

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
我们提出了一个经典的调节被利用的蚂蚁的协议, 允许研究人员研究视觉学习和记忆形成的水平的分析不可能与自由移动的个人。
Abstract
几种昆虫已成为研究学习记忆形成的模型系统。虽然许多研究集中在自由移动的动物, 研究实施经典调理范式与被利用的昆虫是重要的为调查个体学习的确切的提示和神经机制在学习之下和记忆形成。在这里, 我们提出了一个通过经典的条件唤起木材蚂蚁视觉联想学习的协议。在这个范式中, 蚂蚁被利用并呈现出一个视觉提示 (蓝色纸板), 条件刺激 (CS), 配对的食欲糖奖励, 无条件刺激 (美国)。蚂蚁执行上颌-唇伸展反射 (MaLER), 无条件的反应 (UR), 这可以用来作为学习的读数。训练包括10项试验, 用5分钟的 intertrial 间隔时间隔开。蚂蚁还可以在训练后10分钟或1小时内进行记忆保存测试。该协议有可能允许研究人员以精确和控制的方式分析视觉记忆形成的细节以及木蚁学习记忆形成的神经基础。
Introduction
昆虫被广泛地用作学习和记忆形成的模型1。一个特别成功的研究领域涉及使用受约束动物的经典调理, 它允许精确控制所学到的线索, 并允许研究人员研究学习和记忆支撑的神经机制。大多数研究都集中在食欲经典的蜜蜂工作者的调理蜜蜂。蜜蜂工人们被训练成将 CS 和一个唤起美国的人联系在一起。在这个范例中, 最初由武田2和比特曼等开发。3、UR 是喙延伸反射 (每), 美国是糖, CS 是一种气味。蜜蜂学会了 CS 和美国之间的联系, 并且可以形成这个协会的长期记忆。
最初的范式使用的是作为 UR 的, 被用来解开神经通路的基础上的嗅觉学习的蜜蜂4。它已被修改, 以多种方式来测试学习不同的感官刺激, 包括视觉刺激5,6,7, 以纳入厌恶学习使用抑制每8或刺延长反射 (系列)9作为 UR, 并且测试学习在其他种类, 例如熊蜂10和果蝇11。虽然通过经典的调节方法研究了几种模式的记忆形成, 但视觉学习仍然难以用这种方式观察, 即使是在觅食过程中表现出高度视觉学习能力的物种, 例如蜜蜂。
最近的研究应用了类似的方法, 昆虫没有喙, 如蝗虫, 执行下颚开放反应 (12) 和蚂蚁, 执行上颌骨-唇伸展反射 (MaLER)13。这已经揭示了相适应的学习能力, 符合两个不同的沙漠蝗虫表型, 群居和 solitarious 的阶段特定喂养策略, 重新实施记忆形成需要匹配的想法生态需要14。此外, 对蚂蚁嗅觉学习的研究表明, 蚂蚁和蜜蜂在记忆形成和保留方面有相似之处, 长期记忆保持, 训练后72小时, 依赖于蛋白质合成15。
这些原始范式的改编使得学习和记忆的形成可以用多种模式和几种模型来研究。这对于识别昆虫常见的记忆形成机制是至关重要的, 但也可以识别出不同物种的学习和记忆的特定生态学的差异。我们在这里描述的协议的主要目标是提供一种方法来执行经典的调理实验, 使用视觉条件刺激与广泛研究的蚂蚁物种,蚁。本文是从16《木蚁视觉学习》的研究中得到的, 也是视觉经典调理范式的一种改编。
Protocol
1. 维护蚁蚁群
