

Summary
Deze video artikel demonstreert de oprichting van organotypische netvlies wholemount culturen en een Cytospin procedure voor de analyse van exogeen geïnduceerde effecten. Organotypische netvlies wholemount culturen na te bootsen van de
Abstract
Gerichte ablaties van genen en de analyse van dierlijke modellen is de klassieke strategie voor de inschrijving voor specifieke netvlies functie van een gen. Echter, transgene, netvlies-specifieke of conditionele knockout muismodellen vertonen vaak vroeg letaliteit of lijdt aan ernstige misvormingen, het voorkomen van een analyse verder dan embryonale of vroege postnatale fase.
Primaire celkweek is een alternatief voor de effecten van exogeen recombinant toegepaste factoren, overexpressie van genen of siRNA-gemedieerde gen knockdown in een gecontroleerde omgeving te onderzoeken. Gescheiden celcultuur heeft het voordeel dat de endogene signalen bereiken van de doelcellen worden verminderd, waardoor het vergemakkelijken van de identificatie van exogeen veroorzaakt effecten na farmacologische manipulatie. Er zijn echter belangrijke cel-cel interacties in eerste instantie vernietigd door enzymatische digestie of mechanische dissociatie, zelfs als re-geaggregeerde retinospheroid culturen een worden gebruikt.
Daarentegen, organotypische netvlies wholemount culturen zorgen voor een systeem te sluiten bij de fysiologische in vivo situatie met neuronale interacties en verbindingen nog steeds bewaard gebleven 2-5.
In deze video artikel bieden we een stap voor stap demonstratie van (1) de oprichting van in vivo-achtige organotypische netvlies wholemount culturen waaronder dissectie eigenaardigheden van embryonale, postnatale en volwassen muizen ogen en (2) een dissociatie en Cytospin procedure voor de analyse van neuronale apoptose en retinale celproliferatie in organotypische wholemounts, bijvoorbeeld na de cultuur in de aanwezigheid van exogeen recombinant toegepaste factoren.
Protocol
Alle apparatuur en reagentia moeten worden aangeschaft steriel of moet worden warmte of stoom gesteriliseerd of gesteriliseerd met 70% EtOH.
De auteurs stellen dat experimenten op dieren werden uitgevoerd in overeenstemming met de Europese Gemeenschappen richtlijn (86/609/EEG), volgens de richtlijnen van de NIH ten aanzien van de zorg en het gebruik van dieren voor experimentele procedures en de regels uiteengezet door de Institutional Animal set zorg en gebruik Comite (IACUC) aan de Universiteit van Duisburg-Essen (Duitsland).
Deel 1: Enucleatie van muriene ogen van verschillende ontwikkelingsstadia
Enucleatie van embryonale ogen
- Tijd zwanger paringen worden opgezet en de ochtend van de dag waarop een vaginale plug wordt ontdekt bij vrouwen paring is aangewezen zwangerschap dag 0.
- De zwangere vrouw wordt opgeofferd door cervicale dislocatie wanneer de ontwikkeling van de embryo's heeft bereikt op de gewenste fase (hier: embryonale dag (E) 15) en gefixeerd op een wax bord 6.
- De buikwand wordt bevochtigd met 70% EtOH, snijd over de middellijn en de huid flappen worden lateraal bevestigd door de pinnen 6.
- De uterusses worden verwijderd uit de buik, vrijstaande en overgebracht naar een beker met koude PBS 6.
- De embryo's worden gescheiden, overgebracht naar een petrischaal en baarmoeder wand-en embryonale membranen worden zorgvuldig verwijderd door het gebruik van de tang 6.
- Embryo's worden gedood door onthoofding.
- Ogen zijn enucleated worden het gebruik van fijne, gebogen tang, "peeling" de ogen van de oogkas.
Enucleatie van de postnatale en volwassen ogen
- Jonge pups worden gedood door onthoofding, oudere pups en volwassen door cervicale dislocatie.
