

Summary
يصف هذا البروتوكول طريقة غير جراحية لتحديد خلايا المرحلة S بكفاءة لدراسات المجهر المصب، مثل قياس توظيف بروتين إصلاح الحمض النووي عن طريق الإشعاع الدقيق بالليزر.
Abstract
إصلاح تلف الحمض النووي يحافظ على السلامة الوراثية للخلايا في بيئة تفاعلية للغاية. قد تتراكم الخلايا أنواع مختلفة من تلف الحمض النووي بسبب كل من المصادر الذاتية والخارجية مثل الأنشطة الأيضية أو الأشعة فوق البنفسجية. وبدون إصلاح الحمض النووي، يصبح الشفرة الوراثية للخلية عرضة للخطر، مما يقوض هياكل ووظائف البروتينات ويحتمل أن يسبب المرض.
إن فهم الديناميكيات الصدغية للمسارات المختلفة لإصلاح الحمض النووي في مراحل دورة الخلية المختلفة أمر بالغ الأهمية في مجال إصلاح تلف الحمض النووي. توفر تقنيات المجهر الفلورية الحالية أدوات رائعة لقياس حركية التوظيف لبروتينات الإصلاح المختلفة بعد تحريض تلف الحمض النووي. تخليق الحمض النووي خلال مرحلة S من دورة الخلية هو نقطة غريبة في مصير الخلية فيما يتعلق بإصلاح الحمض النووي. وهو يوفر نافذة فريدة لفحص الجينوم بأكمله للأخطاء. وفي الوقت نفسه، تشكل أخطاء تخليق الحمض النووي أيضا تهديدا لسلامة الحمض النووي لا يصادف في الخلايا غير المقسمة. لذلك، تختلف عمليات إصلاح الحمض النووي بشكل كبير في مرحلة S بالمقارنة مع المراحل الأخرى من دورة الخلية، وهذه الاختلافات غير مفهومة بشكل جيد.
يصف البروتوكول التالي إعداد خطوط الخلايا وقياس ديناميكيات بروتينات إصلاح الحمض النووي في مرحلة S في مواقع تلف الحمض النووي المستحثة محليا ، باستخدام مجهر كونفوجكال مسح بالليزر مجهز بخط ليزر 405 نانومتر. يستخدم PCNA الموسومة (مع mPlum) كعلامة دورة الخلية جنبا إلى جنب مع بروتين إصلاح AcGFP المسمى من الفائدة (أي EXO1b) لقياس تجنيد تلف الحمض النووي في مرحلة S.
Introduction
وقد تطورت العديد من مسارات إصلاح الحمض النووي لمعالجة الأنواع المختلفة من آفات الحمض النووي التي يمكن أن تنشأ في الخلايا، وكلها منظمة تنظيما عاليا في كل من المكان والزمان. واحدة من الفترات الأكثر عرضة للخطر من دورة الخلية هي مرحلة S، عندما يحدث تخليق الحمض النووي. وفي حين أن الانتشار أساسي للحياة، فإنه يشكل أيضا تحديا كبيرا. تحتاج الخلايا إلى ضمان تكرار المؤمنين لجينومها لتجنب الطفرات التي تنتقل إلى الأجيال القادمة. وبالتالي، يوفر الانتشار نقطة تدخل علاجية تم استخدامها لتطوير النهج العلاجية في مجال الأورام.
جميع التقنيات الرئيسية المستخدمة لدراسة تجنيد البروتين في آفات الحمض النووي لها نقاط قوتها وحدودها. التشعيع الدقيق له دقة مكانية وزمنية أفضل1 من معظم الطرق البديلة مثل التصوير المناعي الفلوري لل بؤر الإشعاع المؤينة (IRIF) أو الاختناق المناعي الكروماتين (ChIP) أو الكسر الكيميائي الحيوي. ومع ذلك، فإن الإشعاع الدقيق يلتصق بقوة التقنيات المذكورة أعلاه التي يمكن أن تتذوق عددا كبيرا من الخلايا في نفس الوقت.
للتحقيق في إصلاح الحمض النووي في مرحلة S، يجب أن يكون المرء قادرا على تمييز خلايا المرحلة S في مجموعة غير متزامنة من زراعة الخلايا. هناك العديد من الطرق المعروفة لمعالجة هذا الأمر، والتي تنطوي إما على مزامنة الخلايا، أو التصور لمراحل دورة الخلية المختلفة. ومع ذلك، فإن كلا النهجين يفرضان تحديات كبيرة ومصنوعات يدوية محتملة. أساليب التزامن الكيميائية المستخدمة على نطاق واسع لإثراء الخلايا في مرحلة S المبكرة (على سبيل المثال، كتلة الثيميدين المزدوجة، والأفيديكولين، وعلاج الهيدروكسي يوريا) تحقق المزامنة من خلال تحريض إجهاد النسخ المتماثل وتلف الحمض النووي نفسه في نهاية المطاف. وهذا يحد من استخدام هذه الأساليب لدراسة عمليات إصلاح الحمض النووي في المرحلة S2. التزامن من خلال المجاعة في الدم والإفراج ينطبق فقط على عدد محدود من خطوط الخلية، باستثناء خطوط الخلايا السرطانية إلى حد كبير التي تعتمد أقل على عوامل النمو لتطور دورة الخلية مقارنة مع خطوط الخلايا غير المحولة. نظام مؤشر دورة الخلية الفلورية يوبيكيتين (FUCCI) هو أداة مفيدة بشكل خاص لدراسة دورة الخلية، ولكن لديه قيد أساسي عند التفريق بين مراحل دورة الخلية S و G23.
هنا يظهر أن استخدام PCNA الموسومة فلوريا كعلامة غير الغازية للمرحلة S يحد من عيوب أساليب مزامنة دورة الخلايا الكيميائية، مع السماح لمزيد من التحديد والمرونة من نظام FUCCI. كعلامة واحدة، لا يمكن فقط PCNA تسليط الضوء على خلايا المرحلة S في السكان غير متزامن، ولكن يمكن أيضا أن تظهر التقدم الدقيق للخلايا داخل مرحلة S (أي، في وقت مبكر، منتصف، أو أواخر مرحلة S)4. انخفاض مستويات التعبير من PCNA الخارجية، الموسومة يضمن الحد الأدنى من التدخل مع كل من تطور دورة الخلية وعمليات إصلاح الحمض النووي. الأهم من ذلك، PCNA أيضا بمثابة مراقبة داخلية لتحريض تلف الحمض النووي السليم كما أنها تشارك في إصلاح العديد من آفات الحمض النووي ويتم تجنيده لمواقع تلف الحمض النووي المستحثة محليا1،4.
التجارب المعروضة هنا تبين كيفية قياس ديناميات التوظيف من EXO1b في مرحلة S وكيف يتأثر هذا من قبل مثبط PARP راسخة، أولاباريب. نشاط النيوكليز EXO1b هو ذات الصلة لمجموعة واسعة من مسارات إصلاح الحمض النووي بما في ذلك إصلاح عدم التطابق (MMR)، إصلاح استئصال النيوكليوتيدات (NER)، وكسر مزدوجة تقطعت بهم السبل (DSB) إصلاح. في مرحلة S، EXO1b يلعب دورا رئيسيا في إعادة التركيب متجانسة (الموارد البشرية) من خلال تشكيل 3 'ssDNA يتدلى خلال استئصال الحمض النووي5. وقد تورط EXO1b كذلك في تكرار الحمض النووي مع أدوار في تنشيط نقطة تفتيش لإعادة تشغيل شوك الحمض النووي المتوقفة وكذلك إزالة التمهيدي ونضوج جزء أوكازاكي في حبلا متخلفة خلال تشريد حبلا في النسخ المتماثل5. يتم تنظيم التوظيف EXO1b لمواقع الحمض النووي التالفة من خلال التفاعل المباشر مع بولي (ADP-الريبوز) (PAR)6،7. نظرا للعديد من الآثار المحددة لدورة الخلية من EXO1b ، فهو خيار ممتاز لدراسات التوظيف الخاصة ب S-phase باستخدام PCNA.
Protocol
1. زراعة الخلايا المشتقة من هشاشة العظام البشرية (U-2 OS)
ملاحظة: تعتبر خلايا نظام التشغيل U-2 مثالية لهذه الدراسات حيث أن لديها مورفولوجيا مسطحة ونواة كبيرة وتعلق بقوة على العديد من الأسطح ، بما في ذلك الزجاج. ويمكن أيضا استخدام خطوط خلايا أخرى ذات خصائص مماثلة.
- لزراعة خطوط خلايا نظام التشغيل U-2، استخدم وسيط McCoy 5A المكمل بمصل بقري جنيني بنسبة 10٪ (FBS) والمضادات الحيوية (100 U/mL penicillin و 100 ميكروغرام/مل ستريبتومايسين). احتضان الخلايا عند 37 درجة مئوية في جو رطب يحتوي على 5٪ CO2. للدراسات المجهرية، والحفاظ على ثقافة الخلية في طبق 10 سم لتوفير عدد كاف من الخلايا.
- عندما تقترب الخلايا من التقاء 90٪ (~ 7 × 106 خلايا / 10 سم طبق)، وتقسيم الخلايا.
- شطف الخلايا مع برنامج تلفزيوني لغسل مثبطات التربسين الواردة داخل المصل.
