

Summary
神経系の活動は、しばしば特定の集団内のニューロンからの同期活動電位放電を必要とします。例えば、性腺刺激ホルモン放出ホルモン(GnRH)のパルスはおそらくGnRHニューロンの間で協調活動を必要とする。我々は確実にびまん性に分布GnRHニューロンからの同時電気生理学的記録を得るために私たちの方法論的アプローチを提示する。
Abstract
ゴナドトロピン放出ホルモン(GnRH)は、黄体形成ホルモン(LH)と卵胞刺激ホルモン(FSH)の下垂体放出を制御する小さな神経ペプチドである。これらの性腺刺激ホルモンは生殖機能の調節に不可欠です。 GnRHの含有ニューロンは、彼らは下垂体ポータルシステム(1)にその軸索末端からのGnRHの放出正中隆起に視床下部と、プロジェクト全体にびまん性に分布している。ポータル毛細血管では、GnRHは全身循環に性腺刺激ホルモンの放出を刺激する脳下垂体前葉に移動。 GnRHの放出は連続ではなく、むしろエピソードパルスで発生します。それは、GnRH放出の断続的な方法は、再生(2、3)のために不可欠であることが十分に確立されている。
複数のおそらくのGnRHニューロンの基礎となるのGnRHのパルスの活動の調整。 GnRHニューロンの合計ペプチドの含有量は30%のカスタマーが、おそらく放出可能プールで構成され、そのうち、約1.0 PG /セル(4)です。パルス中のGnRHのレベル(5,6)は、複数のGnRHニューロンは、おそらく神経分泌に関与している示唆している。同様に、LH放出時の視床下部のマルチユニット記録から抽出された単一ユニットの活動は、複数のニューロンの活動の変化(7)を示している。 LHパルスの間に記録された活性を有する電極がGnRHのsomaの複数形や繊維(8)のいずれかと関連付けられます。したがって、少なくともこの活動の一部は、GnRHニューロンから生じます。
メカニズムは視床下部のGnRHニューロンの同期発火にその結果が不明です。 GnRHニューロンの発火の調整メカニズムを解明することは複雑な問題です。最初に、GnRHニューロンは数の比較的少ないです。げっ歯類では、800-2500 GnRHニューロンがあります。それはすべてのGnRHニューロンは、エピソードのGnRH放出に関与していることは明らかではない。また、GnRHニューロンは、びまん性(1)配布されます。これは、焼成の調整の我々の理解を複雑にしており、多くの技術的アプローチが手に負えないてきた。私たちは、活動電位の直接検出のための電流クランプモードでゆるいセルに接続されたレコーディングを最適化し、GnRHニューロンのペアからの同時録画を可能にする記録的なアプローチを開発しました。
Protocol
- 視床下部の脳切片を調製し、インキュベートし、GnRHニューロンのGnRHペプチドのプロモーター(10)の制御下に緑色蛍光タンパク質(GFP)を発現動物で(9)以前に詳細な記録室に転送されます。動物からの脳を除去した後、脳は関心のない地域をなくすためにカミソリの刃を使用してブロックされます。視床下部は、可視化できるよう、削除する領域を決定するために、脳はその背面に配置されます。冠状および矢状スライスの方向の両方に、小脳は削除されます。冠状スライス標本の場合は、皮質の吻側部分が削除されます。このスライスの向きで、皮質の外側の領域は、脳をスライスするためのサポートを提供し、録音室内の脳を保護するために銀線を配置するための脳の十分な領域を提供しています。矢状スライスの準備に、脳の外側の領域を除去し、皮質の吻側部分は、スライス脳のためともレコーディングで脳を保護するために銀線の配置のためのサポートを提供している。ブロッキングの手順では、脳が継続的にガラスピペットを用いてコールド人工脳脊髄液(ACSF)で湿らせている。
- スーパー接着剤に少量の最先端のプラットフォーム上に配置されます。脳は、スパチュラで解除されます。過剰ACSFは、ヘラの表面にキムワイプを持参し、脳から過剰なACSFを描画することによって除去される。脳を穏やかに接着剤の上に前方にスライドさせる。それを確保するために脳に触れたり、その上にプッシュダウンしないすることが重要です。脳自体の重量は、通常、それが安全であることを確認するのに十分です。
- 脳と最先端プラットフォームは、振動ミクロトームに固定され、切断はよく風邪ACSFで満たされている。冷溶液には、両方の企業の脳を、切削作業中に損傷から保護するかもしれない。異なる脳の領域は、品質のスライスを最適化する寒さの度合いが異なります。視床下部スライスの調製のためのACSFの温度は、通常0℃です。
- スライスは徐々にカットする必要があり、各スライスは脳から離れて自由にフロートしてください。スライスは、切削時のカールを開始する場合、ブレードが進んでいる速度が遅くなることがあります。一つは、スライスを真っすぐにするために、小さなペイントブラシを使用することができますが、これはスライスのリスクを傷めるおそれがないので、避けるべきである。ゆっくりとブレードの事前の速度は、ペイントブラシを使用してより優先されます。