注: 本研究使用的木蚁 (蚁, L.) 菌落是从英国苏塞克斯森林 (N 51 4.680, E 0 1.800) 收集的。在英国,蚁殖民地应该在7月和8月之间收集。必须清除大量的鸟巢, 包括数以百计的工人和育雏, 以确保该殖民地长期维持和活跃 (最多一年)。在拆巢前应征求有关当局的许可。
- 把木蚁群放在一个宽盒子里 (大约135厘米 x 70 厘米 x 35 厘米), 墙上覆盖着聚四氟乙烯 (PTFE) 树脂 (例如Fluon), 以防止蚂蚁在21°C 的 light:12-h 暗循环下逃逸。
- 用水分配器为蚂蚁提供蔗糖 (333 克/升) 和水。为了确保蚂蚁的饮食富含蛋白质, 每周用冰冻蟋蟀喂养菌落2x。每天用水喷洒巢以保持湿润。
- 改善菌落的卫生条件, 必要时加入松木树脂;松树脂减少寄生在木头蚂蚁殖民地17。在巢内放置一个小容器 (直径约15厘米和5厘米深), 这样蚂蚁就可以将任何死去的殖民地成员存放在里面。
- 在尝试任何实验之前, 在上述条件下保持蚂蚁至少2周。将蔗糖从巢中除去, 在实验前将蚂蚁饿死 2 d, 从而提高它们的动力。
2. 选择和利用蚂蚁
注: 对于利用蚂蚁, 需要一个定制的持有人。这可以建造使用建模粘土和纸板与切口开放附加到它水平。纸板的上部表面应部分涂上蜡, 允许在固定时附着昆虫别针。仔细处理对于本议定书的每一步 (包括维护、运输和实验) 都是必要的, 但特别是在利用蚂蚁时, 要避免在训练前使它们受到高强度的压力。
- 要选择有动机进食的蚂蚁, 在一个小盒子 (大约14厘米 x 8 厘米 x 5 厘米) 内放置一滴蔗糖 (200 克/升), 用 PTF 树脂覆盖的墙壁防止逃逸。不要选择携带木头或死蚂蚁的蚂蚁。取而代之的是, 把爬行在巢壁上的蚂蚁, 因为它们更可能被觅食试图离开巢穴寻找食物。
- 把每只蚂蚁放在盒子里, 等着看它是否在蔗糖滴下进食。如果有, 将其立即转移到另一个空框, 以防止饱足。
- 把每只蚂蚁转移到一个单独的管子上。把每个管子放在冰箱里 1-2 分钟, 或者在被粉碎的冰中放置5分钟, 以固定蚂蚁。
- 将固定的蚂蚁放在支架上, 通过纸板上的切口, 由头部和胸腔之间的接头。确保触角保持在头部一侧, 使用嵌入在纸板顶部蜡层的昆虫别针。
- 用特制的加热元件将昆虫别针的尖端蜡到蚂蚁的头上, 平行于纸板。在此过程中, 请勿用发热元件导线触摸天线, 以免对其造成任何损害。
- 蜡干后, 取出持有触角的昆虫别针, 并小心地将蚂蚁从持有者身上移走。
- 固定的昆虫针持有蚂蚁在一个建模粘土缸和固定在它下面的一个定制的塑料支架, 确保蚂蚁保持一个典型的站立姿势, 整个身体是自由移动, 除了头部。
- 在训练之前, 让被利用的蚂蚁在黑暗中保持至少2小时。
3. 培训和测试
-
设置, 条件刺激和无条件刺激
- 在一个白色的丙烯酸盒子 (50 x 50 x 50 厘米) 进行实验, 在前面打开, 允许实验者进入。用照相机记录蚂蚁的行为。
- 将相机与一个宏透镜直接放在蚂蚁上方, 穿过盒子上表面的一个孔。
- 为了减少任何无关的视觉暗示, 保持房间在黑暗中, 除了一个光源直接指向丙烯酸框顶部 (两个 26 W 灯)。
- 使用作为视觉提示 (CS) 一个明亮的蓝色 (图 S1) 纸板长方形 (60 x 45 毫米) 附有在它的中心到针, 连接到一个注射器 (2 毫升) 与美国 (蔗糖200克/升) 手动交付到蚂蚁 (图 1A和1B).
