

Summary
En snabb
Abstract
Avsökning av föreningar för in vivo-aktivitet kan användas som ett första steg för att identifiera kandidater som kan utvecklas till farmakologisk agens 1,2. Vi utvecklat en ny nanoinjection / elektrofysiologi analys som möjliggör detektering av bioaktiva modulerande effekter av föreningar på funktionen hos en neuronal krets som medierar flyktrespons i Drosophila melanogaster 3,4. Vår analys in vivo, som använder Drosophila Giant fibersystemet (GFS, figur 1) medger screening av olika typer av föreningar, såsom små molekyler eller peptider, och kräver endast minimala mängder för att framkalla en effekt. Dessutom erbjuder Drosophila GFS ett stort antal potentiella molekylära mål på nervceller och muskler. Giant fibrer (GF) synaps elektriskt (gap junctions) och kemiskt (kolinergisk) på en perifer Synapsing interneuron (PSI) och den Tergo trochanter-Muskel neuronen (TTMn) 5 6. Slutligen, den neuromuskulära korsningar (NMJ) av TTMn och DLMn med hopp (TTM) och muskler Flight (DLM) är glutamaterg 7-12. Här visar vi hur du ska injicera metod i nanoliter mängder av en förening, medan få elektrofysiologiska intracellulära inspelningar från Giant Fiber System 13 och hur man övervaka effekterna av föreningen på funktionen av denna krets. Vi visar specificiteten hos analysen med methyllycaconitine citrat (MLA), en nAChR-antagonist, som stör PSI till DLMn förbindelse men inte GF att TTMn anslutning eller funktionen av NMJ vid hopp eller muskler flygning.
Innan du börjar denna video är det viktigt att du noggrant titta på och bekanta dig med Jové video med titeln "Elektrofysiologiska inspelningar från Giant Fiber vägen för D. Melanogaster "från Augustin et al 7, som video presenteras här är avsedd som en expansion till denna befintliga teknik. Här använder vi elektrofysiologiska inspelningar metoden och fokus i detalj endast om tillsättning av de parade nanoinjections och övervakningsteknik.
Protocol
1. Elektrofysiologi Rigg Set-up
- Erforderlig utrustning för elektrofysiologi riggen set-up beskrivs i detalj av Augustin et al. i denna tidskrift 14. Se till denna artikel för en detaljerad förklaring av de nödvändiga elektrofysiologiska apparater.
- Ändra den tidigare beskrivna elektrofysiologi rig set-up 14 genom att lägga till 1/6 mikromanipulator, som håller nanoinjector. För enkel åtkomst till huvudet av djuret bör placeras mellan de två stimulerande elektrod mikromanipulatorer som visas i figur 2.
- Innan du startar experimentet se till att du har en bekväm sortiment på alla axlar rotation med alla mikromanipulatorer, och att alla elektroder samt injektionen mikropipett (se steg 2) kan nå djuret.
2. Nanoinjection Uppsättning
- Den nanoinjection set-up kräver en Nanoliter2000 (World Precisipå instrument, Sarasota, FL, USA) eller liknande typ av injektor som tillåter kontrollerade injektioner metod i nanoliter mängder.
- Förbered injektionsnål med de glas mikropipetter medföljer injektorn genom att dra dem till en resistans på 80-100 MQ med en elektrod avdragare.
- För släta injektioner det är nödvändigt att fasa av Mikropipetter för en 11-17 m öppning i 45 graders vinkel (Figur 3).
- Sakta återfyllning injektionen mikropipett med syntetisk olja med hjälp av en Hamilton spruta enligt instruktioner från Nanoliter2000 manuella och se till att inga luftbubblor finns.
- Försiktigt fast mikropipett på nanoinjector och förbereda den för att ladda föreningen genom att tömma överflödig olja enligt anvisningar från Nanoliter2000 manual.
- Placera injektorn på mikromanipulator och ladda föreningen enligt instruktionerna i Nanoliter2000 manualen. Se till att spetsen på mikropipett inte gå sönder under dettaförfarandet.
- Ställ in önskad mängd nanoliter att injiceras i injektorn kontroll box enligt anvisningar från Nanoliter2000 manual. Observera att den totala mängden injicerade bör inte överstiga 100 nl. Vi fann att större mängder av saltlösning kontrollösningar kan påverka funktionen hos GFS kretsen.
