

Summary
Seriell blokk-ansiktsskanning Elektronmikroskopi (SBEM) påføres bildet og analyserer dendrittiske spines i murin hippocampus.
Abstract
Tredimensjonal elektronmikroskopi (3D EM) gir en mulighet til å analysere morfologiske parametere av dendritiske spines med nanoskala oppløsning. I tillegg kan noen funksjoner i dendrittiske ryggrader, for eksempel volum av ryggraden og postsynaptisk tetthet (PSD) (som representerer postsynaptisk del av synapsen), tilstedeværelse av presynaptisk terminal og glatt endoplasmisk retikulum eller atypisk form for PSD (f.eks. multi-innerverte spines), bare observeres med 3D EM. Ved å bruke seriell blokk-ansikt skanning elektronmikroskopi (SBEM) er det mulig å skaffe 3D EM-data enklere og på en mer reproduserbar måte enn ved utførelse av tradisjonell seriell seksjonering. Her viser vi hvordan du forbereder mus hippocampal prøver for SBEM analyse og hvordan denne protokollen kan kombineres med immunfluorescence studie av dendritiske spines. Mild fikseringsperfusjon gjør at vi kan utføre immunfluorescensstudier med lett mikroskopi på den ene halvdelen av hjernen, mens den andre halvparten var forberedt på SBEM. Denne tilnærmingen reduserer antall dyr som skal brukes til studien.
Introduction
De fleste eksitatoriske synapsene i sentralnervesystemet ligger på dendritiske spines - små fremspring av en nevronmembran. Disse fremspringene danner begrensede biokjemiske rom som styrer intracellulær signaltransduksjon. Strukturell plastisitet av dendritiske spines og synapser er nært knyttet til funksjonelle endringer i synaptisk effekt som ligger til grunn for slike viktige prosesser som læring og minne1,2. Det er viktig å merke seg at elektronmikroskopi (EM) er den eneste teknikken som gjør det mulig å avgjøre om en dendritisk ryggrad har en presynaptisk inngang. EM-oppløsning er også nødvendig for å studere ultrastrukturelle detaljer som form av postsynaptisk tetthet (PSD), som representerer en postsynaptisk del av en synapse, eller dimensjoner av en dendritisk ryggrad, samt størrelsen og formen på en axonal bouton. I tillegg, med EM er det mulig å visualisere synapser og deres omgivelser.
Takket være fremskritt innen bilde- og datateknologi er det mulig å rekonstruere hele nevrale kretser. Volumelektronmikroskopiteknikker, for eksempel seriell seksjonsoverføringselektronmikroskopi (ssTEM), seriell blokk-ansiktsskanning elektronmikroskopi (SBEM), og fokusert ionstråleskanning elektronmikroskopi (FIB-SEM) brukes vanligvis til nevronale kretsrekonstruksjoner3.
I våre studier er SBEM-metoden vellykket brukt til å undersøke den strukturelle plastisiteten til dendritiske spines og PSDs i prøver av musen hippocampus og organotypiske hjerneskiver 4,5. SBEM er basert på installasjon av en miniatyr ultramikrotomi inne i skanning elektron mikroskop kammer6,7,8,9. Toppen av prøveblokken er avbildet, og deretter blir prøven kuttet på en bestemt dybde av ultramikrotomet, og avslører en ny blokk-ansikt, som igjen er avbildet og deretter prosessen gjentas8. Som et resultat er bare bildet av en blokk-ansikt igjen mens stykket som er kuttet, går tapt som rusk. Det er derfor SBEM kalles en destruktiv teknikk, noe som betyr at det ikke er mulig å bilde samme sted igjen. Fordelen med de destruktive on-block-metodene er imidlertid at de ikke lider av fordreiningsproblemer og seksjonstap som kan påvirke datakvaliteten og dataanalysen3betydelig . Videre gir SBEM muligheten til å bilde et relativt stort synsfelt (> 0,5 mm × 0,5 mm) ved høy oppløsning3.
For å bruke SBEM må prøver tilberedes i henhold til en dedikert, svært kontrasterende protokoll på grunn av den bakscattered elektrondetektoren som brukes til å skaffe bilder. Vi viser her hvordan du utfører prøvepreparering i henhold til protokollen basert på en prosedyre utviklet av Deerinck10 (National Center for Microscopy and Imaging Research (NCMIR) metode), ved hjelp av redusert osmium-tiocarbohydrazide-osmium (rOTO) flekker utviklet på 1980-tallet8,11. I tillegg introduserer vi en to-trinns fiksering tilnærming, med mild fiksering perfusjon som gjør det mulig å bruke samme hjerne både for immunfluorescence studier med lys mikroskopi og SBEM.
I protokollen er en musehjerne primært festet med et mildt fikseringsmiddel, og deretter kuttet i halvdeler, og en halvkule er postfixed og forberedt på immunfluorescence (IF), mens den andre for EM-studier (Figur 1).

Figur 1. Skjematisk representasjon av arbeidsflyten for klargjøring av dendrittiske ryggrader for analysen med SBEM. Mus ble ofret og perfundert med et mildt primært fikseringsmiddel. Hjernen ble kuttet i halvdeler, og en halvkule ble postfixed med immunfluorescence (IF)-dedikert fixative, kryorotected, skiver ved hjelp av en kryostat og behandlet for IF-studier, mens den andre halvkule ble postfixed med EM fixative, skiver med vibratom og forberedt for EM-studier. Hjerneskiver for SBEM-studier ble kontrastert, flat innebygd i harpiks, deretter ble en CA1-region av hippocampus montert på pinnen, og avbildet med SBEM (Figur 1). Den delen av protokollen som er uthevet i en gul boks, ble omtalt i videoen. Klikk her for å se en større versjon av denne figuren.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Forskningen ble utført i samsvar med Nencki Institutes retningslinjer og tillatelse fra Den lokale etiske komité. Studiene ble utført i samsvar med Det europeiske fellesskapsråds direktiv av 24. Alle anstrengelser ble gjort for å minimere antall dyr som ble brukt og deres lidelse.
FORSIKTIG: Alle prosedyrer beskrevet nedenfor må utføres i en laboratorierøykhette. På grunn av reagensenes farlige natur. Personlige sikkerhetstiltak som hansker, labfrakk, vernebriller og munnbind er nødvendig.
