna på ballongstadiet (Figur 1A), motsvarande steg 58 på BBCH-skalan för aprikos28, för att undvika oönskad tidigare pollinering.
-
Själv- och korspollinationer i laboratoriet
- Ta bort anthers av blommorna på ballong skede och placera dem på ett papper för att torka vid laboratorietemperatur.
- Efter 24 timmar, sikta pollenkornen med hjälp av ett finmaskigt nät (0,26 mm) (figur 1B).
- Emasculate en grupp på 30 blommor på samma ballong skede för varje självpollinering och korspollinering och placera pistiller på florist skum i vatten vid laboratorietemperatur (figur 1C).
- Hand pollinera pistillerna med hjälp av en pensel med pollen från blommor av samma sort 24 h efter emasculation. Dessutom pollinera en annan uppsättning pistiller av varje sort med pollen från blommor av en kompatibel pollinerare som kontroll (Figur 1D).
- Efter 72 timmar, fixera pistillerna i en fixativ lösning av etanol/ättiksyra (3:1) under minst 24 timmar vid 4 °C29. Kassera sedan fixativet och tillsätt 75 % etanol så att proverna är helt nedsänkta i lösningen. Proverna kan bevaras i denna lösning vid 4 °C tills den används8,,17,,30,,31,32.
-
Utvärdera pollen lönsamhet genom in vitro pollen grobarhet
- För att förbereda groningsmediet, vikt 25 g sackaros, 0,075 g borsyra (H3BO3)och 0,075 g kalciumnitrat (Ca(NO3)2)33.
- Tillsätt komponenterna i mediet i 250 ml destillerat vatten och lös upp helt.
- Stelna mediet lägga 2 g agaros och blanda genom virvlande.
- Kontrollera mediets pH-värde med hjälp av en pH-mätare och justera värdet till 7,0 med NaOH- eller HCl-lösning.
- Autoklav blandningen för att sterilisera mediet.
- Efter autoklavering, kyl ner mediet och fördela det i petriskålar i en steril laminär flödeshuva.
- Sprid pollenkorn av samma sorter som används för kontrollerade pollineringar i det stelnade pollengroningsmediet och observera dem under mikroskopet efter 24 h6.
OBS: För att sterilisera den laminära flödeshuven, rengör ytan med 70% etanol och slå på UV-lampan under 10 min. - Förvara petriskålarna i kylskåp vid 4 °C tills de används.
-
Observationer av mikroskopi
- Tvätta pistillerna tre gånger i 1 h med destillerat vatten och lämna dem i 5% natriumsulfit vid 4 °C. Efter 24 h, autoklav dem på 1 kg/cm2 under 10 min i natriumsulfit för att mjuka upp vävnaderna34.
- Placera de autoklavererade pistillerna över en glasskiva och, med hjälp av en skalpell, ta bort trichomes runt äggstockarna för att få en bättre visualisering av pollenrören. Sedan squash pistillerna med ett täckglas.
- Förbered 0,1% (v / v) aniline blå fläck: blanda 0,1 ml aniline blå i 100 ml 0,1 N kaliumfosfat tribasic (K3PO4). Applicera en droppe aniline blå över förberedelserna för att färga callose depositions under pollen rörtillväxt.
- Observera pollenrören längs stilen med ett mikroskop med UV-epifluorescens med 340-380 bandpass och 425 långpassfilter.
2. DNA-extraktion
- Prov 2-3 blad på fältet på våren. Det rekommenderas att prov bladen i unga stadier eftersom DNA erhålls är av högre kvalitet och lägre nivåer av fenolföreningar jämfört med gamla blad.
- Extrahera genomiskt DNA enligt de steg som beskrivs i ett kommersiellt tillgängligt kit (se Tabell över material).
- Analysera mängden och kvaliteten på DNA-koncentrationer med UV-vis spektrofotometer (260 nm).
3. S-allel identifiering
-
Inställning av PCR-reaktioner
- Bered en 50 ng/μL utspädning i destillerat vatten av varje DNA-extraktionsprov.