注: 蔗糖溶液可以与任何白糖, 只要它没有在水中溶解的颜色和气味。 - 以随机顺序进行平行、配对和不成对的两种训练 (见下文解释)。
注意: 在本研究中, 我们没有注意到视觉提示的性质。只有提示和奖励之间的关联才被考虑进去。颜色和形状未经过测试, 并且有其他功能的 CS 可能产生类似的结果。然而, 蓝色被采摘了, 因为来自同一属的蚂蚁已经被证明对覆盖蓝颜色18的波长敏感。
- 在一个白色的丙烯酸盒子 (50 x 50 x 50 厘米) 进行实验, 在前面打开, 允许实验者进入。用照相机记录蚂蚁的行为。
-
配对训练
注: 在配对训练中, CS 和美国在每一次试验中都呈现, 相互关联 (图 1C)。- 开始记录蚂蚁十年代之前提出 CS, 以确保蚂蚁没有自发地执行 MaLER 之前的演示。如果蚂蚁在这段时间内执行 MaLER, 将试验推迟几秒钟。如果任何 ant 连续显示此行为, 请将其从分析中排除。
- 移动注射器 + CS 在蚂蚁前面为 ~ 十年代, 与尖的针保持在蚂蚁的脸和最大的5毫米以上和每一侧。在这段时间内, 将针的尖端尽可能靠近蚂蚁的头部, 但不接触触角 (图 1B)。
注: 在呈现 CS 时, 运动被包括在内, 因为它已经被证明在蜜蜂6中发挥了视觉联想学习的作用。 - 施加压力, 从注射器尖端渗出一滴蔗糖, 然后将蔗糖滴放在触角和口器旁边, 让蚂蚁检测蔗糖。允许蚂蚁喂养大约5秒。
注: 在试验中, 每项试验中所消耗的蔗糖蚂蚁的数量没有受到控制。蚂蚁饲料中的蔗糖含量应该足够大, 可以让它们自由进食约5的食物。 - 重复此步骤 10x, 以5分钟的。
注意: 在这些实验中, 蚂蚁总是向右转, 右眼靠近丙烯酸盒子的开口边。因此, CS 总是接近蚂蚁从他们的右侧。虽然这并不会使学习失效, 但这些实验可以通过将蚂蚁的一半向左半向右转动来进行, 以避免任何可能的侧向效应。
-
未配对培训
注: 这项训练包括分别向蚂蚁介绍 CS 或美国, 这两种刺激, 因此, 在一段时间内彼此离解 (图 1D)。- 在每次试验前十年代开始记录蚂蚁。
- 以与配对训练相同的方式呈现 CS, 但不将蔗糖传递给蚂蚁。
- 2.5 分钟后, 将美国直接送到口器 (也接触触角) 使用注射器没有 CS 连接。
- 从 CS 试用开始, 并对每种类型的试验重复此过程 10x, 锂2.5 分钟。
-
测试
注: 在上一次训练试验后, 应只进行1x、10分钟或1小时的测试;一个测试, 其中蚂蚁是与 CS, 但不与美国可能导致的联想记忆的消亡以前形成。- 在测试前十年代开始记录蚂蚁。
- 向蚂蚁介绍十年代的 CS。
- 确保蚂蚁在试验后通过提供蔗糖来维持饲料的积极性。
4. 数据收集和分析
- 记录蚂蚁的行为, 从上面, 十年代之前, 每次试验, 并在 CS 和美国的发言。确保所有试验和试验记录后分析。
- 使用在训练和测试中制作的录音, 在十年代的 CS 演示文稿中, 对蚂蚁的反应进行评分。
- 将蚂蚁在 CS 演示过程中的反应分成三种类型的行为, 具体取决于口器的扩展和移动: 与运动 (有限元) 的完全扩展, 如喂养, 完全扩展没有运动 (FE) 或部分延伸 (PE) 的上颌骨-唇或上颌瓣 (图 2A - 2D)。对于分析, 将所有 MaLER 类型分组为单个响应 (图 2E)。
- 排除在每次试验和测试中没有进食的蚂蚁。