- 各スライスは脳から切断されると、それはカッティングチャンバから引き出され、温水浴(約32℃)でのスライスのインキュベーションチャンバー内に配置されます。スライスインキュベーターは、連続して95%O 2および5%CO 2の混合物を用いて酸素が 供給されている。一般に、プリレコーディングスライスのインキュベーションは、30分から2時間の範囲である。
- 前述のようにガラスピペット(MΩ3-6)垂直プーラーを使用して(9)製造される。引っ張りの温度と持続時間は、ピペットプラー目のものの種類、ガラスのいずれかを入力使用と引き手の加熱フィラメントの寿命に基づいて異なります。ピペットの先端の形状はまた、ユーザの好みによって異なります。我々は、0.1 mmの開口部を均一にテーパ状の先端との良好な成功を達成している。ピペットの大きな開口部が小さい開口部は、長時間録音につながることに失敗しながら神経細胞を損傷する傾向がある。
- ピペットは、静電容量を減らすためにSylgardでコーティング。プルピペットチップは軽くSylgard 184は第二ピペットを使用してでコーティングされています。 Sylgardedのヒントは、ヒートガンを使用して熱硬化しています。これらのピペットは、バスの灌流液で満たされている。ピペットチップの可視化を向上させるために、アレクサ- 568少量のピペットを充填する前に、ピペット溶液として使用する浴溶液の部分に追加されます。我々はAlexa - 568の重量を量るのではなく、目でソリューションのピンクをオンにするのに十分使用しないでください。ピペットチップは、テキサスレッドフィルターで可視化することができる。
- 我々は、低抵抗のシールを(説明を参照)を使用して活動電位の検出のための現在のクランプ記録を取得するためのアプローチを開発しました。録音はない適用される現在使用して軸索のインストゥルメントの2B Axoclampと細胞の内因性の静止電位で実行されます。しかしながら、低抵抗のシールに、一つだけ2Bアンプで活動電位を検出できませんでした。信号を高めるために、第2増幅器(AMシステムズ3000)Axoclampの2Bと直列に使用されています。このアプローチは、長期的な録音の緩いシールとユーティリティの使いやすさを持っています。技術的に、人は直接それによって電圧クランプ記録モード(説明を参照)のアプローチで問題を回避して、活動電位を測定することができます。
- 一つは、全細胞記録設定のためのアプローチとして、同様の方法で、同時に二つの以前に選択されたニューロンの表面に2つのピペットを取ります。両方のNEへのアプローチ同時にuronsは独立してペアのそれぞれのニューロンを試みるよりもデュアル録音を得ることに成功している。細胞全体の記録のためのアプローチの間に行われているようにこの時間の間に、電流注入パルスが2Bアンプから渡されます。
- 正圧は、先端の目詰まりを防ぐために両方のピペットで維持する必要があります。デュアル録音のための肯定的な圧力は、最高のチューブの部分の長さによってピペットホルダーに取り付けられた充填されていない3ミリリットル空のシリンジを使用して生成されます。注射器を使用することで、1つは、両方のピペットへの圧力を維持することができます。正圧は、すぐにピペットが録音室の浴溶液に配置されているように適用する必要があります。一つは、順番にピペットで選択されたニューロンに近づく。ニューロンの最大の表面上に直接最初のピペットを配置した後、一つの場所に注射器を残すことによって正の保持して、この位置でピペットを残します。一つは、録音室の上部に戻り、位置に第2の記録ピペットを低下させる。選択されたニューロンは、スライスの異なるレベルで、しばしばです。第一ピペットは、スライスと位置で、より表面的なニューロンの第二ピペット深いですニューロンを介して配置する必要があります。これは、ピペットはスライスを入力すると、スライスは垂直面で若干移動するように、第2ピペットの位置をシフトから二番目ピペットの位置を防ぎます。
- ルースシール(MΩ18〜30)が使用されます。正圧は、それぞれの細胞の表面に小さなくぼみが誘発される。一つは、正圧を解放し、非常にわずかな負圧を印加して、最初のニューロンを封印しようとします。正圧は、注射器を除去することによって解除されます。口による吸引は、チューブの自由端に適用されます。健康的なニューロンの膜は、すぐに正圧のリリースに対応し、ピペットを遵守します。わずかな吸引は、健康な神経細胞の適切な抵抗を持つシールを形成します。