- إضافة 1 مل من تريبسين-EDTA وتأكد من أن طبقة الخلية مغطاة بالتساوي.
- حضانة في 37 درجة مئوية حتى يتم رفع طبقة الخلية قبالة لوحة (حوالي 6 دقائق).
- Resuspend الخلايا جربسينيد في مصل يحتوي على وسائل الإعلام ل inactivate تريبسينوإضافة 1/10 th من حجم (~ 0.7 × 106 خلايا) في لوحة جديدة 10 سم تحتوي على 10 مل من المتوسطة النمو المكمل.
- قبل التجريب، اختبار الخلايا بشكل روتيني للتلوث الميكوبلازما باستخدام عدة الكشف عن الميكوبلازما العالمية بعد توصية الشركة المصنعة.
2. العدوى الرجعية
ملاحظة: بالنسبة لتدابير السلامة BSL-2 وأثناء العمل مع الفيروسات المؤتلفة، يرجى الرجوع إلى: إرشادات المعاهد القومية للصحة، القسم III-D-3: الفيروسات المؤتلفة في زراعة الأنسجة.
- البذور 4 × 106 خلايا HEK293T لتحقيق ~ 60٪ التقاء في غضون 24 ساعة بعد الطلاء في طبق ثقافة 10 سم.
- لزراعة HEK293T يرجى اتباع خطوات زراعة نظام التشغيل U-2 الموصوفة في 1.1-1.3 من هذا البروتوكول. ل HEK293T بديل مكوي 5A المتوسطة لDMEM. تأكد من غسل خلايا HEK293T بلطف دائما لأنها تعلق على لوحات زراعة الأنسجة بشكل ضعيف.
- خلايا ترانسفيكت HEK293T باستخدام كاشف العدوى القائم على الدهون للتغليف الفيروسي من البلازميدات.
- بالنسبة لنواقل الفيروسات الرجعية، اجمع 1.5 ميكروغرام من VSV-G (Addgene #8454) و1.5 ميكروغرام من pUMVC (أدجين # 8449) متجه التعبئة والتغليف جنبا إلى جنب مع 3 ميكروغرام من المتجه الذي يحتوي على جين الفائدة (في العمود الفقري ناقلات الرجعية مع مقاومة بوروميسين) إلى 250 ميكرولتر من وسائل الإعلام أوبتي-MEM خفضت مصل الدم في أنبوب الطرد الدقيق. أضف 1 ميكرولتر من كاشف P3000 لكل ميكروغرام من الحمض النووي المضاف إلى خليط Opti-MEM/DNA (في هذه الحالة 6 ميكرولتر) واخلط بلطف عن طريق النقر. لا دوامة أو ماصة صعودا وهبوطا.
- في أنبوب طرد مركزي آخر، اجمع 2 ميكرولتر لكل ميكروجرام من الحمض النووي (في هذه الحالة 12 ميكرولتر) من كاشف العدوى مع 250 ميكرولتر من وسائط مصل Opti-MEM المخفضة.
- الجمع بين الخلائط اثنين (500 ميكرولتر مجتمعة، لا دوامة، مزيج فقط عن طريق التنصت لطيف) والسماح لها احتضان لمدة 15 دقيقة في درجة حرارة الغرفة.
- بعناية، إضافة الخليط دروبيا إلى خلايا HEK293T المصنف دون فصل الخلايا. دوامة لوحات بلطف.
- العدوى الفيروسية لتوليد خطوط الخلايا مستقرة.
- إزالة الفيروس الذي يحتوي على supernatant من خلايا HEK293T 72 ساعة بعد transfection. قم بتصفية الحل بعناية باستخدام فلتر 0.45 ميكرومتر لإزالة حطام الخلية والخلايا المنفصلة. اختياريا، إضافة 8 ميكروغرام / مل البوليبرين إلى الناسخ الفيروسي لتسهيل العدوى الفيروسية.
- إضافة الفيروسات التي تحتوي على supernatant إلى خلايا نظام التشغيل U-2 في ~ 50٪ التقاء في طبق 10 سم (~ 3 × 106 خلايا). البذور خلايا نظام التشغيل U-2 في اليوم السابق.
- تصيب لمدة 6-16 ساعة قبل إزالة والتخلص من الفيروس المحتوي على supernatant.
ملاحظة: لتحقيق المبلغ المطلوب من التعبير المفرط لجين الفائدة، واحتضان سلسلة من التخفيفات الفيروسية لفترة محددة من الزمن. تحقق من مستويات التعبير من المتحولين جنسيا في كل خط الخلية المنشأة حديثا مع وصمة عار الغربية مقارنتها بمستويات الذاتية. - السماح للخلايا لتحديد في وجود المضادات الحيوية المناسبة (لمدة 3-4 أيام في حالة بوروميسين في التركيز النهائي 2 ميكروغرام / مل) والتحقق من التعبير عن البروتين الفلوري الموسومة الجين من الفائدة تحت المجهر.
- كرر هذه الخطوات لإنشاء خطوط خلايا مزدوجة المسماة. في التجارب المعروضة هنا تم التعبير عن mPlum-PCNA من ناقل للفيروسات الرجعية (pBABE) جنبا إلى جنب مع EXO1B-AcGFP، وأعرب أيضا من ناقل الفيروسات الرجعية (pRetroQ-AcGFP1-N1).
3. إعداد خلايا للإشعاع الدقيق
- طلاء الخلايا: 24 ساعة قبل التجربة، لوحة ما مجموعه 8.0 × 104 خلايا في حجم بين 500 μL-1 مل من وسائل الإعلام (لالتقاء ما يقرب من 70٪) على أربعة coverglass غرف جيدا مع رقم 1.5 البوروسيليكات الزجاج القاع الذي يقدم نتائج مثالية للمجهر confocal عالية التكبير والليزر التشعيع الدقيق. يسمح التقاء الخلايا الأعلى بالمزيد من الخلايا التي تقاس في مجال رؤية واحد (FOV)؛ ومع ذلك سوف الشرائح التقاء تماما إدخال المخالفات دورة الخلية.
- وسائط التصوير: قبل ساعة من التشعيع الدقيق، تبادل متوسط النمو المنتظم لفلوروبريت DMEM تكملها 10٪ FBS، 100 U/mL البنسلين و 100 ميكروغرام / مل ستربتومايسين، 15 MM HEPES (pH = 7.4) و 1 mM الصوديوم بيروفاتي. تساعد وسائط التصوير هذه على زيادة نسبة الإشارة إلى الضوضاء إلى أقصى حد مما يسمح بالكشف عن الفلورسينس الخافت للغاية. وبما أنه يحتوي على HEPES، فإنه يعمل أيضا على استقرار الحموضة في غياب جو CO2 بنسبة 5٪.
- تطبيق أي علاج إضافي قبل التصوير في هذه الخطوة. في التجارب المعروضة هنا، تم معالجة الخلايا قبل ساعة واحدة من التصوير إما مع أولاباريب (مثبط PARP، بتركيز نهائي 1 ميكرومتر) أو التحكم في السيارة (DMSO)1،8،9.
4. إعداد المجهر واختيار خلايا المرحلة S للتصوير.
- استخدم نظام confocal له خصائص مشابهة للنظام الموضح هنا للحصول على أفضل النتائج. أجريت التجارب المعروضة هنا باستخدام مجهر كونفوجكال مثبت على حامل مجهر مقلوب (انظر جدول المواد).
ملاحظة: تم تجهيز المجهر المستخدم هنا مع وحدة ليزر 50 mW 405 نانومتر FRAP، و60x 1.4 NA النفط خطة-apochromat الهدف. كان لدى رأس المسح الضوئي confocal خياران للماسح الضوئي: ماسح ضوئي غالفانو (للحصول على دقة عالية) وماسح ضوئي رنان (للتصوير عالي السرعة).- أدخل استرداد الفلورية بعد الليزر الضوئي (FRAP) للعينة عبر جهاز XY galvano الذي يتم التحكم فيه من خلال برنامج. استخدام خط ليزر 488 نانومتر لإثارة AcGFP و561 نانومتر أو 594 نانومتر خط ليزر لإثارة mPlum.
ملاحظة: تعطي تركيبة الفلتر التالية نتائج مثالية: باستخدام فلتر تمرير طويل 560 نانومتر، تم تمرير ضوء الانبعاثات مع طول موجي أقل من 560 نانومتر من خلال فلتر انبعاثات 525/50 نانومتر ل AcGFP، في حين تم تمرير ضوء الانبعاثات مع طول موجي أعلى من 560 نانومتر من خلال مرشح انبعاثات 595/50 نانومتر لmPlum. أي مجموعة مرشح مناسبة (على سبيل المثال، FITC/TRITC، GFP/mCherry، FITC/TxRed) تضمن الحد الأدنى من النزيف الفلوري.
- أدخل استرداد الفلورية بعد الليزر الضوئي (FRAP) للعينة عبر جهاز XY galvano الذي يتم التحكم فيه من خلال برنامج. استخدام خط ليزر 488 نانومتر لإثارة AcGFP و561 نانومتر أو 594 نانومتر خط ليزر لإثارة mPlum.
- تشغيل الغرفة البيئية ومكونات المجهر.
- قم بتشغيل التدفئة (المرحلة والموضوعية والغرفة البيئية عندما يكون ذلك ممكنا)وإمدادات ثاني أكسيد الكربون ومنظم الرطوبة على الأقل 4 ساعة قبل بدء التجربة لضمان التوازن الحراري للحصول على صورة مستقرة.