- Tina långsamt upp PCR-reagenserna och förvara dem på is. Lämna DNA-polymerasen i frysen tills det behövs.
- Förbered förstärkningsreaktionerna med hjälp av de olika kombinationerna av grundfärger. Skapa PCR-reaktionsmixen genom att kombinera komponenterna i tabell 1. Vortex PCR-reaktionsblandningen väl och fördelar volymen som indikeras för de olika kombinationerna av grundfärger till varje brunn av PCR-plattan. Tillsätt sedan 1 μL av DNA-utspädningen i varje brunn.
- Placera PCR-plattan i termohjulet och kör motsvarande PCR-program som visas i tabell 1.
- Analysera de förstärkta fragmenten. Det finns främst två olika sätt att analysera PCR-förstärkta fragment: kapillärelektrofores (CE) med fluorescerande märkta grundfärger eller som visualisera amplicons av agaros gelelektrofores med inte-märkta grundfärger.
-
Kapillärelektrofores
- För att förbereda lastbufferten, blanda 35 μL av joniserad formamid med 0,45 μL märkvärdstandard. Vortex reagensen för att blanda väl, och sedan fördela 35,5 μL i brunnen på läsaren plattan.
- Tillsätt 1 μl av PCR-produkten i brunnen. Dessutom tillsätt en droppe mineralolja för att förhindra vattenavdunstning.
- Förbered separationsplattan och lägg till separationsbuffert.
- Använd den kommersiella programvara som medföljer genanalysatorn (se Tabell över material). Skapa en ny provplatta och spara provnamnen för alla brunnar på plattan.
- Välj analysmetod. I detta fall, denaturera proverna vid 90 °C för 120 s, injicera vid 2,0 kV för 30 s och separera vid 6,0 kV i 35 min.
- Sätt in de två plattorna i genanalysatorn. Fyll kapillärmatrisen med destillerat vatten.
- Ladda den patenterade linjära polyakrylamiden (LPA) gelen. Klicka slutligen på Kör.
-
Gel Elektrofores
- Förbered en 1% agaros gel lägga 1,5 g molekylärbiologi grad agaros i 150 ml 1x TAE (Tris-acetat-EDTA) elektrofores kör buffert (40 mM Tris, 20 mM ättiksyra och 1 mM EDTA vid pH 8,0). Lös upp agaroset med mikrovågsuppvärmning i 2-3 min.
- För att visualisera DNA, tillsätt 4 μL av en nukleinsyra fläck (se Tabell över material) och blanda försiktigt.
- Tillsätt en gelkam, med tillräckligt med brunnar för stegar, kontroller och prover, i en gelbricka. Häll sedan långsamt blandningen i mitten av gelbrickan och undvik bubblor.
- Låt gelen svalna i 30-45 min i rumstemperatur tills gelen är helt stelnad. Sätt in gelen i elektroforeskammaren, ta bort gelkammen och fyll kammaren med tillräckligt med 1x TAE-buffert för att täcka gelen.
OBS: Kontrollera gelens placering. Brunnarna bör placeras nära den negativa polen eftersom negativt laddade DNA migrerar mot katoden. - Tillsätt 5 μl lastbuffert (0,1 % (v/v) bromfenolblå) till PCR-produkterna och blanda väl.
- För att uppskatta storleken på banden, ladda 5 μl DNA-molekylviktstege (se Tabell över material).
- Ladda proverna i gelens extra brunnar.
- När alla prover och DNA-molekylviktsstegen är lastade, kör gelen vid 90 V i 1-1,5 timmar, tills den blå färglinjen är ungefär 75% av gelens längd.
- Visualisera banden i en transilluminator för nukleinsyror.
