

Summary
Мы описываем методологию объединения автоматизированных культивирования клетки с высоким содержанием изображений для визуализации и количественной оценки нескольких клеточных процессов и структур, в высокой пропускной образом. Такие методы могут помочь в дальнейшей функциональной аннотации геномов, а также выявление заболеваний генных сетей и потенциальных лекарственных препаратов.
Protocol
1. Автоматизированная спаечного процесса культуре клеток (рис. 1) 20
- Подготовка автоматизированной системы клеточных культур для ввода пластин клеточной культуры. Нагрузка расходные материалы (например, наконечники для пипеток, тарелки культуре клеток, анализа пластин) в системе с помощью графического интерфейса пользователя (GUI). Обеспечить достаточную медиа культуре клеток, фосфатным буферным раствором (PBS) и трипсина в роботизированной системы.
- Вручную семена два omnitray плиты с 2 х 10 6 клеток на чашку, из SH-SY5Y линии клеток нейробластомы. Поддерживать клетки в Opti-MEM с 10% эмбриональной бычьей сыворотки (FBS). Положите пластины в робота культуре клеток с помощью графического интерфейса. Клетки будут инкубировали при 37 ° С и 5% CO 2.
- Выберите, какой культуре клеток протокола должна быть начата 20. Можно выбрать из спаечного процесса клеточной культуры 22, культивирования и расширения эмбриональных стволовых (ЭС) клеток на мышь фидерных клеток 23 или культивирование подвески челLs 22.
- Выберите протокол приверженцем культуры клеток (рис. 1) и обеспечить приверженцем клеточной линии определенные файлы параметров подбираются таким образом, что слияние порог (площадь пластинки omnitray, которая содержит клетки) установлена на уровне 70%. Установить общее время трипсинизации до двух минут.
- Поручить робот для подготовки новых пластин omnitray, с рядом посева ячейки 2 х 10 6 клеток на чашку.
- Входной omnitray пластин в системе культуры клеток с использованием автоматизированной приверженцем протокол культуры клеток (рис. 1). Этот протокол включает в себя следующие этапы: Планшеты инкубируют и отображаемого пока они не достигнут предопределенного слияния порога. Если клетки не достигают порога слияния в течение 5 чтений, тарелки будут удалены из системы. По достижении определенного пользователем слияния порог, клетки промывают, трипсином и подсчитаны. Заранее определенного числа клеток добавляются новые пластины omnitray и если Есть достаточное число клеток, указанное онемениеэ пробирного пластины транспортируются на палубу и определенное количество клеток разливали в каждую лунку. Анализ пластин может быть напрямую отображаемого с помощью встроенного микроскопа или выход из системы для дальнейшей обработки.
- Поручить системы для подготовки анализа 4 пластины на пластину omnitray, в общей сложности 5000 клеток на лунку.
2. shRNA производства вирусов и покрытие в анализе пластин (Необходимое время: 6 дней)
- Рост запасов бактериальной глицерин содержащие shRNA векторов (Open Biosystems, TRC1) в течение ночи в 2 мл Лурия-Бертани среда сред, содержащих 100 мкг / мл ampicilin (Sigma-Aldrich).
- Извлечение плазмид следующий протокол производителя (Promega Мастер MagneSil TFX).
- Продукция вирусов с помощью РНК-интерференции Консорциум высокой пропускной лентивирусов Производство (96 лунками) протокола 24. Работа с лентивирус относительно безопасно, потому что вирусные частицы используются для трансдукции являются репликации с дефицитом и сплит-стратегии ген упаковки используются для их производства. Тем не менее, при работе с лентивирус, дополнительных процедур биобезопасности необходимо свести к минимуму риск для себя и других 25. Все эксперименты должны проводиться в лабораторном уровне безопасности MLII или BSL2. Все пластмасс (пипетки, пластиковой посуды, средств массовой информации), которые были в контакте с лентивирус частиц должна быть инкубировали с хлоркой в течение 24 часов перед удалением.
- Рассчитать множественности заражения (МВД) из лентивирусов, определяя процент GFP-положительных клеток использованием pLKO.1 GFP плазмиды (Sigma-Aldrich).