- Tot postnatale fase P15, het tijdstip wanneer de muizen hun ogen open, oog gleuven moeten mechanisch worden geopend door het gebruik van de tang en vergroot door twee dwarse sneden van de oogleden met veer schaar.
- Ogen zijn enucleated door de hulp van gebogen pincet, druk uitoefenen op de baan.
Opmerking: Zoals op postnatale dag 2, de orbitale botten zijn nog kraakbeen, is het belangrijk niet te veel druk uit te oefenen terwijl het proberen om de ogen te verwijderen.
Ter vergelijking, in de volwassen muizen, de orbitale botten stevig. Dus, om ophelderen de ogen is het voldoende om de druk op de baan toe te passen zonder uitbreiding van het oog spleten op voorhand.
Deel 2: Dissectie van embryonale en volwassen muizen postnatale netvliezen
Dissectie van netvliezen
- De ogen zijn geplaatst in een kleine petrischaal met steriele PBS en de omliggende ogen lagen zijn verwijderd onder een dissectie microscoop.
- Om de buitenste lagen oog te verwijderen, in postnatale stadia alsook tegen volwassen ogen van de oogzenuw te worden afgesneden door de hulp van de lente schaar of afgeknepen door de tang zo dicht mogelijk bij de basis mogelijk te maken.
- Draai het oog, zodat de achterkant met het gat waar oorspronkelijk de oogzenuw verbleven naar u toe. Geef de subretinale ruimte tussen netvlies en pigment epitheel met de toppen van twee zeer fijne pincet van de plaats waar de oogzenuw het oog doorgedrongen lagen.
Let op: Meestal is het pigment epitheel gemakkelijk kunnen worden geïdentificeerd door zijn donkere kleur. In sommige muis mutante stammen - vooral in albino dieren - dit pigment laag kan echter niet worden gekleurd en dus niet onmiddellijk kunnen worden op te sporen. - Verwijder het pigment epitheel met de bijgevoegde vaatvlies membraan en sclera door zorgvuldig te scheuren aan beide zijden met beide tang.
- Verwijder de lagen tot op het niveau van het hoornvlies, draai de oogschelp om de lens te verwijderen en samen het hoornvlies met pigment epitheel, choroidea en membraan sclera, terwijl het resterende netvlies beker door de andere tang.
- Samen Pak het glasvocht met de kleine lens en terwijl scheuren met een pincet, de retinale wholemount zijn plaats te houden met de tweede tang.
Opmerking: Als het ontleden van embryonale ogen, zorg er dan volledig de driehoekige, tent-achtige capillaire plexus samen te verwijderen onder het glasachtig lichaam met het glasvocht.
In de volwassen oog, het glasachtig lichaam dient te worden begrepen aan de zijkanten en de zorg moet worden genomen niet te doorboren het glasvocht met de toppen van de tang als de inhoud ervan is stroperig en kleeft aan de tang, belemmeren de verwijdering ervan. - Voor organotypische wholemount de cultuur van de retina bekers worden verzameld in een 96-well plaat met 200 ul Dulbecco's gemodificeerd Eagle's medium (zie hieronder).
Let op: Tussen de dissectie van de individuele netvlies, houdt de 96-well plaat collectie met daarin het kweekmedium in de incubator als de pH van het kweekmedium wordt veroorzaakt door CO 2 via de carbonaat-systeem.
Deel 3: Murine organotypische netvlies wholemountcultuur
- Bereid 500 ml cultuur door middelgrote weging van 7.8g Dulbecco's gemodificeerd Eagle's / voedingsstoffen mengsel F-12 HAM (DMEM) en 0,6 g NaHCO 3 en op te lossen, zowel in MiIliQ water. Stel de pH op 7,15. Voeg 50mg apo-transferin, 50μl putrescin (stock: 60mg/ml), 50μl natriumseleniet (stock: 52μg/ml), progesteron (stock: 60μg/ml), en 2,5 ml gentamicine (200 mm) onder de motorkap. Meng en filtreer door een fles top filter. Onmiddellijk voorafgaand aan het gebruik toe te voegen 10μl glutamine (200 mm) per ml kweekmedium.