- تهيئة مصادر الضوء جنبا إلى جنب مع خطوط الليزر على الأقل 1 ساعة قبل نقل الخلايا إلى المجهر.
- حدد خلايا S-phase في مجموعة سكانية غير متزامنة باستخدام PCNA الموسومة بالفلورسنت كعلامة. قم بذلك باتباع الخطوات التالية.
- ابحث عن نمط التعريب الفريد ل PCNA الموسوم ب mPlum في مرحلة S مما يجعل تحديد مرحلة دورة الخلية هذه ممكنا. PCNA لديه توزيع متجانسة تماما في النواة في G1 و G2 مراحل دورة الخلية، في حين يجري استبعادها من النوى. في مرحلة S، PCNA يشكل بؤر في موقع replisomes في النواة. ويبين الشكل 1 الأنماط المختلفة لم بؤر PCNA طوال مرحلة S، مما يجعل من الممكن التمييز حتى في وقت مبكر، منتصف، وأواخر مرحلة S.
- ننظر من خلال العين لتحديد FOV التي لديها ما يكفي من الخلايا المرحلة S للإشعاع الدقيق. خلايا OS U-2 غير المتزامنة عادة ما يكون 30-40٪ من سكانها في مرحلة S.
- في محاولة لتجنب التطرف في مستويات التعبير (الخلايا الساطعة والخافتة على حد سواء) لكل من PCNA وبروتين الاهتمام (POI)، في هذه الحالة EXO1b-AcGFP، والتي يمكن أن تؤدي إلى التحف التجريبية.
- عند العثور على FOV مناسبة، في محاولة لتجنب مسح الحقل لفترة طويلة للحد من photobleaching وتلف الحمض النووي غير المرغوب فيها.
- تعيين منطقة الاهتمام المطلوبة (ROI) للتشعيع الجزئي. باستخدام البرامج المرتبطة (راجع جدول المواد)،قم بتعيين عائد الاستثمار المطلوب عن طريق إدراج الخطوط الثنائية أولا (تعيين العدد المطلوب من الأسطر والتباعد). انقر فوق ثنائي، ثم انقر فوق إدراج سطر | دائرة | القطع الناقص لرسم العدد المطلوب من الخطوط.
- تحويل هذه الخطوط الثنائية إلى ROIs وتحويل هذه ROIs أخيرا إلى ROIs التحفيز. للقيام بذلك، انقر أولا على ROI، ثم انقر فوق نقل ثنائي إلى ROI، ثم انقر بزر الماوس الأيمن على أي من ROIs وحدد استخدام ك ROI التحفيز: S1. ضع هذه الخطوط في FOV لتمرير من خلال نواة الخلايا. ROIs بطول 1024 بكسل التي امتدت FOV بأكملها استخدمت في جميع أنحاء البروتوكول.
5. التشعيع الدقيق لتلطيخ immunofluorescence أو التصوير الفاصل الزمني.
- تحديد إعدادات الإشعاع الدقيق الأمثل.
- قبل التشعيع الدقيق للخلايا ، خذ صورة أعلى دقة ل FOV لتحديد بؤر PCNA لتحليلها لاحقا. بدلا من المسح المتسلسل، سجل في نفس الوقت كلا القناتين البصريتين المستخدمتين (الأخضر والأحمر)، لتجنب حركة الخلايا بين المسح الضوئي عند الطولين الموجيين. للحصول على الدقة المناسبة للنقطة استخدم ما لا يقل عن 1024 × 1024 بكسل / دقة الحقل مع تكبير 1x (حجم بكسل 0.29 ميكرومتر على نظام التصوير المستخدم هنا) ، مع سرعة مسح 1/8 إطار / ثانية (4.85 ميكروس / بكسل) بمتوسط 2x. بمجرد تعيين هذه المعلمات في واجهة المستخدم الرسومية A1 LFOV Compact وإطارات منطقة المسح الضوئي A1 LFOV، اضغط على زر الالتقاط لتسجيل FOV.
ملاحظة: من المهم الحفاظ على نفس حجم البكسل خلال التجارب لضمان نتائج قابلة للمقارنة. - لإعداد الإشعاع الدقيق، افتح علامة التبويب تحفيز ND في برنامج التصوير للوصول إلى الجدول الزمني (A1 LFOV / جهاز Galvano). هذا يستخدم الماسحات الضوئية galvano للحصول على سلسلة من الصور قبل التحفيز، وتحفيز (باستخدام الليزر LUN-F 50 mW 405 نانومتر FRAP)، ومن ثم الحصول على سلسلة من الصور بعد التحفيز مرة أخرى باستخدام الماسحات الضوئية غالفانو. إعداد ثلاث مراحل في إطار الجدول الزمني أولا. في العمود Acq/Stim حدد | الاستحواذ | التبييض اقتناء المراحل الثلاث على التوالي. بالنسبة لمرحلة التبييض، قم بتعيين S1 ك ROI.
ملاحظة: في التجربة المعروضة هنا، لم يتم الحصول على أي صور خلال مرحلة التحفيز. - في إطار Galvano XY، قم بإعداد العوامل الرئيسية للإشعاع الدقيق: إخراج طاقة ليزر 405 نانومتر ، وقت الإسهاب (التكرار هو 1 افتراضيا على هذا النظام). في التجارب المعروضة هنا، تم تشعيع الخلايا بالليزر FRAP 405 نانومتر (50 كيلوواط في طرف الألياف) في إنتاج الطاقة 100٪ مع 1000-3000 ميكروس يسكن الوقت.
ملاحظة: نظرا لأن وقت الشغل بالليزر هو على أساس كل بكسل، طالما أن حجم البكسل لا يزال كما هو، فإن العلاقة بين وقت الشغل وكثافة الطاقة ستكون قابلة للمقارنة بين FOVs المختلفة. يوضح الشكل 2A استخدام بروتينات محددة لمسار الاستجابة لضرر الحمض النووي (DDR) (FBXL10 ل DSBs و NTHL1 للضرر الأساسي التأكسدي) لتحسين إعدادات طاقة الليزر لتحريض تلف محدد. تم إنشاء خطوط الخلايا المستقرة هذه بعدوى فيروسية بعد القسم 2 من البروتوكول.
- قبل التشعيع الدقيق للخلايا ، خذ صورة أعلى دقة ل FOV لتحديد بؤر PCNA لتحليلها لاحقا. بدلا من المسح المتسلسل، سجل في نفس الوقت كلا القناتين البصريتين المستخدمتين (الأخضر والأحمر)، لتجنب حركة الخلايا بين المسح الضوئي عند الطولين الموجيين. للحصول على الدقة المناسبة للنقطة استخدم ما لا يقل عن 1024 × 1024 بكسل / دقة الحقل مع تكبير 1x (حجم بكسل 0.29 ميكرومتر على نظام التصوير المستخدم هنا) ، مع سرعة مسح 1/8 إطار / ثانية (4.85 ميكروس / بكسل) بمتوسط 2x. بمجرد تعيين هذه المعلمات في واجهة المستخدم الرسومية A1 LFOV Compact وإطارات منطقة المسح الضوئي A1 LFOV، اضغط على زر الالتقاط لتسجيل FOV.
- التصوير الفاصل الزمني.
- إعداد التصوير الفاصل الزمني للنافذة الزمنية المطلوبة والفواصل الزمنية باستخدام الجدول الزمني و GUI المدمج A1 LFOV و الإطارات منطقة المسح الضوئي A1 LFOV. في التجارب المعروضة هنا ، تم تصوير توظيف EXO1b و PCNA لمدة 12 دقيقة ، مسح FOV كل 5 ثوان ، في 1024 × 1024 بكسل / حقل ، وذلك باستخدام 1x التكبير (مما أدى إلى حجم بكسل 0.29 ميكرومتر على نظام التصوير المستخدمة هنا) مع 0.35 إطار / ثانية سرعة المسح الضوئي (1.45 ميكرو / بكسل) دون متوسط للحد من تبييض الصور.
- تحسين قوة الليزر ٪، وكسب وإزاحة إعدادات للحد من تبييض الصور أثناء التصوير في إطار واجهة المستخدم الرسومية المدمجة A1 LFOV. إذا كان أحد يهدف إلى قياس كل من POI وPCNA، استخدم المسح الضوئي المتزامن بدلا من المسح التسلسلي لتجنب حركة الخلية بين مسح الحقل للفلوروفوريس المنفصلين.
- تم استخدام نظام التصوير مع الإعدادات التالية. لخط الليزر 488 نانومتر (20 كيلوواط): 7٪ قوة الليزر, كسب: 45 (GaAsP كاشف) مع وإزاحة من 2, لخط الليزر 561 نانومتر (20 كيلوواط): 4٪ قوة الليزر, كسب 40 (GaAsP كاشف) مع وإزاحة 2.
- اعتمادا على الحركية من البروتين، تمديد أو تقصير الفاصل الزمني بين الصور أو مدة الفاصل الزمني الإجمالي. في إطار جدول زمني تعيين الفاصل الزمني المطلوب والمدة لصف اقتناء المرحلة الثالثة.
- اضغط تشغيل الآن لتنفيذ التشعيع الدقيق والتصوير الفاصل الزمني اللاحق.