- Пластина лентивирус в анализе пластин, с МВД 3.
3. Лентивирусов трансдукции и нейронных дифференциации SH-SY5Y клеток (Необходимое время: 6 дней)
- Клетки будут добавлены в анализе пластин (см. шаг 1.7). Нагрузка анализа чашки, содержащие shRNA лентивирус в автоматизированной системе культуры клеток.
- После 24 часов, средств массовой информации наАнализ пластин будет изменен на Opti-MEM, содержащую 0,5% FBS и 0,1 мкМ ретиноевой кислоты, чтобы начать процесс дифференциации. Дифференциация SH-SY5Y клетки позволяет выявлять neuritic структур и синхронизирует деление клеток.
- Продолжить инкубации анализа пластин в дифференциации средств массовой информации в течение 5 дней. Это обеспечивает максимальное нокдаун целевых экспрессии генов.
- На 5 день, добавить 50 мкМ H 2 O 2 до половины анализа пластин в течение 24 часов, чтобы стимулировать транслокацию DJ1 в митохондриях.
- На 6-й день, добавьте Mitotracker CmxROS (Invitrogen) для элементов, в конечной концентрации 200 нМ на лунку и инкубировать при температуре 37 ° С в течение 30 минут.
- Можно указать системе, чтобы изображение пластины непосредственно с помощью тепловизора HC или пластины могут быть экспортированы из системы для дальнейшей обработки.
4. Автоматизированная иммуноокрашивания пробирного пластин (Необходимое время: 2 дня)
Качество изображения paramounт для ведения оперативной и надежной ЖКХ. Повреждение сотовой монослоя из-за неточности пипетки может привести к ухудшению качества изображения и невоспроизводимые результаты. Чтобы свести к минимуму повреждения клеток слой, иммунной был проведен с использованием роботов станции. Процедура очень похожа на тот, который был описан ранее 26, но был настроен для увеличения пропускной способности и снижения расходных использования.
- Fix клеток с 100 мкл 4% параформальдегида предварительно нагревают до 37 ° C. Инкубируйте в течение 20 минут при комнатной температуре.
- Вымойте клеток с 200 мкл PBS в течение 5 минут, 3 раза.
- Инкубируйте анализа пластин с 200 мкл PBS, содержащего 0,1% Тритон (PBST) в течение 10 минут.
- Вымойте клеток с 200 мкл PBS в течение 5 минут, 3 раза.
- Инкубируйте анализа пластин с 200 мкл буфера блок (PBST с 5% FBS) в течение 1 часа при комнатной температуре.
- Вымойте клеток с 200 мкл PBS в течение 5 минут, 3 раза.
- Инкубируйте с Фоллявследствие первичных антител ночи при 4 ° С:
- Коза DJ1 N20 (Санта-Крус, 5 мкг / мл)
- Кролик β-III тубулина (Sigma-Aldrich, 1 мкг / мл)
- На следующий день, мыть ячейки с 200 мкл PBS в течение 5 минут, 3 раза.
- Инкубируйте анализа пластин со следующими вторичными антителами 1 час при комнатной температуре:
- AlexaFluor 488 осла анти-козел (Invitrogen, 2 мкг / мл)
- AlexaFluor 647 козий анти-кролик (Invitrogen, 2 мкг / мл
- Вымойте клеток с 200 мкл PBS в течение 5 минут, 3 раза.
- Инкубируйте клетки с Hoechst (Invitrogen; 1 мкг / мл) в течение 10 минут.
- Вымойте клеток с 200 мкл PBS в течение 5 минут, 3 раза.
- Магазин пластин при 4 ° С, пока они не могут быть отображены.
5. Высокое содержание получения изображений и анализа изображений (Необходимое время: 5 дней)
- Изображение в общей сложности 30 полей в хорошо использование 20x объектив. Визуализируйте DJ1 с FITC набор фильтров, митохондрии с TRITC набор фильтров, β-III тубулина с Cy5 набор фильтров и ядер с использованием УФ-фильтром набор (рис. 3).
- Анализ изображений с помощью Compartmental Bioapplication анализа (Cellomics, ThermoFisher), чтобы определить среднюю интенсивность Mitotracker сигнала в митохондрии. (4B рис, F).