注: 为分析, 建议将单个蚂蚁考虑在内的统计测试, 从而对个体内的变异性进行核算。当在训练中以二进制方式对蚂蚁的反应进行分类 (1 为阳性反应, 0 为无反应) 时, 采用混合效应的逻辑回归是可取的16、19。为了比较每项训练试验或试验中的蚂蚁比例, 根据分析16、20的蚂蚁数量, 建议进行G检验或费舍尔的精确测试。
Representative Results
在经典的调理实验中, CS 不能诱导动物自发反应。在我们进行的实验中, 只有 3%-4% 的蚂蚁在训练前对第一次试验的视觉提示进行 MaLER。正在进行配对训练的蚂蚁对 CS 的反应越来越 MaLER (图 3A; 逻辑回归, N = 51, 自由度 (df) = 507, z = 5.949, p < 0.01)。从第三次试验开始, 对 CS 停滞不前反应的配对蚂蚁的比例为50% 左右。相反, 未配对的蚂蚁在训练过程中 MaLER 没有显著增加 (图 3B; 逻辑回归, N = 29, df = 287, z = 0.758, p = 0.45)。MaLER 在对视觉提示的反应中的发生显著高于配对训练 (逻辑回归, N = 80, df = 796, z =-5.306, p < 0.01), 这是真实的每一个试验, 除了第一 (表 1)。
为了检查他们的短期和中期记忆15, 蚂蚁测试了10分钟或1小时后, 最后一次训练试验。对于这两项测试, 在对 CS 进行 MaLER 反应时, 蚂蚁的比例明显高于配对训练 (图 3C和3D;表 1)。
在训练过程中, 个体蚂蚁显示了 MaLER 的数量和类型的巨大变化 (图 4A和4B)。从第二次或第三次试验开始的每项试验中, 只有14% 的配对蚂蚁反应, 而大多数蚂蚁在他们反应的试验和他们没有的实验之间交替。在蚂蚁反应的训练试验中, 他们延长和移动口器的程度各不相同。因此, 我们将 MaLER 分为三种不同类型: 有限元、FE 或 PE。通常情况下, 蚂蚁比铁更经常地执行有限元或 PE。然而, 只有少数蚂蚁的反应是一致的;在大多数情况下, 蚂蚁对他们所执行的 MaLER 的类型没有什么一致性 (图 4)。

图 1: 实验设置和培训方案.(A) 将蚂蚁与蜡固定在一个建模粘土圆筒 (橙色) 的昆虫别针上, 然后将一个特制的支架放在它下面, 以允许自然主义的姿态。把它放在一个由两个光源照亮的白色丙烯酸盒子里, 直接放在相机下面。使用作为条件刺激 (CS) 一个纸板 (蓝色正方形) 附着在注射器, 提供无条件刺激 (美国), 糖奖励。该插图显示了在持有人的蚂蚁的特写视图。(B) 注射器针的尖端被移动尽可能接近蚂蚁的头部, 但不接触的触角, 如本示意图所示。训练蚂蚁通过 a (C) 配对或 (D) 不成对训练。这个数字已经从费尔南德斯等修改。16.请点击这里查看这个数字的更大版本.

图 2: 木蚁的上颌骨唇伸展反应.录像中的个别帧显示蚂蚁在训练过程中的嘴巴部分运动。(A) 本小组不显示任何答复。(B) 本小组显示在舌终止的上颌骨唇的完全延伸 (FE)。(C) 此面板显示部分延伸 (PE), 只有上颌 palpus 可见。(D) 本小组显示上颌骨-唇结构的部分延伸 (PE)。(E) 接受配对训练的蚂蚁 (N = 51) 执行完全伸展运动 (有限元; 深褐色), 铁 (中褐色) 或 PE (浅褐色)。这个数字已经从费尔南德斯等修改。16.请点击这里查看这个数字的更大版本.