1. Fremstilling av fikseringsmiddelet for perfusjon (2% wt/vol paraformaldehyd (PFA) og 0,5% vol/vol glutaraldehyd (GA) i 0,1 M fosfatbuffer (PB), pH 7,4)
MERK: Forbered den fikseringsløsningen på samme dag som den skal brukes, og ikke oppbevar den lenger enn i 3 timer. Ved tidsmangel, forbered 2% PFA i 0,1 M PB dagen før, lagre den ved 4 °C og tilsett fersk GA kort tid før perfusjonen.
- Ta 400 ml sterilt dobbelt destillert vann (ddH2O) og varm det opp til 60 °C ved hjelp av en omrørende kokeplate. Tilsett deretter 20 g PFA. Tilsett dråper på 1 M NaOH til PFA er helt oppløst og la blandingen avkjøles.
- Tilsett 500 ml 0,2 M PB (pH 7,4).
- Filtrer oppløsningen for å fjerne eventuelle avleiringer og avkjøl den til 4 °C.
- Like før perfusjon, tilsett 20 ml 25% GA til løsningen og fyll deretter opp volumet med ddH2O til 1 L.
2. Fremstilling av postperfusjonsfiksiv for SBEM (2% wt / vol PFA og 2,5% vol / vol GA i 0,1 M PB, pH 7,4)
- Ta 50 ml av fixativet for perfusjon (2% PFA og 0,5% GA i 0,1 M PB).
- Tilsett 5 ml 25 % GA.
3. Fremstilling av postperfusjonsfiksiv for IF-farging (4% PFA i fosfatbufret saltvann (PBS))
- Løs opp en tablett med 1x PBS (pH 7.4) i et renset og deionisert vann (H2O) i henhold til produsentens instruksjoner.
- Bruk en omrørende kokeplate for å varme opp oppløsningen til 60 °C og tilsett 40 g PFA.
- Tilsett dråper på 1 M NaOH til PFA er fullstendig oppløst og la blandingen avkjøles.
- Juster pH-en til løsningen til 7,5 med 1 M HCl og fyll deretter volumet med H2O til 1 L.
- Filtrer løsningen for å fjerne eventuelle innskudd.
4. Transkardiale perfusjon av dyr
MERK: Alt PFA- og GA-avfall må samles inn og oppbevares for avhending i henhold til lokale forskrifter. Anestesi og perfusjon bør følge lokale forskrifter. I den beskrevne protokollen voksne 3 måneder gamle og 20±1 måneder gamle kvinnelige Thy1-GFP (M) mus (Thy1-GFP +/-)12 uttrykker grønt fluorescensprotein (GFP) i en tynt distribuert populasjon av glutamatergiske nevroner ble brukt, men alle andre kan også brukes. Dyr ble oppdrettet som heterozygoter med C57BL /6J-bakgrunn i Dyrehuset til Nencki Institute of Experimental Biology.
- Før perfusjon bedøver du en mus ved administrering av en ketamin/xylazinblanding (opptil 90 mg/kg kroppsvekt ketamin og 10 mg/kg kroppsvekt xylazin) via intraperitoneal injeksjon (27-gauge nål).
- Vurder om dybden av anestesi er tilstrekkelig ved å sjekke refleksen til smertestimuli (klemming) og hornhinnen refleks (mysing).
- Etter 20 minutter utføre en intraperitoneal injeksjon (27-gauge nål) av natrium pentobarbital (50 mg / kg kroppsvekt).
- Perfuse en mus i henhold til en perfusjon kirurgi protokoll beskrevet av Gage et al.13 (se punkt 4; Figur 5-6). Ved hjelp av en perfusjonspumpe starter du med en 0,1 M fosfatbuffer, pH 7,4 (30 ml) i 3 minutter og fortsetter med 2 % PFA og 0,5 % GA på 0,1 M PB, pH 7,4 i 6 minutter (80 ml).
- Disseker hjernen forsiktig fra skallen og del den i to (se figur 9-10 i Gage et al., 201213). Legg ett stykke i et hetteglass som inneholder fikseringsmiddel for SBEM og det andre i hetteglasset med 4 % PFA/PBS for HVIS-farging.
- Hold halvkule i fikseringsmiddelet ved 4 °C over natten.
5. Hjerneskiver forberedelse for elektronmikroskopi
- Velg vibratominnstillinger (bladreisehastighet: 0,075 mm/s, skjærefrekvens: 80 Hz).
- Plasser skjærekammeret i holderen, fest det til vibratomet og omgi det med is. Plasser deretter et barberblad i vibratombladholderen.
- Bruk en skje, eller lignende gjenstand og plasser den avkjølte hjernen (dorsal overflate opp) på en tøff skjæreflate (f.eks. et petriskållokk i glass). For å forberede en koronalskive av hippocampus gjør et vinkelrett kutt mellom hjernehalvøya og cerebellum med barberblad eller skalpell, og dermed fjerne cerebellum. Den olfaktoriske pæren kan også fjernes.
- Påfør cyanoakrylatlim på den tørre plattformen til vibratomet.
- Plukk opp hjernen med tang og tørk den forsiktig på filterpapir.
- Lim halvkule til plattformen nær skjærebladet med rostralspissen oppover. Fest plattformen til holderen og fyll den umiddelbart med iskald 0,1 M PB, pH 7.4. Hvis det vinkelrette kuttet er riktig laget, vil halvkule stå rett opp og gi 90 ° vinkel som kreves for å lage et symmetrisk koronalsnitt som inneholder hippocampus. Forsikre deg om at hjernen er dekket med PB.
- Plasser vibratombladet foran halvkule og senk det til koronalsiden av halvkule. Senk bladet til 400 μm videre i kaudal retning og begynn å kutte. Fortsett å kutte til de to første skivene er helt skilt fra vevsblokken.
- Trekk tilbake bladet og senk ytterligere 100 μm, og skjær deretter opp igjen.
- Når hippocampus blir synlig (bruk musen hjerne atlas Paxinos og Franklin, 200414) samle skiver med den lille pensel eller utvidet plast Pasteur pipette.
- Overfør skivene til en 12-brønns plate fylt med kald 0,1 M PB, pH 7,4.
- Disseker hippocampus i et glass Petri-tallerken fylt med 0,1 M PB, pH 7,4 ved hjelp av et barberblad eller skalpell, og legg det i hetteglass med samme fosfatbuffer.