- في نهاية التصوير الفاصل الزمني ، حفظ ROIs التحفيز كصور منفصلة ، والتي ستكون مساعدة مفيدة لتحديد إحداثيات الإشعاع الدقيق في أي برنامج المصب المستخدمة للتحليل.
- تلطيخ المناعة.
ملاحظة: توضح الخطوة 5.1.3 والشكل 2A استخدام بروتينات إصلاح الحمض النووي المعروفة لتقييم أنواع آفات الحمض النووي التي أدخلها الإشعاع الدقيق. يمكن أيضا اكتشاف بعض آفات الحمض النووي باستخدام أجسام مضادة محددة بعد إصلاح الخلايا. ومن الممكن أيضا الكشف عن تجنيد POI عن طريق الكشف عن الأجسام المضادة للبروتين الذاتية. يتم عرض التصور من γH2A.X للتحقق من وجود DSBs أدناه (الشكل 2B). ويبين الشكل 3 اتساق توطين ال PCNA وتوظيفها طوال دورة الخلية لكل من PCNA الموسومة الذاتية والخارجية.- بعد الخطوة 5.1.3، خذ صورة واحدة فقط بعد التشعيع الدقيق لضمان حدث FRAP المناسب على أساس توظيف mPlum-PCNA. يحيط علما الإحداثيات الدقيقة للFOV للعثور على الحقل في وقت لاحق بعد وضع العلامات immunofluorescent.
- أخرج غرفة زراعة الخلايا من المجهر واحتضن الخلايا عند 37 درجة مئوية في جو رطب يحتوي على 5٪من ثاني أكسيد الكربون لمدة 5-10 دقائق.
ملاحظة: Paraformaldehyde (PFA) سامة، وينبغي أن يتم العمل في منطقة جيدة التهوية أو غطاء الدخان. سيتم إجراء جميع عمليات الغسيل والحضانة اللاحقة بأحجام 0.5 مل في شريحة غرفة البئر 4. بعد وقت الحضانة، اغسل الخلايا ب 0.5 مل من برنامج تلفزيوني (137 mM NaCl، 2.7 mM KCl، 8 mM Na2HPO4، و 2 mM KH2PO4)وإصلاح مع 0.5 مل من 4٪ PFA في PBS لمدة 10 دقائق في درجة حرارة الغرفة (RT). - غسل الخلايا مرة واحدة مع برنامج تلفزيوني، ثم غسلها مع 50 mM NH4Cl لإرواء PFA المتبقية.
- Permeabilize الخلايا لمدة 15 دقيقة في RT مع 0.1٪ تريتون X-100 في برنامج تلفزيوني.
- منع العينات لمدة 1 ساعة مع حظر العازلة (5 ٪ FBS ، 3 ٪ جيش الشعب البسا ، 0.05 ٪ تريتون X - 100 في برنامج تلفزيوني).
- إزالة حل حظر وإضافة الأجسام المضادة الأولية المخفف (anti-γH2A.X، 1:2000) في حظر العازلة لمدة 1 ساعة في RT.
- غسل الآبار مع حظر العازلة 3 × 10 دقيقة.
- إضافة الأجسام المضادة الثانوية المخففة (المضادة للفأرة اليكسا 488 زائد اقتران، 1:2000) في حظر العازلة لمدة 1 ساعة في RT.
- غسل الآبار مع حظر العازلة 3 × 10 دقيقة.
- مضادةالنواة مع 1 ميكروجرام / مل DAPI الحل في برنامج تلفزيوني لمدة 15 دقيقة.
- غسل الخلايا مرة واحدة مع برنامج تلفزيوني. يمكن إجراء التصوير مباشرة في برنامج تلفزيوني أو حل برنامج تلفزيوني مع الكواشف المضادة للضفد (على سبيل المثال، AFR3) للحد من photobleaching.
6. تحليل التوظيف
ملاحظة: يظهر الشكل 4A صورا تمثيلية لتوظيف Exo1b وPCNA في وجود DMSO أو olaparib. يظهر الشكل 4B صورة تمثيلية لتحليل البيانات. تم حساب متوسط قيم الفلورسينس عن طريق قياس متوسط كثافة AcGFP باستخدام مستطيل على طول مسار الليزر الذي تم تمييزه بواسطة mPlum-PCNA (مستطيلات صفراء) عبر نقاط زمنية مختلفة باستخدام فيجي. يمكن أن يعمل PCNA كتحكم داخلي لتسليط الضوء على الإشعاع الناجح على طول إحداثيات عائد الاستثمار. وبالمثل، تم حساب قيم مضان AcGFP المتوسطة أيضا للمناطق غير التالفة من النواة (B والمستطيلات الزرقاء). تم قياس كثافة إشارة الخلفية في المناطق غير المأهولة (C والمستطيلات الحمراء) وتم طرحها من متوسط قيم الفلورسنت(الشكل A و B). وهكذا، تم حساب متوسط وحدة الفلورسنت النسبية لكل نقطة جمع بيانات بواسطة المعادلة RFU = (A − C) / (B − C)8،9. يتم تطبيع قيم RFU الناتجة عن المنطقة المشععة الدقيقة إلى قيم RFU قبل التشعيع الدقيق.
- لتحديد المنطقة A للموقع المشعع الدقيق، استبعاد المناطق النووية، بؤر النسخ المتماثل، والمناطق النووية غير النظامية للخلية من القياس. عقد مفتاح التحول بين رسم اثنين من ROIs في فيجي إلى المجموعة منطقتين منفصلتين كواحد.
ملاحظة: سوف يتفاوت توظيف البروتين بين الجينات المختلفة وظروف التشعيع؛ وبالتالي، يجب تحديد حجم المنطقة ألف بشكل فردي. بمجرد تحديد عرض بكسل المنطقة A، يجب أن يظل ثابتا لأي عمليات توظيف مقارنة. في التجارب المعروضة هنا، تم استخدام مستطيلات بعرض 7 بكسل. - استبعاد الخلايا التي انتقلت خلال مدة مقاطع الفيديو المسجلة من التحليل. لتضمين خلايا متحركة للغاية، يجب إجراء التحليل الموصوف من إطار إلى آخر.
- لتصور ملف تعريف التوظيف، رسم قيم RFU العادية مع الوقت باستخدام برنامج إحصائي.
- احسب الفرق عند نقطة زمنية مشار إليها بين علاج DMSO وolaparib (n=31) باستخدام اختبار Mann-Whitney.
Representative Results
تعالج الخلايا كل نوع من آفة الحمض النووي بطريقة محددة تعتمد أيضا على مرحلة دورة الخلية التي هي فيها. على سبيل المثال، بعد التشعيع الدقيق، ستتم معالجة فواصل مزدوجة تقطعت بها السبل (DSB) إما عن طريق الانضمام إلى نهاية غير متجانسة (NHEJ) أو الموارد البشرية اعتمادا على مرحلة دورة الخلية. النوى التي تعمل على نطاق واسع خلال مرحلتي S و G2 من دورة الخلية تخلق تراكمات الحمض النووي التي تعتبر حاسمة للموارد البشرية المناسبة. لتعزيز تقييم الخلايا في مرحلة S، تم استخدام PCNA كعلامة دورة الخلية ذات اللون الواحد. يوضح الشكل 1A ملف تعريف التعريب ل mPlum-PCNA أثناء تطور دورة الخلية. PCNA لديه توزيع متجانسة تماما في النواة في المرحلة G1 و G2 (في حين يجري أيضا استبعاد معظمها من النيوكليولي). في مرحلة S، PCNA مترجمة إلى مواقع النسخ المتماثل الحمض النووي، والتي يمكن تصورها على أنها نقاط مضيئة في النواة. في خلايا المرحلة S المبكرة ، تكون البقع صغيرة نسبيا وتوزع بالتساوي في جميع أنحاء نواة الخلية. تتقدم إلى منتصف مرحلة S، تصبح البقع غير واضحة وترجمة أكثر نحو محيط النواة والنوكليولي. في مرحلة S المتأخرة، تقلل البقع في الأرقام ولكنها تصبح كبيرة بشكل متزايد حيث يركز PCNA في مواقع النسخ المتماثل المتأخرة(الشكل 1B). والأهم من ذلك، كان تعبير PCNA الخارجي من العمود الفقري ناقلات pBABE أقل من المستويات الذاتية ولكنه كان كافيا للكشف عن طريق المجهر الذي يقلل من القطع الأثرية المحتملة في تطور دورة الخلية وDDR. ويبين الشكل 1C مدى فرط التعبير عن PCNA مقارنة بالمستويات الذاتية. يرجى ملاحظة أن الفرقة المقابلة ل mPlum-PCNA تهاجر أبطأ بسبب حجمها الأكبر.