- Для определения коэффициента среднего перекрытия между DJ1 и митохондриях, анализ изображений с помощью Cellomics bioapplication Colocalisation (Cellomics, ThermoFisher). Определить регионы интереса (ROI) следующим образом: ROI - ядра (рис. 4а, E), рентабельность инвестиций B - митохондрии (рис. 4 B, F). Исключить ROI от ROI B обеспечить анализ только цитоплазмы. Определите, как митохондрии целевом регионе я, как и DJ1 целевом регионе II (рис. 4С, G).
- Анализ изображений с помощью нейронных bioapplication Профилирование (Cellomics, Thermofisher) проследить средней длины нейритов от β-III тубулина окрашивания (рис.4D, H).
- Изображение пластины использованием оперы LX автоматизированных конфокальной читателя (Perkin-Elmer). Изображение в общей сложности 30 полей в хорошо использование 60x объектив с погружения в воду. Визуализируйте митохондрии с 561 нм и ядер с УФ возбуждением.
- Анализ изображений с помощью Spot-Edge-Ридж (ГЭЭ) алгоритм текстуры особенности. SER-Ридж фильтр передает интенсивности в пикселях формирования гребня-подобных моделей. Более фрагментированной митохондрий, тем выше SER-Ридж балл (рис. 8).
6. Данные нормализации и анализа
- Импорт данных из программного обеспечения для анализа изображений в Bioconductor CellHTS2 пакет для программной среды R (R версии 2.11.1, Bioconductor версия 2.6).
- Логарифм по основанию (2) преобразования данных до на чашку средний основан нормализацию 27, 28. Не применять дисперсию регулировки на чашку.
- Чтобы определить модификаторов фенотипа, используйте два способа ANOVA между differeнт группах т.е. Яичница инфицированных по сравнению с необработанными клетками платные токсин обработанных клеток по сравнению с гена-мишени по сравнению с необработанными клетками гена-мишени обработанных клеток (рис. 5-7).
7. Представитель результаты
Мутации в DJ1 приводят к раннему развитию паркинсонизма-рецессивным 21, но неясно, как потеря DJ1 порождает болезнь фенотип. Известно, что клетки недостаточно из DJ1 более восприимчивы к окислительным стрессом гибель клеток и в ответ на окислительный стресс, DJ1 перемещает из цитоплазмы в митохондрии 29, 30. Построив HC анализы, чтобы контролировать эти фенотипы, мы можем идентифицировать гены, которые регулируют или затрагивают фенотип связаны с DJ1. Этот подход может помочь расшифровать пути, в котором DJ1 функции и которые могут быть задействованы в патогенезе болезни.
Пример эпистатических взаимодействия с DJ1 (рис. 5): Нокдаун от DJ1 в клетках, подвергавшихсядля токсина приводит к большей потере жизнеспособности клеток (БАР-B: образ-Б) по сравнению с клетки, инфицированные с яичницей лентивирусов (БАР-: изображения). Нокдаун целевой ген имеет такой же эффект, что наблюдается в клетках с DJ1 нокдаун (БАР-C: изображение-C). Нокдаун как DJ1 и целевого гена приводит к значительно большей потере жизнеспособности клеток, чем потеря любого гена в одиночку (БАР-D:-D изображение). Это говорит о том эпистатических взаимодействия между DJ1 и гена-мишени А.
Пример ген регулирующий DJ1 транслокации (рис. 6): Когда клетки подвергаются воздействию токсинов, DJ1 перемещает из цитоплазмы в митохондрии, которые можно измерить с помощью выше коэффициент перекрытия между DJ1 и митохондриях (BAR-A: изображение по сравнению с БАР-C: изображение C). В клетках, где B гена-мишени был замолчать, менее DJ1 перемещает в митохондриях, когда клетки подвергаются воздействию токсинов. Это означает, что B гена-мишени участвует в перевозке DJ1 в митохондриях. (БАР-B: изображение B и БАР-D: изображение D)
Пример ген, участвующий в нейронных результатом (рис. 7): Нокдаун целевой ген С в дикого типа SH-SY5Y клеток приводит к значительному увеличению длины аксонов (БАР-B: образ-Б) по сравнению с клетки, инфицированные лентивирус выражения вскарабкался shRNA (BAR-A: изображение). Этот эффект теряется в клетки инкубировали с токсином (БАР-C и D).
