

Summary
A robust way to study neuronal avalanches, i.e. scale-invariant spatio-temporal activity bursts, indicative of critical state dynamics in cortex. Avalanches emerge spontaneously in developing superficial layers of cultured cortex which allows for long-term measurements of the activity with planar integrated multi-electrode arrays (MEA) under precisely controlled conditions.
Abstract
The cortex is spontaneously active, even in the absence of any particular input or motor output. During development, this activity is important for the migration and differentiation of cortex cell types and the formation of neuronal connections1. In the mature animal, ongoing activity reflects the past and the present state of an animal into which sensory stimuli are seamlessly integrated to compute future actions. Thus, a clear understanding of the organization of ongoing i.e. spontaneous activity is a prerequisite to understand cortex function.
Numerous recording techniques revealed that ongoing activity in cortex is comprised of many neurons whose individual activities transiently sum to larger events that can be detected in the local field potential (LFP) with extracellular microelectrodes, or in the electroencephalogram (EEG), the magnetoencephalogram (MEG), and the BOLD signal from functional magnetic resonance imaging (fMRI). The LFP is currently the method of choice when studying neuronal population activity with high temporal and spatial resolution at the mesoscopic scale (several thousands of neurons). At the extracellular microelectrode, locally synchronized activities of spatially neighbored neurons result in rapid deflections in the LFP up to several hundreds of microvolts. When using an array of microelectrodes, the organizations of such deflections can be conveniently monitored in space and time.
Neuronal avalanches describe the scale-invariant spatiotemporal organization of ongoing neuronal activity in the brain2,3. They are specific to the superficial layers of cortex as established in vitro4,5, in vivo in the anesthetized rat 6, and in the awake monkey7. Importantly, both theoretical and empirical studies2,8-10 suggest that neuronal avalanches indicate an exquisitely balanced critical state dynamics of cortex that optimizes information transfer and information processing.
In order to study the mechanisms of neuronal avalanche development, maintenance, and regulation, in vitro preparations are highly beneficial, as they allow for stable recordings of avalanche activity under precisely controlled conditions. The current protocol describes how to study neuronal avalanches in vitro by taking advantage of superficial layer development in organotypic cortex cultures, i.e. slice cultures, grown on planar, integrated microelectrode arrays (MEA; see also 11-14).
Protocol
1. Sterile, Sealable Glass Chamber with MEA for Long-term Recordings
- Threaded Glass cylinders with Teflon-plastic cap (Ace Glass), required for secure and tight culture chamber closure, are cut (Aceglass) approximately 2 mm from bottom of the thread (Fig. 1A, B). Clean glass rings by rinsing with water (3x) and boiling for 5 min in 200 proof ethyl-alcohol, let dry.
- Aliquot silicon solution required to attach glass rings to MEA surface. Mix 15 ml of parts A & B of Sylgard 184 Silicone Elastomer Kit thoroughly, let sit for 15 min to remove air bubbles, store in 1 ml aliquots at -20 °C.
- Glue glass ring to the MEA (8x8 grid w/internal ground electrode, 30 μm electrode diameter, 200/100 μm inter-electrode distance for rat/mouse) (Fig. 1A, B). Take up 1 ml of silicon (23 °C) in syringe with small gauge needle. Apply silicone to unpolished cut surface of glass ring, center glass ring on MEA, apply an additional layer of silicon around the outside of the ring for a stronger seal, let cure for 1 - 2 hrs at ~60 °C on hot plate.
- Sterilize MEA chamber and chamber caps in a laminar flow hood by 3x rinse in deionized water followed by 70% alcohol (3 x; for last rinse let sit for 10 min in alcohol) followed by 10 min exposure of chamber and cap interior to UV light. Autoclave MEA chamber (120 °C; wet; 45 min) and let dry.
- Coat MEA surface inside culture chamber with poly-D-lysine. For new MEAs, which are rather lipophilic, coat by repeated droplet aspiration of the solution from the electrode grid. For used MEAs, cover chamber bottom with solution, aspirate excess liquid, allow to evaporate under sterile conditions inside laminar hood. Attach cap to seal MEA chamber for storage and future use.