- Det är viktigt att ha injektorn kopplas bort från manöverboxen under inspelning förvärvet med undantag för själva injektionen, eftersom strömmen av nanoinjector stör inspelningar som syns som bakgrundsbrus (Figur 4). Men du inte koppla ifrån strömmen till styrenheten som det kommer åter ställa in den.
3. Drosophila melanogaster Framställning
- Bedöva 2 till 6 dagar gamla flugor med CO 2 eller på is som tidigare beskrivits 14,15.
- När orörliga, använd en pincett för att överföra djuret till en liten skylt wie mjuka dentalvax genom att plocka upp det från sina ben. Observera att en manlig bananfluga väger ca 1,0 mg och en kvinnlig bananfluga väger ca 1,2 mg, vilket förening till kroppsvikt förhållandet är annorlunda i manliga kontra kvinnliga flugor. Därför rekommenderas att endast använda ett kön för experimenten.
- Som tidigare beskrivits 14,15 försiktigt montera flugan ryggsidan uppåt och se till att bröstkorgen och huvudet är immobiliserade med mjuk dentalvax placeras runt kroppen. Försiktigt sprida vingarna ut så att de låg vinkelrätt mot bröstkorgen (figur 2, C). Flugan ska monteras med så lite skada som möjligt.
4. Parade Nanoinjection / elektrofysiologi
- Placera den monterade flugan på elektrofysiologi riggen med huvudet mot stimulerande elektroderna.
- Spetsa djuret med motsvarande stimulerande, mark och elektroder inspelning som tidigare beskrivits 14,15 (FIGUR 2, C). Om inte annat så önskas, placera en inspelning elektrod i DLM och den andra till TTM muskeln. Den DLM muskeln är beläget i bröstkorgen mellan främre Dorso-Central hår och mittlinje flugan. TTM muskeln ligger nära vingens fästen, mellan den bakre och främre Supra-Alar hår av flugan 7.
- Anpassa injektion mikropipett som innehåller föreningen med centrum av de tre ocelli belägen på den mediala bakre delen av huvudet, men som inte ännu injicera (figur 2, C).
- Innan föreningen injektion få en baslinje inspelning av GF för att TTM och GF för att DLM vägar för Giant Fiber System (GFS, figur 1) via hjärnstimulering. För att göra detta, aktivera Giant fibrer (GFS) med 10 tåg med 10 stimuli (40-50 mV) vardera ges för en period av 0,03 ms vid 100 Hz med en 1 sekund fördröjning mellan tågen 14,15 (Figur 4). Vildtyp flyger shOuld kunna följa en-till-en i denna takt av stimulering för både DLM och vägar TTM. Kassera flugan om GF till DLM och GF för att TTM vägar inte följer 100 Hz stimulering vid ett 1:1 förhållande.
- Växla till kontinuerlig stimulering av GF med enkla pulser vid 1 Hz (Figur 4).
- Snabbt ansluter injektorn i kontrollboxen. Även om bakgrundsljud stör de 1 inspelningarna Hz stimulering, inte avbryta det.
- Försiktigt in injektion mikropipett in i huvudet kapsel av flugor strax under cuticule, och injicera den önskade mängden av föreningen i hemolymfa av flugor under bibehållande av 1 Hz stimulering (figur 4). På grund av den öppna cirkulationssystem fluga, kommer hela nervsystemet exponeras för föreningen inom några sekunder. Även en viss injektionsstället är inte kritisk för att leverera föreningar i hemolymfa, finner vi den region i ocelli, som lokaliserard mediala vid den mest dorsala sidan av huvudet kapsel, för att vara ett lämpligt ställe som medger en enkel injektion som leder till snabb och jämn fördelning av föreningen.
- Omedelbart ta bort injektionsstället mikropipett från injektionsstället och dra ur injektorn från kontrollboxen men fortsätter det 1 Hz GF stimulering upp till 1 min efter injektion (Figur 4).
- För att visa mer subtila effekter av föreningar på GFS, betona Giant fibrer (GFS) med 10 tåg med 10 stimuli på 100 Hz med en 1 sekund fördröjning mellan tågen. Att fortsätta att testa funktionen av GF vägar med denna paradigm var 5 minuter upp till 15 minuter (Figur 4). Men kortare intervaller eller längre perioder övervakning är också möjliga.