MERK: For langtidslagring av skiver supplement 0,1 M PB med 0,05% natriumazid (NaN3).
6. Hjerneprøvepreparering for immunstaining
- Etter nattfiksering i 4% PFA / PBS-løsning i en avtrekkshette, legg hjernevevet i kryopreserveringsløsningen (30% sukrose i PBS med 0,05% NaN3) og hold det ved 4 °C i 2 dager (til det senkes).
- Forbered en frostvæskeløsning (15 % sukrose/30 % etylenglykol/0,05 % NaN3/PBS).
- Still inn kabinetttemperaturen på kryostaten ved -19 °C og sørg for at temperaturen er nådd før du fortsetter videre. Under seksjonering må du sørge for at kabinetttemperaturen forblir mellom -18 °C og -20 °C.
- Bruk en skje, eller lignende gjenstand og plasser den avkjølte hjernen (dorsal overflate opp) på en tøff skjæreflate (f.eks. et petriskållokk i glass). For å forberede en koronalskive av hippocampus gjør et vinkelrett kutt mellom hjernehalvøya og cerebellum med barberblad eller skalpell, og dermed fjerne cerebellum.
- Velg en forhåndskjølt prøveskive, dekk den med et medium for frysing på en frihylle og bruk tang fest halvkule til platen med en rostralspiss oppover og vent til prøven er helt frossen. Sett prøveskiven inn i et prøvehode.
- Monter et blad i en bladholder inne i kryokammeret og kutt 40 μm tykke skiver.
- Overfør skiver med en liten pensel til 96-brønnsplaten fylt med kald frostvæskeløsning og rull forsiktig ut seksjonene (samle en skive etter hver runde med kutting for å forhindre at de går seg vill i skjærekammeret).
MERK: Hjerner eller seksjoner kan lagres i en frostvæskeoppløsning ved -20 °C i lang tid.
7. Immunostaining av hjerneskiver
MERK: Alle fargingstrinn ble utført i en 24-brønnsplate på en plattform shaker.
- Vask skivene med PBS tre ganger, hver gang i 6 minutter.
- Inkuber skiver i 300 μL blokkeringsløsning (5% normalt eselserum (NDS)/0,3% Triton X-100) i 1 time med skånsom risting på en rotator.
- Inkuber skivene med det primære antistoffet mot PSD-95 fortynnet 1:500 i 5% NDS / 0,3% Triton X-100 / PBS (300 μL per brønn) over natten ved 4 °C. Den endelige konsentrasjonen av det primære antistoffet er 2 μg/ml.
- Vask skivene med 0,3% Triton X-100/PBS ved romtemperatur (RT) tre ganger, hver gang i 6 minutter.
- Inkuber skiver med sekundært antistoff fortynnet 1:500 i 300 μL av 0,3% Triton X-100 / PBS i 90 minutter. Den endelige konsentrasjonen av sekundært antistoff er 4 μg / ml.
- Vask skivene med PBS tre ganger, hver gang i 6 minutter.
- Monter stykkene på lysbilder ved hjelp av et monteringsmedium, og lukk dem deretter med en dekselsklie.
- Undersøk prøvene ved hjelp av et konfokalt mikroskop (63 × oljemål, NA 1,4, pikselstørrelse 0,13 μm × 0,13 μm).
MERK: For langvarig lagring: Oppbevar prøvene ved 4 °C, beskytt dem mot lys.
8. SBEM prøvepreparering
FORSIKTIG: På grunn av reagensens farlige natur må alle prosedyrene beskrevet nedenfor utføres i en laboratorierøykhette. Før du bruker disse kjemikaliene, må du lese nøye gjennom materialsikkerhetsdatabladene fra produsentene og spørre sikkerhetsansvarlig om de lokale reglene for å sikre sikker håndtering og avfallshåndtering.
- Eksempel på kontrastering
MERK: Skiver vasker og romtemperatur inkubasjon bør utføres med mild risting (f.eks. på en plattform shaker). Autoklavavgasset vann ble brukt.- Vask skivene med kaldt 0,1 M PB, pH 7,4 fem ganger, hver gang i 3 minutter.
- Forbered en 1:1 blanding av 4% vandig osmium tetroxide og 3% kalium ferrocyanid (1:1 av vol). Sluttproduktet blir brunt. Senk prøvene i denne blandingen, legg dem på is, og fra dette stadiet og utover er det viktig å beskytte dem mot lys. Inkuber dem med mild risting i 1 time.
- I mellomtiden forberede en tiokarbohydrazid (TCH) løsning. Bland 10 ml dobbeltdestillert H2O (ddH2O) og 0,1 g TCH og legg den i en ovn satt til 60 °C i 1 time. Det er viktig å virvle løsningen fra tid til annen (f.eks. hvert 10. minutt). Når den er klar, avkjøl den til romtemperatur.
- Vask skivene med ddH2O fem ganger, hver gang i 3 minutter.
- Filtrer TCH-oppløsningen med et 0,22 μm sprøytefilter og senk prøven ned i filtrert oppløsning. Skivene blir svarte. Inkuber dem i 20 minutter ved romtemperatur.
- Vask prøvene med ddH2O fem ganger, hver gang i 3 minutter.
- Inkuber prøvene med en 2% vandig løsning av OsO4 i 30 minutter ved romtemperatur.
- Vask prøvene med ddH2O fem ganger, hver gang i 3 minutter.
- Plasser prøvene i filtrert 1% vandig uranylacetat og inkuber dem ved 4 °C over natten. Bruk 0,22 μm sprøytefilter for filtrering.
- Forbered L-aspartinsyreoppløsning ved å blande 0,4 g L-aspartinsyre og 80 ml ddH2O, juster pH til 3,8 for enklere oppløsning, og fyll deretter på med vann til 100 ml.
- Neste dag begynner waltons ledelse som15 forberedelser. Bland 0,066 g blynitrat med 10 ml L-aspartiginsyreoppløsning (punkt 8.1.10) forvarmet til 60 °C og juster pH til 5,5 (målt ved 60 °C) med 1 M NaOH. Lukk hetteglasset med bly aspartat og la det stå ved 60 °C i 30 minutter i et vannbad. Løsningen skal være klar. Hvis det blir overskyet, må det kastes og en ny må være forberedt.