-
Kapillärelektrofores
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Pollineringsstudier i aprikos kräver användning av blommor i slutet av ballongstadiet en dag före antes (figur 1A). Detta skede anses vara den mest gynnsamma för både pollen och fisktillsamling, eftersom blomstrukturer är nästan mogna, men anther dehiscence har ännu inte inträffat. Detta förhindrar inblandning av oönskad pollen, inte bara av pollen från samma blomma utan också från andra blommor, eftersom de slutna kronbladen hindrar ankomsten av insekter som bär yttre pollen. Pollenkornen är lätt siktade genom ett finmaskigt(figur 1B)från dehisced anthers som tidigare placerats på ett papper för 24 timmar vid rumstemperatur eller med lätt extra värme. På samma sätt erhålls pistiller från blommor i ballongstadiet efter avlägsnande av kronblad, foderblad och ståndare med hjälp av pincett eller naglar (figur 1C). Pistiller kan vara själv- och korspollinerade med en fin borste (figur 1D).
Hermafrodditiska blommor av aprikos har fem mörkröda foderblad, fem vita kronblad (figur 1A), en enda pistill (figur 2A) och 25-30 ståndare. Pistillen har tre huvudstrukturer: stigma, stil och äggstock. Äggstocken har två ägglossning, och befruktningen av minst en av dem krävs för fruktinställning. Under pollinering överför insekter, främst bin, pollenkorn till stigmatiseringen (figur 1A), där de gror (figur 2B) inom 24 timmar efter pollinering. Ett pollenrör produceras från varje groning pollenkorn, som växer genom pistillstrukturerna för att nå äggstocken efter 3-4 dagar och befrukta en av de två ägglossarna efter cirka 7 dagar. I själv oförenliga sorter där S-allelen av pollenkornet är densamma som en av de två av pistillerna, slutar pollenröret växa i övre stil, förhindra befruktning (figur 2C). S Pollenrören från en kompatibel sort, med en annan S-allel, kan dock växa genom stilen (figur 2D), nå äggstocken (figur 2 S E) och befrukta en av de två ovulerna.
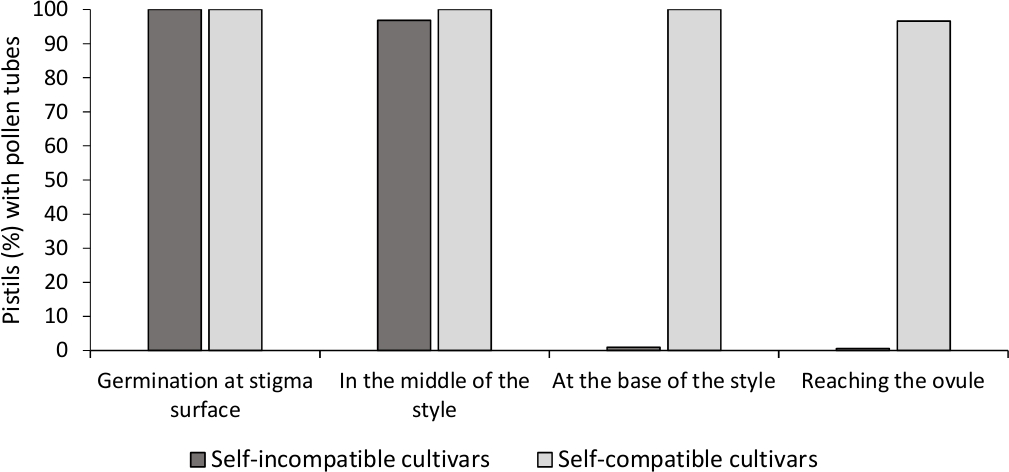
Analysen av in vitro pollengroning visade god pollen livsduglighet i alla sorter analyseras här, eftersom de flesta pollen rör var längre än längden på pollen korn efter 24 h i odlingsmediet. Grodda pollenkorn observerades vid stigmatytan(figur 2B)i pistiller från alla pollineringar, vilket tyder på adekvat pollinering (figur 3).
För att bestämma själv-(i)kompatibilitet för varje sort, pollen rör beteende i själv- och korspollineringar görs i laboratoriekontrollerade förhållanden observerades under fluorescensmikroskopi. Pollenröret tillväxt registrerades längs stilen i alla pistiller undersökta. Sorter ansågs vara självkompatibla när pollenrörstillväxten greps längs stilen i de flesta självpollinerade pistiller (figur 2C, figur 3) och självkompatibel när minst ett pollenrör nådde basen för stilen i de flesta av de pistiller som undersökts (figur 2E, figur 3).