2. Ingredients Required for the Preparation and Growth of Organotypic Cultures
- Dissolve sterile agar in 0.9% NaCl, pour into sterile petri dish (Falcon, 100 x15; ˜5 mm level), let cool, and sterile wrap with Parafilm for storage. Cut 20 x 10 x 5 mm blocks from solid agar for use.
- Store super glue, e.g. Devcon Super Glue II, with the packaging wiped down with 70% EtOH before opening, inside the laminar flow hood to preserve sterility.
- Prepare 50% D-Glucose (SIGMA ultra, G7528) by adding 40 g of glucose to 40 ml of de-ionized culture water (Sigma). Store in 2 ml aliquots at -20°C.
- Add 4 ml of 50% D-glucose to 500 ml of Gey's Balanced Salt solution, and chill to slush (mixture of liquid/ice crystals) in freezer before use.
- Reconstitute chicken plasma in 5 ml de-ionized culture water (shake gentle, avoid formation of bubbles), let solution settle for 5 – 10 min, gently swirl, and decantate the clear content into a sterile Petri dish. Sterile-filter (0.22 μm pore filter; protein) plasma solution, aliquot 350 μl into cryotubes (NuncTM), store at -20°C.
- Reconstitute thrombin from bovine plasma accordingly, sterile-filter (0.22 μm pore filter), aliquot 40 μl into cryotubes (NuncTM ), store at -20°C. For working solution, dilute 40 μl of the thrombin solution in 375 μl of Gey’s balanced salt solution w/D-Glucose.
- Prepare 400 ml of culture medium by mixing 100 ml of horse serum, 200 ml Basal Medium Eagle, 100 ml Hank’s Buffered Saline Solution to which 4 ml of 50% glucose and 2 ml of 200 mM L-glutamine are added. Can be stored 4 – 8 weeks in 100 ml PYREX bottles at 4°C.
- Prepare mitosis inhibitor by mixing 0.3 mM uridine, 0.3 mM ARA-C cytosine-β-D-arabinofuranoside, and 0.3 mM 5-fluoro-2’-deoxyuridine, sterile filter, aliquot 200 μl and store at -20°C for 6 – 12 months.
3. Cortex and Ventral Tegmental Area (VTA) Tissue Dissection (time: < 1hr)
- Procedure yields cortex and VTA tissue sections for ˜12 co-cultures from rats or mice, and is prepared inside a laminar flow hood under sterile conditions. Total time for tissue collection should be than 1 hr.
- Take healthy, well nourished pups (litter size ˜10; presence of an abdominal ‘milk spot’) at 1 – 2 postnatal days (PND). Hold a pup gently by the snout, allow it to hang freely, and quickly decapitate at the base of the neck with sharp scissors.
- For brain removal, remove skin (two lateral scissor cuts), cut skull open with fine eye scissor (1 sagital midline cut; 1 coronal cut at cortex/cerebellum junction). Flip back all 4 skull flaps. With a sharpened spatula, cut frontally through the olfactory bulb, advance spatula caudally underneath the brain. Gently lift the brain out of the skull and let it slide into sterile, chilled, Gey’s solution for rapid cooling and temporary storage. Repeat the steps 3.2 to 3.3 for 2 more brains (total time: <20 min).
- To obtain VTA tissue, transfer the brains onto a sterile, dry Petri dish using a small spatula. Further remove excess fluid by gently sliding each brain about 1 cm sideways. Remove brain stem by a coronal, vertical cut at the level of the cerebellum using a razor blade.
- Glue agar block on mounting disc for mechanical stabilization of brains during slicing procedure. Place a thin line of superglue a few millimeters in front of the agar block on the disk (avoid glue touching the agar).
- Using a small spatula, transfer and mount each brain, frontal pole down. Ensure that frontal poles are glued to mounting disc and that ventral sides touch the agar without any glue residues in order to achieve proper mechanical stabilization during cutting and easy lift-off of cut slices.