لقد استهدفنا إدخال DSBs أثناء الإشعاع الدقيق للتحقيق في تجنيد PARP1/2 المعتمد على EXO1b لهذه الآفات في مرحلة S. ويبين الشكل 2 ألف أن جرعات منخفضة من الطاقة (1000 ميكروس يسكن الوقت) لا تحفز تجنيد EGFP-FBXL10، وهو المستجيب DSB (مكون من مجمع FRUCC 8)،في حين كان كافيا للحث على تجنيد NTHL1-mCherry، إصلاح قاعدة (BER) بروتين المسار، والتجنيد لمواقع تلف الحمض النووي التأكسدي10،11،12. في 3000 ميكروس يسكن الوقت، كل من EGFP-FBXL10 وNTHL1-mCherry تجنيد، مما يدل على إخراج الليزر الذي يولد كل من الآفات التأكسدية وDSBs. تعزيز هذه النتائج، الشكل 2B يظهر تلطيخ immunofluorescence ضد γH2A.X (علامة DSB)، والذي هو بوضوح أكثر وضوحا عند استخدام جرعات الطاقة العالية. PCNA بمثابة علامة دورة الخلية وعلامة للتشعيع الدقيق الناجح ، حيث أنه يجند بشكل كاف مع كل من إعدادات وقت السكن بالليزر. الأهم من ذلك، يمكن استخدام كل من الفلورسنت الخارجي و / أو الذاتية البروتين الموسومة PCNA لهذه الوظيفة مراسل لأنها تتصرف بالمثل(الشكل 3). تم تصميم PCNA الموسومة داخليا عن طريق إدراج mRuby في الإطار مع أول إكسون في أليل واحد من موقع PCNA13 (كان خط الخلية هدية من نوع يورغ مانسفيلد).
ويبين الشكل 4A والشكل 4C توظيف EXO1b الموسومة ب AcGFP في خلايا المرحلة S. يصل EXO1b إلى أقصى مستوى من التراكم في مواقع الإشعاع الدقيق حوالي دقيقة واحدة ثم يبدأ ببطء في فك الارتباط من آفات الحمض النووي بعد ذلك. ويشار إلى عمليات التخصيب في مواقع الإشعاع الجزئي بوحدة مضان نسبية واحدة > على الرسم البياني. في وجود olaparib ، تراكم EXO1b في شريط الليزر في دقيقة واحدة هو أقل بكثير مقارنة مع التحكم في السيارة. هذه النتائج متفقة مع الأدب6،7. ويبين الشكل 4 باء المناطق التمثيلية للقياس الكمي (المناطق ألف وباء وجيم) على النحو المبين في النقطة 6 من البروتوكول. ويبين الشكل 4D مستويات التعبير القابلة للمقارنة من EXO1b الذاتية والخارجية EXO1b-AcGFP في الخلايا المستخدمة في الإشعاع الدقيق.
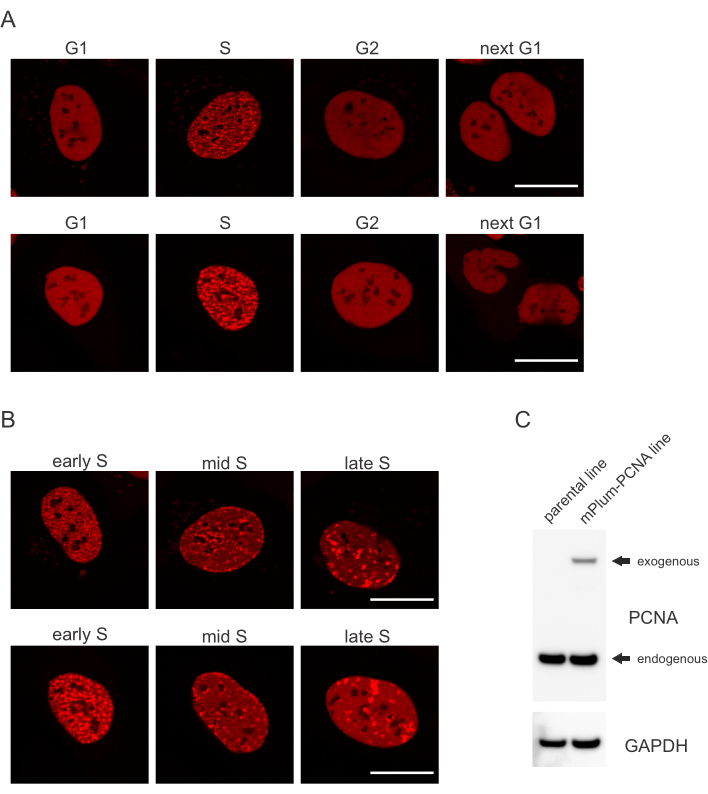
الشكل 1: نمط توطين PCNA. (أ) تظهر الصور نمط توطين PCNA المتكامل بشكل ثابت والخارجي طوال دورة الخلية في خلايا نظام التشغيل U-2. (ب)تظهر الصور أنماط بؤر PCNA في مراحل مختلفة من مرحلة S (في وقت مبكر، منتصف، وفي وقت متأخر) في خلايا نظام التشغيل U-2. (ج) لطخة الغربية تظهر مستويات الذاتية والخارجية من PCNA في خلايا نظام التشغيل U-2 المستخدمة للتصوير. يمثل شريط المقياس 20 ميكرومتر. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 2: يمكن تحسين تحريض DSBs من خلال إخراج الطاقة بالليزر الأمثل. (أ) إعدادات الليزر للحث على أشكال مختلفة من تلف الحمض النووي. تم استخدام خلايا نظام التشغيل U-2 التي تعبر بشكل ثابت عن كل من EGFP-FBXL10 و NTHL1-mCherry لتحديد DSBs ومواقع الآفات التأكسدية ، على التوالي. تم إجراء التشعيع الدقيق مع خط ليزر 405 نانومتر على خلايا OS U-2 غير المتزامنة مع إما 1000 ميكرومتر أو 3000 ميكروس من وقت الإقامة. مقياس شريط يمثل 20 ميكرومتر. (ب) Immunofluorescent تلطيخ ضد γH2A.X تم القيام به على خلايا ظهارية صبغ الشبكية البشرية (hTERT RPE-1) وجود mRuby الموسومة PCNA الذاتية. تم إصلاح الخلايا ومعالجتها بعد 5 دقائق من الإشعاع الدقيق إما مع 1000 ميكرو ميكروس أو 3000 ميكروس من وقت السكن. يمثل شريط المقياس 20 ميكرومتر. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 3:توظيف مماثلة من mRuby PCNA الذاتية والخارجية mPlum-PCNA لمواقع الإشعاع الدقيق في 1000 ميكرو ميكرو أو 3000 ميكرو ثانية ليزر يسكن الوقت. كل من الذاتية والخارجية الموسومة PCNA شكل بؤر النسخ المتماثل خلال مرحلة S. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 4: PARP1/2-تعتمد على توظيف EXO1b في مرحلة S. U-2 خلايا نظام التشغيل التي تعبر بشكل ثابت EXO1b-AcGFP و mPlum-PCNA كانت مشعة بشكل دقيق مع خط ليزر FRAP 405 نانومتر باستخدام 3000 ميكروجرام من وقت الإقامة. (أ) صور تمثيلية للخلايا المشععة الدقيقة في النقاط الزمنية المشار إليها بعد المعالجة المسبقة إما مع التحكم في المركبة (DMSO) أو أولاباريب (1 ميكرومتر). يمثل شريط المقياس 20 ميكرومترا (B) الصور التمثيلية للمناطق المحددة من مناطق A وB وC لتحليل التوظيف. يمثل شريط المقياس 20 ميكرومتر (C) تم التقاط ديناميكيات توظيف تلف الحمض النووي عن طريق تصوير الخلايا الحية. تم الحصول على قيم وصور متوسطة نسبيا كل 5 ق لمدة 12 دقيقة. لكل حالة، تم تقييم ≥30 خلية. تم رسم قيم الفلورسينس النسبية (الخطوط السوداء الصلبة) والخطأ القياسي (النطاق الذي تصوره منطقة مظللة) مقابل الوقت. يظهر الخط المتقطع قيم التوظيف في دقيقة واحدة بعد الإشعاع الدقيق. تم حساب الفرق بين DMSO (n = 32) وolaparib (n = 31) العلاج باستخدام اختبار مان ويتني. أستريكس يدل على p<0.0001. (D) لطخة الغربية يقارن مستويات التعبير من EXO1b الذاتية والخارجية EXO1b-AcGFP في الخلايا المستخدمة للتشعيع الدقيق. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
Discussion
الخطوات الهامة واستكشاف الأخطاء وإصلاحها المحتملة للبروتوكول/تعديلاته
أنسجة السليم وعاء زراعة للإشعاع الدقيق أمر بالغ الأهمية للنجاح. تم تحسين معظم أنظمة التصوير عالية الدقة لسمك زجاج الغطاء 0.17 مم. استخدام غرف التصوير سمك أعلى أو أقل أو تلك المصنوعة من البوليمرات البلاستيكية (غير الأمثل للتصوير 405 نانومتر)، يمكن أن تقلل بشكل كبير من جودة الصورة. عند استخدام الأسطح الزجاجية، تأكد من أنها زراعة الأنسجة تعامل لتعزيز التصاق الخلايا. إذا لم يتم علاجها من الأنسجة والثقافة، وهذه الغرف تحتاج إلى أن تكون مغلفة، على سبيل المثال، مع بولي-د-ليسين قبل بذر الخلايا. عند طلاء الخلايا في coverglass الغرفة، كثافة الخلية المثالية أمر بالغ الأهمية لتجنب المخالفات دورة الخلية والإجهاد إضافية للخلايا. التوازن الحراري السليم لمكونات المجهر قبل التجريب للحفاظ على درجة حرارة مستقرة أمر بالغ الأهمية للحفاظ على التركيز على حد سواء طوال التصوير الفاصل الزمني وضروري أيضا لضمان DDR متجانسة عبر الزمن والعينات.
