

Summary
在这里,我们描述了一个基本的协议,以图像和量化现场后,转染哺乳动物组织培养细胞的有丝分裂的时机。
Abstract
蜂窝组织和染色体在有丝分裂过程中发生的动态变化是紧密协调,以确保准确的基因和细胞内容继承。有丝分裂,如染色体运动的标志性赛事,可以很容易地跟踪单个单元格,使用时间推移,哺乳动物细胞表达特定的蛋白GFP标记线的荧光显微镜的基础上。结合RNAi为基础的枯竭,这可能是一个功能强大的方法,针对后一种特定的蛋白质的水平已经降低缺陷发生有丝分裂阶段(S)。在这个协议中,我们提出了一个消耗上有丝分裂的时机的一个潜在的有丝分裂调控蛋白的效果评估的基本方法。是与siRNA的转染细胞,放置在舞台上方的孵化室,和使用自动荧光显微镜成像。我们介绍了如何使用软件设置一个失效时间的实验中,如何处理图像序列,使静止图像的蒙太奇或电影,以及如何量化和分析有丝分裂阶段使用一个细胞系表达的时序mCherry标记组蛋白H2B。最后,我们讨论设计一个时间推移实验的重要因素。这种策略是其他方法的补充和提供的优势1)在寻找作为一个人口和2)分析有丝分裂的细胞,而不需要使用药物治疗的细胞周期同步时,可能无法观察到的动力学变化的敏感性。从这样的成像实验的视觉信息不仅可以使有丝分裂的子阶段进行评估,而且还可以提供意想不到的洞察力,不会明显流式细胞仪细胞周期分析。
Protocol
siRNA转染和细胞制备
- 准备加入的纤维连接蛋白(10μg/mL)0.5ML,以及每个将用于4以及LabTek II腔玻璃罩。孵育10-15分钟室温。
- 根据制造商的指示,准备反向转染的siRNA /脂质体RNAiMAX(Invitrogen)的配合。使用推荐量为每室24道菜要使用(100μL总)。一个类似的产品/转染的协议可以被替换。
- 取出从腔玻璃罩井中的纤维连接蛋白。以及每个将用于添加的siRNA /脂质体RNAiMAX的复合物100μL。
- 分装20000组蛋白H2B - mCherry表达细胞,每孔含转染混合物(500μL)。
- 允许细胞孵化前转染混合物取代常规的细胞培养液1毫升〜24小时。
- 孵育细胞图像采集前24小时(转染后48小时内)。
显微镜设置和图像采集
- 图像采集设置有三个主要部分:1)一个细胞的孵化器,适合在显微镜舞台上和潮湿的环境中保持恒定的37℃,5%的CO 2; 2)倒置显微镜配备有一个自动化的阶段,自动荧光和明帘,自动滤光轮; 3)软件包,集成和控制的阶段,百叶窗,和滤光轮。在我们的成像设置,我们使用一个定制OKO实验室细胞培养箱LabTek二腔玻璃罩可以使用。控制的阶段,百叶窗,滤光轮MetaMorph软件提示:OKO实验室阶段顶部的孵化器系统包括了各种不同的盘,碟,或盖玻片大小的适配器。
- 位置LabTek第二腔玻璃罩在孵化器打开盖子的预热和平衡的孵化器,适配器放置在墓室玻璃罩,并在与橡皮泥的地方举行。所以留在腔玻璃罩盖细胞培养液中不会干出来的。盖上盖子,整个孵化器和显微镜舞台上的地位,确保孵化器是牢固地举行。
- 使用Metamorph,成立了自己的实验,在软件的菜单栏选择“应用程序/多维收购”。这将弹出一个窗口,在其中你可以选择的频率和长度的timelapse的,每种颜色的曝光时间(如果做多个设置),数量和舞台位置的位置,和Z -节数(如需要的话)。选择在哪里保存数据文件提示的功能,如果使用其他软件比Metamorph,可能会略有不同,但整体的概念是相同的。事无巨细,一个开源的基于ImageJ收购与大多数硬件设置兼容的软件,是一个很好的替代。
- 对于本议定书的目的,我们将在两个波长(明和mCherry)为8小时,在15级职位提示每15分钟获取图像:如果需要更好的时间分辨率,那么你可以收购之间的时间间隔缩短,但,较少的阶段职位,可以使用,从而较少的有丝分裂细胞将可用于分析。 时,考虑的时间间隔要考虑到过滤器轮之间切换过滤器设置所需的时间,这一点很重要 。
- 确定最佳曝光时间为每个波长的第一重点领域的使用明照明细胞。在软件中,调整自动对焦功能的唯一的明通道(这将允许该软件在每次收购之前,每个阶段的位置进行自动对焦) 提示:我们用一个40X干相衬的目标,给出色的分辨率没有对石油的需求。视光学所需的分辨率,可用于其他目标 。
- 切换到mCherry波长和捕捉图像,使用50毫秒的曝光。调整的曝光时间,看到了良好的形象所需的最低。关键是要限制的光的照射,以确保持续的细胞活力量。作为一般规则,这最能实现一个中性密度滤光镜(减少激发光通量)和一个较长的相机的曝光时间,而不是通过增加光照强度。重复此过程要使用的任何额外的波长提示:要确定一个长期的细胞活力为您的实验给予曝光,你可以预览图像所需timelapse的总人数,一个细胞可以生存,暂时调整时间点间距从几分钟到秒(即10分钟变成10秒)。如果你的细胞存活100-200风险(或适当数量),那么您的曝光罚款。如果不是,减少您的激发光,曝光时间,或总数的图像模拟您完整的实验。