Studien av S-locusen vid PCR-analys tillåtet att karakterisera S-genotypen av varje sort. För det Sförsta identifierades S-allelerna genom förstärkning av den första S-RNase-intronen med hjälp av grundfärgerna SRc-F/SRc-R (tabell 2). Storleken på de förstärkta fragmenten analyserades av kapillärelektrofores (figur 4A) och användes för att klassificera de genotyper som analyserades i motsvarande inkompatibilitetsgrupp (I.G.) (Tabell 3).
Vissa par av alleler, såsom S1 och S7 eller S6 och S9, visade liknande fragmentstorlekar för den första intronen. Differentieringen av dessa alleler gjordes således genom att förstärka en region i RNases andra intron med grundfärgerna Pru-C2/PruC4R, SHLM1/SHLM2 och SHLM3/SHLM4 (tabell 2). PruC2/PruC4R primer-kombinationen användes för att skilja mellan S6 och S9. För S6förstärktes ett fragment på 1300 bp medan ett fragment på omkring 700 bp observerades för S9-allelen ( figur4B, tabell 3). De specifika grundfärgerna SHLM1/SHLM2 och SHLM3/SHLM4 förstärkte ett fragment på cirka 650 bp i S1-allelen och 413 bp i S7-allelen ( figur4C, Tabell 3).
De primers AprFBC8-(F/R) som förstärker V2 och HVB variabla regioner i SFB genen användes för att skilja Sc och S8 alleler eftersom båda allelerna visar identiska RNase sekvens. S8-allelen visade ett PCR-fragment på cirka 150 bp medan ett fragment på 500 bp motsvarade Sc-allelen (figur 4D, tabell 3).c När SS-genotypen fastställdes för alla sorter, självokompatibla sorter tilldelades deras motsvarande inkompatibilitetsgrupper baserat på deras S-alleler (tabell 3). S
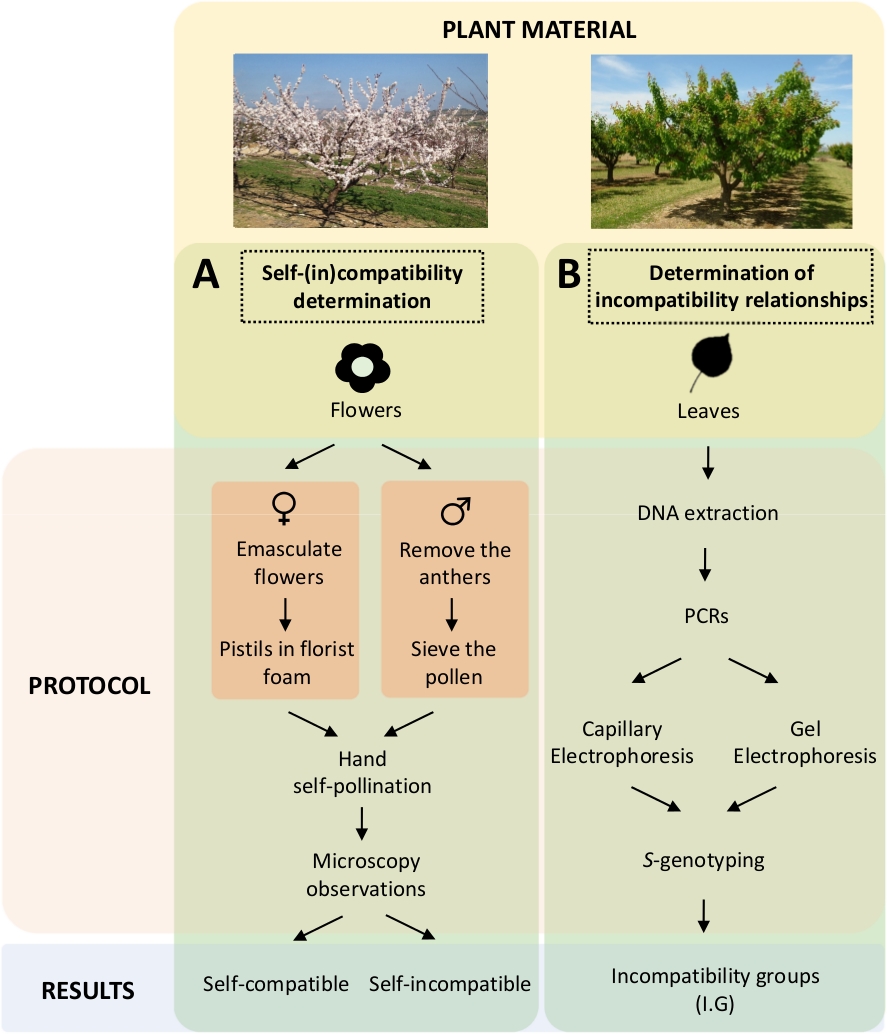
Detta tillvägagångssätt kräver att man fastställer varje sorters själv-(in)kompatibilitet genom kontrollerade själv- och korspollineringar i laboratoriet (figur 5A)samtidigt med karakteriseringen av S-genotypen genom genetisk analys (figur 5B). S Som ett resultat kan pollineringskraven för varje sort och inkompatibilitetsrelationer mellan aprikossorter bestämmas.

Figur 1. Experimentell uppsättning för bestämning av själv-(i)kompatibilitet i aprikos.