من المهم أن تكون الخلايا في حالة صحية قبل التشعيع الدقيق لتقليل البيانات اليدوية. إذا كانت الخلايا لديها مورفولوجيا غير منتظمة بعد العدوى / الاختيار ، فسمح للخلايا بالتقدم من خلال مقاطع متعددة حتى يعود مورفولوجيا إلى وضعها الطبيعي. تأكد دائما من أن خطوط الخلايا المستخدمة خالية من التلوث الميكوبلازما. من بين العديد من الآثار السلبية لعدوى الميكوبلازما ، فإنه يسبب أيضا تلف الحمض النووي للخلايا المضيفة ويمكن أن يؤثر على مسارات DDR الخاصة بهم14،15. الطريقة الأكثر حساسية للكشف عن الميكوبلازما في ثقافة الخلية هي من خلال PCR (مقابل الكشف مع DAPI أو Hoechst).
وينبغي أن يكون التعبير الأمثل عن البروتين إصلاح الفائدة مماثلة لمستويات الذاتية، ومع ذلك، عالية بما يكفي للكشف. يمكن تعديل المروج المستخدم على ناقلات الفيروسات ، والتيتر الفيروسي أثناء العدوى ، وطول وقت العدوى لمستويات التعبير المثالية. للحصول على نتائج متسقة، قم بعزل استنساخ الخلايا الفردية لضمان مستويات التعبير المتجانسة ومورفولوجيا الخلايا العادية. فمن المستحسن استخدام ناقلات يبني التي لا تفرط في التعبير PCNA الموسومة في مستويات أعلى من الذاتية لدورة الخلية المناسبة ووظيفة علامة إصلاح الحمض النووي. حتى مستويات منخفضة من التعبير المفرط PCNA كافية للتمييز S-الخلايا المرحلة. وقد استخدمت ناقلات pBABE الرجعية بنجاح لهذا الغرض (أدجين # 1764، # 1765، # 1766، # 1767). يمكن وضع علامة PCNA مع أي بروتينات فلورية خضراء أحادية(على سبيل المثال ، mPlum ، mCherry ، mRuby ، إلخ) أو بروتينات فلورية خضراء أحادية (على سبيل المثال ، mEGFP ، AcGFP ، mWasabi ، mNeonGreen ، mEmerald ، وما إلى ذلك) والتي يمكن دمجها بعد ذلك مع POI الموسومة بالتناوب. الإفراط في التعبير عن POI الموسومة الفلورسنت لديه بعض القيود والاعتبارات. قد تعطل العلامات الفلورية وظيفة البروتين العادية والتوطين. وبالتالي، يجب النظر في موقع العلامة (N أو C-terminal). استخدم دائما البروتينات الفلورية أحادية الألوان ، حيث يمكن أن يؤثر احتكار القلة للمتغيرات غير أحادية الشكل على وظيفة POI.
يجب تحديد إعدادات الليزر لكل نظام تصوير لأن العديد من مكونات المسار البصري ستؤثر على الطاقة الفعلية التي يتم توصيلها إلى الخلايا. الليزر التشعيع الدقيق يمكن أن يسبب عدة أنواع من آفات الحمض النووي اعتمادا على الطول الموجي الإثارة، وإنتاج الطاقة من الليزر FRAP وإذا تم استخدام أي عوامل ما قبل التوعية (مثل Bromodeoxyuridine أو Hoechst). 405 نانومتر الليزر يمكن أن يسبب تلف الحمض النووي التأكسدي, فواصل تقطعت بهم السبل واحد ومزدوج16,17. باستخدام إعدادات إخراج ليزر أعلى، يزيد مقدار DSBs. في هذا البروتوكول لم تستخدم أساليب التوعية المسبقة، ولكن هذه التقنيات مغطاة إلى حد كبير في الأدبيات وإعادة توج في المناقشة أدناه. في رأينا ، فإن أفضل طريقة لاختبار ما إذا كانت الآفة المطلوبة يتم إنشاؤها عن طريق اختبار لتوظيف جينات محددة معروفة لمسار تلف الحمض النووي. تجنيد NTHL1 أو OGG1، مكونات مسار BER، يشير إلى تحريض قواعد الحمض النووي المؤتأكسدة10،11،17،18،19،في حين FBXL10 أو XRCC5 تشير إلى وجود DSBs8،20،21. تجنيد XRCC1 يمكن أن تشير إلى وجود كل من قواعد الحمض النووي المؤتأكسدة وفواصل واحدة تقطعت بهم السبل (SSB)22،23. XPC (أي، RAD4) هو مؤشر جيد من NER أن يزيل إضافات الحمض النووي الضخمة التي تم إنشاؤها بواسطة الأشعة فوق البنفسجية (UV)17،24. لأن تجنيد البروتينات الخارجية قد يدخل بعض المخالفات، يمكن أن تلطيخ immunofluorescent من البروتينات إصلاح الحمض النووي الذاتية أو علامات (مثل γH2A.X فواصل تقطعت بهم السبل مزدوجة) تأكيد وجود آفات الحمض النووي محددة. وبدلا من ذلك، يمكن أيضا استخدام الأجسام المضادة التي تثار ضد أنواع محددة من آفات الحمض النووي. لضبط طاقة الليزر التي تم تسليمها، يمكن تغيير كل من وقت السكن وقوة الليزر.
بمساعدة النمذجة الرياضية ، يمكن إجراء تحليل حركي مفصل يمكن أن يوفر رؤى قيمة في خصائص التوظيف في POI (على سبيل المثال ، مساهمة مجالات ربط الحمض النووي المتعددة ، والحساسية تجاه أحداث الإشارات المختلفة ، وما إلى ذلك). يمكن الجمع بين تقييم التوظيف الآلي وتتبع الخلايا لإنشاء مهام سير عمل قوية 1و25.
مزايا وقيود الحمض النووي قبل التوعية
قبل التوعية من الحمض النووي قبل التشعيع الدقيق هو أداة شائعة الاستخدام لإصلاح الحمض النووي البروتين التوظيف16،17. إن توعية الحمض النووي قبل التشعيع الدقيق تجعلها أكثر عرضة ل DSBs. الطريقتان الأكثر شيوعا للتوعية المسبقة للحمض النووي هما المعالجة المسبقة للخلايا إما بصبغة بروموديوكسيوريدين (BrdU) أو صبغة Hoechst. بالنسبة للأنظمة غير القادرة على التشعيع الدقيق في قوى الليزر العالية ، قد تكون هذه الأساليب ضرورية لتحفيز آفات الحمض النووي مثل DSBs. بالإضافة إلى ذلك ، في غياب كاشف ضوء منقول أو إشارة فلورية تسلط الضوء على نواة الخلية (على سبيل المثال ، عند دراسة توظيف بروتينات إصلاح الحمض النووي الذاتية غير الموسومة) ، يعمل Hoechst كأداة توعية مسبقة ووصمة عار نووية فلورية. ومع ذلك، يمكن أن يؤدي التحسيس المسبق للحمض النووي إلى مضاعفات كبيرة. BrdU (تستخدم بتركيز نهائي قدره 10 ميكرومتر) يجب أن تضاف إلى الخلايا 24 ساعة (أو ما يعادل الوقت لدورة الخلية الكاملة في خط الخلية المستخدمة) لدمج بشكل صحيح في الحمض النووي ويمكن أن يسبب تدخل دورة الخلية26. Hoechst 33342 (يستخدم بتركيز نهائي قدره 1 ميكروغرام / مل) هو سام للخلايا بعد فترات حضانة طويلة ولكنه يتطلب وقتا كافيا لتشبع النواة بالصبغة. ولذلك، ينبغي أن تطبق فقط 15-20 دقيقة قبل التشعيع الدقيق. وإلا فإن بيانات التوظيف لن تكون متسقة. الخلايا الملطخة بهذه الطريقة لا يمكن أن تبقى في الثقافة لأكثر من بضع ساعات27،28. تأكد من عدم استخدام Hoechst 33358 ، والتي ليست خلية نفاذية مثل صبغة Hoechst 33342. يمكن أن تقدم التوعية المسبقة أيضا تباينا غير ضروري بين التجارب وتجعل التجربة أكثر حساسية للاختلافات في كثافة الخلايا (لأن هذا سيؤثر على كمية الصبغة / الخلية المدمجة).
مزايا وقيود المجهر البؤري
يمكن أن تكون سرعة التصوير بالمنظار المجهري الكونفوجكال محددة بالمقارنة مع المجهر واسع النطاق. ومع ذلك ، يمكن للمجهر confocal مجهزة ماسح ضوئي رنانة تحسين سرعة التصوير بشكل كبير (على حساب القرار) تقترب من سرعات المجهر الغزل القرص. ثلاث ميزات تجعل من نظام A1R HD25 confocal خيارا ممتازا للبروتوكول المعروض هنا. أولا، يجعل FOV 25 مم من النظام من الممكن تصوير ما بين 15-20 خلية في حقل واحد ممسوحة ضوئيا (مقابل 5-10 خلايا في الاجهزة العادية)، والحد من عدد عمليات الاستحواذ اللازمة للحصول على ما يكفي من الخلايا للتحليل الإحصائي. ثانيا، تتيح وحدة FRAP واثنين من رؤوس المسح الضوئي تصوير الخلايا وتشعيعها في وقت واحد، وليس فقط بالتتابع. وأخيرا، فإن مرونة وجود كل من الماسحات الضوئية الرنانة والجالفانو توفر القدرة على التبديل بسهولة بين التصوير عالي الدقة الزمنية بسرعة استثنائية تقلل من إخماد الفلوروفوريس، والتصوير عالي الدقة المكانية الذي يستخدم سرعات مسح أبطأ لإنتاج صور ذات نسبة إشارة أعلى إلى الضوضاء. وفي حين أن النظام المستخدم يسمح بالمرونة المذكورة أعلاه، لتشبه تكوينات المجهر الكونفوجال المتاحة على نطاق أوسع، لم يستخدم سوى الماسح الضوئي الجلفانو في التجارب المعروضة (لكل من التشعيع الدقيق والتصوير اللاحق).
