

Summary
Генетические исследования у дрожжей могут быть использованы для исследования молекулярных и клеточных функций генов человека в клеточный метаболизм ДНК. Описаны методы генетической характеристике человека
Abstract
Понимание роли человеческих белков репарации ДНК в генетических путей является сложной задачей для многих исследователей. Генетические исследования млекопитающих системы были ограничены из-за отсутствия доступных инструментов, включая генетический мутант определены клеточных линий, систем регулирования выражения, а также соответствующие выбор маркеров. Чтобы обойти эти трудности, модель генетических систем в низших эукариот стали привлекательным выбором для изучения функционально сохраняется репарации ДНК белков и путей. Мы разработали модель системы дрожжей учиться плохо определенными генетическими функциями синдром Вернера геликазы-нуклеазы (
Protocol
1. Штаммов дрожжей
Штаммы с диким типом SGS1 TOP3 (WT; W303-1A, генотип, ВСУ ade2-1 CANL-100 his3-11, 15 leu2-3, 112 TRPL-л URA3-1) [2], sgs1 мутант (W1292-3C ; генотип MAT SUP4-о:: URA3 sgs1-25 ade2-1 can1-100 his3-11, 15 leu2-3, 112 TRP1-1 URA3-1 rad5-535) и sgs1 топ3 мутант (W1058-11C, генотип, ВСУ SUP4-о:: URA3 sgs1-25 топ3-2:: HIS3 ade2-1 can1-100 his3-11, 15 leu2-3, 112 TRP1-1 URA3-1 rad5-535) были охарактеризованы [3] и были любезно предоставлены д-р Родни Ротштейн (Колумбийский университет).
2. Плазмиды конструкций ДНК
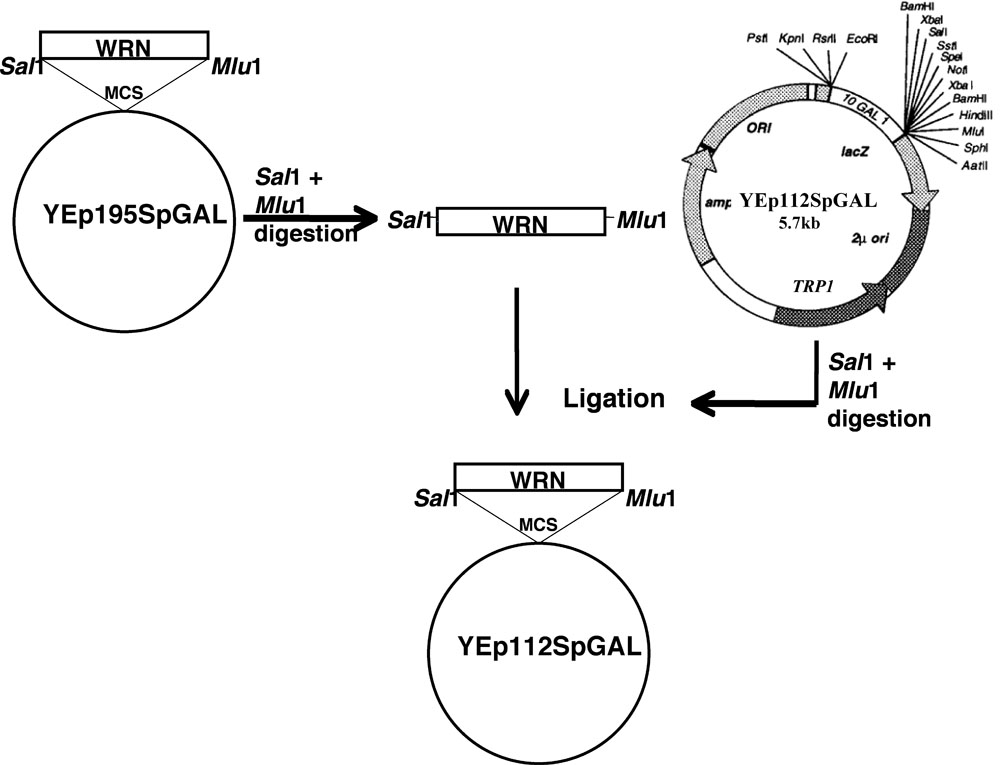
Ген WRN была клонирована в YEp112SpGAL по схеме, как описано.