Opmerking: Dit serum-en insuline-vrij medium kan worden opgeslagen bij 4 ° C gedurende maximaal twee weken en gebruikt voor apoptose inductie experimenten als er geen insuline gaat de effecten. Als incubatie van wholemounts langer dan 24 uur is gewenst en celdood tarieven zullen niet worden geëvalueerd, insuline serum (bijv. foetaal kalf serum; FCS) moet of supplementen worden toegevoegd aan de overlevingskansen te verbeteren. - Voordat u begint met de cultuur, de retina wholemounts pre-geïncubeerd gedurende 15 min bij 37 ° C met 200 ul warm, pH-gebalanceerde DMEM dat 0.5mg/ml hyaluronidase vooraf verteren de hyaluronidase met binnenste en buitenste membraan van het beperken van M ller glia cellen, het vergemakkelijken van de penetratie van exogeen toegepaste stoffen.
- Netvliezen worden overgebracht naar een 24-wells plaat met zo weinig hyaluronidase mogelijk en gekweekt als organotypische wholemounts in 2 ml chemisch welbepaald Dulbecco's gemodificeerd Eagle medium.
Opmerking: Voor het overbrengen van het netvlies, gebruik dan een 1 ml pipet en de pipetpunt een paar millimeter gesneden om de opening te verbreden. Voor de embryonale ogen, een 200 ul pipetpunt is voldoende. De snijkanten moeten worden gladgestreken door het inbrengen en het verdraaien van een seconde pipetpunt.
Voor 24-48 uur op korte termijn culturen, kunnen alle stappen worden uitgevoerd op de bank, maar als verontreiniging van de culturen blijkt te zijn een probleem, moet men het werk onder de motorkap. - Culturen worden onderhouden voor 24 - 48 uur bij 37 ° C in een 5% CO 2 atmosfeer en bijvoorbeeld onderworpen aan een farmacologische behandeling met recombinant factoren.
Deel 4: Dissociatie van gekweekte netvlies wholemounts
- Nadat de gewenste cultuur tijd, zijn netvlies verzameld in 2 ml Eppendorf buizen met ronde bodem met 850 ul PBS en 50 ui bovine serum albumine (BSA; 30 mg / ml).
- Plaats Eppendorf buizen met netvliezen in een verwarmings-blok, opgewarmd bij 37 ° C.
- Voeg 25 ul collagenase (200 U / ml) en 25 ul hyaluronidase (20mg/ml) om elke eppendorfbuisje en start dissociëren het netvlies in enkele celsuspensie met 3 gaat door een gesiliconiseerde Pasteur pipet.
- Voeg 10 ul trypsine (1mg/ml), wacht 3-5 minuten, en dan langzaam pipet 3-5 keer op en neer met gesiliconiseerde Pasteur pipet mechanisch dissociëren het weefsel.
- Voeg 10 ul DNase I (5mg/ml), weer wachten op 3-5 minuten, langzaam dan 3-5 keer pipet op en neer met gesiliconiseerde Pasteur pipet.
Opmerking: De incubatietijd voor enzymatische dissociatie varieert en is afhankelijk van de grootte van de ogen en de ontwikkelingsfase, respectievelijk. Controleer het stadium van de enzymatische vertering van het weefsel door voorzichtig en neer te pipetteren. - Als de celsuspensie niet homogeen is inmiddels maar nog steeds bevat belangrijke cel aggregaten, voeg extra 10 pl trypsine en 10 pi DNase I.
- Wanneer de celsuspensie homogeen is, vertering van het weefsel wordt gestopt door toevoeging van 10 ul EDTA (0,5 M), Eppendorf-buisjes worden verwijderd uit de kachel en de celsuspensies zijn vastgesteld voor 1 uur door de toevoeging van 1 ml verse, ijskoude 8% paraformaldehyde (PFA) bij kamertemperatuur op een rotatie-shaker.