- Carefully submerge and secure mounting disk with brain assembly in a vibratome tray (e.g. Leica VT1000) filled with sterile, chilled Gey’s solution. With a carefully cleaned razor blade (90% EtOH), cut coronal slices of the midbrain at highest vibration frequency and relatively low forward speed at a thickness of 400 500 μm. Using an inverted Pasteur pipette with suction bulb, transfer and collect slices containing the VTA in 35 x 10 mm Petri dishes filled with sterile, chilled Gey’s solution (Fig. 1C; see also coronal plate 18 – 20 at E22 in15).
- For cortex sections, repeat steps 3.2 – 3.6, but apply vertical cut between cortex and cerebellum, and mount forebrains with frontal pole up. About 3 coronal slices (350 μm thickness) starting at the level of the striatum are collected for future cortex dissection.
- Using a micro-knife made of broken razor blades, dissect ˜2 mm wide coronal section of frontal cortex and midbrain areas containing the VTA (Fig. 1C) under a stereomicroscope. Collect tissue sections separately in small dishes (e.g. chamber slides) filled with chilled Gey’s solution.
4. Mounting Cortex and VTA Tissue Slices on the MEA (Time: <1 hr)
- Position MEA at room temperature under a stereo microscope with the electrode array in focus. Center a 25 μl droplet of plasma on the clean, dust-free, and sterile electrode array matrix. Using small spatulae, carefully slide a cortex and VTA section into the plasma droplet.
- Place MEA on cooling plate, refocus view, let chill for ˜15 s, then add 25 μl of thrombin into the plasma droplet. Using the thrombin pipette tip, carefully spread the plasma/thrombin mixture with small circular movements across the MEA. Do not touch the brittle electrode array directly. Gently position the cortex on the array with dorsal border along the second electrode row of the array. This way, the developing superficial layers will eventually cover the reminder of the array. The VTA is placed adjacent to the ventral border of the cortex section (Fig. 1D).
- Cap and loosely close the MEA chamber to retain high humidity while the MEA/culture assembly sits for ˜5 min inside the hood at room temperature to allow for plasma/thrombin coagulation. Meanwhile, repeat step 4.1 – 4.3 for 3 more cultures.
- Carefully add 600 μl of culture medium in small droplets to the culture chamber using a 1 cc syringe with 25 x 5/8 needle.
- Tightly close the MEA chamber and place MEA/culture assembly on the rocking storage tray inside the incubator (Fig. 1B). To speed up the procedure, 3 – 4 MEAs can be assembled in overlapping sequences. Assembly time for 12 MEAs should be <1hr.
- After 2 days in vitro (DIV), add 10 μl of mitosis inhibitor. Refresh culture media by 60% at 4 DIV and every 4 days thereafter.
5. Electrophysiological Recording and Stimulus Generation
- To establish the relationship between significant deflections in the local field potential (LFP) and the tendency of neurons to fire action potentials, after ˜1 week 5,6 record spontaneous activity at 24 kHz for ˜10 min from each electrode of the MEA (Hardware: MEA1060 w/ blanking circuit, x1200 gain, 12 bit A/D, range 0–4096 mV, MultiChannel Systems; Software: MC_Rack). Ground is provided either through the internal ground electrode, or externally by adding a Ag/AgCl-half cell.
- Separate the LFP with a band pass filter of 1 – 200 Hz from extracellular spike activity (band pass 300 – 3,000 Hz). Spike activity can be further classified into single and multi-unit activity using off-line spike sorters (e.g. Plexon Inc.). Calculate spike triggered averages for each electrode. For cortex cultures, most averages will identify negative LFP deflections (nLFP) as the preferred time of neuronal spiking in the culture.
- Calculate for each electrode a threshold of -3 standard deviations of the noise (SD) from the LFP traces (Fig. 2), determine peak times and amplitudes of nLFPs that cross threshold (Fig. 2B, C). Chose a time bin Δt (e.g. between 2 – 8 ms) and identify spatiotemporal nLFP clusters on the array by concatenating nLFPs from all electrode that are in the same of successive time bines of length Δt (Fig. 2D; for details see 2,4,5).