- Сайт-направленный мутации WRN (E84A экзонуклеазная мертвых, K577M геликазы мертвых, R834C и K1016A) в плазмиды YEp195SpGAL-WRN [4] были построены с использованием мутагенных праймеров и стандартный протокол от QuickChange II XL сайт-направленного мутагенеза комплект (Stratagene) по Lofstrand лаборатории (Gaithersburg, MD).
- Фрагменты ДНК кодирования WRN и связанные с ней варианты были гель очищенный от соответствующих YEp195SpGAL-WRN конструкций после двойного переваривания Сал я и MLU I.
- Гель очищенные фрагменты затем были клонированы в Сал я MLU сайтов, которые я векторных YEp112SpGAL, 2 мкм мульти-копии плазмиды, содержащей TRP1 селективного маркера для построения YEp112SpGAL WRN или WRN варианты под контролем гал-индуцируемых промоторов, как показано на рисунке 1 .
- Полученные конструкции были названы YEp112SpGAL-WRN, YEp112SpGAL-WRN E84A, YEp112SpGAL-WRN K577M, YEp112SpGAL-WRN K1016A и YEp112SpGAL-WRN R834C.
3. Преобразование
Дрожжи культуры выращивали с использованием стандартных протоколов и преобразования проводились с использованием литий ацетат основе протокола Gietz и др. [5]. Короче говоря, следующие шаги были выполнены.
- Культуры выше штаммы выращивали в течение ночи в 5 мл YPD (дрожжевой экстракт-пептон-глюкоза) среде. Культуры reinoculated в 50 мл среды YPD и им было позволено расти, пока OD 600 в 1,0 - 1,2 было достигнуто.
- Культуры собирали, промывали 20 мл воды, и ресуспендировали в 1 мл 100 мМ ацетат лития (LiAc). Клетки инкубировали при 30 ° С в течение 10 мин.
- Сотовые осадок ресуспендировали в 100 мМ LiAc до конечного объема 500 мкл, что достаточно сделать десять преобразований. 50 мкл клеточной суспензии аликвоты и осадок клеток была собрана с помощью центрифугирования.
- Чтобы осадок клеток была добавлена в следующем порядке: 250 мкл 50% полиэтиленгликоля (ПЭГ), 36 мкл 1 М LiAc, 25 мкл 2 мг / мл оцДНК (ДНК спермы лосося), 5 мл ДНК плазмиды (100 нг 1000 нг) и 50 мкл воды. Сотовые осадок перемешивают в течение 1 мин.
- Затем клетки инкубировали при 30 ° С в течение 30 мин, а затем путем теплового шока при 42 ° С в течение 20 мин.
- Клетки собирали и ресуспендировали в 1 мл воды. Поскольку плазмиды YEp112SpGAL содержит ГТО в качестве селективного маркера, все мутантные штаммы укрывательство построить бы ГТО опытным. Таким образом, трансформанты были отобраны покрытия подвеска SC (Синтетические полное) глюкозы средах без Trp и инкубирования пластин при 30 ° С в течение 3-4 дней, пока колонии появляются.
4. Генетический анализ медленное восстановление фенотипа рост WRN преобразован sgs1 топ3 напряжения.
1. Подряд анализа:
- Чтобы изучить влияние WRN выражение роста дикого типа родительского (W303-1A), sgs1 или sgs1 топ3 штаммы, соответствующие штаммы с YEp112SpGAL или YEp112SpGAL-WRN были прожилками на SC минус Trp пластины, содержащие либо 2% глюкозы ( GLU) или 2% галактозу (GAL). В качестве контроля sgs1 топ3 / YEp112SpGAL-SGS1 был включен. Пластины инкубировали при 30 ° С в течение 2 дней.