Deel 5: Het wassen van gedissocieerde celsuspensies
- De celsuspensie wordt gecentrifugeerd gedurende 5 minuten bij 4 ° C en bij 0,2 RCF in een koel-centrifuge.
- Het supernatant wordt verwijderd en de pellet wordt opnieuw gesuspendeerd in 1 ml PBS met 3mg/ml BSA.
- Na het herhalen van deze wasstappen twee keer, is het eindelijk pellet geresuspendeerd in 500 pi PBS met 3mg/ml BSA, 5mM EDTA en 0,1% natriumazide.
Opmerking: De toevoeging van natrium-zuur maken opslag van de celsuspensie voor meerdere dagen bij 4 ° C. Echter, als een immunocytochemische kleuring zal volgen, doen natrium-zuur niet toe te voegen aan de resuspensie buffer, omdat dit leidt tot verlies van kwaliteit vlekken.
Deel 6: Cytospin van celsuspensies voor de kwantitatieve analyse van apoptose en proliferatie
- Een matte-end microscoop glijbaan, een Cytospin filter met een of twee gaten en een Cytospin trechter worden ingevoegd in een Cytospin glijbaan clip. De dia clip is gesloten en geplaatst in het Cytospin rotor.
- De gedissocieerde celsuspensie wordt gehomogeniseerd door zachtjes en neer te pipetteren.
Opmerking: Afhankelijk van het ontwikkelingsstadium van het netvlies, kan de celsuspensie van de dissociatie procedure moetenworden verdund met PBS tot een aftelbaar aantal cellen te verkrijgen. - Een aliquot (100 ul) van de celsuspensie wordt toegepast op een Cytospin trechter.
Opmerking: Als pipetteren de celsuspensie in de trechter, de punt van de pipet moet helemaal te bereiken tot op de bodem van de trechter. Het is belangrijk om niet door te drukken de tweede druk punt van de pipet, omdat dit zorgt voor luchtbellen, die zichtbaar zijn in de cel plek na de Cytospin en belemmert de cellen telt. - De celsuspensie is gespot op een glijbaan bij 700 omwentelingen per minuut gedurende 7 minuten.
- Voor de bepaling van het effect van exogeen toegepaste factoren op apoptose niveaus cellen kunnen worden gekleurd met 4 ',6-diamidino-2-phenylindole (DAPI, 2μg/ml), gemonteerd met fluorescerende montage medium. Veranderingen in de cel apoptose kan worden bepaald door het tellen van minimaal 1000 cellen (bestaande uit ten minste 10 pycnotic kernen) in de cel Cytospin plekjes en de cel sterftecijfer wordt berekend als percentage van de totale cellen telt 3,4.
Opmerking: Als alternatief, de verdeling van apoptotische kernen kan worden geëvalueerd in flatmounts 3 of cryostaat sections4 van gekweekte netvlies wholemounts door TdT-gemedieerde dUTP nick end labeling (TUNEL). - Voor de opsporing van celproliferatie, kan BrdU (5 uM) toegevoegd 6h voor het einde van de cultuur en de BrdU incorporatie gevisualiseerd in cytospins van celhomogenaten door immunocytochemische kleuring met behulp van een anti-BrdU antilichaam (bijv. Developmental Studies Hybridoma Bank, Iowa, USA).
- Het effect van de behandeling op verschillende retina celtypes gebruik kunnen worden gevisualiseerd in cytospins door neuron specifieke antilichamen, zoals Brn3a (ganglion cel marker) of opsin (afdrukband marker) en tegenkleuring met DAPI.