- In order to identify neuronal avalanches, calculate the size of each nLFP cluster, e.g. number of active electrodes or sum of nLFP amplitudes, construct a size histogram, and plot in double-logarithmic coordinates. For neuronal avalanches, the size distribution follows a power law approximated by a straight line in double-logarithmic coordinates2 (Fig. 2E,F). See 16 for statistical tests on power laws.
- Elicit evoked responses in the tissue by selecting an electrode through which current-controlled stimuli with amplitude S are applied (Stimulus generator STG 1008, Multichannel Systems). To reduce electrode damage, use a range limited, charge-neutral stimulation of single shocks with bipolar square waveform: 50 μs with amplitude -S followed by 100 μs with amplitude +S/2 and S between 10 – 200 μA. See owner’s manual for further details.
- To record the dynamic range9, record stimulus recorded LFP responses at 4 kHz sampling rate on all electrodes following 500 ms after stimulation. Use blanking circuitry (Multi-channel systems) which disconnects the head stage amplifiers during stimulation to reduce stimulus artifacts and to prevent pre-amplifier saturation.
6. Representative Results:
With new MEAs about 8 – 9 out of 10 cultures will survive for many weeks. Most of our long-term recordings take place inside the incubator in culture medium , which allows us to follow the development of individual cultures over the course of many weeks5. Based on our experiments, LFP recordings can be reliably obtained with MEAs used for more than 100 culture days. In contrast, extracellular spike activity is more reliably measured with relatively new MEAs (<40 culture days). In a typical experiment, we transfer an MEA from the storage tray (Fig. 1B, right) to the tray with the head stage attached (Fig. 1B, left) keeping the culture chamber sealed. For the cortex5, cortex-VTA co-cultures6, as well as in the anesthetized rat6 and the awake monkey in vivo 7, neuronal firing during neuronal avalanches in superficial layers occurs predominantly close to the peak negative deflection of the LFP (nLFP). Thus, the spatiotemporal organization of locally synchronized neuronal groups can be estimated by measuring the occurrence of nLFPs in space and time on the array 17.