Пример ген, участвующий в митохондриальной морфологии (рис. 8): Инфекция дикого типа SH-SY5Y клеток с shRNA таргетинг гена D приводит к снижению митохондриального SER-Ридж сегментации значение (рис. 8, изображение В и Г) по сравнению с клетки, инфицированные с яичницей лентивирусов (рис. 8, изображения и В).
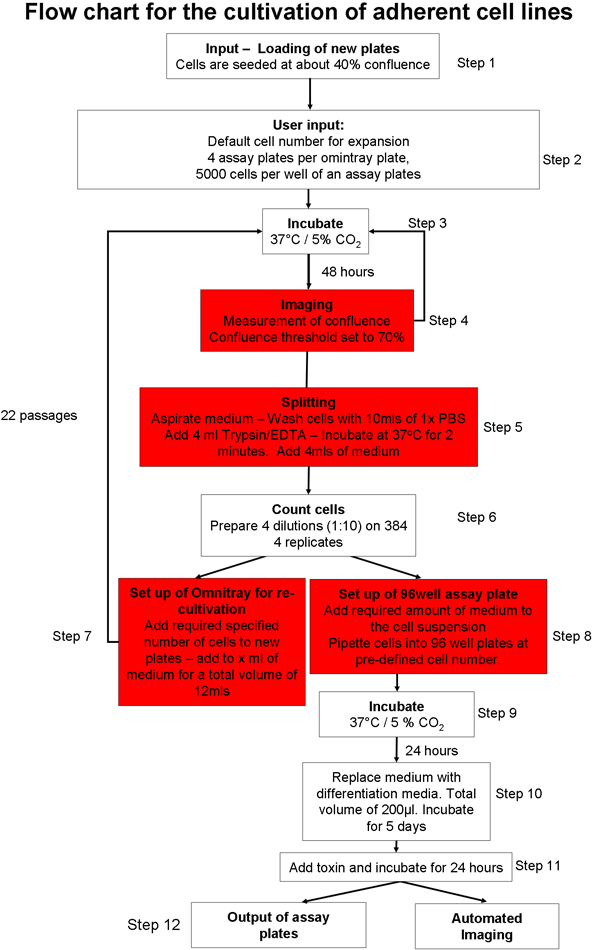
Рисунок 1 План автоматизированный протокол культуре клеток.:

Рисунок 2 Схема обзор процесса отбора, анализа изображений и статистических методов, используемых в процессе скрининга. Я) Клетки культивируются, пока они не сливающийся, а затем высевали в чашки, содержащие анализ лентивирус shRNA. Клетки дифференцированы в течение 5 дней и токсинного затем добавляется к пластинам в течение 24 часов. Анализ пластины выводятся из системы и immunostained. Время, затраченное для каждого из процессов, указанных в скобках. II) Данные получены с помощью тепловизора HC (жизнеспособность клеток, белков транслокации и нейронных вырост) и автоматизированных тепловизор конфокальной (митохондриальная морфологии). Данные экспортируются в CellHTS2 продукта в пределах R, базовые (2) журнал преобразован и нормированы. Двусторонний ANOVA используется для идентификации значимых взаимодействий между различными переменными.
Рисунок 3. Композитных изображений клеток, приобретенных HC изображений. ) Необработанных клетках, В) клеток, обработанных H 2 O 2. DJ1 помечена зеленым цветом, митохондрии в красном и ядер в синий цвет. Нейритов окрашивания не подсвечивается.

Рисунок 4. Количественная оценка несколько сотовых функции получена из ЖКХ. Н.э.) необработанных SH-SY5Y клеток. EH) H 2 O 2 рассматривается SH-SY5Y клеток. , E) сегментация ядра и определение рентабельности, B, F) выявление и количественное определение митохондрии, ROI B, C, G) определение DJ1, Target канал II, D, G) Определение и расчет средней длины аксонов. Клетки близко к краю изображения, исключаются из анализа. Вставка изображений изображения до анализа.