- Чтобы определить способность WRN влиять темпы роста sgs1 топ3 напряжение при более низких уровней WRN экспрессии белка, sgs1 топ3 напряжение трансформируется с YEp112SpGAL, YEp112SpGAL-WRN, или YEp112SpGAL-SGS1 был прожилками на SC минус Trp чашек, содержащих различные концентрации гал от ниже, чем 0,005% до достигать 2%. Планшеты инкубировали при 30 ° С в течение 2 дней.
- Для оценки функциональных требований WRN восстановить медленный фенотип рост sgs1 топ3 фон, sgs1 топ3 трансформированных YEp112SpGAL-WRN (E84A, K577M, R834C и K1016A) были streaked на SC минус Trp пластины, содержащие либо Glu или гал в указанных концентрациях. Для элементов управления, sgs1 топ3 напряжение трансформируется с YEp112SpGAL, YEp112SpGAL-WRN или YEp112Sp GAL-SGS1 был включен. Планшеты инкубировали при 30 ° С в течение 2 дней.
2. Жидкие анализ культуры:
- Для оценки роста трансформированных sgs1 топ3 штаммов в жидкой культуре, дрожжевые клетки были выращены в СК раф минус Trp при температуре 30 ° С в течение ночи. Культуры reinoculated в СК раф минус Trp и выросли на 30 ° С до ранней стадии журнал (OD 600 из ~ 0,5). Культуры затем reinoculated на OD 0,05 в СМИ СК, содержащий 2% галлон и рост сопровождался измерения абсорбции при указанных интервалов времени.
5. Генетический анализ гидроксимочевины или methylmethane-сульфонат чувствительности в WRN преобразован штаммов дрожжей.
- Для определения влияния WRN выражение MMS и HU чувствительность sgs1 или sgs1 топ3 штаммы, штаммы трансформированных YEp112SpGAL или YEp112SpGAL-WRN были выращены в СК раф минус Trp при температуре 30 ° C.
- Культуры reinoculated в СК раф минус Trp и выросли на 30 ° С до ранней стадии журнал (OD 600 из ~ 0,6 до 0,8).
- Десять раз серийные разведения этих штаммов были приготовлены с использованием SC среда отсутствует Trp. С экспоненциально растущей культуры, 5 х 10 6 клеток были взяты и разводили серийно до 10000 раз. Четыре мкл каждого разведения был замечен на SC гал минус пластин Trp содержащие указанных концентраций HU или MMS. Пластины инкубировали при 30 ° C.
- В качестве контроля дикого типа родительского штамма (W303-1A) трансформированных YEp112SpGAL или YEp112SpGAL-WRN, или sgs1 топ3 / YEp112SpGAL-SGS1 рассматривалась как описано выше.
6. Клеточного цикла распределения WRN преобразован sgs1 топ3 напряжения.
- Для изучения распределения клеточного цикла WRN преобразован sgs1 топ3 штамма, культуры sgs1 топ3 трансформированных YEp112SpGAL, YEp112SpGAL-WRN, или YEp112SpGAL-SGS1, а вектор-преобразовано дикого типа родительского штамма (W303-1A) были выращены в СК раф минус ГТО при температуре 30 ° С в течение ночи.
- Культуры reinoculated в OD 0.05 и позволяют расти сезам ранней стадии журнал (OD 600 0,5 -0,6). WRN выражение было вызвано добавлением галлон до 2% конечной концентрации. Культуры то позволено расти при температуре 30 ° C в течение 6 ч.
- Культуры затем обрабатываются для DAPI (4 ', 6-diamidino-2-фенилиндола) окрашивания.
- Культур собирают центрифугированием при 5800 мкг в течение 10 мин при комнатной температуре (RT).
- Клетки промывали один раз 1X фосфатным буферным раствором (PBS), а затем были зафиксированы в 70% этаноле в течение 20 мин при комнатной температуре.
- Фиксированные клетки дважды промывали 1 X PBS, и, наконец, ресуспендировали в 1X PBS.