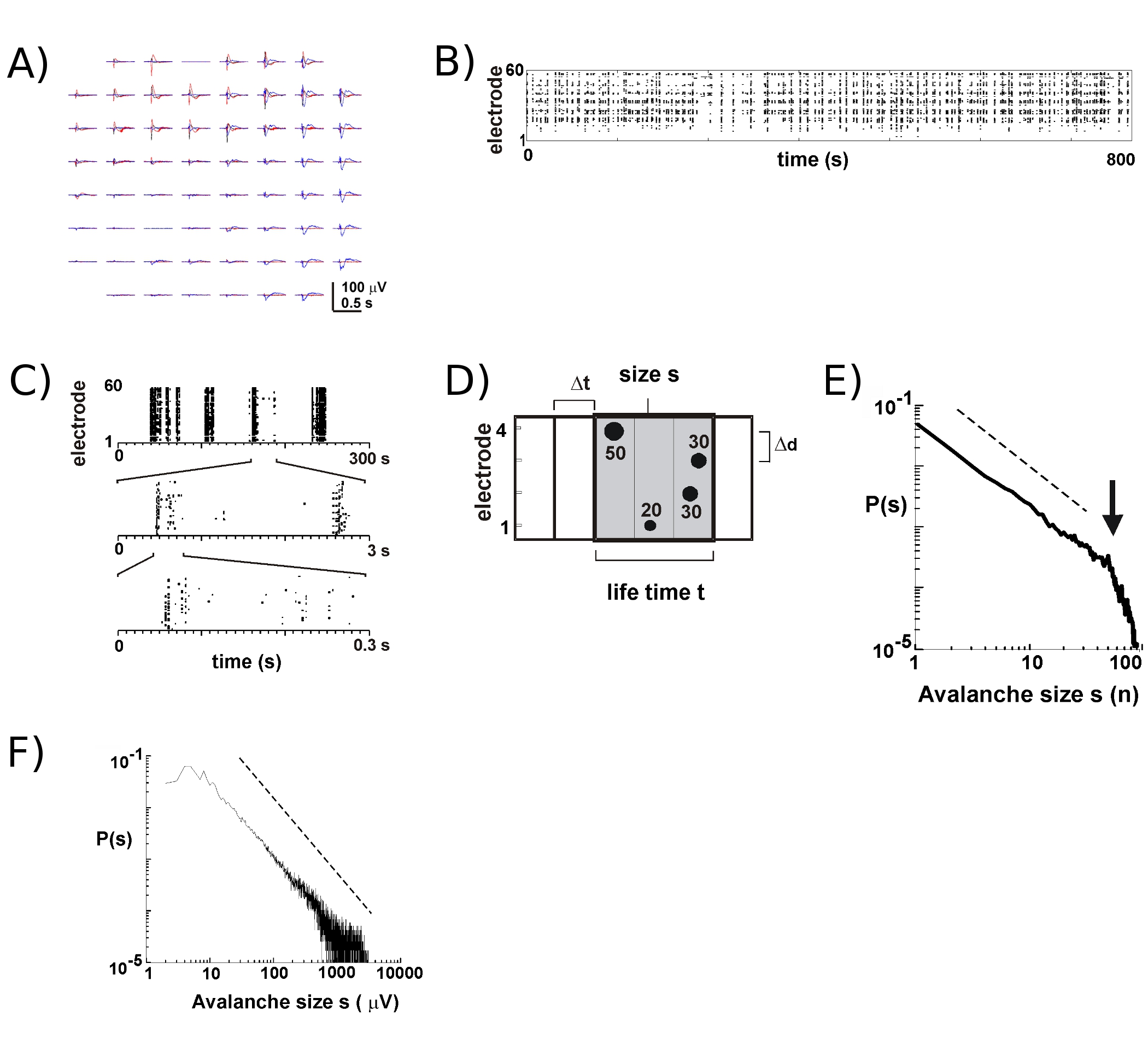
Activity on the MEA tends to emerge in temporal clusters, such that activity at one electrode is accompanied by activity at other sites. Typical waveforms of the LFP during such activity periods are shown in figure 2A, by over plotting 3 clusters occurring several seconds apart. For each cluster, negative field deflections can be seen at several electrodes within a window of 1 s. When extracting nLFP peaks that cross a threshold of multiple negative SD, the activity in the form of nLFP peak times is conveniently visualized in a raster in which ‘columns’ of dots represent near coincident nLFPs at various electrodes (Fig. 2B). The spatiotemporal organization of this activity is rather complex; ‘columns’, which appear more or less homogenous at low temporal resolution, are composed of separate clusters at higher temporal resolution and so on (Fig. 2C). In fact, the emergence of spatiotemporal nLFP clusters is highly organized in cortical networks. More specifically, the organization is scale-invariant for neuronal avalanches. This is demonstrated by calculating the probability of cluster sizes at a given temporal resolution Δt. Here, clusters are composed of nLFPs that occur in the same or successive time bins (Fig. 2D). When the size of such a cluster is expressed in total number of nLFPs per cluster, or integrated nLFP amplitudes per cluster, the cluster size distributions reveals a power law, whose slope has been shown to be -1.5 2,4,5,7 (Fig. 2E,F). Note that this distribution identifies a scale-invariant ordering of cluster sizes that is the ratio of sizes s to k x s , where k is a constant factor, is k-1.5 , which is independent of s. This power law organization is independent of the size of the array 2, the temporal resolution Δt 2, and the threshold used to identify significant nLFP deflections 7. Because nLFP amplitude scales with neuronal group size 7, the scale-invariant organization of nLFPs reflects a scale-invariant, i.e. fractal, ordering of locally synchronized neuronal groups that include all sizes.