Рисунок 6. Определение гена регулирующего DJ1 транслокации (письма от бара соответствует надписи на изображениях). Изображения через D являются SH-SY5Y клетки помечены для DJ1 (зеленый), митохондрии (красный) и ядра (синий), которые были использованы для количественного определения DJ1 транслокации в митохондрии.

Рисунок 7. Идентификация генов, участвующих в нейронных результат (письма от бара соответствует надписи на изображение). Изображения через D являются SH-SY5Y клетки помечены для β-III тубулина (зеленый) и ядра (синий), которые были использованы для количественного определения нейритов длины.

Рисунок 8. Определение генов участвуют в регуляции митохондриальной морфологии. Изображение А и С являются составными образами SH-SY5Y клетки, инфицированные с яичницей shRNA или shRNA таргетинг гена D соответственно. Митохондрии окрашены в красный в то время как ядра окрашены в синий цвет. Изображение B и D. визуализации SER-Ридж количественной оценке.
Discussion
С уменьшением расходов на HT / HC системах сотовой скрининга, в сочетании с наличием мощной генома инструменты для изменения функции генов, HT / HC экраны становятся обычным явлением в академических кругах. Этот подход уже был успешно применен в различных областях исследований, таких как идентификация лекарственных мишеней в 9 рака, 31-33 и 34-36 эмбрионального развития и даже имеет потенциал для применения в расшифровке путей, участвующих в нервно-психических расстройств 37,38. Однако реализация такой системы требует значительных затрат времени и усилий благодаря оптимизации процессов часто принимает не менее 6 месяцев. Все шаги, такие как трипсинизации раз, пипетки скорости и плотности посева, должны быть скорректированы, чтобы гарантировать, что клетки здоровы и растут постоянно. Предупреждение бактериального загрязнения является одной из самых сложных задач, стоящих перед автоматизированной культуре клеток с еженедельная уборка протоколов сombination с постоянной промывки всех жидких линий подшипник с 70% этанола являются необходимыми для загрязнения свободной культуры. Это будет также необходимо для улучшения робототехники, так что дополнительные инструменты, такие как конфокальные микроскопы для более высокого разрешения и -80 ° C морозильники для хранения соединения могут быть интегрированы.
Существуют также ограничения, которые необходимо решить для улучшения чувствительны, скорость и полезности этого метода для изучения генных сетей и выявить гены, вовлеченные в патогенные молекулярных путей.
Провести HT / HC экран и убедиться, что надежные данные собираются, некоторые аспекты должны быть оптимизированы. Во-первых, надежность измерений имеет первостепенное значение и зависит от устойчивости и чувствительности анализа. Например, тесты, описанные выше, подходит для небольших экранов, но трудно реализовать на геном масштабе, по причине большого числа этапов обработки, требуется до получения изображений.Таким образом, можно было бы построить стабильные клеточные линии выражения гена-репортера, который позволил бы для прямой визуализации и привести к снижению изменения из-за сокращения числа этапов обработки. В настоящее время проектирование анализа, который точно изображает и надежно количественно фенотип интерес является основным узким местом в процессе скрининга HC.
Многие экраны проводятся в клетках млекопитающих с использованием различных библиотек RNAi, все из которых страдают от вне целевых эффектов, ограниченное эффективности глушителей гена и неполный охват генома. Таким образом библиотеки должны быть сделаны, которые являются более конкретными, сильнодействующих и лучшего покрытия сетей. Предпринимаются усилия для создания такой есть надежда, такие усилия улучшат воспроизводимость HT / HC scre ан хитов.