- För att testa om föreningen har en effekt på den neuromuskulära korsningar (NMJs) i GFS, och eventuellt begränsa effekterna av föreningen, fortsätt att aktivera motorn neuroner riktningtly av bröstkorg stimulering. För detta ta bort stimulerande elektroderna från ögonen och ersätta dem på främre sidorna av bröstkorgen för att stimulera de motoriska nervceller med 10 tåg med 10 stimuli vid 100 Hz.
OBS: elektrofysiologi spår som visas i videon motsvarar inte effekterna av rent färgämne injektion.
5. Representativa resultat
Effekt av en antagonist på PSI för att DLM synaps av Giant Fiber System
Methyllycaconitine citrat (MLA) är en nAChR-antagonist som är specifik för a7 nAChR-subtyp. PSI till DLMn synaps i GF-DLM reaktionsvägen är beroende på Dα7 nAChR-subtypen för korrekt funktion, medan genetisk avlägsnande av Dα7 nAChR-subtypen har ingen effekt på GF-TTM reaktionsvägen 5,6. För att visa specificitet och sensitivitet för vår analys har vi injiceras MLA vid olika koncentrationer (0, 0,02, 0,04, 0.08, 0,12 ng / mg, 46 nl injiceras) i huvudet på djuret (n = 10 per förening behandling, n = 15 för saltlösning behandling). Endast manliga flugor (av vildtyp genotypen vild 10E) användes, och effekten av föreningen övervakades under totalt 15 minuter efter injektion.
Figur 5 visar skillnaden mellan baslinjen inspelningar som erhållits före injektion och de som erhållits efter injektion som svar på MLA och saltlösningskontroll lösning. Vi fann att injektion av MLA resulterade i oförmåga GF-DLM väg att följa en-till-en vid 100 Hz med stimuli av GF i hjärnan medan GF-TTM vägen förblev opåverkade. (Figur 5, Top och mitten spår, t-test utförs mellan saltlösningskontroller [0 ng / mg] och de olika koncentrationer av MLA vid varje tidpunkt såvida uppgifterna är icke-parametrisk [normalitet och lika varianser testade], annars använder vi en Mann-Whitney Rank Sum Test. * p <0,001). Emellertid en en-till-ett response av DLM observerades när de motoriska nervceller stimulerades direkt (Figur 5, botten spår), visar att NMJ funktion DLM och TTM inte påverkas av MLA. MLA tycktes nå sin maximala effekt 1 minut efter injektion för 0,04, 0,08 och 0,12 ng / mg MLA injiceras, eftersom inga ytterligare väsentliga förändringar observerades under de följande 15 minuterna av testperioden. Dessutom nådde föreningen en maximal effekt vid 0,08 ng / mg sedan starkare svar observerades inte med den högre dosen av 0,12 ng / mg.

Figur 1. The Giant Fiber System Diagram över Giant Fiber System (GFS). De Giant fibrer (GF, som visas i rött) synaps elektriskt (gap junctions) samt kemiskt (kolinerg) på en Peripheral Synapsing interneuron (PSI, som visas i grönt) och av Tergo trochanter Muscle neuron (TTMn, visas i gult) 5. PSI till DLMn (Dorsal längsgående muskeln neuron, som visas i blått) anslutning beror på Dα7 nAChR-subtypen 6. Slutligen neuromuskulära förbindelsen (NMJ) av TTMn och DLMn på hoppet (TTM, som visas i lila) och muskler flygning (DLM, som visas i lila) är glutamaterg.
Obs: GF för att PSI-anslutningen är både elektriska och kemiska. I shakB mutanter (som saknar kanalförbindelser), kan inget svar registreras från DLM vid stimulering av GF i hjärnan, vilket visar att den kemiska komponenten i frånvaro av elektriska anslutningar är inte tillräcklig för att framkalla en aktionspotential i PSI 5,16-18. Eftersom GF att PSI-anslutning är kanalförbindelsekonduktans beroende, visar denna figur endast gap junction vid synapsen för enkelhets skull.

Figur 2.
Mikromanipulatorer set-up.