- I mellomtiden vasker du prøvene med avgasset ddH2O fem ganger, hver gang i 3 minutter. Hold dem deretter i ovnen satt til 60 °C i 30 minutter.
- Senk prøvene ned i den nylagde bly aspartatløsningen og inkuber dem i ovnen som er satt til 60 °C i 20 minutter.
- Vask prøvene med avgasset ddH2O fem ganger, hver gang i 3 minutter.
- Dehydrering og harpiksinnbygging
- Forbered epoksyharpiks. Vei ingrediensene (33,3 g komponent A/M, 33,3 g komponent B og 1 g komponent D) og bland harpiksbrønnen (f.eks. rist den på en roterende shaker i et 15 ml rør) i minst 30 minutter før du tilsetter 16 dråper akselerator DMP 30. Rør igjen i ytterligere 10 minutter.
MERK: Vei ingrediensene under avtrekkshetten. Denne mengden komponenter gir ca. 60 ml harpiks det som er nok for 15 hetteglass med prøver. Du kan tilberede mindre eller lagre resten av harpiksen i en sprøyte ved 4 °C og bruke den neste dag. Husk å forsegle spissen av sprøyten. - Forbered hetteglass med graderte fortynninger av etanol (30%, 50%, 70%, 90%, 100%, 100% etanol i vann og 100% tørket på en molekylær sikt).
- Bland harpiksen med 100% etanol i 1:1 andel for å få 50% harpiks. Bland det godt.
- Dehydrer prøvene i 5 minutter i hver fortynning av etanol fra 30% og slutter med 100% vannfri etanol tørket på molekylær sikt. Husk at prøvene aldri må tørke helt.
- Infiltrer prøvene først i 50% harpiks i 30 minutter, neste i 100% harpiks i en time, og deretter igjen i 100% harpiks over natten. Utfør alle infiltrasjonstrinn med konstant langsom risting.
- Neste dag legger du prøvene i frisk 100% harpiks i en time og legger dem deretter inn mellom fluoropolymerinnbyggingsplater. Avfette biter av innebyggingsark med etanol, og flat bygge inn prøvene mellom to lag av det, ved hjelp av to glasssklier som støtte. Prøv å unngå luftbobler i harpiks på eller i nærheten av prøven.
- Herd prøvene i ovnen ved 70 °C i minst 48 timer.
- Forbered epoksyharpiks. Vei ingrediensene (33,3 g komponent A/M, 33,3 g komponent B og 1 g komponent D) og bland harpiksbrønnen (f.eks. rist den på en roterende shaker i et 15 ml rør) i minst 30 minutter før du tilsetter 16 dråper akselerator DMP 30. Rør igjen i ytterligere 10 minutter.
- Trimming og montering
- Skill innbyggingsplatene og klipp et stykke av den innebygde prøven (ca. 1 mm x 1 mm) med et barberblad. Overfør den til en parafilm. Dette vil minimere faren for å miste prøven på grunn av elektrostatikk.
- Ta en aluminiumspinne som har blitt avfettet med etanol. Bland en ledende epoksybrønn og bruk litt av den til å montere prøven på pinnen. Herd ledende epoksy ved 70 °C i 10 minutter.
- Trim hver side av prøveblokken med diamantkniven og poler deretter ansiktet på blokken til vevet er utsatt.
MERK: På dette trinnet bekrefter du tilstedeværelsen av en interesseregion ved å samle noen seksjoner og utføre toluidinblå farging16 eller sjekke under elektronmikroskopet. - Reduser størrelsen på prøven så mye som mulig. Jord deretter prøven til pinnen med ledende maling og kurer den (i 24 timer ved romtemperatur eller i ovnen ved 65 °C i 40 minutter).
- For å minimere ladeartefakter, sputter frakk prøvene med et tynt lag gull eller gull / palladium.
9. SBEM-avbildning
- Plasser pinnen med en prøve i kammeret til det serielle blokk-ansiktsskanningselektronmikroskopet. Juster prøven etter kniven. Lukk kammeret og angi parametrene.
- Samle en stabel med bilder ved ønsket forstørrelse, pikselstørrelse, stykketykkelse, akselerert spenning (EHT), blenderåpning, trykk, etc.
MERK: Parametrene avhenger av prøven og målet med eksperimentet. Eksemplariske innledende bildebehandlingsinnstillinger er inkludert i materialtabellen.
10.3D rekonstruksjoner
MERK: For trinnene nevnt nedenfor bruker vi programvare med åpen tilgang, for eksempel FijiJ17 (ImageJ versjon 1.49b), Microscopy Image Browser (MIB)18 og Reconstruct19, men forskjellige andre programvarer kan brukes.
- Konverter digitale mikrograffiler (dm) til TIFF-format. Importer først bildesekvensen (i FijiJ: Fil > Importer > Bildesekvens > Velg 8-biters format) og lagre deretter som TIFF (i FijiJ: Fil > Lagre som > Bildesekvens > Velg Tiff).
- Juster lysstyrken og kontrasten til bildestakken (i FijiJ: Image > Juster lysstyrken/kontrasten >) og denoise den (bruk om nødvendig DenoisEM-plugin for FijJ20 : Plugins > DenoiseEM > Denoise).
- Juster stakken (i FijiJ: Plugins > StagReg eller MIB: Datasett > Justeringsverktøy).
- Segment dendrittiske spines og synapser (I MIB- eller Reconstruct-programvare er omfattende opplæringsprogrammer tilgjengelige (se Tabell over materialer for detaljer).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Ved hjelp av metoden beskrevet ovenfor høykontrast, kan du få bilder med god oppløsning av musens hjernevev. Et stort synsfelt levert av SBEM-teknikken letter presis utvelgelse av interesseområdet. Det store bildet av CA1-regionen i hippocampus ble tatt for å måle lengden på stratum radiatum (SR) (Figur 2A) og for å angi bildet nøyaktig i midten (Figur 2B). Deretter ble bunken med bilder anskaffet og gjenstandene av interesse, for eksempel dendritter, dendritiske spines, postsynaptiske tettheter av synapsene ble segmentert (Figur 2C, D).
Bruken av SBEM-teknikken gir muligheten til å analysere et relativt stort volum vev og gjenkjenne sjeldne hendelser som en multi-innervated dendritisk ryggrad (Figur 2E) - en bestemt type synapser som er preget av flere presynaptiske terminaler som kontakter samme postsynaptiske ryggrad21.