- 膜は非常にきれいなガラスにシール以来一ニューロンと障害が発生した場合を封印しようとすると、ピペットは再使用できません。その代わりに、人はよく灌流の上に離れてスライスの表面からピペットを(と顕微鏡対物)が発生、バスからピペットを削除する、クリーンなピペットに変更し、別のセルを封止するために試みるようにスライスして返します。 。これらの理由から、それは録音のための複数の潜在的な神経細胞を選択することが重要です。選択可能な以下の4月6日録音品質のニューロンとスライスの場合は、デュアル録音を得るための成功は限られている。
- 抵抗に基づいて、緩いシールを確立した後、2Bアンプからの電流注入パルスが終端されます。 2Bのampliferは、3000シリーズのampliferと直列に配置されています。この構成では、2Bのampliferの出力は、3000シリーズのampliferのための入力として機能します。後者ampliferは、データ取得のための信号を提供します。
- シール(ステップ4)とamplifersの再構成(ステップ5)、2番目のニューロンごとに繰り返されます。

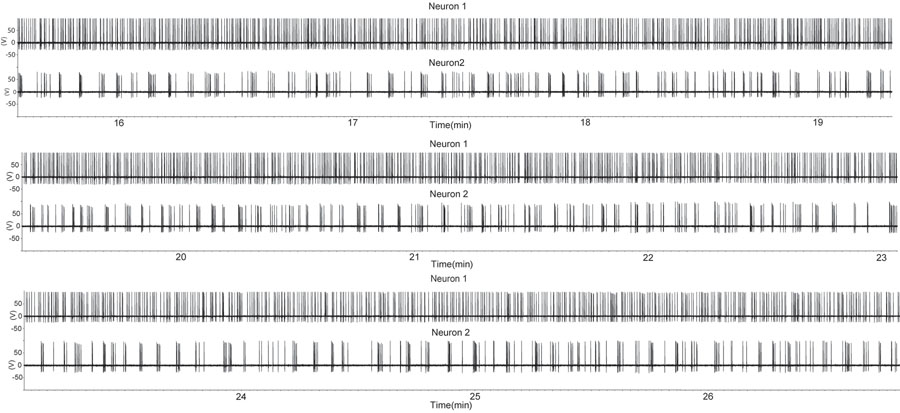
図1の期間は、矢状スライス標本で緩いシール添付の録画設定を使用して、2つのGnRHニューロンの発火を増加した。スライスは、去勢された男性から派生した。それぞれ上向きたわみは活動電位を示している。第二のGnRHニューロンは、断続的なバーストを示す、一方のGnRHのニューロン(上部トレース)がほぼ連続的な発火を示すことに注意してください。単一のGnRHニューロンからの活動のこのパターンは、 生体内でホルモンの分泌(7)中に、マルチユニット記録から抽出された単一のユニットのものと似ています。してくださいここをクリックして図1の拡大バージョンを参照すること。
Discussion
GnRHニューロン(ホルモンの分泌に基づく)を含むいくつかのニューロンへの関心の活動は、時間の時間スケール(5-7)で発生します。したがって、全体のセル構成は、全細胞記録モードでの細胞内メッセンジャーの透析に起因するいくつかの実験的な目標のための最良の選択ではありません。二次的に、全細胞記録は、一般的に年齢の約120日未満の動物に制限されています。加齢とともに、神経細胞膜は高抵抗のシールを達成することが困難になり、堅くように見えます。さらに、一方のパッチがピペットと膜の間の穴を残して、シールを破壊する破裂、高抵抗のシールを取得します。これにより、使用不能の記録と迅速イオン不均衡が原因で死亡するニューロンにつながります。定期的な卵巣周期、および従って、GnRHのパルス発生器の安定的な活動は、その後の人生で(C57BL6女性の年齢の70から10ヶ月、11、12)まで発生していないものが合理的に細胞全体の記録を得ることが予想できるときに年齢を超えて、確実に。最後に、細胞全体の記録は、内部と外部のイオン濃度の内因性比を破壊する。全細胞記録と、どんなイオンの内部濃度は、 ピペット溶液中のイオンの濃度に等しい。 /は、セルの比較的小規模な内因性のボリュームを置き換えるとピペット溶液が比較的大量に迅速に平衡に達するためです。
緩いセル添付のアプローチは、全細胞記録の多くの制限を回避することができます。最初に、低抵抗のシール(MΩ15〜30)を使用することができます。これらは、さらに古い動物から神経細胞で形成することが比較的容易です。第二に、1は破裂膜の密封されたパッチをしません。したがって、緩いセル添付の録音は技術的に細胞全体の録音よりもはるかに簡単です。さらに、細胞膜が無傷であるため、細胞内成分の透析は発生しませんし、内因性のイオン比率が保持されます。一つは、シナプス電流を研究するための緩いセル添付のアプローチを使用することはできませんが、比較的非侵襲的に神経細胞からの長期的な録音に最適です。セルに接続された録音は、またピペット内の任意の標準的な細胞内液を用いて行うことができます。これは、長期の記録が完了すると、膜のパッチを破壊し、細胞内のマーカーでニューロンを標識する付加的な利点を提供しています。
緩い細胞添付アプローチは、 電圧クランプ記録モードで使用されています。しかし、緩いセル接続構成の電圧クランプ記録は、いくつかの手法上の問題があります。最初に、記録された信号は、活動の間接的な尺度です。 (いわゆるアクション現在のように)測定される信号は、 膜 (13) 充電する容量性の電流である。これは非常に重要な方法論の問題です。記録ピペットの容量と抵抗値が記録された信号をフィルタリングすることができます。それは、小さなアクションの電流が原因でヘッドステージの高抵抗に、適切に、ほとんどのアンプで補償することができない、ピペットの容量を充電するに失われる可能性が非常に高いです。これらの信号が検出されないままとする場合、ニューロンの見かけの発火パターンは、真の発火パターンを反映していません。同様に、補正されていないピペットとシールの抵抗は、このようなアクションの流れが表現されている場合など、変化(13)中の測定で大きな誤差を引き起こす。一部のアンプは、信号の損失を制限するピペットやシール、のための静電容量と抵抗"補償"を提供していますが、ほとんどのアンプの高抵抗ヘッドのステージは、最適な補償を妨げる。第二に、人工的な状況は、セルに課されている。電圧クランプモードでは、細胞膜の周辺には固定電位、これらの研究では、0 mVに保持されます。これは、細胞膜に適用される電流は存在しないという意味ではありません。電圧クランプで測定された信号は、実際には固定電位を維持するために膜に印加される電流の量です。したがって、この印加電流は、細胞の活動を変更することができます。