- 下一步,选择和适当数量标志着一个阶段岗位,让您可以实现细胞的人口有足够的采样。 Metamorph将记录的位置,将返回每个收购提示:一般,我试图找到岗位,其中有大量的细胞图像,但是没有那么多,他们过于密集 。 此外,重要的是选择足够的位置,所以,你将有一个很好的机会观察,通过有丝分裂进展的细胞大量。
- 毕竟timelapse,波长的曝光时间,和舞台上的地位的信息已被记录,选择“预览”屏幕上的按钮,以确定是否该软件是适当控制设备。
- 一旦你感到满意,选择“获取”按钮在屏幕上进行收购。然后坐下来暗示 ,使计算机和显微镜做的工作:通过观察至少有一个收购的丰满圆润的自动化,以确保一切正常工作 。
图像处理和分析
- 收购完成后,下一步就是转移到一个形式,是比较容易处理的图像数据。对于本议定书的目的,我将展示如何使用Metamorph电影,那么如何使用ImageJ程序生成图像的蒙太奇。两种格式将被用于量化的目的。
- 第一步是审查数据。要做到这一点,选择菜单栏中的“应用程序/审查多维数据”。选择文件的位置,然后选择“查看”。这将允许你从每个舞台上的地位的图像的堆栈单独审查。选择所需的舞台上的地位,然后选择“载入图像”。您将能提前通过数据帧帧。
- 对于那些阶段职位中,细胞通过有丝分裂(DNA和细胞形态确定),您可以使电影的蒙太奇使用Metamorph,或与其他程序,如ImageJ(通过美国国立卫生研究院免费提供)。
- 要在Metamorph电影中,选择“过程/保存电影”,在菜单栏中。您将可以选择使用帧的速度,和文件类型的电影。然后选择“保存电影”。
- 为了使蒙太奇,我。STK文件保存为图像的堆栈,它可以用于其他程序,如ImageJ。
- 在ImageJ打开该文件,并提前帧,直到找到一个细胞通过有丝分裂。作物的面积和周围的细胞选择“图像/重复” 提示:确保所有帧重复。
- 为了蒙太奇,选择“图像/垛/蒙太奇”在菜单栏中。在这里您可以指定您要包括,蒙太奇的大小和形状的帧,以及是否要帧之间的边界。选择这些热播的“好”。 。保存为TIF文件的蒙太奇,并做进一步处理ImageJ或Adobe Photoshop中的任何提示:更改Metamorph使用更方便的图像处理,以8位格式的 16位格式的文件蒙太奇。
- 要计算出在不同阶段的有丝分裂的时间,确定T = 0(DNA凝结或细胞四舍五入的第一个迹象),计数染色体在赤道(在前期/前中期的时间),帧帧的数量,直到对齐,直到染色体开始隔离(在中期的时间),帧,直到细胞开始变平(后期/末期/胞质)。在每个有丝分裂阶段的计算时间取决于每帧之间的时间间隔。
代表性的成果

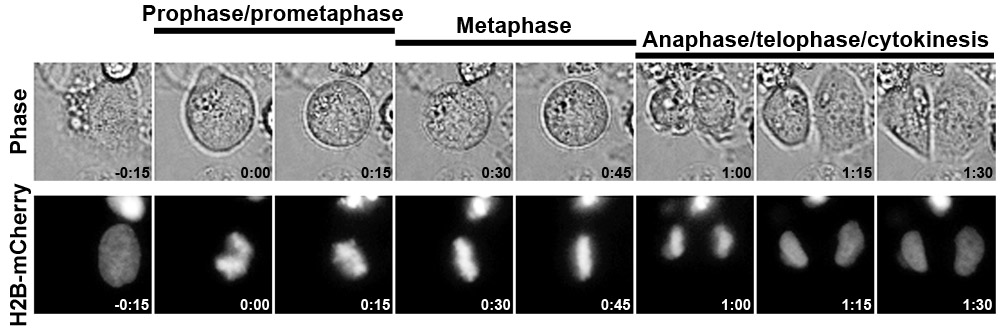
图1说明了一个典型的对照siRNA转染实验中使用的HeLa细胞系表达组蛋白H2B - mCherry结果。这种方法已被用来有效地量化的有丝分裂的时机,揭示了一个意想不到的作用nucleoporin Nup153在有丝分裂1月底的时间。请点击这里看到图1的放大版本。
Discussion
在成像和荧光蛋白技术的最新进展,实时成像已经成为常规的细胞分析2方面。一个各种绿色荧光蛋白(和其他颜色的变种3)标签蛋白被广泛使用或相对容易构造的,可以用来追踪一个细胞分裂的标志,包括DNA / 染色体的动态4,5,中心体复制,细胞周期蛋白B动态7,核信封细分和重组8,9,10有丝分裂纺锤体的形成, 胞质 11的各个阶段。标签蛋白,可单独或联合使用时的激发/发射光谱,荧光标记是兼容的,允许特定事件之间的协调,以进行评估。如果更大的空间和/或时间分辨率/所需,是否可以使用激光扫描,旋转盘,或共振扫描共聚焦显微镜,并与日益增加的分辨率复杂的显微镜选项列表中继续增长。在哺乳动物细胞的有丝分裂的实时成像也已适应在高通量的RNAi和小分子屏幕12-14使用。因此,一个S最初的实验中,使用这种技术,而可能是相对简单的,这一战略的基础上的可能性是多方面的。
Acknowledgments
这项工作是由美国癌症协会(PF - 07 - 103 - 01 - CSM),美国国立卫生研究院(R01 GM61275和P30 CA042014),白血病和淋巴瘤协会,Huntsman癌症基金会的支持。
Materials
| Name | Company | Catalog Number | Comments |
| Lipofectamine RNAiMAX | Invitrogen | 13778-075 | |
| Lab Tek II Chambered Coverglass (4-well) | Thermo Fisher Scientific, Inc. | 12-565-337 | Additional chamber sizes are available |
| Fibronectin | Sigma-Aldrich | F1141 | |
| Stage-top cell incubator | OKO Lab | Can use any appropriate chamber | |