Figur 2. Dendritiske spines i mus hippocampus CA1-regionen. Representativt bilde av mus hippocampal CA1-regionen. Stratum radiatum (SR) vises med en hvit pil (A). Området midt i SR ble valgt for avbildning (B). Rekonstruksjon av et stykke dendrit med dendrittiske spines (C). Rekonstruksjon av dendrittiske spines i et volum av vev (D). Rekonstruksjon av en multi-innervated dendritisk ryggrad (E). To presynaptiske terminaler som kontakter samme postsynaptiske ryggrad vises (i magenta og grønt). Klikk her for å se en større versjon av denne figuren.
Den presenterte protokollen for SBEM prøvepreparering tillot oss å skaffe bilder av hjernevev av høy kvalitet med sterkt kontrasterte membraner, noe som muliggjør enklere segmentering og rekonstruksjon av membran-omringede strukturer. Standard fiksering med høyere konsentrasjon av PFA (Figur 3B) og to-trinns fiksering tilnærming resulterte i lignende vev morfologi (Figur 3A).

Figur 3. Morfologiske detaljer. Med bruk av protokollen presentert her god kvalitet bilder som viser morfologi av synaptiske vesikler, PSD og mitokondrier ble oppnådd. Hjernevev festet med to-trinns tilnærming (A) eller med en standard fixative (B). Skalalinje 2 μm. Klikk her for å se en større versjon av denne figuren.
Resultatene er reproduserbare, noe som betyr at hvert eksemplar som er utarbeidet ved hjelp av denne protokollen, ikke bare hjernevev, men også monolayer av dyrkede celler, var godt kontrastert.
Bruken av mild primær fiksering gir mulighet til å bruke vevet også til immunfluorescensstudier som involverer et lysmikroskop. Når det gjelder PSD-95-antistoffet, observerte vi ikke forskjellen i immunstainingsresultater mellom prøvene som er festet med totrinns fiksering (mild primær og deretter sekundær med et tradisjonelt fikseringsmiddel) og den som oppnås ved hjelp av tradisjonell fiksering med henholdsvis 4% PFA (figur 4A og 4B).

Figur 4. Sammenligning av PSD-95 immunfluorescens i prøver festet med ulike protokoller. Representative mikrofotografier av PSD-95 immunostaining i stratum radiatumlaget av CA1-området i dorsal hippocampus. (A) Konfokal bilde etter mild primær fiksering etterfulgt av postfixation med en standard fixative. (B) Konfektbilde etter standard, sterkere fiksering. Skalastenger: 5 μm. Klikk her for å se en større versjon av denne figuren.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Det er mange varianter av den primære NCMIR-metoden beskrevet av Deerinck i 201010. De grunnleggende prinsippene forblir de samme, men avhengig av hvilken type materiale som studeres, implementeres små endringer. Det ble tidligere beskrevet at forskjellige harpikser kan brukes til å legge inn prøver for SBEM, og for eksempel når det gjelder planter, er Spurrs den valgte harpiksen på grunn av sin lave viskositet som gir bedre infiltrasjon gjennom celleveggene22,23. Videre kan ulike buffere brukes til fiksering (f.eks. kaktocylatbuffer, HEPES eller fosfatbuffer samt en annen konsentrasjon av glutaraldehyd (2-3%) og paraformaldehyd (2-4%)23,24,25,26,27,28). Bruk av tannsyre for å forbedre farging av prøver rik på ekstracellulær matrise29 eller ruthenium rød for å forbedre farging av plantemateriale23,30 er en annen endring som kan brukes. Når det gjelder dehydrering, anbefales vanligvis etanol og/eller aceton for epoksyharpiks24,28.
Den samme protokollen som beskrevet her, eller veldig lik, har blitt brukt før av teamet vårt og andre grupper4,5,24. Eksempler som behandles i henhold til denne metoden, er kompatible med TEM imaging31. Sammenlignet med den opprinnelige NCMIR-protokollen som ble publisert i 201010, introduserer vi noen mindre endringer som utskifting av giftig kaktocylatbuffer under fiksering og osmium postfikseringstrinn med en fosfatbuffer. Som nevnt ovenfor kan forskjellige buffere brukes under EM-forberedelse, og ifølge vår erfaring gir giftfri PB tilfredsstillende resultater.
En annen endring er en erstatning av aceton med mindre brukerskadelig etanol i det siste trinnet av dehydrering. Etanol ble også brukt som harpiksløsningsmiddel under innbyggingsprosessen. Videre ble DMP-30 brukt i stedet for epoksyharpikskomponent C, som publisert før32,33.
På grunn av behovet for å bruke de samme hjernene til immunfluorescence studier ble to-trinns fiksering tilnærming introdusert. Først, dyrene ble parfymert med en buffer som inneholdt en lavere konsentrasjon av glutaraldehyd (mild primær fiksering ved perfusjon med en buffer som inneholdt 2% PFA og 0,5% GA) og etter det var bare halvparten av hjernen dedikert til EM ytterligere postfixed med en høyere konsentrasjon av glutaraldehyd (2% PFA og 2,5% GA), mens den andre halvdelen av hjernen ble postfixed med en standard fixative for immunostaining (4% PFA). Bruken av EM fixative med lavere konsentrasjon av PFA ble publisert før av andre og vår gruppe4,5,31,34. Totrinnsfikseringen som presenteres her, er like tilstrekkelig som en tradisjonell etttrinnsmetode (Figur 3 og Figur 4). Men siden vi foreslår at hver halvdel av hjernen kan behandles separat, gjør vår protokoll det mulig å redusere antall dyr som brukes. Mild primær fiksering opprettholder vellykket vevsantigenicitet, noe som muliggjør ytterligere immunfluorescens som vist for antistoffet mot PSD-95 protein. Det må imidlertid understrekes at nytten av vår tilnærming må valideres for andre antistoffer.