Deel 7: representatieve resultaten

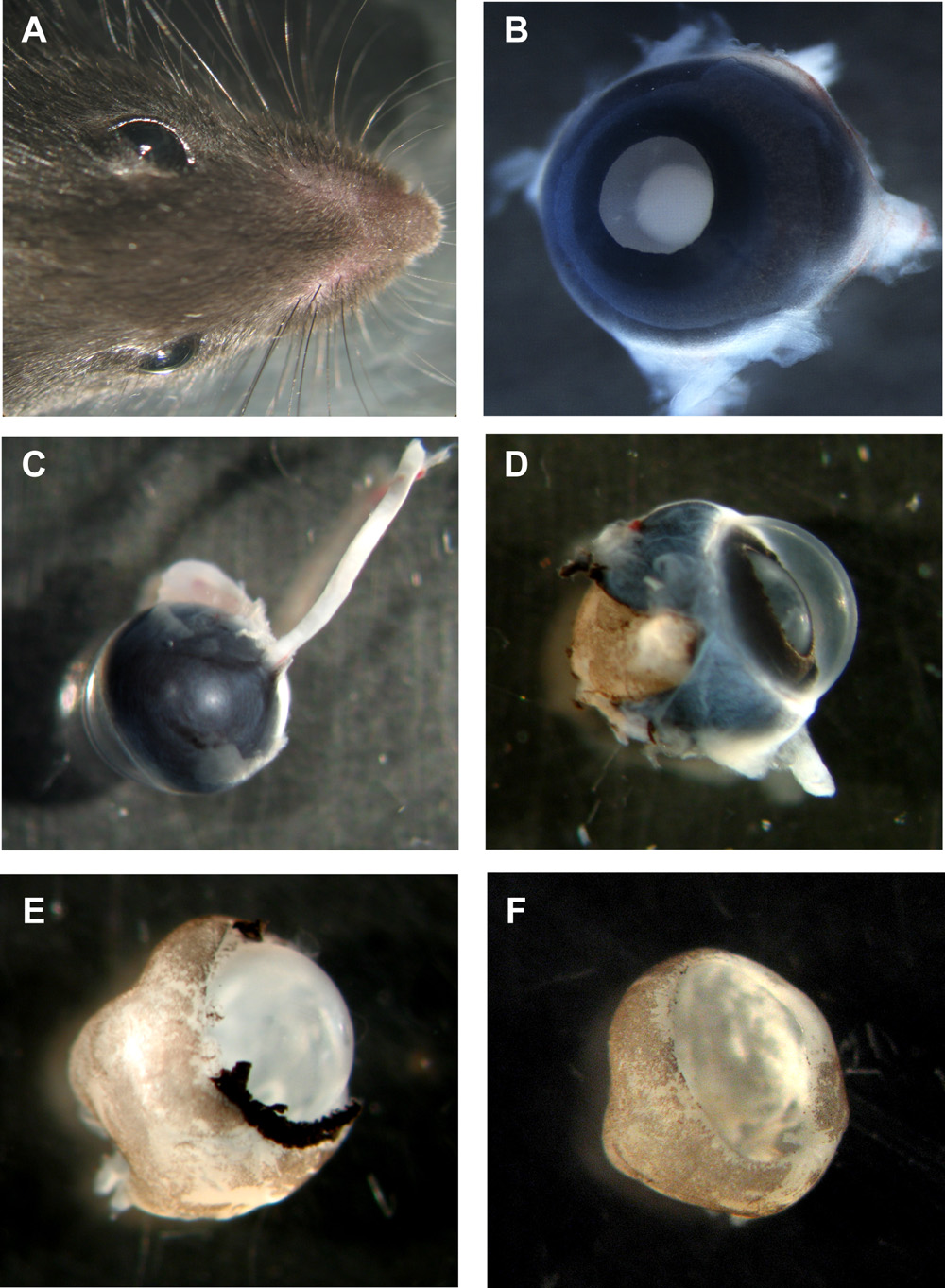
Figuur 1: Stappen in de voorbereiding van muriene organotypische netvlies wholemounts
Een hoofd van de muis met beide ogen. B Murine oog met lens naar boven, alle lagen nog op zijn plaats. C Murine oog van de achterzijde met de oogzenuw er nog aan. D Murine oog met sclera en pigment epitheel gedeeltelijk verwijderd. E Murine netvlies met hoornvlies, sclera en pigment epitheel volledig verwijderd, maar lens en het glasvocht nog op zijn plaats. F Murine netvlies wholemount beker met lens en het glasvocht verwijderd. Gelieve Klik hier voor een grotere versie van figuur 1 te zien.