图 3: 木蚁形成联想记忆的视觉线索配对与糖奖励.(A) 对 MaLER 的配对蚂蚁 (N = 51) 的比例在培训过程中显著增加。(B) 在未配对训练中, 执行 MaLER 的蚂蚁的百分比没有显著增加 (N = 29)。蚂蚁被测试了 (C) 10 分钟 (配对: n = 15; 配对 : n = 15) 或 (D) 1 小时 (配对: n = 15; 对: n = 14) 在最后训练试验之后。在配对或未配对训练期间, 蚂蚁对 CS 反应的比例分别以褐色或灰色表示。三种类型的 MaLER 以暗 (有限元)、中 (FE) 和光 (PE) 音调表示。这个数字已经从费尔南德斯等修改。16.请点击这里查看这个数字的更大版本.

图 4: 蚂蚁在训练过程中的个体表现.这些面板显示了 (A) 配对蚂蚁和 (B) 不成对蚂蚁的个体表现。三种类型的 MaLER 代表在黑暗 (有限元), 中等 (FE), 和光 (PE) 棕色或灰色。这个数字已经从费尔南德斯等修改。16.请点击这里查看这个数字的更大版本.
| 试验 | N | Df | G (已调整) | P |
| 1 | 80 | 那 | 那 | > 0。1 |
| 2 | 80 | 1 | 3.86 | < 0.05 |
| 3 | 80 | 1 | 8.41 | < 0.01 |
| 4 | 80 | 1 | 6.63 | < 0.01 |
| 5 | 80 | 1 | 8.41 | < 0.01 |
| 6 | 80 | 1 | 7。5 | < 0.01 |
| 7 | 80 | 1 | 10.69 | < 0.01 |
| 8 | 80 | 1 | 11.76 | < 0.01 |
| 9 | 80 | 1 | 17.13 | < 0.01 |
| 10 | 80 | 1 | 17.13 | < 0.01 |
| 10分钟 | 59 | 1 | 5。5 | < 0.05 |
| 1小时 | 59 | 1 | 4.42 | < 0.05 |
表 1: 对每项试验和试验进行配对和未配对训练的蚂蚁之间 MaLER 反应的比较.显示蚂蚁的数量 (N), 自由度 (df), g-独立测试 (g) 和p值。第一次试验是用费舍尔的精确测试分析的。这张桌子已经从费尔南德斯等改了。16。
Discussion
经典的调理是研究学习和记忆的最完善的范例之一。我们在这里提出的协议是对蜜蜂工作者2,3和随后用于其他几个物种, 如熊蜂, 果蝇的模式的适应, 这也使用每作为学习的读数10 ,11, 蝗虫和蚂蚁, 使用的是 MaLER, 分别为12,13。使用这个协议, 可以训练被利用的木头蚂蚁学习视觉提示和糖奖励之间的联系并且分析这个短的 (10 分钟) 和中期 (1 小时) 记忆16的保留。
在任何行为实验中, 都有必要考虑到能够最大限度地减少动物反应变异的关键步骤。在这里提出的议定书中, 采取了若干措施, 以尽量减少培训前后的变化。在实验开始之前, 蚁群需要至少两天的时间, 蚂蚁应该根据他们愿意吃糖滴的意愿来选择。以这种方式选择蚂蚁, 是为了最大限度地培养有动力喂养蚂蚁的机会。细心的处理也是一个重要的考虑因素, 因为它可以帮助减少压力水平, 这会扰乱学习, 如果它是太激烈21。为此, 蚂蚁在被利用的时候应该被寒冷麻醉, 以保持静止不动, 因为在这个过程中任何运动 (为了逃跑) 可能是压力的源泉。此外, 蚂蚁和蜡之间的接触应该是最小的, 避免接触的触角和热蜡或电线, 这可能会造成损害。虽然这些观察没有被正式分析过, 但在学习过程中, 触角似乎以特定的模式移动。
在实验中, 小心地交付糖对保持蚂蚁的积极性也很重要。再次, 虽然这还没有正式分析, 突然的食物传递似乎给蚂蚁带来了额外的压力, 这反过来又导致缺乏动力和学习。此外, 在训练期间提供的蔗糖应减少浓度 (200 克/升), 以避免在训练和测试结束前的饱足。这使得 MaLER 成为一个优秀的候选者无条件的回应, 因为, 连同对视觉线索的这种反应的低自发性能, 它也不会饱和超过试验。最后, 相反到多数古典制约研究2,3,5,6,7,8,9,10,11,12,13, 我们训练了一只蚂蚁, 直到实验结束, 把它放在试验之间, 而不是把它移到测试另一只蚂蚁上。训练数只蚂蚁在一起似乎产生了更多的可变结果, 这可能是由于景观的完全变化引起的视觉信息的重音和/或冲突的增加。为了减少每项实验的持续时间, 我们使用了5分钟的模拟, 而不是10分钟的使用在大多数经典的调理研究16。尽管所有这些考虑都应该有助于确保蚂蚁在训练过程中有动力去喂养和学习, 但有些变异是不能避免的。我们建议使用似乎有正常的社会, 食欲和运动行为的蚂蚁, 并排除蚂蚁从分析的那一刻, 他们没有进食的训练试验或测试。