مزايا والقيود المفروضة على الإشعاع الجزئي
وفي حين أن الإشعاع الجزئي يوفر دقة مكانية وزمنية لا مثيل لها، فإنه لا يخلو من القيود. تلف الحمض النووي عن طريق الليزر التشعيع الدقيق تتجمع بشكل كبير إلى أجزاء محددة من النواة بالمقارنة مع العوامل الضارة التي تحدث بشكل طبيعي. وبالتالي، قد تختلف استجابة الكروماتين بسبب الإشعاع الدقيق مقارنة بالضرر الموزع بشكل متجانس. بالإضافة إلى ذلك ، فإن الإشعاع الدقيق يستغرق وقتا طويلا ولا يمكن إجراؤه إلا على بضع عشرات من الخلايا ، في حين أن الطرق الكيميائية الحيوية الكبيرة القائمة على السكان (تجزئة الكروماتين ، وسرعة المناعة ، وChIP) يمكن أن توفر قوة متزايدة من خلال دراسة آلاف الخلايا في وقت واحد. 10- إن التحقق من الملاحظات التي يجريها التشعيع الدقيق بالتقنيات الكيميائية الحيوية التقليدية هو استراتيجية فعالة للتوصل إلى استنتاجات موثوقة. على الرغم من أن التشعيع الدقيق المتزامن للعديد من الخلايا في FOV معينة ممكن ، فإن نظام التصوير سيحتاج إلى مزيد من الوقت لأداء المهمة. ولذلك، فإن قياس ديناميات البروتينات التي تجند بسرعة كبيرة لآفات الحمض النووي يحد من عدد ROIs المحتملة للتشعيع الدقيق المستخدم في وقت واحد. على نظام التصوير المستخدمة لهذا البروتوكول، والإشعاع الدقيق من واحد 1024 بكسل طويل ROI يستغرق 1032 مللي ثانية باستخدام 1000 ميكروس يسكن الوقت و 3088 مللي ثانية باستخدام 3000 ميكروس يسكن الوقت لإكمال. استخدام خطوط متعددة من ROIs سيزيد بشكل كبير من الوقت اللازم لإنهاء الإشعاع الجزئي (على سبيل المثال، 7 × 1024 بكسل طويل ROI يستغرق 14402 مللي ثانية باستخدام 1000 ميكروس يسكن الوقت و 21598 مللي ثانية باستخدام 3000 ميكروس يسكن الوقت). هذه المرة تضيع من الحصول على صورة ويجب أن تؤخذ في الاعتبار. عند تصوير أحداث التوظيف السريع، استخدم أقصر عائد استثمار ممكن وفقط خلية صغيرة مشععة في كل مرة.
مزايا وقيود على أساليب المزامنة
بالنسبة للدراسات الخاصة بدورة الخلية، تتضمن الطرق الحالية إما مزامنة الخلايا في مراحل محددة لدورة الخلية أو استخدام المراسلين الفلوريين لتحديد مرحلة دورة الخلية المحددة للخلية. ومع ذلك، كل من هذه الطرق يوفر التحديات الخاصة بهم والقيود.
نظام FUCCI3 (الاعتماد على البروتين الفلوري الموسومة أشكال مبتورة من CDT1 والجوزمين) هو أداة مفيدة بشكل خاص لدراسات دورة الخلية ولكن لديه قيود عندما يتعلق الأمر بالتفرقة بين مراحل S و G2 من دورة الخلية. مستويات الجوزاء مرتفعة بالفعل من منتصف مرحلة S وتبقى عالية حتى مرحلة M، مما يجعل من الصعب فصل هذه المراحل. استخدام نظام FUCCI يعني أيضا أنه لا يمكن استخدام قناتين بصريتين للمجهر لتصوير POI.
يمكن مزامنة خطوط الخلايا غير السرطانية في G0 عن طريق إزالة عوامل النمو الموجودة في المصل (تجويع المصل) مما يسبب ضررا ضئيلا أو معدوما للحمض النووي للخلايا. ومع ذلك، فإن معظم خطوط الخلايا السرطانية تستمر جزئيا في التقدم من خلال دورة الخلية حتى من دون كميات كافية من المصل في وسائل الإعلام الخاصة بهم. بالإضافة إلى ذلك، تبدأ الخلايا جزئيا في فقدان المزامنة بواسطة أواخر G1، مرحلة S المبكرة. بالإضافة إلى المجاعة المصل، وهناك العديد من الطرق الكيميائية لتحقيق تزامن دورة الخلية. Hydroxyurea، المنديكولين، وكتل الثيميدين هي طرق لوقف تكرار الحمض النووي لمزامنة الخلايا في مرحلة S في وقت مبكر. في حين أن هذه الأساليب رخيصة وبسيطة ، فإنها تدخل إجهاد النسخ المتماثل مما يؤدي إلى تلف الحمض النووي. وقد ثبت أن مثبطات تكرار الحمض النووي هذه تحفز على الفوسفور H2A. X، علامة معروفة من DSBs2،29. طريقة استخدام الموسومة PCNA كعلامة لخلايا المرحلة S يقلل من احتمال التحف الناجمة عن التزامن الكيميائي ويمكن تطبيقها على مجموعة واسعة من خطوط الخلايا مقارنة مع المجاعة المصل.
استنتاج
تلف الحمض النووي هو القوة الدافعة للأمراض الوراثية حيث الآفات المطفرة يمكن أن يؤدي إلى التحول الخبيث للخلايا. استهداف آلات تخليق الحمض النووي هو استراتيجية علاجية أساسية في علاج الأمراض المفرطة الانتشار مثل السرطان. من أجل علاج هذه الأمراض بطريقة أكثر استهدافا، نحن بحاجة إلى فهم أفضل للبروتينات التي تصلح آفات الحمض النووي. ويساعد البروتوكول الوارد وصفه هنا الدراسات القائمة على الإشعاع الجزئي في مرحلة S عن طريق التقليل إلى أدنى حد من التحديات التي تطرحها أساليب المزامنة التقليدية للحد من القطع الأثرية المحتملة وزيادة إمكانية استنساخ التجارب.
Disclosures
ويقول المؤلفان إن نشر العمل المعروض كان برعاية شركة نيكون. ويعلن المؤلفان أنه لا توجد مصالح متنافسة.
Acknowledgments
يشكر المؤلفون م. باغانو على دعمه المستمر وكذلك د. سيمونشي وأ. مارزيو وجي تانغ على مراجعتهم النقدية للمخطوطة. ب. ميواتاني مينتر يشكر ر. ميواتاني وب. مينتر على دعمهما المستمر. ج. رونا يشكر ك. رونان جوراز وجي رونا على دعمهما المستمر.