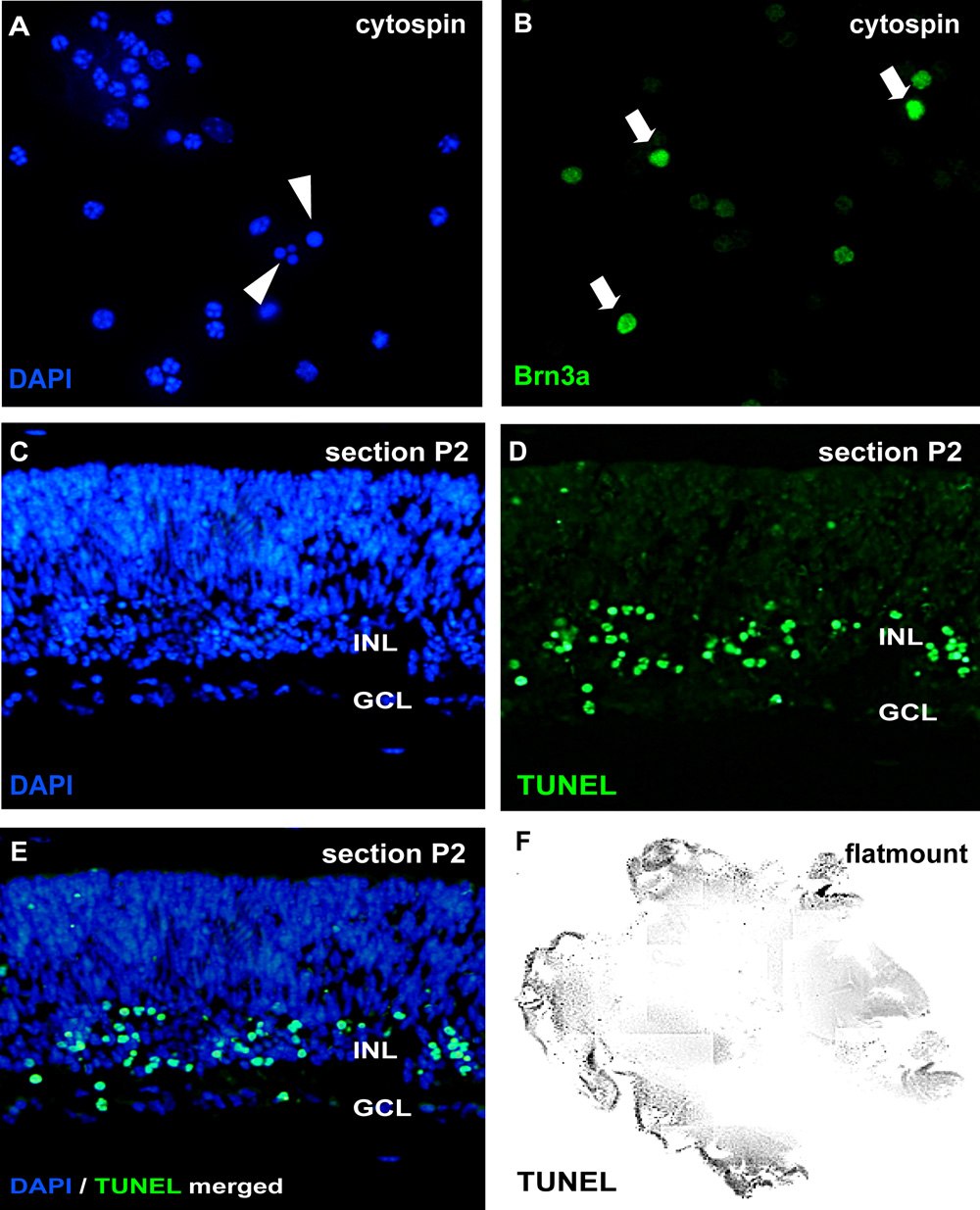
Figuur 2: Analyse van organotypische netvlies wholemount culturen door Cytospin en secties
Voor de analyse van apoptose, zijn cytospins van gescheiden celsuspensies gekleurd door DAPI en pycnotic kernen kunnen worden onderscheiden door nucleaire fragmentatie of chromatine condensatie (pijlpunten in A). Als alternatief, wholemount secties (CE, muizen netvlies postnatale dag (P) 2) of retinale flatmount (F) kan worden onderworpen aan een test en TUNEL tegengekleurd met DAPI (E). Het effect van de behandeling op verschillende retina celtypes gebruik kunnen worden gevisualiseerd in cytospins door neuron specifieke antilichamen, zoals het ganglion cel marker Brn3a (pijlen in B) GCL, ganglion cel laag;. INL, prospectieve binnenste nucleaire laag. Gelieve Klik hier voor een grotere versie van figuur 2 te zien.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Het voordeel van muizen organotypische netvlies wholemount culturen 2-5 meer dan dissociatie, monolaag, retinospheroid of opnieuw geaggregeerde 3D sferoïde culturen een ligt in het behoud van de neuronale interacties en verbindingen, het nabootsen van de in vivo situatie. In vergelijking met het vroegere rapporten 2, onze video artikel geeft een gedetailleerde demonstratie van de eigenaardigheden in enucleatie van muriene ogen en de dissectie van het netvlies van de verschillende ontwikkelingsstadia met inbegrip van verwijdering van de lens en het glasachtig lichaam zonder beschadiging van het netvlies. De verwijdering van de lens