Ограничение многих крупных масштабах ячейке на основе экранов является то, что они проводятся в клетках нейробластомы, потому что они могут быть генетически модифицированных и культурной большому числу с относительной легкостью. Тем не менее, актуальность "хитов", определенные в бывшей модели ячейки естественных условиях культуры в естественных условиях функции сомнительна, тем более, что мозг состоит из высоко специализированных типов клеток, которые образуют плотный и сложный сети синаптических связей функционировать как высоко интегрированную единицу. Как следствие, он является общим, который бьет идентифицируются с помощью скрининга подход, описанный выше, проверяются в средних экранов с помощью дополнительных методов и в более физиологическое соответствующих моделей 39. Для улучшения перевод хитов, выявленные в ходе ЖКХ, более представительным и сложных моделей, например, первичных элементов и дифференцированные стволовые клетки или со-культуре системы должны быть разработаны и адаптированы для HT / HC подходов.
ntent "> В сочетании с автоматизированной культивирования клеток и HC изображений можно быстро получить новые знания о том, нейроны функцию и определить, какие пути имеют важное значение для развития болезни. Объединение ЖКХ / HTS данных информацию, полученную от подходов других" omics ", он будет Тогда можно построить обзор системной биологии заболеваний головного мозга, способствуя тем самым терапевтический развития.Disclosures
Нам нечего раскрывать
Acknowledgments
Мы благодарим Гамильтон программистов и специалистов для постоянной поддержки и Ева Blaas для оказания технической помощи. Эта работа была поддержана двумя NWO инвестиционных грантов (911-07-031 и 40-00506-98-10011), Prinses Беатрикс Fonds Wetenschapsprijs 2009 и неврологии Campus Амстердаме; SJ поддерживается Ti-Фарма: T5-207.
Materials
| Name | Company | Catalog Number | Comments |
| AI.CELLHOST | Hamilton Co | http://www.hamiltonrobotics.com/en-uk/applications/cellomics/ | |
| OPTI-MEM | Invitrogen | 31985-054 | |
| RETINOIC ACID | Sigma-Aldrich | R2625 | |
| OMNITRAY PLATES | Nalge Nunc international | 465219 | |
| 96 WELL CULTURE PLATES | Greiner Bio-One | 655086 | |
| DJ1 N20 ANTIBODY | Santa Cruz Biotechnology, Inc. | SC27004 | |
| BETA-III TUBULIN ANTIBODY | Sigma-Aldrich | T3952 | |
| MITOTRACKER CMXROS | Invitrogen | M-7512 | |
| H–CHST-33342 | Invitrogen | H1399 | |
| HYDROGEN PEROXIDE | Sigma-Aldrich | 216763-100ML | |
| TRYPSIN | Invitrogen | 25050014 | |
| DULBECCO’S PHOSPHATE BUFFERED SALINE | Invitrogen | 14190086 | |
| PROMEGA WIZARD MAGNESIL TFX | Promega Corp. | A2380 | |
| SHRNA CLONES | Open Biosystems | http://www.openbiosystems.com/RNAi/shRNALibraries/ TRCLibraryDetails/ | |
| CELLOMICS BIOAPPLICATIONS | Thermo Fisher Scientific, Inc. | http://www.thermo.com/hcs |
References
- Geschwind, D. H., Konopka, G. Neuroscience in the era of functional genomics and systems biology. Nature. 461, 908-915 (2009).
- Ge, H., Walhout, A. J., Vidal, M. Integrating 'omic' information: a bridge between genomics and systems biology. Trends. Genet. 19, 551-560 (2003).
- Zhu, H., Snyder, M. 'Omic' approaches for unraveling signaling networks. Curr. Opin. Cell. Biol. 14, 173-179 (2002).
- Jain, S., Heutink, P. From single genes to gene networks: high-throughput-high-content screening for neurological disease. Neuron. 68, 207-217 (2010).
- Manolio, T. A. Finding the missing heritability of complex diseases. Nature. 461, 747-753 (2009).
- Nalls, M. A. Imputation of sequence variants for identification of genetic risks for Parkinson's disease: a meta-analysis of genome-wide association studies. Lancet. 377, 641-649 (2011).
- Simon-Sanchez, J. Genome-wide association study confirms extant PD risk loci among the Dutch. Eur. J. Hum. Genet. 19 (6), 655-661 (2011).
- Van Regenmortel, M. H. Reductionism and complexity in molecular biology. Scientists now have the tools to unravel biological and overcome the limitations of reductionism. EMBO. Rep. 5, 1016-1020 (2004).
- Aherne, G. W., McDonald, E., Workman, P. Finding the needle in the haystack: why high-throughput screening is good for your health. Breast. Cancer. Res. 4, 148-154 (2002).