- Клетки были помещены на слайд и монтируется в Vectashield монтажа среде с DAPI (1 мкг / мл) (Vector, США). Клетки были рассмотрены с Axiovert 200 M микроскоп (Zeiss, линзы 100x) и композитных отличие вмешательства дифференциала (DIC). Флуоресцентные изображения были проанализированы с использованием AxioVision, версия 3.0 программы (Zeiss).
7. Вестерн-блот анализ.
1. Для определения экспрессии белка из WRN или WRN варианты в трансформированных sgs1 или sgs1 топ3 штаммов, лизаты были подготовлены следующий метод и белков проанализированы immunblotting.
- Преобразованный sgs1 или sgs1 топ3 штаммы были выращены в СК раф минус Trp при температуре 30 ° С в течение ночи.
- Культуры reinoculated и инкубировали при 30 ° С до начала фазы журнал (OD 600 из 0,5 - 0,8), а затем индуцированные, добавив галлон до 2% конечной концентрации. Культуры то позволено расти при температуре 30 ° C в течение 6 ч.
- Клетки (3 мл) собирали центрифугированием и промывали PBS. Сотовые осадок ресуспендировали в щелочном буфере лизис [50 мМ NaOH, рН 10,5, 2 мМ ЭДТА, 1 мМ phenylmethylsulfonyl фторида (PMSF), 2% SDS, 10% глицерина, 5% 2-меркаптоэтанола и ингибиторы протеазы (Рош молекулярной Biochemicals)] , кипятят 5 мин, уточнил центрифугированием, и нейтрализовали 1 М HCl.
- Белки из эквивалентных количеств Клеточный лизат решались на 8-16% полиакриламидном геле SDS.
2. Для определения уровня экспрессии белка WRN под влиянием различных концентраций галактозы, культур были обработаны следующим образом.
- Преобразованный sgs1 топ3 штаммы были выращены в СК раф минус Trp при температуре 30 ° С в течение ночи.
- Культуры reinoculated и инкубировали при 30 ° С до начала фазы журнал (OD 600 из 0,5 - 0,8), а затем индуцированные, добавив гал до 2% конечной концентрации. Культуры то позволено расти при температуре 30 ° C в течение 6 ч.
- Клетки собирали центрифугированием и ресуспендировали в буфере (50 мМ Трис-Cl, рН 7,5, 1% дезоксихолата натрия, 1% тритона Х-100, 0,1% додецилсульфата натрия [SDS]), содержащий ингибиторы протеазы (0,05% PMSF, 0,05 мкг / мл leupeptin).
- Стеклянные бусы (425 до 600 мкм) были добавлены к смеси, и клетки были разбиты вортексе.
- В результате Гомогенат центрифугировали в течение 5 минут при 15000 мкг и надосадочную фракцию собирают.
- Белок в лизат фракций оценивали по методу Брэдфорда.
- Равные концентрации белка из разных фракций лизата был решен на 8-16% полиакриламидном геле SDS.
3. Выражение WRN или WRN белков мутанта определялась иммуноблоттинга с помощью мыши WRN моноклональные антитела, направленные против эпитопов в очищенной С-концевой фрагмент WRN [6] (1:1000, Спринг-Валли Labs).
4. Первичная инкубации антител с последующей инкубацией с вторичной (пероксидаза хрена, конъюгированных) антител (1:5000). Блоты обрабатываются с ECL Plus Западная система обнаружения в соответствии с протоколом производителя.
5. Для количественного Вестерн-блот анализе, повышение концентрации очищенного рекомбинантного Его с метками полнометражный WRN белка были включены гели, такие, что при количественной оценке пятно стандартной линейной кривой может быть сгенерирован. Кроме того, ячейки выписка из эквивалентное количество клеток или 20 мкг образцов дрожжей лизат были погружены на гели для оценки концентрации WRN.
6. ImageQuant анализ был тогда осуществляется на гели и стандартной линейной кривой был сгенерирован.
7. Концентрация WRN белка в различных концентрациях гал оценивали по стандартной линейной кривой, полученной выше.