Den presenterte metoden resulterer i høykontrastbilder av hjernevevet med tydelig synlige membraner. Det er imidlertid verdt å nevne at det er de samme trinnene som er utsatt for feil, som i tilfelle av noen annen metode for SBEM-prøveforberedelse, og spesiell oppmerksomhet må gis til dem. Kvaliteten på vevsmorfologien i seg selv avhenger for det meste av perfusjonen. Det er et kritisk skritt, da bare vellykkede faste prøver gir tilfredsstillende resultater. Dessverre påvirkes fikseringsperfusjon av individuell variasjon blant dyr, og dermed kan kvaliteten på bildene variere fra ett dyr til et annet. Videre må man være forsiktig så man ikke tørker prøven under dehydrering; Ellers vil prøven ikke bli infiltrert riktig med harpiksen. Et annet viktig aspekt er prøvemontering til pinnen. Epoksyharpiksene er ikke-konduktive, og derfor samler de elektroner under avbildning av hva som forårsaker lading av artefakter. For å redusere ladeeffekten i SBEM, er det nødvendig å forsiktig fjerne overflødig harpiks på hver side av prøven og nøyaktig jorde prøven med sølvmaling og sputterbelegg.
Den nåværende protokollen fokuserer på hjernevev; Det kan imidlertid tilpasses for cellemonolayer, organotypiske skiver eller andre vev. I likhet med den opprinnelige NCMIR-protokollen resulterer den som presenteres her i svært kontrasterte prøver som ikke bare passer for SBEM, men også for FIB-SEM- og ATUM-SEM-eksperimenter.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingenting å avsløre.
Acknowledgments
SBEM-avbildning, lysmikroskopiavbildning og elektronmikroskopiprøvepreparering ble utført ved bruk av utstyret til Laboratory of Imaging Tissue and Function som fungerer som et bildekjerneanlegg ved Nencki Institute of Experimental Biology.
For fremstilling av figur 1 ble bildet av en mus (Souris_02) og et hetteglass fra https://smart.servier.com/ brukt.
Dette arbeidet ble støttet av National Science Centre (Polen) Grant Opus (UMO-2018/31/B/NZ4/01603) tildelt KR.
Materials
| Name | Company | Catalog Number | Comments |
| Anesthetic: | |||
| Ketamine/xylazine mixture (Ketamina/Sedazin) | Biowet Pulawy, Pulawy, Poland | ||
| Sodium pentobarbital (Morbital) | Biowet Pulawy, Pulawy, Poland | ||
| Fixatives: | |||
| Glutaraldehyde (GA) | Sigma-Aldrich,St. Louis, MI, USA | G5882 | Grade I, 25% in H2O, specially purified for use as an electron microscopy fixative |
| Hydrochloric acid (HCl) | POCH, Gliwice, Poland | 575283115 | pure p.a. |
| Paraformaldehyde (PFA) | Sigma-Aldrich,St. Louis, MI, USA | 441244 | prilled, 95% |
| Phosphate buffered saline (PBS), pH 7.4 | Sigma-Aldrich,St. Louis, MI, USA | P4417-50TAB | tablets |
| Sodium hydroxide (NaOH) | Sigma-Aldrich,St. Louis, MI, USA | S5881 | reagent grade, 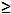 98%, pellets (anhydrous) 98%, pellets (anhydrous) |
| Sodium phosphate dibasic (Na2HPO4) | Sigma-Aldrich,St. Louis, MI, USA | S3264 | |
| Sodium phosphate monobasic (NaH2PO4) | Sigma-Aldrich,St. Louis, MI, USA | S3139 | |
| Perfusion: | |||
| Large blunt/blunt curved scissors (~14.5 cm) | Fine Science Tools, Foster City, CA, USA | 14519-14 | |
| Micro-spatula (double 2" flat ends, one rounded, one tapered to 1/8") | Fine Science Tools, Foster City, CA, USA | 10091-12 | |
| Needle tip, 15 GA, blunt (perfusion needle) | KD Medical GmbH Hospital Products, Berlin, Germany | KD-FINE 900413 | 1.80 x 40 mm |
| Pair of fine (Graefe) tweezers | Fine Science Tools, Foster City, CA, USA | 11050-10 | |
| Perfusion pump | Lead Fluid | BQ80S | |
| Plastic vials | Profilab, Warsaw, Poland | 534.02 | plastic vials with blue cap for tissue storage, 20 ml, 31 x 48 mm |
| Straight iris scissors (~9 cm) | Fine Science Tools, Foster City, CA, USA | 14058-11 | |
| Brain slices preparation for EM: | |||
| 12-well plate | NEST, Rahway, NJ, USA | 712001 | |
| Cyanoacrylic glue | Fenedur, Montevideo, Uruguay | ||
| Glass vials | Electron Microscopy Sciences, Hatfield, PA, USA | 72632 | 20 ml Scintillation Vial, a pack of 100 |
| Pasteur pipette | VWR, Radnor, PA, USA | 612-4545 | LDPE, disposable, 7.5 ml |
| Razor blade | Wilkinson Sword, London, UK | Classic double edge safety razor blades | |
| Scalpel blade | Swann-Morton, Sheffield, UK | No. 20 | |
| Vibratome | Leica Microsystems, Vienna, Austria | Leica VT1000 S | |
| Brain slices preparation for IF: | |||
| 96-well plate | NEST, Rahway, NJ, USA | 701101 | |
| Criostat | Leica Microsystems, Vienna, Austria | Leica CM 1950 | |
| Ethylene glycol | Bioshop, Burlington, Canada | ETH001 | |
| Low-profile disposable blade 819 | Leica Biosystems Inc., USA | 14035838925 | |
| Scalpel blade | Swann-Morton, Sheffield, UK | No. 20 | |
| Sodium azide (NaN3) | POCH, Gliwice, Poland | 792770426 | |
| Sucrose | POCH, Gliwice, Poland | 772090110 | |
| Tissue freezing medium for cryosectioning, OCT-Compound | Leica Biosystems, Switzerland | 14020108926 | |
| Immunostaining: | |||
| 24-well plate | NEST, Rahway, NJ, USA | 702001 | |
| Anti-Post Synaptic Density Protein 95 Antibody | Merck-Millipore, Burlington, MA, USA | MAB1598 | |
| Confocal microscope | Zeiss, Göttingen, Germany | Zeiss Spinning Disc microscope (63 × oil objective, NA 1.4, pixel size 0.13 µm × 0.13 µm) | |