- An, W. F., Tolliday, N. J. Introduction: cell-based assays for high-throughput screening. Methods. Mol. Biol. 486, 1-12 (2009).
- Mayr, L. M., Bojanic, D.
- Hertzberg, R. P., Pope, A. J. High-throughput screening: new technology for the 21st century. Curr. Opin. Chem. Biol. 4, 445-451 (2000).
- Conrad, C., Gerlich, D. W. Automated microscopy for high-content RNAi screening. J. Cell. Biol. 188, 453-461 (2010).
- Thomas, N. High-content screening: a decade of evolution. J. Biomol. Screen. 15, 1-9 (2010).
- Arrasate, M., Finkbeiner, S. Automated microscope system for determining factors that predict neuronal fate. Proc. Natl. Acad. Sci. U. S. A. 102, 3840-3845 (2005).
- Dragunow, M.
- Varma, H., Lo, D. C., Stockwell, B. R. High throughput screening for neurodegeneration and complex disease phenotypes. Comb. Chem. High. Throughput. Screen. 11, 238-248 (2008).
- Durr, O. Robust hit identification by quality assurance and multivariate data analysis of a high-content, cell-based assay. J. Biomol. Screen. 12, 1042-1049 (2007).
- Miller, J. Quantitative relationships between huntingtin levels, polyglutamine length, inclusion body formation, and neuronal death provide novel insight into huntington's disease molecular pathogenesis. J. Neurosci. 30, 10541-10550 (2010).
- Jain, S., Sondervan, D., Rizzu, P., Bochdanovits, Z., Caminada, D., Heutink, P. The Complete Automation of Cell Culture. Journal of Biomolecular Screening. 16 (8), (2011).
- Bonifati, V. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science. 299, 256-259 (2003).
- Ricardo, R., Phelan, K. Trypsinizing and Subculturing Mammalian Cells. J. Vis. Exp. (16), e755-e755 (2008).
- Kent, L. Culture and Maintenance of Human Embryonic Stem Cells. J. Vis. Exp. (34), e1427-e1427 (2009).
- Moffat, J. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 124, 1283-1298 (2006).
- Volksgezondheid, M. V. Integrale versie van de Regeling genetisch gemodificeerde organismen en het Besluit genetische gemodificeerde. Organismen. , (2004).
- Anderl, J. L., Redpath, S., Ball, A. J. A Neuronal and Astrocyte Co-Culture Assay for High Content Analysis of Neurotoxicity. J. Vis. Exp. (27), e1173-e1173 (2009).
- Wiles, A. M., Ravi, D., Bhavani, S., Bishop, A. J. An analysis of normalization methods for Drosophila RNAi genomic screens and development of a robust validation scheme. J. Biomol. Screen. 13, 777-784 (2008).
- Canet-Aviles, R. M. The Parkinson's disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. U. S. A. 101, 9103-9108 (2004).
- Blackinton, J. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 284, 6476-6485 (2009).
- Gupta, P. B. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138, 645-659 (2009).
- Luo, J. A genome-wide RNAi screen identifies multiple synthetic lethal interactions with the Ras oncogene. Cell. 137, 835-848 (2009).
- Mouchet, E. H., Simpson, P. B. High-content assays in oncology drug discovery: opportunities and challenges. IDrugs. 11, 422-427 (2008).
- Vogt, A. Automated image-based phenotypic analysis in zebrafish embryos. Dev. Dyn. 238, 656-663 (2009).
- Pardo-Martin, C.
- Vogt, A., Codore, H., Day, B. W., Hukriede, N. A., Tsang, M. Development of automated imaging and analysis for zebrafish chemical screens. J. Vis. Exp. (40), e1900-e1900 (2010).
- Ross, P. J., Ellis, J. Modeling complex neuropsychiatric disease with induced pluripotent stem cells. F1000. Biol. Rep. 2, 84-84 (2010).
- Ebert, A. D., Svendsen, C. N. Human stem cells and drug screening: opportunities and challenges. Nat. Rev. Drug. Discov. 9, 367-372 (2010).
- An, W. F., Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 45, 180-186 (2010).