8. Представитель Результаты
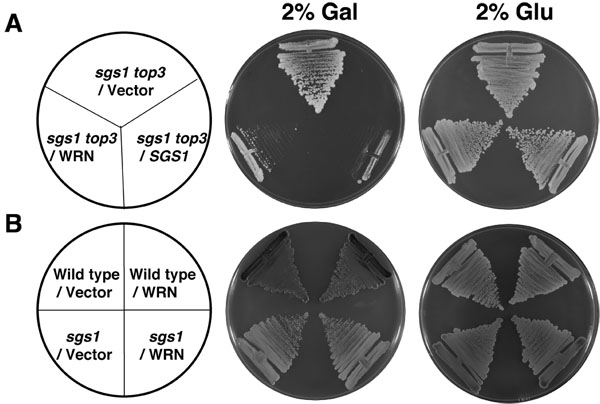
Все штаммы, используемые в настоящем исследовании, Trp ауксотрофы. Таким образом, sgs1, sgs1 топ3 и дикого типа W303-1A штаммы трансформированных YEp112SpGAL или YEp112SpGAL-WRN были отобраны на основе их способности расти в присутствии SC минус Trp СМИ. Чтобы изучить влияние WRN выражение роста трансформированных sgs1 топ3 мутантных клеток, культуры были прожилками на чашки, содержащие SC минус Trp средах с 2%-гал, чтобы побудить WRN выражения. TOP3 decatenates переплетаются молекулы ДНК порожденных Sgs1 геликазы во время репликации [1, 2], поэтому при отсутствии TOP3, торсионное напряжение не освобождается в результате медленного роста и гипер-рекомбинации. Генетической функции sgs1 заключается в подавлении медленный фенотип рост мутанта топ3. Если WRN может заменить SGS1 в генетическом взаимодействии с топ3, восстановление топ3 медленный фенотип рост sgs1 топ3 можно было бы ожидать.
Как показано на рис 2А, WRN преобразован sgs1 топ3 штамма росли значительно медленнее по сравнению с вектором преобразован sgs1 топ3 напряжения. Sgs1 топ3 трансформированных YEp112SpGAL-SGS1 плазмиды был включен в качестве положительного контроля (рис. 2А), демонстрируя, что дикого типа Sgs1 выражается в sgs1 топ3 мутантов удалось генетически дополнением роста фенотипа. Преобразовано sgs1 топ3 штаммы росли так же в отсутствие гал, как показано в течение 2 дней (рис. 2А). Выражение WRN не влияло на рост родительской дикого типа (W3031A) или sgs1 штаммов (рис. 2В), что свидетельствует о влиянии WRN на рост клеток был специально для этого наблюдаются в sgs1 топ3 мутант фоне. Генетические исследования производится с помощью WRN вариантов показал, что WRN геликазы но не экзонуклеазная активность была необходима для восстановления роста топ3 фенотип (рис. 3В). Естественные миссенс полиморфизма в WRN, что мешает геликазы деятельности отменили его способность восстанавливать топ3 медленный фенотип роста.
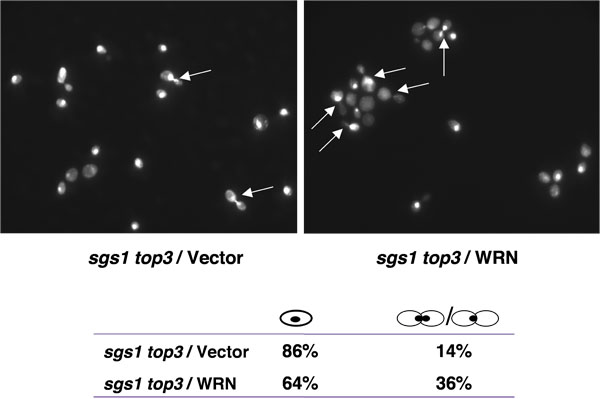
топ3 мутантные штаммы с задержкой в поздней стадии S/G2 клеточного цикла [1], характерно, что можно объяснить их медленный рост. Мутация гена SGS1 в фоновом режиме топ3 подавляет задержка в S/G2 фазе клеточного цикла. Если WRN выражение, может вернуть задержка в S/G2 фазе клеточного цикла в sgs1 топ3, повышенный населения крупных расцветший клетки с ядрами безраздельно можно было бы ожидать для sgs1 топ3 мутанта клеток, экспрессирующих WRN. Как показано на рисунке 4, процент крупных расцветший клетки была выше sgs1 топ3 / WRN чем sgs1 топ3 / вектор, предполагая восстановление S/G2 де лежал характеристика топ3.