(A) Blommor på ballong scenen (svarta pilar) i fältet. B)Sikt av pollenkorn med hjälp av ett finmaskigt nät. (C) Pistiller placeras på florist skum i vatten. (D)Handpollinering av pistillerna med hjälp av en pensel. Klicka här för att se en större version av denna siffra.

Figur 2. Diagrammatisk representation av gametophytic inkompatibilitet relationer i aprikos blommor.
(A)I Gametophytic Self-Inkompatibilitet (GSI), både kompatibla och oförenliga pollenkorn grorar på stigmatisering. Pollenkornet bär en av två S-alleler av den ursprungliga genotypen, i detta fall antingen S1 eller S2. S Om SS-allelen av pollenkornet matchar en av de två S-allelerna i pistillen, i detta fall S1S3, hämmas pollenrörstillväxten i den övre tredjedelen av stilen. S B)Grobarhet av pollenkorn på stigmatytan. (C) Pollen röret greps i den stil som anger ett oförenligt beteende. (D)Pollenrör växer längs stilen. (E) Pollenrör vid basen av den stil som anger ett kompatibelt beteende. Skala barer, 100 μm. Klicka här för att se en större version av denna siffra.

Figur 3. Representativa resultat av pollengroning och pollenrör tillväxt genom stil för självkompatibla och själv-oförenliga sorter efter självpollineringar.
Andel pistiller med pollenkorn som gror på stigmatytan, med pollenrör i halvvägs stil, vid basen av stilen, och nå äggulan. Klicka här för att se en större version av denna siffra.

Bild 4. PCR fragment förstärkning med hjälp av fem primer Spar kombinationer för identifiering av S-alleler.
(A) Genanalysator utgång för SRc-(F /R) grundfärger som visar storleken på de två förstärkta fragment av RNase första intron regionen motsvarar S-allelerna. S (B)PCR förstärkning med hjälp av primers PruC2/PruC4R för identifiering av S6 och S9 alleler. (C)PCR-produkter som erhållits med hjälp av de specifika grundfärgerna SHLM1 och SHLM2 för differentiering av S1-allelen och SHLM3 och SHLM4 för att skilja S7-allelen. (D)PCR-förstärkning med AprFBC8-(F/R) grundfärger för identifiering av Sc- och S8-alleler. MI:1 kb DNA Stege. MII: 100 bp DNA Stege. Klicka här för att se en större version av denna siffra.