GnRHのシステムでのデュアル録音は特にGnRHニューロンとそれらの拡散分布の限られた数のために挑戦しています。デュアルレコーディングを成功させるには、マニピュレータは非常に安定している必要があります。電極のわずかな動きがニューロンから滑り落ち、録音を終了するにはピペットを引き起こす可能性があります。また、セル上にピペットの動きは(例えば、再配置の動きを補償するために)発火パターンを変更することができます。いくつかのイオンチャネルは、N型カルシウムチャネルは、機械的に敏感です:膜のストレッチは、全細胞と細胞接着型の記録の構成(14)の両方で反復的な活動を引き起こす。最後に、マニピュレータシステムは、非常に細かいと滑らかな動きに対応していなければなりません。デュアル録音で、前述のように、人は同じ時間と封印しようとする試みでいた2つの選択されたニューロンの表面に2つのピペットを取ります一つのニューロン。成功した場合は、一方の試みは、番目のセルをシールする。一般に、人はすべての試行で高品質の録音を密封していることを期待することはできません。これは、しかし、二重録音で、特定の問題を作成します。一方が、ニューロンと成功するが、2番目で失敗した場合、一つはピペットを変更し、別のセルを試みる必要があります。したがって、人は成功した密閉ニューロンを中断することなく、よく灌流(ピペットを変更する)の上に顕微鏡の浸対物レンズとピペットの両方を移動できるようにする必要があります。
デュアル録音のための緩いセルに接続されたアプローチの私たちの開発と利用は、GnRHニューロンを研究する上で主要な技術的な進歩です。それは、メカニズムが拍動性ホルモンの分泌の結果という協調活動の根底にあるものの重大な問題の文脈で前方にフィールドを移動するのに役立つ有用な結果を生成する可能性があります。
Acknowledgments
私はロナルドL.カラブレーゼ、ディーターイエガー(エモリー大学)および有用な技術的な議論のための病棟Yuhas(アクソンインスツルメンツ)に感謝しています。
References
- Silverman, A. J. The gonadotropin releasing-hormone (GnRH) neuronal systems: immunocytochemistry. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Freeman, M. E. The neuroendocrine control of the ovarian cycle of the rat. In: The physiology of reproduction. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Belchetz, P. E., Plant, T. M., Nakai, Y., Keogh, E. J., Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 202, 631-633 (1978).
- Maurer, J. A., Wray, S. Luteinizing hormone-releasing hormone quantified in tissues and slice explant cultures of postnatal rat hypothalami. Endocrinology. 140, 791-799 (1999).
- Harris, G. C., Levine, J. E. Pubertal acceleration of pulsatile gonadotropin-releasing hormone release in male rats as revealed by microdialysis. Endocrinology. 14, 163-171 (2003).
- Sisk, C. L., Richardson, H. N., Chappell, P. E., Levine, J. E. In vivo gonadotropin-releasing hormone secretion in female rats during peripubertal development and on proestrus. Endocrinology. 142, 2929-2936 (2001).