| Cover slide | Menzel Glaser, Braunschweig, Germany | B-1231 | 24 x 60 mm |
| Donkey anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor 555 | Invitrogen, Carlsbad, CA, USA | A31570 | |
| Fluoromount-G Mounting Medium, with DAPI | Invitrogen, Carlsbad, CA, USA | 00-4959-52 | |
| Microscope slide | Thermo Scientific, Waltham, MA, USA | AGAA00008 | SuperFrost |
| Normal donkey serum (NDS) | Jackson ImmunoResearch Laboratories, West Grove, PA, USA | 017-000-121 | |
| Shaker | JWElectronic, Warsaw, Poland | KL-942 | |
| TritonT X-100 Reagent Grade | Bioshop, Burlington, Canada | TRX506 | |
| Electron microsocpy sample preparation | |||
| Potassium hexacyanoferrate(II) trihydrate | POCH, Gliwice, Poland | 746980113 | |
| Aclar 33C Film | Electron Microscopy Sciences, Hatfield, PA, USA | 50425 | Fluoropolymer Film embedding sheet |
| DMP-30, 2,4,6-Tris(dimethylaminomethyl)phenol | Sigma-Aldrich,St. Louis, MI, USA | T58203 | Epoxy embedding medium accelerator |
| Durcupan ACM single component A, M | Sigma-Aldrich,St. Louis, MI, USA | 44611 | Durcupan ACM single component A, M epoxy resin |
| Durcupan ACM single component B | Sigma-Aldrich,St. Louis, MI, USA | 44612 | Durcupan ACM single component B, hardener 964 |
| Durcupan ACM single component D | Sigma-Aldrich,St. Louis, MI, USA | 44614 | Durcupan ACM single component D , plasticizer |
| Ethyl alcohol absolute | POCH, Gliwice, Poland | 64-17-5 | Ethyl alcohol absolute 99.8 % pure P.A.-BASIC |
| Genlab laboratory oven | Wolflabs, York, UK | Mino/18/SS | Oven Genlab MINO/18/SS 18l volume, no fan circulation, no digital display, standard temperature gradient, standard recovery rate, no timer, 250°C maximum temperature, 240V electrical supply |
| L-Aspartic acid | Sigma-Aldrich,St. Louis, MI, USA | A-9256 | reagent grade, 98% (HPLC) |
| Lead (II) nitrate | Sigma-Aldrich,St. Louis, MI, USA | 467790 | 99.95% trace metals basis |
| Osmium tetroxide | Sigma-Aldrich,St. Louis, MI, USA | 75632 | for electron microscopy, 4% in H2O |
| pH meter | Elmetron, Zabrze, Poland | CP-5-5 | |
| Rotator | BioSan, Józefów, Poland | Multi Bio RS-24 | rotator Multi Bio RS-24 |
| Sodium hydroxide (NaOH) | Sigma-Aldrich,St. Louis, MI, USA | S5881 | reagent grade, 98%, pellets (anhydrous) |
| Sunflower mini shaker | Grant bio, Shepreth,UK | PD-3D | |
| Syringe filter | Millipore, Burlington, MA, USA | SLGP033NB | 0,22 µm pore size |
| Thiocarbohydrazide | Sigma-Aldrich,St. Louis, MI, USA | 88535 | purum p.a., for electron microscopy, 99.0% (N) |
| Uranyl acetate | Serva, Heidelberg, Germany | 77870 | Uranyl acetate·2H2O, research grade |
| Water bath | WSL, Swietochlowice, Poland | LWT | |
| Specimen mounting for SBEM | |||
| 96-well culture plate | VWR, Radnor, PA, USA | 734-2782 | 96-well plates, round bottom, non treated |
| AM Gatan 3View stub handling tweezers | Micro to Nano, Haarlem, Netherlands Netherlands |
50-001521 | |
| Binocular | OPTA-TECH, Warsaw, Poland | X2000 | |
| Conductive glue | Chemtronics, Georgia, USA | CW2400 | conductive eopxy |
| Gatan 3View sample pin stubs | Micro to Nano, Haarlem, Netherlands Netherlands |
10-006003 | |
| Parafilm | Sigma-Aldrich,St. Louis, MI, USA | P7793 | roll size 20 in. × 50 ft |
| Pelco conductive silver paint | Ted Pella, Redding, CA, USA | 16062-15 | PELCO® Conductive Silver Paint, 15g |
| Razor blades double edge | Electron Microscopy Sciences, Hatfield, PA, USA | 72000 | Stainless Steel "PTFE" coated. PERSONNA brand .004" thick, wrapped individually, 250 blades in a box. |
| Scanning Electron Microscope | Zeiss, Oberkochen, Germany | Sigma VP with Gatan 3View2 chamber, acceleration voltage 2.5 kV, variable pressure 5 Pa, aperture 20 µm, dwell time 6 µs, slice thickness 60 nm, magnification 15 000 x, image resolution 2048 x 2048 pixels, pixel size 7.3 nm | |
| trim 90° diamond knife | Diatome Ltd., Nidau, Switzerland | DTB90 | |
| Ultramicrotome | Leica Microsystems, Vienna, Austria | Leica ultracutR | |
| Software | webpage | tutorials | |
| FijiJ | https://fiji.sc/ | ||
| Microscopy Image Browser | http://mib.helsinki.fi/ | http://mib.helsinki.fi/tutorials.html | |
| Reconstruct | https://synapseweb.clm.utexas.edu/software-0 | https://synapseweb.clm.utexas.edu/software-0) | |
| Animals | |||
| Mice | Adult 3-month old and 20±1 month old female Thy1-GFP(M) mice (Thy1-GFP +/-) (Feng et al.,2000) which express GFP in a sparsely distributed population of glutamatergic neurons. Animals were bred as heterozygotes with the C57BL/6J background in the Animal House of the Nencki Institute of Experimental Biology. |
References
- Bosch, M., Hayashi, Y. Structural plasticity dendritic spines. Current Opinion in Neurobiology. 22 (3), 383-388 (2012).
- Borczyk, M., Radwanska, K., Giese, K. P. The importance of ultrastructural analysis of memory. Brain Research Bulletin. 173, 28-36 (2021).
- Wanner, A. A., Kirschmann, M. A., Genoud, C. Challenges of microtome-based serial block-face scanning electron microscopy in neuroscience: challenges of SBEM in neuroscience. Journal of Microscopy. 259 (2), 137-142 (2015).