Figure 1. (A) Side and top views of the MEA with threaded glass ring mounted, and corresponding cap. (B) Inside view of the incubator. Left: headstage mount allowing for recording from a single culture under incubator condition. Right: Tray holding numerous MEAs for culture growth. Side wheels: stepping motor controlled rocking device for alternating submerged and atmosphere-exposed phase required for culture growth. (C) Schematic drawing for the coronal rat slices used for cortex-VTA co-cultures. Cortex sections (left) and midbrain sections (middle, right) containing the VTA ventral tegmental area (vta; grey) are obtained by cutting along the broken lines. Ctx: cortex; wm: white matter; cpu: striatum; vta: pons: pontine area. See also corresponding coronal plates 8, 18, and 20 by 15 . (D) Placement and growth of a single cortex-VTA co-cultures on the MEA and its development over the first 9 DIV in culture. Note the flattening of the culture and its progressive expansion on the array. Reflective tissue parts indicate degenerated cells and tissue debris. Healthy tissue is opaque and greyish under transillumination with visible light.

Figure 2. Neuronal avalanches in cortical organotypic cultures. (A) Overplot of three period of spontaneous activity on the array, separated by several seconds. Note that each activity period consists of negative LFP deflections on many electrodes on the array (each color labels one activity period). (B) Negative peak times of nLFPs from each electrode are assembled into a raster of activity. ‘Column’-like structures indicated periods of near synchronous activity. (C) Note that columns that appear highly synchronized at one time scale consist of multiple columns at the higher temporal scales (3 temporal scales shown). (D) Schematic representation of the neuronal avalanche algorithm. On a 2 x 2 electrode array peak time and amplitude of negative LFP deflections (nLFP) crossing a threshold of –x SD of the noise are identified. Spatiotemporal organization of nLFPs is clustered into successively active time bins of width Δt. The size of a cluster is identified by either the number of active sites, i.e. electrodes with nLFP (s = 4), or the integrated sum of nLFP amplitudes (s = 130 μV). The life time is measured in multiples of Δt. (E, F) Power law in cluster size distribution identifies clusters as neuronal avalanches. Note that the choice of a particular interelectrode distances Δd for the array (here 200 μm) introduces a particular Δt at which the dynamics should be observed. More specifically, the ratio in which Δd/Δt approximates the average propagation velocity in the network, at which the slope α of the power law is approximates -1.5 for neuronal avalanches 2,4,5. Please click here to see a larger version of figure 2.
Discussion
1. Technical issues:
- Sterile technique. The preparation of the MEAs and culture preparation are all performed inside a laminar flow hood under sterile conditions. Antibiotics, which affect neuronal activity, are not used at any time during the preparation and culture process.
- Plasma/thrombin coagulation and tissue adherence on the MEA. Tissue survival on the MEA requires a careful balance between the time required for plasma/thrombin coagulation and the time of tissue exposure to the atmosphere. A short coagulation time risks premature detachment of sections from the MEA, while prolonged exposure to atmosphere triggers tissue degeneration. Because the strength of the thrombin solution determines the speed of the coagulation process, it is a very important parameter for successfully attaching, healthy cultures to the MEA surface. We obtain best results using 1000 units (1KU; 1 NIH unit = 0.324 ± 0.073 mg). Importantly, incomplete mixing of the plasma/thrombin solution results in spatially heterogeneous coagulation promoting plasma-ruptures during culturing. These plasma ‘holes’ severely affect culture health and detach the culture from microelectrodes, thereby compromising the quality of the electrophysiological recording. Working with cooling plates during the MEA/tissue assembling slows the coagulation and allows for homogenous mixing of the plasma/thrombin solution and proper positioning of the tissue sections.
Similarly, by adding medium in single droplets to the culture chamber to submerge the culture after only 5 min of coagulation, greatly reduces the risk of tissue detachments due to surface tensions. A successful culture will flatten on the MEA and slightly expand showing healthy growth for weeks, without any major signs of incomplete tissue-MEA contact or tissue degeneration (e.g. Fig. 1D). - Tissue dissection. Micro knifes have greatly improved our dissection process. We use double edge razor blades (Fine Science Tools – breakable scalpel blades – 100050-00), from which we split ˜2 mm wide ‘blades’ using pliers, and hold them with a scalpel holder. Tissue samples dissected from the coronal slice using a smooth, vertical firm motion of the blade thereby greatly reducing mechanical stress due to pulling of the tissue overall improving culture health.