- En modifierad inställning av en tidigare publicerad protokoll nr 14 används för att passa injektionen mikromanipulator för parade GFS inspelningar med samtidiga nanoinjections. Den monterade fluga preparatet är orienterad med huvudet på flugan mot försöksledaren. Injektionen mikromanipulator (# 1) är placerad framför den som utför experimentet mellan de två manipulatorer för volfram stimulerande elektroder (2 #-och # 3). De två mikromanipulatorer för elektroderna glas inspelning (# 4 och # 5) är placerade på vänster och höger sida, respektive. Den mikromanipulator för volfram jordelektroden (# 6) är placerad längst i ryggen, antingen på vänster sida (visas här) eller på höger sida.
- En nära bild upp från toppen av arrangemang av olika elektroder och injektion mikropipett.
- En korrekt monterad D. melanogaster spetsad med elektroder och injektion mikropipettspetsar klar för injektion. Lägg märke till att djurets kroppmonteras med sin bröstkorg horisontellt och vingarna utspridda. Vaxet är säkert lindas runt sin kropp att förhindra djuret från att röra sig. Dessutom är jordelektroden (# 6, i buken), de glas registreringselektroderna (# 4 och # 5 i bröstkorgen, lyfts fram av mörka konturer), och stimulerande elektroder (# 2 och # 3, en i varje öga ) är spetsad på plats, såsom tidigare beskrivits 14. Injektionen mikropipett (# 1) är korrekt inriktad med centrum av de tre ocelli (inringad). Insättning av injektion mikropipett bör placeras i detta område.

Figur 3. Avfasade injektion mikropipett. Ett diagram över en korrekt fasad mikropipett visas här. Elektroden öppning bör vara avfasade vid en 45 graders vinkel och har en öppning mellan 11 till 17 pm. En riktig avfasade injektion mikropipett är avgörande för en smidig injektion med minimal dAmage till flugan.

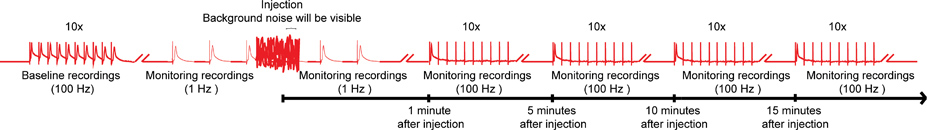
Figur 4. Övergripande syftet med nanoinjection / elektrofysiologi protokollet. En representant diagram över hela systemet för att nanoinjection / elektrofysiologi protokollet. Börja med att få ett utgångsvärde inspelning genom att stimulera Giant fibrer (GFS) vid 100 Hz med 10 tåg med 10 stimuli vardera (endast ett tåg visas här). Före injektion börjar de 1 Hz stimuli en sekunds mellanrum. Under injektion tiden (när injektor är ansluten till kontrollboxen), kommer du att följa betydande bakgrundsljud, men inte avbryta inspelningarna. Efter injektion (och injektor är utdragen från kontroll box), fortsätta 1 Hz stimulering för ca 1 minut till. Slutligen, fortsätt att betona GF med 10 tåg med 10 stimuli på 100 Hz och fortsätter att testa funktionen hos de GF vägar med denna paradigm var 5 minuter upp till 15 minuter. Obs: inspelningar were manipuleras för att skapa den övergripande planen och inte utgör ett specifikt resultat erhålls. Inte skala, är inte alla spår visas. Klicka här för större bild .

Figur 5. Effekterna av MLA i GFS.
- En grafisk avbildning av effekten av α7 nAChR-antagonisten Methyllycaconitine citrat (MLA) i GF-DLM reaktionsvägen av bananfluga vid olika koncentrationer (0, 0,02, 0,04, 0,08, 0,12 ng / mg n = 10 per förening behandling.; n = 15 för saltlösning behandling). Endast en minut efter injektion, var en signifikant och omedelbar effekt sågs med 0,04 ng / mg av MLA. En signifikant effekt observerades också med 0.8ng/mg och 0.12ng/mg av MLA vid 100 Hz stimulering av GF. Ingen signifikant skillnad noterades mellan saltlösningskontroller och 0,02 ng / mg MLA. Dessutom, ingen förändring in effekt sågs efter 1 minut efter injektionen under tiden testas (15 minuter). En t-test utfördes mellan saltlösningskontroller (0 ng / mg) och olika koncentrationer av MLA vid varje tidpunkt. Nivåer är rapporterade som medelvärde + / - SEM, * p <0,001.