本研究未对 CS 的性质进行测试。虽然我们使用了一个蓝色的视觉刺激, 因为同一属的蚂蚁对这些波长18敏感, 其他颜色也可能被学习与奖励联系在一起。将需要进一步的实验, 以充分描述在这个设置中看到和学到的颜色。对于不同形状和大小的视觉提示, 这也是正确的。我们还没有测试蚂蚁的空间分辨率是否足以区分这里所呈现的视觉刺激与蚂蚁眼睛的距离。虽然木蚂蚁的复眼已经被描述的大小和数量的方面22, 据我们所知, 他们的空间分辨率尚未充分描述。然而, 这是为Melophorus magoti23计算的。类似于木材蚂蚁或其他被测昆虫的眼睛的描述将有助于对动物观察和学习的视觉线索的特征进行清楚的调查。此外, 我们在向蚂蚁展示视觉刺激时包括了运动, 因为它已经被证明在经典调理6中扮演蜜蜂联想学习的角色。然而, 这也没有在本研究中进行试验, 由于飞行昆虫与行走昆虫的运动性质不同, 蜜蜂和蚂蚁视觉经典调理的区别可以观察到。
最后一点, 我们无法检查长期记忆保留, 因为蚂蚁在训练后无法存活很长时间。然而, 在随后的一系列实验中, 我们让蚂蚁活着, 并在被利用的时候有动力去吃和学习, 并在黑暗和潮湿的环境中过夜 (把盒子放在上面)。因此, 这个范式可以用来解开木材蚂蚁的长期记忆保留, 除了短期和中期记忆。
借助于一般经典调理范式的这一简单程序, 可以研究在利用自由运动动物的范式中广泛研究的视觉记忆的获取和保留。这种范式有可能被用来分析视觉学习的神经基础, 在一个非常完善的昆虫导航模型。
Disclosures
作者没有什么可透露的。
Acknowledgments
作者感谢汤姆科雷特和科妮莉亚 Buehlmann 分享关于收集和维护木蚁殖民地的宝贵信息。作者还感谢贾斯汀 Crevel 对本文先前版本的评论。
Materials
| Name | Company | Catalog Number | Comments |
| Fluon | Blades Biological Ltd, Edenbridge, UK | ACS 109; ACS 112; ACS 114 | For preventing insects from scaping |
| Crickets | Blades Biological Ltd, Edenbridge, UK | LZJ 217 | Given to the ant colonies as protein source |
| Natural Pine Rosin/Resin | Minerals-water Ltd, Rainham, UK | 500g | Given to the ant colonies for sanitation |
| Austerlitz Insect Pin | Fine Science Tools GmbH, Heidelberg, Germany | 26000-40 | For harnessing ants |
| High speed camera | Edmund Optics Inc., Barrington, USA | eo-13122M | MaLER recordings during training and testing |
| Macrolens | Cannon, Surrey, UK | EF 100 mm f/2.8 L Macro IS USM | MaLER recordings during training and testing |
| Software | IDS Imaging Development Systems GmbH | uEye64 | MaLER recordings during training and testing |
| Blue Cardboard | john smith's at Union Store, University of Sussex | JACK-PJM41358 | Constitutes de conditional stimulus |
| Syringe | Fisher Scientific LTD, Loughborough, UK | BD Plastipak 300185case; Product Code.12369289 | US and CS (attached) presentation |
| Needle (0.5 x 16 mm) | Fisher Scientific LTD, Loughborough, UK | BD Microlance 300600; Product Code:10442204 | US and CS (attached) presentation |
References
- Chittka, L., Niven, J.
- Takeda, K. Classical conditioned response in the honey bee. Journal of Insect Physiology. 6, 168-179 (1961).