- Cardenas, H., Ordog, T., O'Byrne, K. T., Knobil, E. Single unit components of the hypothalamic multiunit electrical activity associated with the central signal generator that directs the pulsatile secretion of gonadotropic hormones. Proc Natl Acad Sci U S A. 90, 9630-9634 (1993).
- Silverman, A. J., Wilson, R., Kesner, J. S., Knobil, E. Hypothalamic localization of multiunit electrical activity associated with pulsatile LH release in the rhesus monkey. Neuroendocrinology. 44, 168-171 (1986).
- Roberts, C. B., O'Boyle, M. P., Suter, K. J. Dendrites determine the contribution of after depolarization potentials (ADPs) to generation of repetitive action potentials in hypothalamic gonadotropin releasing-hormone (GnRH) neurons. J Comput Neurosci. 26, 39-53 (2009).
- Spergel, D. J., Kruth, U., Hanley, D. F., Sprengel, R., Seeburg, P. H. GABA- and glutamate-activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 19, 2037-2050 (1999).
- Nelson, J. F., Felicio, L. S., Randall, P. K., Sims, C., Finch, C. E. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 27, 327-339 (1982).
- Gee, D. M., Flurkey, K., Finch, C. E. Aging and the regulation of luteinizing hormone in C57BL/6J mice: impaired elevations after ovariectomy and spontaneous elevations at advanced ages. Biol Reprod. 28, 598-607 (1983).
- Anson, B. D., Roberts, W. M. Loose-patch voltage-clamp technique. Neuromethods, Patch-clamp analysis: Advanced techniques. Walz, W., Boulton, A. A., Baker, G. B. 35, Humana Press. Place Unknown. 265-286 (2002).
- Calabrese, B., Tabarean, T., Juranka, P., Morris, C. E. Mechanosensitivity of N-type calcium channel currents. Biophys J. 83, 2560-2574 (2002).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Rapid killing of single neurons by irradiation of intracellularly injected dye. Science. 206, 702-704 (1979).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Lucifer Yellow slows voltage-gated Na+ current inactivation in a light-dependent manner in mice. J Physiol. 550, 159-167 (2003).

{kind=link}