Figur 5. Schema för den experimentella designen för att klarlägga kompatibilitetsförhållandena mellan sig och mellan (i) i aprikossorter.
a)Arbetsflöde för självkompatibilitetsbestämning genom kontrollerade pollineringar i laboratoriet. (B)Arbetsflöde av S-allelidentifieringen vid molekylärt att närma sig. Klicka här för att se en större version av denna siffra.
| PCR Master Mix | Termocyklar | ||||||
| Komponenter | Slutlig koncentration | 15 μL reaktion | Cykelsteg | Temperatur | Tid | Cykler | |
| 10x NH4 reaktionsbuffert | 10x (10x) | 1,5 μl | Inledande denatration | 94 °C | 3 min | 1 | |
| 50 mM MgCl2 Lösning | 25 mM | 1,2 μl | Denatureringen | 94 °C | 1 min | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 μl | Glödgning | 55 °C | 1 min | ||
| Primer SRc-F | 10 μm | 0,6 μl | Förlängning | 72 °C | 3 min | ||
| Primer SRc-R | 10 μm | 0,6 μl | Slutlig förlängning | 72 °C | 5 min | 1 | |
| 500 U Taq DNA-polymeras | 0.5 U | 0,2 μl | 4 °C | Hålla | |||
| H2O | 8,3 μl | ||||||
| Komponenter | Slutlig koncentration | 25 μl reaktion | Cykelsteg | Temperatur | Tid | Cykler | |
| 10x PCR-buffert | 10x (10x) | 2,5 μl | Inledande denatration | 94 °C | 2 min | 1 | |
| 5x Q-lösning | 5x (5x) | 5 μl | Denatureringen | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 μl | Glödgning | 55 °C | 2 min | ||
| Primer PruC2 | 10 μm | 0,2 μl | Förlängning | 68 °C | 2 min | ||
| Primer C4R | 10 μm | 0,2 μl | Denatureringen | 94 °C | 10 s | 25 | |
| 250 U Taq DNA-polymeras | 10 U | 0,13 μl | Glödgning | 58 °C | 2 min | ||
| H2O | 15,5 μl | Förlängning* | 68 °C | 2 min | |||
| Slutlig förlängning | 72 °C | 5 min | 1 | ||||
| 4 °C | Hålla | ||||||
| * med 10 s läggas varje cykel till 68 %C förlängning steg. | |||||||
| Komponenter | Slutlig koncentration | 25 μl reaktion | Cykelsteg | Temperatur | Tid | Cykler | |
| 10x PCR-buffert | 10x (10x) | 2,5 μl | Inledande denatration | 94 °C | 2 min | 1 | |
| 5x Q-lösning | 5x (5x) | 5 μl | Denatureringen | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 μl | Glödgning | 62 °C | 1,5 min | ||
| Primer SHLM1 | 10 μm | 0,2 μl | Förlängning | 72 °C | 2 min | ||
| Primer SHLM2 | 10 μm | 0,2 μl | Slutlig förlängning | 72 °C | 5 min | 1 | |
| 250 U Taq DNA-polymeras | 10 U | 0,13 μl | 4 °C | Hålla | |||
| H2O | 15,5 μl | ||||||
| Komponenter | Slutlig koncentration | 20 μL reaktion | Cykelsteg | Temperatur | Tid | Cykler | |
| 5x PCR-buffert | 5x (5x) | 4 μl | Inledande denatration | 98 °C | 30 s | 1 | |
| dNTP (DNTP) | 2,5 mM | 1,6 μl | Denatureringen | 98 °C | 10 s | 35 | |
| Primer SHLM3 | 10 μm | 1 μl | Glödgning | 51 °C | 30 s | ||
| Primer SHLM4 | 10 μm | 1 μl | Förlängning | 72 °C | 1 min | ||
| 100 U DNA Polymeras | 5 U (på) | 0,2 μl | Slutlig förlängning | 72 °C | 5 min | 1 | |
| H2O | 12,4 μl | 4 °C | Hålla | ||||
| Komponenter | Slutlig koncentration | 25 μl reaktion | Cykelsteg | Temperatur | Tid | Cykler | |
| 10x PCR-buffert | 10x (10x) | 2,5 μl | Inledande denatration | 94 °C | 2 min | 1 | |
| 100 mM dNTP | 2,5 mM | 2 μl | Denatureringen | 94 °C | 30 s | 35 | |
| Primer FBC8-F | 10 μm | 1 μl | Glödgning | 55 °C | 1,5 min | ||
| Primer FBC8-R | 10 μm | 1 μl | Förlängning | 72 °C | 2 min | ||
| 250 U Taq DNA-polymeras | 10 U | 0,125 μL | Slutlig förlängning | 72 °C | 5 min | 1 | |
| H2O | 17,4 μl | 4 °C | Hålla | ||||
Tabell 1. Reaktions- och cykelförhållanden för olika primerkombinationer som används i detta protokoll.