- Exempel på spår av DLM svaren vid 100 Hz stimulering. Top spår visar svaren från muskeln innan MLA injektion vid GF stimulering i hjärnan. Lägg märke till att muskeln kan svara på varje stimulus ett-till-en vid 100 Hz. Mellanöstern spår visar svaren på DLM efter MLA injektion (0,12 ng / mg). Lägg märke till att muskeln inte kan svara på varje stimulus ett-till-en vid 100 Hz. (Asterisker). Bottenspår visar svaren från DLM av samma preparatet (0,12 ng / mg) vid direkt stimulering av motoriska nervceller i bröstkorgen. Eftersom DLM svarar en-till-en på 100 Hz med bröstkorg stimulering, kan misslyckandet av svaren med hjärnan stimuli hänföras till kolinerg PSI-DLMn anslutning.
- En grafisk avbildning av effekten med olika koncentrationer MLA (0, 0,02, 0,04, 0,08, 0,12 ng / mg) på GF-TTM reaktionsvägen. Inga signifikanta effekter sågs mellan saltlösning (0 ng / mg) och injektioner sammansatta vid någon tidpunkt. En t-test utfördes mellan saltlösningskontroller (0 ng / mg) och olika koncentrationer av MLA vid varje tidpunkt, * p <0,001.
- Exempel på spår av TTM svaren vid 100 Hz stimulering. Top spår visar svaren från muskeln innan MLA injektion vid GF aktivering med stimulering i hjärnan. Lägg märke till att muskeln kan svara på alla stimuli vid 100 Hz. Mellanöstern spår visar svaren från DLM efter MLA injektion (0,12 ng / mg). Svar från TTM muskeln att stimulering av GF i hjärnan förblir en-till-en. Bottenspår visar svaren från TTM i samma beredning till 100 Hz stimulering av motoriska nervceller i bröstkorgen (0,12 ng / mg).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Den nanoinjection / elektrofysiologi bioassay presenteras här möjliggör en snabb screening av föreningar i nervsystemet av bananfluga. Detta är en ny in vivo-teknik som kräver små mängder av en förening för att framkalla en effekt på en mängd olika molekylära mål i en väl karakteriserad neuronala krets. Denna metod kan användas för att testa bioaktiviteten hos olika föreningar, av okända toxiner med kommersiellt tillgängliga farmakologiska medel.
Här har vi visat att funktionen av vår analys med användning av MLA, som hade en effekt på Giant fibersystemet (GFS) av bananfluga (figur 5). Vi fann att det selektivt störde GF att DLM väg men inte GF till TTM väg. Aktivering av motoriska nervceller direkt via bröstkorg stimulering visat att det fel i GF för att DLM väg berodde inte på en dysfunktion vid den neuromuskulära förbindelsen (NMJ) men det var förenligt med den antagonistiska effektivtT av MLA vid Dα7 nAChR subtyper närvarande vid PSI-DLMn synaps (figur 1). Fastän GF att TTMn förbindelse visades vara kolinerga, är det okänt huruvida Dα7 nAChR subenheter är närvarande vid denna synaps. Vidare, den genetiska frånvaro av kolinacetyltransferas (Cha)-genen eller Dα7 nAChR-subtypen (Dα7)-genen inte stör funktionen hos GF-TTMn anslutningen på grund av den samtidiga närvaron av en elektrisk koppling 5,6,17,19, 20, vilket gör den reaktionsväg osannolikt att påverkas av MLA.