- Bitterman, M. E., Menzel, R., Fietz, A., Schäfer, S. Classical conditioning of proboscis extension in honeybees (Apis mellifera). Journal of Comparative Psychology. 97 (2), 107-119 (1983).
- Menzel, R., Muller, U. Learning and memory in honeybees: from behavior to neural substrates. Annual Review of Neuroscience. 19 (1), 379-404 (1996).
- Hori, S., Takeuchi, H., Kubo, T. Associative learning and discrimination of motion cues in the harnessed honeybee Apis mellifera L. Journal of Comparative Physiology A. 193 (8), 825-833 (2007).
- Balamurali, G. S., Somanathan, H., De Ibarra, N. H. Motion cues improve the performance of harnessed bees in a colour learning task. Journal of Comparative Physiology A. 201 (5), 505-511 (2015).
- Mujagić, S., Würth, S. M., Hellbach, S., Dürr, V. Tactile conditioning and movement analysis of antennal sampling strategies in honey bees (Apis mellifera L). Journal of Visualized Experiments. (70), e50179 (2012).
- Wright, G., et al. Parallel reinforcement pathways for conditioned food aversions in the honeybee. Current Biology. 20 (24), 2234-2240 (2010).
- Vergoz, V., Roussel, E., Sandoz, J. C., Giurfa, M. Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex. PloS One. 2 (3), e288 (2007).
- Laloi, D., et al. Olfactory conditioning of the proboscis extension in bumble bees. Entomologia Experimentalis et Applicata. 90 (2), 123-129 (1999).
- Chabaud, M. A., Devaud, J. M., Pham-Delègue, M. H., Preat, T., Kaiser, L. Olfactory conditioning of proboscis activity in Drosophila melanogaster. Journal of Comparative Physiology A. 192 (12), 1335-1348 (2006).
- Simões, P., Ott, S. R., Niven, J. E. Associative olfactory learning in the desert locust, Schistocerca gregaria. Journal of Experimental Biology. 214 (15), 2495-2503 (2011).
- Guerrieri, F. J., d'Ettorre, P. Associative learning in ants: conditioning of the maxilla-labium extension response in Camponotus aethiops. Journal of Insect Physiology. 56 (1), 88-92 (2010).
- Simões, P. M., Niven, J. E., Ott, S. R. Phenotypic transformation affects associative learning in the desert locust. Current Biology. 23 (23), 2407-2412 (2013).