- Śliwińska, M. A., et al. Long-term Memory Upscales Volume of Postsynaptic Densities in the Process that Requires Autophosphorylation of αCaMKII. Cerebral Cortex. 30 (4), 2573-2585 (2020).
- Borczyk, M., Śliwińska, M. A., Caly, A., Bernas, T., Radwanska, K. Neuronal plasticity affects correlation between the size of dendritic spine and its postsynaptic density. Scientific Reports. 9 (1), 1693 (2019).
- Leighton, S. B. SEM images of block faces, cut by a miniature microtome within the SEM - a technical note. Scanning Electron Microscopy. , Pt 2 73-76 (1981).
- Denk, W., Horstmann, H. Serial Block-Face Scanning Electron Microscopy to Reconstruct Three-Dimensional Tissue Nanostructure. PLoS Biology. 2 (11), 329 (2004).
- Smith, D., Starborg, T. Serial block face scanning electron microscopy in cell biology: Applications and technology. Tissue and Cell. 57, 111-122 (2019).
- Titze, B., Genoud, C. Volume scanning electron microscopy for imaging biological ultrastructure: Volume scanning electron microscopy. Biology of the Cell. 108 (11), 307-323 (2016).
- Deerinck, T. J., Bushong, E. A., Thor, A., Ellisman, M. H. NCMIR methods for 3D EM: A new protocol for preparation of biological specimens for serial block face scanning electron microscopy. , 6-8 (2010).
- Willingham, M. C., Rutherford, A. V. The use of osmium-thiocarbohydrazide-osmium (OTO) and ferrocyanide-reduced osmium methods to enhance membrane contrast and preservation in cultured cells. Journal of Histochemistry & Cytochemistry. 32 (4), 455-460 (1984).
- Feng, G., et al. Imaging Neuronal Subsets in Transgenic Mice Expressing Multiple Spectral Variants of GFP. Neuron. 28 (1), 41-51 (2000).
- Gage, G. J., Kipke, D. R., Shain, W. Whole Animal Perfusion Fixation for Rodents. Journal of Visualized Experiments. (65), e3564 (2012).
- Paxinos, G., Franklin, K. B. J. The mouse brain in stereotaxic coordinates. , Acad. Press. San Diego, Calif. (2004).
- Walton, J. Lead aspartate, an en bloc contrast stain particularly useful for ultrastructural enzymology. Journal of Histochemistry & Cytochemistry. 27 (10), 1337-1342 (1979).
- Mercer, E. H. a scheme for section staining in electron microscopy. Journal of the Royal Microscopical Society. 81 (3-4), 179-186 (1963).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Belevich, I., Joensuu, M., Kumar, D., Vihinen, H., Jokitalo, E. Microscopy Image Browser: A Platform for Segmentation and Analysis of Multidimensional Datasets. PLOS Biology. 14 (1), 1002340 (2016).
- Fiala, J. C. Reconstruct: a free editor for serial section microscopy. Journal of Microscopy. 218 (1), 52-61 (2005).
- Roels, J., et al. An interactive ImageJ plugin for semi-automated image denoising in electron microscopy. Nature Communications. 11 (1), 771 (2020).
- Radwanska, K., et al. Mechanism for long-term memory formation when synaptic strengthening is impaired. Proceedings of the National Academy of Sciences. 108 (45), 18471-18475 (2011).
- Kittelmann, M., Hawes, C., Hughes, L. Serial block face scanning electron microscopy and the reconstruction of plant cell membrane systems: SBFSEM Methods for Plant Cells. Journal of Microscopy. 263 (2), 200-211 (2016).
- Fendrych, M., et al. Programmed Cell Death Controlled by ANAC033/SOMBRERO Determines Root Cap Organ Size in Arabidopsis. Current Biology. 24 (9), 931-940 (2014).
- Russell, M. R. G., et al. 3D correlative light and electron microscopy of cultured cells using serial blockface scanning electron microscopy. Journal of Cell Science. 130 (1), 278-291 (2017).
- Płachno, B. J., Świątek, P., Jobson, R. W., Małota, K., Brutkowski, W. Serial block face SEM visualization of unusual plant nuclear tubular extensions in a carnivorous plant (Utricularia, Lentibulariaceae). Annals of Botany. 120 (5), 673-680 (2017).
- Genoud, C., Titze, B., Graff-Meyer, A., Friedrich, R. W. Fast Homogeneous En Bloc Staining of Large Tissue Samples for Volume Electron Microscopy. Frontiers in Neuroanatomy. 12, (2018).
- Puhka, M., Joensuu, M., Vihinen, H., Belevich, I., Jokitalo, E. Progressive sheet-to-tubule transformation is a general mechanism for endoplasmic reticulum partitioning in dividing mammalian cells. Molecular Biology of the Cell. 23 (13), 2424-2432 (2012).
- Gluenz, E., Wheeler, R. J., Hughes, L., Vaughan, S. Scanning and three-dimensional electron microscopy methods for the study of Trypanosoma brucei and Leishmania mexicana flagella. Methods in Cell Biology. 127, 509-542 (2015).
- Starborg, T., et al. Using transmission electron microscopy and 3View to determine collagen fibril size and three-dimensional organization. Nature Protocols. 8 (7), 1433-1448 (2013).
- Hughes, L., Borrett, S., Towers, K., Starborg, T., Vaughan, S. Patterns of organelle ontogeny through a cell cycle revealed by whole-cell reconstructions using 3D electron microscopy. Journal of Cell Science. 130 (3), 637-647 (2017).
- Bojko, A., et al. Improved Autophagic Flux in Escapers from Doxorubicin-Induced Senescence/Polyploidy of Breast Cancer Cells. International Journal of Molecular Sciences. 21 (17), 6084 (2020).
- Knott, G. W., Holtmaat, A., Trachtenberg, J. T., Svoboda, K., Welker, E. A protocol for preparing GFP-labeled neurons previously imaged in vivo and in slice preparations for light and electron microscopic analysis. Nature Protocols. 4 (8), 1145-1156 (2009).
- Glauert, A. M., Lewis, P. R. Biological specimen preparation for transmission electron microscopy. , Princeton University Press. Princeton. (2014).
- Genoud, C. Altered Synapse Formation in the Adult Somatosensory Cortex of Brain-Derived Neurotrophic Factor Heterozygote Mice. Journal of Neuroscience. 24 (10), 2394-2400 (2004).