Efter förening injektion ska lösningen sänk genast hela nervsystemet hos djuret på grund av dess öppna cirkulationssystemet 21. Om rätt injiceras når föreningen vanligtvis bröstkorgen och buken på några sekunder, men en homogen dispersion kan ta upp till en minut. Om emellertid föreningen ej insprutas ordentligt i hemolymfa (dvs. att injicera mikropipett för deep gå in i hjärnvävnaden) sedan långsammare spridning över hela djuret observeras. Även färg kan användas för att utöva ett korrekt injektionsteknik som visas i videon, är det inte rekommenderat att samarbeta injicera den blå karamellfärg med en förening som ska testas eftersom det kan ändra egenskaperna för föreningen och därmed dess bioaktivitet. Dessutom, eftersom de flesta lösningar används som lösningsmedel är tydliga i färg (saltlösning, DMSO, etc), är det svårt att se huruvida föreningen ut från injektionsnålen. Därför, när upplösning av en specifik förening är det viktigt att säkerställa att den går helt i lösning, i annat oupplösta partiklar kommer snabbt sätta igen insprutningsspetsen, vilket förhindrar från någon utmatning av fluid. Även om föreningen spridning kan ske omedelbart hela hemolymfa, når målen i det centrala nervsystemet, samt når sitt maximum dosering, kan ta längre tid baserat på föreningens kemiska egenskaper, såsom storlek och Polarity, och dess förmåga att genomsyra flugans blod-hjärnbarriären. 22 Därför är det viktigt att övervaka potentiella effekter av okända föreningar flera minuter efter injektionen eftersom olika föreningar kan ha variation i tider av debut effekter som kan öka med tiden i vissa fall . Starka och omedelbara effekter av föreningen som helt blockera funktionen hos neuroner kan redan ses med de utlösta svaret vid 1 Hz, medan stimulering av GFS vid högre frekvenser (100 Hz) används för att detektera mer subtila effekter på grund av lägre dosering eller potens av en förening. Om inga effekter observeras efter förening injektion kan det bero på antingen små läkemedelsdos eller det faktum att föreningen specifika molekylära mål inte är närvarande i GFS.
Dessutom, vid användning av bioanalys som presenteras här som ett screeningsverktyg för nya föreningar (såsom konotoxiner) är det viktigt att notera att analysen är begränsad till de molekylära mål som finns iGFS av flugan. Även om analysen i sig inte tillåter att lokalisera de faktiska molekylära mål för den injicerade föreningen medger det för minska ner av potentiella mål inom GFS. Ytterligare tester, såsom patch clamp på neuroner eller muskler eller genetiska interaktionsstudier med Drosophila melanogaster mutanter, kan göras för att bestämma den specifika målet för dessa föreningar. Slutligen presenterades inspelningen protokollet för att upptäcka antagonistiska effekter på funktion GFS. Kan dock inspelningen protokollet kan enkelt justeras för att övervaka agonistiska effekter genom att passivt övervaka för svar induceras av föreningen i stället för att testa om GFS inte kan svara ett tillförlitligt sätt när kretsen stimuleras av försöksledaren.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Inga intressekonflikter deklareras.
Acknowledgments
Vi skulle vilja tacka ledamöterna i Mari lab och Godenschwege lab, i synnerhet Aline Yonezawa, för kommentarer och hjälp med detta protokoll. Detta arbete har finansierats av det nationella institutet för neurologiska sjukdomar och stroke bidrag R21NS06637 till FM och TAG, AB har finansierats av National Science Foundation Award antalet 082.925, URM: Integrativ biologi för framtida forskare.
Materials
| Name | Company | Catalog Number | Comments |
| Recording glass electrodes: borosilicate glass capillaries | World Precision Instruments, Inc. | 1B100F-4 | 1.0mm OD, 0.58mm ID |
| Stimulator | Grass Technologies | Model S48 | |
| Amplifier | Getting Instruments, Inc. | Model 5A | |
| Data acquisition Software: Digidata | Molecular Devices | Model 1440A | |
| Data collection software: pCLAMP | Molecular Devices | Version 10 | |
| Stereomicroscope with fiber optic microscope ring illuminator | AmScope | SM-4T Model HL250-AR | |
| Dissecting scope for mounting | AmScope | SM-2TZ | |
| Kite Manual Micromanipulator & Tilting Base | World Precision Instruments, Inc. | Model # M3301 Kite: Model # KITE-M3-L | |
| Drosophila melanogaster Wild 10E genotype (wild type strain) | Bloomington Stock center | Stock # 3892 | |
| Vertical pipette puller | David Kopf Instruments | Model 700c | |
| Injection glass micropipettes: Borosilicate glass capillaries | World Precision Instruments, Inc. | Catalogue # 4878 | 1.14mm OD, 0.5mm ID |
| Silicon oil | Fisher Scientific | Catalogue # S159-500 | |
| Beveler | Sutter Instrument Co. | K.T. Brown Type Model # BV-10 | |
| Nanoliter2000 | World Precision Instruments, Inc. | Catalogue # B203XVY | |
| Blue food coloring | McCormick & Co. | N/A | Ingredients: Water, Propylene Glycol, FD&C Blue 1, and 0.1% Propylparaben (preservative). |
| Methyllycaconitine citrate (MLA) | Tocris Bioscience | Catalogue # 1029 | |
| Plastic wax sticks | Hygenic Corporation (Akron Ohio USA) |
References
- Koehn, F. E., Carter, G. T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 4, 206-220 (2005).