- Guerrieri, F. J., d'Ettorre, P., Devaud, J. M., Giurfa, M. Long-term olfactory memories are stabilised via protein synthesis in Camponotus fellah ants. Journal of Experimental Biology. 214 (19), 3300-3304 (2011).
- Fernandes, A. S. D., Buckley, C. L., Niven, J. E. Visual associative learning in wood ants. Journal of Experimental Biology. 221 (3), 173260 (2018).
- Christe, P., Oppliger, A., Bancala, F., Castella, G., Chapuisat, M. Evidence for collective medication in ants. Ecology Letters. 6 (1), 19-22 (2003).
- Aksoy, V., Camlitepe, Y. Behavioural analysis of chromatic and achromatic vision in the ant Formica cunicularia (Hymenoptera: Formicidae). Vision research. 67, 28-36 (2012).
- Bates, D. M. lme4: Mixed-Effects Modeling with R. , Springer. New York, NY. (2010).
- Sokal, R. R., Rohlf, F. J. Biometry (3rd edn). , W.H. Freman and Company. New York, NY. (1995).
- Bateson, M., Desire, S., Gartside, S. E., Wright, G. A. Agitated honeybees exhibit pessimistic cognitive biases. Current Biology. 21 (12), 1070-1073 (2011).
- Perl, C. D., Niven, J. E. Differential scaling within an insect compound eye. Biology letters. 12 (3), 20160042 (2016).
- Schwarz, S., Narendra, A., Zeil, J. The properties of the visual system in the Australian desert ant Melophorus bagoti. Arthropod Structure & Development. 40 (2), 128-134 (2011).
Tags
行为 问题 140 视觉提示 食欲调理 学习 记忆 MaLER,蚁Erratum
Formal Correction: Erratum: Visual Classical Conditioning in Wood Ants
Posted by JoVE Editors on 12/31/1969.
Citeable Link.
An erratum was issued for: Visual Classical Conditioning in Wood Ants. The Acknowledgments section was updated, and a supplemental figure was added.
The Acknowledgments sections was updated from:
The authors thank Tom Collett and Cornelia Buehlmann for sharing valuable information regarding collection and maintenance of wood ant colonies. The authors also thank Justine Crevel for commenting on previous versions of this article.
to:
The authors thank Tom Collett and Cornelia Buehlmann for sharing information regarding collection and maintenance of wood ant colonies. The authors also thank Justine Crevel for commenting on previous versions of this article, and Nora Nevala for measuring the spectrum intensity of the visual stimulus. This work was supported by a BBSRC grant to JEN (grant number BB/R005036/1). All the data pertaining to this manuscript are published in the University of Sussex Research Data Repository online database (10.25377/sussex.5794386).
The following supplemental figure was added to the end of the Representative Results section:
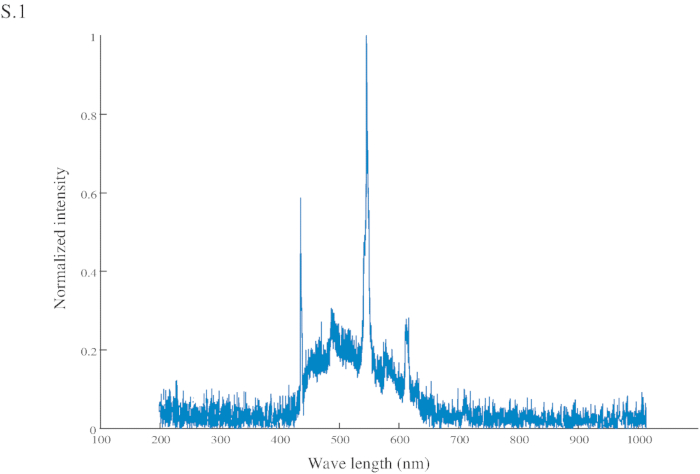
Figure S1: Normalized intensity of the conditional stimuli (CS). The CS has a peak intensity at 545 nm (in the green range) and another at 435 nm (in the blue range).