| Grundfärger | Sekvens | Referens |
| SRc-F | 5'-CTCGCTTTCCTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Beskär-C2 | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCATCTACTCTTAG-3' | 26 |
Tabell 2. Primers används i detta protokoll, sekvens och referens för S-genotyp karakterisering i Prunus armeniaca.
| Sorten | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotyp | Inkompatibilitetsgrupp (I.G) |
| Undrar Cot8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Magiska Barnsäng8 | 334, 420 | 749 | S2S9 | Xx | |||
| Guldstrike8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ⅰ | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| Cooper Cot8 (På andra) | 274, 408 | 650 | S1S3 | Xviii | |||
| Apriqueen (apriqueen) | 358, 358 | 500 | ScSc | - | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | - | |||
| Våren Blush8 | 274, 358 | 150 | S3S8 | Xxi |
Tabell 3. S-genotypning av aprikossorter med fem primerpar som används i detta protokoll och inkompatibilitetsgrupptilldelning. De olika polymeras kedjereaktionsproduktstorlekarna för S-alleler som förstärks med SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 och AprFBC8-(F/R) primers visas i tabellen. S
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Traditionellt var de flesta kommersiella aprikos europeiska sorter självkompatibla36. Ändå har användningen av nordamerikanska själv oförenliga sorter som föräldrar i avelsprogram under de senaste decennierna resulterat i frisläppandet av ett ökande antal nya själv oförenliga sorter med okänd pollinering krav7,8,37. Således är fastställandet av själv- och inter-(in)kompatibilitetsrelationer i aprikossorter allt viktigare. Detta accentueras i de områden där vintern kylning minskar, eftersom hög år till år variationer i tiden för blomning hindrar sammanträffande i blomningen av sorter och deras pollenizers i många fall, särskilt i sorter med höga kylning krav38. Den metod som beskrivs häri, som kombinerar handpollinerande, mikroskopi och genetiska analyser har varit mycket användbar för att bestämma själv(i)kompatibilitet för varje sort och att fastställa dess potentiella pollinerare sorter.
Pollineringskraven kan fastställas genom fältkontrollexperiment under fruktodlingsförhållanden11,39. Utläggningen till yttre faktorer inklusive meteorologiska ogynnsamma förhållanden kan dock orsaka pollinering misslyckande10, vilket kan resultera i felaktiga diagnoser av självinkompatibilitet. Den metod som beskrivs häri gör det möjligt att utvärdera själv-(i)kompatibilitet mer exakt genom mikroskopi observationer av handpollinerade blommor i laboratoriekontrollerade förhållanden, undvika miljöpåverkan. Dessutom gör detta tillvägagångssätt att analysera ett högre antal sorter per år, eftersom endast ett litet antal blommor krävs i stället för flera vuxna träd för varje sort som krävs i fältexperiment40.
Inkompatibilitetsrelationer kan upprättas som kombinerar handpollinationer och mikroskopi14. Pollinationer kan dock endast utföras under en kort period under blomningssäsongen på våren, och vuxna träd nära laboratoriet behövs, eftersom livslängden på de insamlade blommorna är mycket kort. Således är antalet inkompatibilitetsrelationer som kan analyseras genom kontrollerade handpollinationer under varje säsong mycket lågt. Karakteriseringen av de gener som kodas av S-locushar möjliggjort utvecklingen av PCR-baserade metoder för S-allelgenotypning18,41. Detta tillvägagångssätt Saccelererar S-allel identifiering eftersom det inte kräver blommor, och experimenten kan utföras med någon vegetativ vävnad42. Detta förlänger den period under vilken växtmaterial, vanligtvis unga blad, kan samlas in43. Dessutom kan bladen lyofiliseras eller frysas, så att analysen kan göras när som helst på året, till skillnad från pollineringar som endast kan göras på färska blommor under blomningssäsongen44. En ytterligare fördel är att löv kan samlas in från unga träd redan innan blomning ålder, underlätta insamling av prover och tidig erhålla resultat45.