- Tissue cooling. Proper cooling of the slices and tissue sections during the preparation is essential for culture success. We use custom-made cold plates built of Peltier elements attached underneath to a metal disc. The heat produced by the Peltier element is removed through cold water perfusion. This greatly reduces preparation time and standardizes cooling during each stage of the preparation (Dold Labs & Engineering. 131 Plantation Dr. Seguin, TX 78155; (830) 560-1471)).
- Incubator condition. A custom-made incubator with precise internal rocking conditions has been crucial to our success in cultivating slices on MEAs. Based on an original in-house design by Multichannelsystems (currently not available commercially), the internal rocking device consists of trays that are attached to two large side wheels. Stepping motors and computerized controls allow for a precise rocking trajectory (rocking angle, rocking velocity, and intermittent pauses). Ultimately, slice cultures need to be exposed to atmosphere and culture medium in slow alternation. The traditional approach is to place slice cultures into narrow tubes that slowly rotate along there longest axis. Here, slow rotation does not produce mechanical stress due to the rotation itself, and the rotation velocity is high enough to obtain an optimal ‘feeding/breathing’-cycle of about 5 – 10 min duration. The more compact interior of the MEA chamber, its total volume of ˜2 cc, and the small culture medium volume required for medium conditioning by the tissue itself, poses a significant challenge. By rocking the MEA between ±˜70° angle (cycle time: ˜200 s), slowing down the rocking velocity as the culture transitions between the liquid phase and atmosphere, and halting the rocking at the extreme angles for extended exposure to the atmosphere has been essential for culture survival.
2. Developmental age of cortex cultures to study neuronal avalanches
Acute slices from rat cortex are commonly taken at PND 0 – 1 and cultured for many weeks on the MEA. Early studies have clearly demonstrated that single cortex slice cultures, after several weeks in vitro, maintain a layered structure with identifiable cell types that can be easily compared to in vivo cell classes 18,18-21. The layered organization in this in vitro system has been conveniently used to study thalamic innervation of cortex during development 22-24, as well as for driving subcortical structures such as the striatum 25,26. In fact, the specificity in the formation of neuronal connections within and across brain regions allows for the construction of complex in vitro systems that recapture numerous detailed projection systems, e.g. that of the cortex-basal ganglia circuitry 27-30.
After 4 – 6 weeks in vitro, single cortex slices 31 and cortex slices co-cultured with striatum 26 or thalamus 32 show spontaneous up- and down states typically found in vivo in the urethane anesthetized rat 33. The fine temporal organization of these up-states bears the hallmark of nested θ- and γ-oscillations indicative of an electrophysiologically mature network of pyramidal neurons and fast-spiking GABAergic interneurons 31. Importantly, in the absence of dopamine D2-receptor stimulation, maturation of parvalbumin-positive cortical interneurons is delayed by about 2 weeks in cortex slice cultures 34. In line with these findings, the developmental time course of nested θ-, β- and γ-oscillations is matched to that in vivo when cortex slices are co-cultured with the ventral tegmental area (VTA), which contains dopaminergic neurons projecting to the cortex 6.
These studies indicate that when studying neuronal avalanches, which crucially depend on mature fast GABAergic inhibition and are located in superficial layers of cortex 2,4, great care has to be taken to assure proper maturation of the cortical tissue. While neuronal avalanches arise in single cortex cultures over the time course of 2 – 5 weeks 4, when requiring a developmental time course that is matched to the in vivo development, cortex slices need an appropriate dopamine receptor stimulation, e.g. by co-culturing cortex slices with the VTA 6.
Disclosures
No conflicts of interest declared.
Acknowledgments
This study was funded by the Division of Intramural Research Program (DIRP) of the National Institute of Mental Health, National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}