| Automated inverted fluorescence microscope | Olympus Corporation | Can use any appropriate microscope | |
| Software package | Metamorph | Can use any appropriate software |
References
- Mackay, D. R., Elgort, S. W., Ullman, K. S. The nucleoporin Nup153 has separable roles in both early mitotic progression and the resolution of mitosis. Mol Biol Cell. 20, 1652-1660 (2009).
- Khodjakov, A., Rieder, C. L. Imaging the division process in living tissue culture cells. Methods. 38, 2-16 (2006).
- Shaner, N. C., Steinbach, P. A., Tsien, R. Y. A guide to choosing fluorescent proteins. Nat Methods. 2, 905-909 (2005).
- Meraldi, P., Draviam, V. M., Sorger, P. K. Timing and checkpoints in the regulation of mitotic progression. Dev Cell. 7, 45-60 (2004).
- Mora-Bermudez, F., Ellenberg, J. Measuring structural dynamics of chromosomes in living cells by fluorescence microscopy. Methods. 41, 158-167 (2007).
- Prosser, S. L., Fry, A. M. Fluorescence imaging of the centrosome cycle in mammalian cells. Methods Mol Biol. 545, 165-183 (2009).
- Malureanu, L. A. BubR1 N terminus acts as a soluble inhibitor of cyclin B degradation by APC/C(Cdc20) in interphase. Dev Cell. 16, 118-131 (2009).
- Anderson, D. J., Hetzer, M. W. Reshaping of the endoplasmic reticulum limits the rate for nuclear envelope formation. J Cell Biol. 182, 911-924 (2008).
- Beaudouin, J., Gerlich, D., Daigle, N., Eils, R., Ellenberg, J. Nuclear envelope breakdown proceeds by microtubule-induced tearing of the lamina. Cell. 108, 83-96 (2002).
- Marcus, A. I. Visualization of spindle behavior using confocal microscopy. Methods Mol Med. 137, 125-137 (2007).
- Steigemann, P. Aurora B-mediated abscission checkpoint protects against tetraploidization. Cell. 136, 473-484 (2009).
- Draviam, V. M. A functional genomic screen identifies a role for TAO1 kinase in spindle-checkpoint signalling. Nat Cell Biol. 9, 556-564 (2007).
- Neumann, B. High-throughput RNAi screening by time-lapse imaging of live human cells. Nat Methods. 3, 385-390 (2006).
- Stegmeier, F. Anaphase initiation is regulated by antagonistic ubiquitination and deubiquitination activities. Nature. 446, 876-881 (2007).

{kind=link}