en het glasvocht is essentieel voor de farmacologische manipulaties als zowel de toegang van stoffen belemmeren het netvlies lagen. In tegenstelling tot andere gemelde muizen explantatie cultuur-systemen, gebruiken we geen ondersteunende materialen, zoals polycarbonaat membranen 2, voor onze organotypische cultuur, maar de cultuur van de retina wholemount cups vrij zwevend, bovendien het vergemakkelijken van de toegankelijkheid voor exogeen toegepaste stoffen 3-5.
Met behulp van een chemisch welbepaald, serum-en aanvullen-vrije cultuur medium zonder insuline staat slechts voor korte tijd cultuur (24 - 48 uur), maar tegenwicht effecten van insuline op apoptose niveau 3 of growth factor-nabootsen effecten van de FCS en supplementen worden vermeden.
De meeste apoptose en proliferatie studies maken gebruik van MTT testen of FACS-analyse voor de kwantificering van effecten. We echter een stap voor stap demonstratie van de dissociatie van gekweekte netvliezen voor de kwantitatieve apoptose en proliferatie analyse door Cytospin. Voor zover onze ervaring gaat, handmatige telling van de kernen in DAPI of BrdU gekleurde cel plaatsen - ook al vervelend en tijdrovend - is de meest nauwkeurige methode voor het kwantificeren van het netvlies apoptose en proliferatie, in het bijzonder in muriene netvlies.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
De auteurs willen graag E. de la Rosa en AI Valenciano bedanken voor de eerste hulp bij de oprichting van de organotypische culturen en U. Laub en U. Gerster voor technische bijstand.