Sgs1 топ3 двойной мутант менее чувствителен к MMS или HU чем топ3 одного мутанта [2]. С WRN пострадавших рост клеток из sgs1 топ3, мы затем рассмотрели его влияние на чувствительность к methylmethane сульфонат (MMS, алкилирующий агент) и гидроксимочевины (HU, ингибитор репликации). Sgs1 топ3 / WRN отображается чувствительность к обеим наркотиков сопоставимо с sgs1 топ3 / SGS1 (рис. 5). Генетический анализ WRN вариантов показал, что WRN геликазы / АТФ-азы, но не WRN деятельности экзонуклеазная, было необходимо для его влияние на чувствительность к MMS и HU.

Рисунок 1. Схематическое представление для клонирования WRN / WRN вариантов YEp112SpGAL вектор. Пожалуйста, нажмите здесь для увеличения фигуры 1.

Рисунок 2. WRN выражение в sgs1 топ3 восстанавливает медленный фенотип рост топ3. Группы, sgs1 топ3 напряжение трансформируется с YEp112SpGAL или YEp112SpGAL WRN были штрихом на SC-Trp пластины, содержащие либо 2% Glu или 2% гал. В качестве контроля sgs1 топ3 напряжение трансформируется с YEp112SpGAL SGS1 был прожилками на обеих пластинках. Пластины инкубировали при 30 ° С в течение 2 дней, а затем сфотографировали. Группы B, Дикий тип родительского штамма W303-1A или sgs1 напряжение трансформируется с YEp112SpGAL или YEp112SpGAL WRN были штрихом на SC-Trp пластины либо, содержащий 2% Glu или 2% гал . Пластины инкубировали при 30 ° С в течение 4 дней, а затем сфотографировали. Состав пластин, как в панели и панели B соответственно. Пожалуйста, нажмите здесь , чтобы видеть большую версию рисунке 2.

Рисунок 3. WRN АТФазы / геликазы, но не экзонуклеазная деятельности, требуемое для восстановления медленный фенотип рост топ3 в sgs1 топ3 фоне. Sgs1 топ3 напряжение трансформируется с АТФазы / геликазы-мертвых (YEp195SpGAL WRN K577M), экзонуклеазная-мертвых (YEp195SpGAL- WRN E84A), RQC мутант (YEp195SpGAL-WRN K1016A), или полиморфных мутант (YEp195SpGAL-WRN R834C) был штрихом на SC-Trp пластины, содержащие либо 2% Glu (группа С) или 2% галлон (группа В). Пластины инкубировали при 30 ° С в течение 2 дней, а затем сфотографировали. Состав пластин как в Панели А. Пожалуйста, нажмите здесь , чтобы видеть большую версию рисунке 3.

Рисунок 4. WRN выражение вызывает S/G2 ареста в sgs1 топ3 клеток. Логарифмически растущие культуры sgs1 топ3 напряжение трансформируется с YEp112SpGAL, YEp112SpGAL WRN, или YEp112SpGAL SGS1, а вектор-преобразовано дикого типа родительского штамма были вызваны на уровне 2% гал концентрации в течение 6 ч. Культуры были собраны, обработаны для DAPI окрашивания, как описано в Материалы и методы и наблюдались с помощью Axiovert 200 M микроскоп (Zeiss, линзы 100x). Показана DAPI окрашивание sgs1 топ3 трансформированных YEp112SpGAL (вверху слева) и с YEp112SpGAL WRN (справа вверху). Стрелками показаны ячейки с безраздельной ядер. Распределение клеток в G1 (отдельных клеток) и S/G2 (расцветший клетки) показан на нижней панели. Пожалуйста, нажмите здесь , чтобы видеть большую версию рисунке 4.