- Miljanich, G. P. Ziconotide: neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 11, 3029-3040 (2004).
- Layer, R. T., Wagstaff, J. D., White, H. S. Conantokins: peptide antagonists of NMDA receptors. Curr. Med. Chem. 11, 3073-3084 (2004).
- Lewis, R. J. Conotoxins as selective inhibitors of neuronal ion channels, receptors and transporters. IUBMB Life. 56, 89-93 (2004).
- Allen, M. J., Godenschwege, T. A., Tanouye, M. A., Phelan, P. Making an escape: development and function of the Drosophila giant fibre system. Semin. Cell Dev. Biol. 17, 31-41 (2006).
- Fayyazuddin, A., Zaheer, M. A., Hiesinger, P. R., Bellen, H. J. The nicotinic acetylcholine receptor Dalpha7 is required for an escape behavior in Drosophila. PLoS biology. 4, e63 (2006).
- Jan, L. Y., Jan, Y. N. L-glutamate as an excitatory transmitter at the Drosophila larval neuromuscular junction. The Journal of physiology. 262, 215-236 (1976).
- Usherwood, P. N., Machili, P., Leaf, G. L-Glutamate at insect excitatory nerve-muscle synapses. Nature. 219, 1169-1172 (1968).
- Marrus, S. B., Portman, S. L., Allen, M. J., Moffat, K. G., DiAntonio, A. Differential localization of glutamate receptor subunits at the Drosophila neuromuscular junction. The Journal of neuroscience : the official journal of the Society for Neuroscience. 24, 1406-1415 (2004).
- Petersen, S. A., Fetter, R. D., Noordermeer, J. N., Goodman, C. S., DiAntonio, A. Genetic analysis of glutamate receptors in Drosophila reveals a retrograde signal regulating presynaptic transmitter release. Neuron. 19, 1237-1248 (1997).
- Qin, G. Four different subunits are essential for expressing the synaptic glutamate receptor at neuromuscular junctions of Drosophila. The Journal of neuroscience : the official journal of the Society for Neuroscience. 25, 3209-3218 (2005).
- Schuster, C. M. Molecular cloning of an invertebrate glutamate receptor subunit expressed in Drosophila muscle. Science. 254, 112-114 (1991).
- Tanouye, M. A., Wyman, R. J. Motor outputs of giant nerve fiber in Drosophila. Journal of. 44, 405-421 (1980).
- Augustin, H., Allen, M. J., Partridge, L. Electrophysiological Recordings from the Giant Fiber Pathway of D. melanogaster. J. Vis. Exp. (47), e2412 (2011).
- Allen, M. J., Godenschwege, T. Drosophila Neurobiology. Zhang, B., Freeman, M. R., Waddell, S. , Cold Spring Harbor Laboratory Press. 215-224 (2010).
- Blagburn, J. M., Alexopoulos, H., Davies, J. A., Bacon, J. P. Null mutation in shaking-B eliminates electrical, but not chemical, synapses in the Drosophila giant fiber system: a structural study. J. Comp. Neurol. 404, 449-458 (1999).
- Thomas, J. B., Wyman, R. J. Mutations altering synaptic connectivity between identified neurons in Drosophila. J. Neurosci. 4, 530-538 (1984).
- Baird, D. H., Schalet, A. P., Wyman, R. J. The Passover locus in Drosophila melanogaster: complex complementation and different effects on the giant fiber neural pathway. Genetics. 126, 1045-1059 (1990).
- Gorczyca, M., Hall, J. C. Identification of a cholinergic synapse in the giant fiber pathway of Drosophila using conditional mutations of acetylcholine synthesis. J. Neurogenet. 1, 289-313 (1984).
- Allen, M. J., Murphey, R. K. The chemical component of the mixed GF-TTMn synapse in Drosophila melanogaster uses acetylcholine as its neurotransmitter. The European journal of neuroscience. 26, 439-445 (2007).
- Mejia, M. A novel approach for in vivo screening of toxins using the Drosophila Giant Fiber circuit. Toxicon. 56, 1398-1407 (2010).
- Stork, T. Organization and function of the blood-brain barrier in Drosophila. J. Neurosci. 28, 587-597 (2008).

{kind=link}