Materials
| Name | Type | Company | Catalog Number | Comments |
| Mice | Animal | Charles River Laboratories | ||
| Dissection microscope | Tool | Carl Zeiss, Inc. | ||
| PBS | Reagent | Sigma-Aldrich | PBS should be cold (> 4°C) and sterile | |
| Dulbecco`s modified eagle`s medium / nutrient mixture F-12 Ham | Reagent | Sigma-Aldrich | D 8900 | DMEM / F-12 |
| Apo-transferin | Reagent | Sigma-Aldrich | T 1147 | |
| Putrescin | Reagent | Sigma-Aldrich | P 5780 | |
| Sodium selenite | Reagent | Sigma-Aldrich | S 9133 | |
| Progesterone | Reagent | Sigma-Aldrich | P 6149 | |
| Gentamicine | Reagent | Invitrogen | ||
| L-Glutamine | Reagent | Invitrogen | 25030-024 | 200 mM (100X), liquid |
| Bovine serum albumine (BSA) | Reagent | Carl Roth Gmbh | 8076.3 | 30 mg/ml |
| Collagenase | Reagent | Sigma-Aldrich | C 0773 | 200 U/ml |
| Trypsin | Reagent | Sigma-Aldrich | T4799 | From porcine pancreas; 1 mg/ml |
| Hyaluronidase | Reagent | Sigma-Aldrich | H 3884 | 200 mg/ml |
| DNase I | Reagent | Roche Group | 1 284 932 | 10 mg/ml |
| EDTA | Reagent | Sigma-Aldrich | E 6511 | |
| Silicone solution | Reagent | SERVA Electrophoresis | 35130 | |
| Paraformaldehyde (PFA) | Reagent | Sigma-Aldrich | P6148 | 8% PFA in 0.1M phosphate buffer (pH 7.4). |
| 4’,6-diamidino-2-phenylindole dihydrochloride | Reagent | Sigma-Aldrich | D 0542 | DAPI |
| Fluorescent Mounting Medium | Reagent | Dako | S3023 | |
| BrDU | Reagent | Sigma-Aldrich | B 9285 | |
| 96-well plates | Tool | Falcon BD | 3072 | |
| 24-well plates | Tool | Falcon BD | 3047 | |
| Pasteur pipettes | Tool | Brand GmbH | 747720 | |
| Forceps DUMONT #5 | Tool | Fine Science Tools | 11252-30 | bevelled very fine shanks (0.05 mm x 0.02 mm tip) |
| Forceps DUMONT #7 | Tool | Fine Science Tools | 11271-30 | curved shanks (0.07 mm x 0.10 mm tip) |
| Spring scissors,straight, 8cm | Tool | Fine Science Tools | 15000-00 | fine, small straight blades |
| Standard scissors, straight, sharp/blunt | Tool | Fine Science Tools | 14007-14 | Use for decapitation or cervical dislocation |
| Eppendorf tubes | Tool | Eppendorf | 2ml; round bottom for better precipitation of pellet during centrifugation /cytospin | |
| Cooling centrifuge | Tool | Eppendorf | ||
| Rotation shaker | Tool | CAT | ||
| Cytospin | Tool | Thermo Fisher Scientific, Inc. |
References
- Rieke, M., Gottwald, E., Weibezahn, K. -F., Layer, P. G. Tissue reconstruction in 3D-spheroids from rodent retina in a motion-free, bioreactor-based microstructure. Lab. Chip. 8, 2006-2213 (2008).
- Donovan, S. L., Dyer, M. A. Preparation and square wave electroporation of retinal explant cultures. Nature Protocols. 1, 2710-2718 (2006).
- Duenker, N., Valenciano, A. I., Franke, A., Hernandez-Sanchez, C., Dressel, R., Behrendt, M., de Pablo, F., Krieglstein, K., de la Rosa, E. J. Balance of pro-apoptotic transforming growth factor-beta and anti-apoptotic insulin effects in the control of cell death in the postnatal mouse retina. Eur. J. Neurosci. 22, 28-38 (2005).
- Franke, A. G., Gubbe, C., Beier, M., Duenker, N. Transforming growth factors beta and Bone morphogenetic proteins: Cooperative players in chick and murine programmed retinal cell death. J. Comp. Neurol. 495, 263-278 (2005).
- de la Rosa, E. J., Díaz, B., De Pablo, F.
- Dohle, D. S., Pasa, S. D., Gustmann, S., Laub, M., Wissler, J. H., Jennissen, H. P., Duenker, N. Chick ex ovo culture and ex ovo CAM assay: How it really works. J Vis Exp. 32, (2009).

{kind=link}
{kind=link}