Materials
| Name | Company | Catalog Number | Comments |
| Ammonium chloride | Sigma-Aldrich | A9434-500G | For quenching formaldehyde |
| Anti-EXO1 Rabbit Polyclonal Antibody | Proteintech | 16253-1-AP | primary antibody |
| Anti-phospho-Histone H2A.X (Ser139) Antibody, clone JBW301 | Millipore | 05-636 | primary antibody |
| Bovine Serum Albumin | Sigma-Aldrich | 3117332001 | BSA for blocking |
| BrdU (5-Bromo-2'-deoxyuridine) | Merck | 19-160 | pre-sensitizing agent |
| Citifluor™ Mountant Solution AFR3 | Electron Microscopy Sciences | 17973-10 | antifade containing PBS solution for imaging |
| DAPI | Sigma-Aldrich | D9542-1MG | nucleic acid stain |
| DMEM Medium | Thermo Fisher Scientific | 10569010 | Cell culture medium for HEK293T cells |
| DMSO | Sigma-Aldrich | D2650-100ML | Vehichle control and dissolution solvent |
| EGFP-FBXL10 | Addgene | #126542 | viral expression vector for EGFP-FBXL10 |
| EXO1b-AcGFP (in pRetroQ) | custom cloning | na | EXO1b cDNA was cloned in the NheI, BamHI sites of pRetroQ-AcGFP1-N1 vector. |
| Fetal Bovine Serum | Gibco | 16140071 | Media supplement |
| FluoroBrite DMEM | Thermo Fisher Scientific | A1896701 | Phenol red free medium for microscopy |
| Goat anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 | Thermo Fisher Scientific | A32723 | secondary antibody |
| HEK293T cells | ATCC | ATCC CRL-3216 | Cell line for viral packaging |
| HEPES | Sigma-Aldrich | H0887-100ML | Buffering agent to supplement live cell imaging medium |
| Hoechst 33342 | Thermo Fisher Scientific | H3570 | pre-sensitizing agent |
| Lipofectamine 3000 | Thermo Fisher Scientific | L3000015 | Transfection reagent |
| McCoy’s 5A (Modified) Medium | Life Technologies | 16600-108 | Cell culture medium for U-2 OS cells |
| mCherry-PCNA | Addgene | #55117 | non-viral PCNA construct suitable for cell cycle marker |
| mPlum-PCNA | Addgene | #55994 | non-viral PCNA construct suitable for cell cycle marker |
| mPlum-PCNA (in pBABE) | custom cloning | na | mPlum-PCNA cDNA was cloned from Addgene #55994 in the BamHI, SalI sites of pBABE (puro) |
| Nikon A1R-HD25 Confocal Scanhead and Controller | Nikon | na | confocal imaging system |
| Nikon LUN4 laser unit | Nikon | na | excitation system |
| Nikon LUN-F 50 mW 405 nm FRAP laser unit | Nikon | na | FRAP laser unit |
| Nikon NIS Elements Confocal Controller Software | Nikon | na | Confocal controlling software |
| Nikon Ti2-E Inverted Microscope | Nikon | na | inverted epifluorescent microscope base |
| Nikon Ti2-LAPP Modular Illumination System | Nikon | na | illumination system |
| NTHL1-mCherry (in pRetroQ) | custom cloning | na | NTHL1 cDNA was cloned in the NheI, SalI sites of pRetroQ-mCherry-N1 vector. |
| Nunc Lab-Tek II Chambered Coverglass (4 well) | Thermo Fisher Scientific | 155382PK | Live cell microscopy cell culture chamber |
| Olaparib | Selleck Chemicals | S1060 | PARP inhibitor |
| Opti-MEM reduced serum media | Thermo Fisher Scientific | 31985062 | Dilution medium for transient transfection |
| Paraformaldehyde aqueous solution (32%) | Thermo Fisher Scientific | 50-980-494 | Fixative |
| pBABE (hygro) | Addgene | #1765 | retroviral expression vector (for low expression levels) |
| pBABE (neo) | Addgene | #1767 | retroviral expression vector (for low expression levels) |
| pBABE (puro) | Addgene | #1764 | retroviral expression vector (for low expression levels) |
| pBABE (zeo) | Addgene | #1766 | retroviral expression vector (for low expression levels) |
| PCNA Antibody (PC10) | Santa Cruz | sc-56 | primary antibody |
| Penicillin-Streptomycin-Glutamine (100x) | Gibco | 10378016 | Media supplement |
| polybrene | Sigma-Aldrich | TR-1003 | Increase viral infection efficiency |
| pRetroQ-AcGFP-C1 | Takara | 632506 | retroviral expression vector |
| pRetroQ-AcGFP-N1 | Takara | 632505 | retroviral expression vector |
| pRetroQ-mCherry-C1 | Takara | 632567 | retroviral expression vector |
| pRetroQ-mCherry-N1 | Takara | 632568 | retroviral expression vector |
| pUMVC | Addgene | #8449 | Viral packaging vector |
| Sodium-pyruvate | Thermo Fisher Scientific | 11360070 | Supplement for live cell imaging medium |
| Triton X-100 aqueous solution (10%) | Sigma-Aldrich | 11332481001 | Dilute in PBS for cell permeabilization buffer |
| Trypsin-EDTA Solution 10X | Sigma-Aldrich | 59418C-100ML | Dilute in PBS to split cells |
| U-2 OS Cells | ATCC | HTB-96 | Optimal cell line for microscopy experiments |
| Universal Mycoplasma Detection Kit | ATCC | 30-1012K | PCR based Mycoplasma detection kit |
| VSV-G | Addgene | #8454 | Viral protein envelope vector |
References
- Aleksandrov, R., et al.
- Darzynkiewicz, Z., Halicka, H. D., Zhao, H., Podhorecka, M. Cell synchronization by inhibitors of DNA replication induces replication stress and DNA damage response: Analysis by flow cytometry. Methods in Molecular Biology. 761, 85-96 (2011).
- Sakaue-Sawano, A., et al. Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell. 132 (3), 487-498 (2008).
- Herce, H. D., Rajan, M., Lattig-Tunnemann, G., Fillies, M., Cardoso, M. C. A novel cell permeable DNA replication and repair marker. Nucleus. 5 (6), 590-600 (2014).
- Keijzers, G., et al. Human exonuclease 1 (EXO1) regulatory functions in dna replication with putative roles in cancer. International Journal of Molecular Sciences. 20 (1), (2018).
- Cheruiyot, A., et al. Poly(ADP-ribose)-binding promotes Exo1 damage recruitment and suppresses its nuclease activities. DNA Repair (Amsterdam). 35, 106-115 (2015).
- Zhang, F., Shi, J., Chen, S. H., Bian, C., Yu, X. The PIN domain of EXO1 recognizes poly(ADP-ribose) in DNA damage response. Nucleic Acids Research. 43 (22), 10782-10794 (2015).
- Rona, G., et al. PARP1-dependent recruitment of the FBXL10-RNF68-RNF2 ubiquitin ligase to sites of DNA damage controls H2A.Z loading. elife. 7, (2018).
- Young, L. M., et al. TIMELESS forms a complex with PARP1 distinct from its complex with TIPIN and plays a role in the dna damage response. Cell Reports. 13 (3), 451-459 (2015).
- Kong, X., et al. Laser microirradiation to study in vivo cellular responses to simple and complex dna damage. Journal of Visualized Experiments. (131), e56213 (2018).
- Kong, X., et al. Condensin I recruitment to base damage-enriched DNA lesions is modulated by PARP1. PLoS One. 6 (8), 23548 (2011).
- Lan, L., et al. Novel method for site-specific induction of oxidative DNA damage reveals differences in recruitment of repair proteins to heterochromatin and euchromatin. Nucleic Acids Research. 42 (4), 2330-2345 (2014).
- Zerjatke, T., et al. Quantitative cell cycle analysis based on an endogenous all-in-one reporter for cell tracking and classification. Cell Reports. 19 (9), 1953-1966 (2017).
- Ji, Y., Karbaschi, M., Cooke, M. S. Mycoplasma infection of cultured cells induces oxidative stress and attenuates cellular base excision repair activity. Mutation Research. 845, 403054 (2019).
- Sun, G., et al. Mycoplasma pneumoniae infection induces reactive oxygen species and DNA damage in A549 human lung carcinoma cells. Infection and Immunity. 76 (10), 4405-4413 (2008).
- Gassman, N. R., Wilson, S. H. Micro-irradiation tools to visualize base excision repair and single-strand break repair. DNA Repair (Amsterdam). 31, 52-63 (2015).
- Muster, B., Rapp, A., Cardoso, M. C. Systematic analysis of DNA damage induction and DNA repair pathway activation by continuous wave visible light laser micro-irradiation. AIMS Genetics. 4 (1), 47-68 (2017).
- Ikeda, S., et al. Purification and characterization of human NTH1, a homolog of Escherichia coli endonuclease III. Direct identification of Lys-212 as the active nucleophilic residue. Journal of Biological Chemistry. 273 (34), 21585-21593 (1998).
- Rosenquist, T. A., Zharkov, D. O., Grollman, A. P. Cloning and characterization of a mammalian 8-oxoguanine DNA glycosylase. Proceedings of the National Academy of Science U. S. A. 94 (14), 7429-7434 (1997).
- Reid, D. A., et al. Organization and dynamics of the nonhomologous end-joining machinery during DNA double-strand break repair. Proceedings of the National Academy of Science U. S. A. 112 (20), 2575-2584 (2015).
- Taccioli, G. E., et al. Ku80: product of the XRCC5 gene and its role in DNA repair and V(D)J recombination. Science. 265 (5177), 1442-1445 (1994).
- Marsin, S., et al. Role of XRCC1 in the coordination and stimulation of oxidative DNA damage repair initiated by the DNA glycosylase hOGG1. Journal of Biological Chemistry. 278 (45), 44068-44074 (2003).
- Thompson, L. H., Brookman, K. W., Jones, N. J., Allen, S. A., Carrano, A. V. Molecular cloning of the human XRCC1 gene, which corrects defective DNA strand break repair and sister chromatid exchange. Molecular and Cell Biology. 10 (12), 6160-6171 (1990).
- Scharer, O. D.
- Oeck, S., et al. High-throughput evaluation of protein migration and localization after laser micro-irradiation. Science Reports. 9 (1), 3148 (2019).
- Mistrik, M., et al. Cells and stripes: A novel quantitative photo-manipulation technique. Science Reports. 6, 19567 (2016).
- Durand, R. E., Olive, P. L. Cytotoxicity, mutagenicity and dna damage by hoechst 33342. Journal of Histochemistry and Cytochemistry. 30 (2), 111-116 (1982).
- Tobey, R. A., Oishi, N., Crissman, H. A. Cell cycle synchronization: reversible induction of G2 synchrony in cultured rodent and human diploid fibroblasts. Proceedings of the National Academy of Science U. S. A. 87 (13), 5104-5108 (1990).
- Podhorecka, M., Skladanowski, A., Bozko, P. H2AX phosphorylation: Its role in DNA damage response and cancer therapy. Journal of Nucleic Acids. 2010, (2010).