Den genetiska analysen möjliggör en bättre differentiering av självinkompatibilitetsakreter eftersom den ger exakta resultat av amplifierade fragmentstorlekar21,46. Hittills har trettiotre S-alleler identifierats i aprikos12,,18,20,,21,,22,,23,24, som har gjort det möjligt att upprätta 36 inkompatibilitetsgrupper baserade på S-genotyp8,,9,,17,, S25,,26,27. Å andra sidan är en nackdel med denna metod att olika alleler i samma intervallstorlek eller mutationer felaktigt kan identifieras som samma allel. Således är Sc och S8 alleler identiska för RNase sekvensen men en 358-bp insättning finns i SFB genen i Sc19. På samma sätt är den första intronregionen i allelerna S1 och S7 identiska och omöjliga att skilja från med hjälp av grundfärgerna SRc-F/SRc-R. Dessutom har flera homologer, såsom S6 och S528 eller S20 och S55, och S7, S13 (EF062341) och S4617, hittats eftersom vissa av dessa alleler delvis har sekvenserats eller av fel under PCR-förstärkning och följaktligen krävs ytterligare arbete för att särskilja dem korrekt.
PCR-analys och S-RNase-sekvensering är tillräckliga för att upprätta oförenlighetsförhållanden genom identifiering av S-alleler Soch tilldelning av sorter i motsvarande inkompatibilitetsgrupp8,,17,,26,27. Denna metod har dock begränsningen att förhindra att självkompatibiliteten för särskilda aprikossorter fastställs. Självkompatibilitet (SC) har associerats till särskilda S-alleler i andra Prunus arter47, som mandel(Sf)48,,49 eller sötkörsbär(S4')50,51. S Men i aprikos, Sc allel, som har associerats till SC21, kan felaktigt identifieras som S8, en själv-oförenliga allele19,22, och eventuella mutationer som inte är kopplade till S locus, som M-locus12,52, conferring SC har identifierats. Nyligen har MM-locus genotypats med SSR-markörer12. Därför behöver den genetiska identifieringen av SC för aprikos genotyper ytterligare forskning och, för att undvika misstag på grund av faktorer som inte är kopplade till S locus, i detta arbete karakterisering av själv-(i)kompatibilitet har också fastställts genom att fenotypera beteendet hos pollenrören genom pistill av självpollinerade blommor.
Den metod som beskrivs häri som kombinerar bestämning av självpollinentering genom handpollinationer under laboratorieförhållanden med efterföljande observation av pollenrörens beteende i pistillen Sav kontrollerade självpollinerande ämnen under fluorescensmikroskopin och identifiering av S-genotyp genom PCR-analys gör det möjligt att fastställa pollineringskraven för aprikossorter. Detta ger en värdefull information för odlare och uppfödare, eftersom det gör det möjligt att upprätta oförenlighet relationer mellan sorter att välja lämpliga pollinatörer i utformningen av nya fruktträdgårdar samt att välja lämpliga föräldrar att utforma nya kors i aprikos avelsprogram.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna har inget att avslöja.
Acknowledgments
Denna forskning finansierades av Ministerio de Ciencia, Innovación y Universidades-European Regional Development Fund, Europeiska unionen (AGL2016-77267-R och AGL2015-74071-JIN); Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-00); Gobierno de Aragón-Europeiska socialfonden, Europeiska unionen (Grupo Consolidado A12_17R), Fundación Biodiversidad och Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R. Mechanisms of self-incompatibility in flowering plants. Cellular and Molecular Life Sciences. 58, 1988-2007 (2001).
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}