Рисунок 5. Влияние WRN выражение MMS и HU чувствительность sgs1 топ3 напряжения. Логарифмически растущие культуры sgs1 топ3 напряжение трансформируется с YEp112SpGAL, YEp112SpGAL WRN, экзонуклеазная-мертвых (YEp112SpGAL-WRN E84A), АТФ-азы / геликазы-мертвых (YEp112SpGAL WRN K577M), RQC мутант (YEp112SpGAL-WRN K1016A), полиморфные мутант (YEp112SpGAL-WRN R834C), YEp112SpGAL SGS1 и векторных преобразован дикого типа родительских штаммов были замечены в десять раз серийные разведения на SC-Trp чашки, содержащие Glu или гал и либо MMS или HU в указанных концентрациях. Пластины инкубировали при 30 ° С в течение 2 дней (контроль пластин) и 4 дня (MMS и HU пластин), а затем сфотографировали. Пожалуйста9/1639fig5.jpg "> нажмите здесь, чтобы посмотреть большую версию рисунке 5.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Одной из сильных сторон использования дрожжей в качестве модельной системы является наличие мутантов в определенных репликации ДНК и ремонт путей, которые сохраняются между дрожжей и людей. Дальнейший отбор трансформантов укрывательство определенных генов просто и надежно, как лабораторных штаммов являются ауксотрофных мутантов и векторы с ауксотрофных маркеров легко доступны. С помощью этих векторов экспрессии гена продуктов можно регулировать, размещая их под контролем индуцируемого промотора (например, галлон индуцибельной промоутера и т.д.). Учитывая эти преимущества, мы разработали систему дрожжей модель для изучения функциональных требований человеческий ген WRN дефекты Вернер синдром в путь, который является потенциально сохраняется между человеком и дрожжами. Использование описанных подходов, мы предоставили первые доказательства того, что WRN может функционировать в генетический путь, который влияет на топ3 связанных фенотипов. Наши наблюдения строке дальнейшего исследования возможности того, что WRN функционально взаимодействует с Top3α в клетках человека во время клеточного репликации ДНК или рекомбинации. Предположительно, в БЛМ с нарушениями условий, WRN может частично заменить BLM через свои партнерские отношения с белком топоизомеразы.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Эта работа была поддержана в полном объеме Внутренние программы исследований NIH, Национальный институт по проблемам старения. Мы благодарим д-р Родни Ротштейн (Колумбийский университет) для штаммов дрожжей и доктор Брэд Джонсон (Университет Пенсильвании, Школа медицины, Филадельфия, штат Пенсильвания) для выражения SGS1 плазмиды.
References
- Aggarwal, M., Brosh, R. M. Jr WRN helicase defective in the premature aging disorder Werner Syndrome genetically interacts with Topoisomerase 3 and restores the top3 slow growth phenotype of sgs1 top3. Aging. 1, 219-233 (2009).
- Gangloff, S., McDonald, J. P., Bendixen, C., Arthur, L., Rothstein, R. The yeast type I topoisomerase Top3 interacts with Sgs1, a DNA helicase homolog: a potential eukaryotic reverse gyrase. Mol Cell Biol. 14, 8391-8398 (1994).
- Shor, E., Gangloff, S., Wagner, M., Weinstein, J., Price, G., Rothstein, R. Mutations in homologous recombination genes rescue top3 slow growth in Saccharomyces cerevisiae. Genetics. 162, 647-662 (2002).
- Sharma, S., Sommers, J. A., Brosh, R. M. Jr In vivo function of the conserved non-catalytic domain of Werner syndrome helicase in DNA replication. Hum Mol Genet. 13, 2247-2261 (2004).
- Gietz, R. D., Schiestl, R. H., Willems, A. R., Woods, R. A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast. 11, 355-360 (1995).
- von Kobbe, C., Thoma, N. H., Czyzewski, B. K., Pavletich, N. P., Bohr, V. A. Werner syndrome protein contains three structure specific DNA binding domains. J Biol Chem. 278, 52997-53006 (2003).

{kind=link}
{kind=link}
{kind=link}
{kind=link}