

Summary
Dopamin erstatning farmakoterapi ved hjelp av L-DOPA er den mest brukte symptomatisk behandling av Parkinsons sykdom, men er ledsaget av bivirkninger, inkludert ufrivillige unormale bevegelser, kalt dyskinesi
Abstract
MALDI bildebehandling massespektrometri (IMS) er en kraftig tilnærming som muliggjør romlig analyse av molekylære arter i biologiske vevsprøver 2 (Fig.1). En 12 mikrometer tynne vevet delen er dekket med en MALDI matrise, noe som letter desorpsjon og ionisering av intakte peptider og proteiner som kan oppdages med en masse analysator, vanligvis ved hjelp av en MALDI TOF / TOF massespektrometer. Vanligvis hundrevis av topper kan vurderes i et enkelt rotte hjernevev delen. I motsetning til vanlige imaging teknikker, ikke denne tilnærmingen ikke krever forkunnskaper i molekyler av interesse og gir mulighet for ukontrollert og omfattende analyse av flere molekylære arter samtidig opprettholde høy molekylær spesifisitet og sensitivitet to. Her beskriver vi en MALDI IMS tilnærming for å belyse region-spesifikke distribusjon profiler av nevropeptider i rottehjernen av en dyremodell Parkinsons sykdom (PD).
PD er en vanlig nevrodegenerativ sykdom med en prevalens på 1% for personer over 65 av alder 3,4. Den vanligste symptomatisk behandling er basert på dopamin utskifting bruke L-DOPA 5. Men dette er ledsaget av alvorlige bivirkninger, inkludert ufrivillige unormale bevegelser, betegnes L-DOPA-dyskinesier (LID) 1,3,6. En av de mest fremtredende molekylær endring i LID er en oppregulering av opioid forløperen prodynorphin mRNA 7. De dynorphin peptider modulere neurotransmission i hjernen områder som er vesentlig involvert i bevegelsen kontroll 7,8. Imidlertid har hittil nøyaktig opioide peptider som stammer fra behandlingen av neuropeptid forløperen ikke blitt karakterisert. Derfor benyttet vi MALDI IMS i en dyremodell av eksperimentell Parkinsons sykdom og L-DOPA indusert dyskinesi.
MALDI bildebehandling massespektrometri viste seg å være spesielt fordelaktig med hensyn til neuropeptid characterizasjon, ettersom vanlige antistoff baserte tilnærminger rettet mot kjente peptid-sekvenser og tidligere observerte posttranslasjonelle modifikasjoner. I motsetning MALDI IMS kan rakne romanen peptid bearbeiding produkter og dermed avsløre nye molekylære mekanismer neuropeptid modulering av neuronal overføring. Mens den absolutte mengden av nevropeptider ikke kan avgjøres ved MALDI IMS, den relative overflod av peptid ioner kan avgrenses fra masse spektra, gir innsikt om hvordan du endrer nivåer i helse og sykdom. I eksemplene som presenteres her, ble peak intensitet av dynorphin B, alfa-neoendorphin og substans P funnet å være signifikant økt i dorsolateral, men ikke dorsomedial, striatum av dyr med alvorlig dyskinesi involverer ansikts, bagasjerommet og orolingual muskler (Fig. 5). Videre avslørte MALDI IMS en sammenheng mellom dyskinesi alvorlighetsgrad og nivåer av des-tyrosin alpha-neoendorphin, representerer en tidligere ukjent mekanisme funksjonell inactivasjon av dynorphins i striatum som fjerning av N-terminal tyrosin reduserer dynorphin har opioid-reseptor binding kapasitet 9. Dette er den første studien på neuropeptid karakteriseringen i LID med MALDI IMS og resultatene markere potensialet av teknikken for anvendelse i alle felt av biomedisinsk forskning.
Protocol
Protokollen er justert for formålet av statistisk analyse av MALDI IMS data fra flere rottehjernen seksjoner, typisk 20-30 seksjoner, og består av fem ulike trinn bestående vev forberedelse, matrise program, MALDI-TOF MS analyse, data evaluering, og neuropeptid identifikasjon. Prosedyrene er beskrevet og omtalt i mer detaljert nedenfor:
1. Tissue forberedelse
Denne prosedyren omfatter innsamling av de respektive vevsprøver samt vev seksjonering for IMS analyse. Et spesielt mål i protein og peptid analyse er å unngå proteolytisk degradering. Derfor er det viktig å jobbe raskt og flittig under vev disseksjon.
- Ofre rotter (typisk 250-300 g) ved halshogging, fjerne rottehjernen innen maksimalt post-mortem tid <30-årene og fryse på pulverisert tørris før de overføres til -80 ° C fryser. Raskere frysing med flytende nitrogenøke risikoen for microtears i hjernevev, som vil negativt påvirke matrise krystallisering og dermed redusere MS kvalitet (Fig. 2D). Hele hjerner kan lagres i flere år før seksjonering uten tap av MS signalkvalitet.
- Skjær frosne vev på en cryostat mikrotomen til 12 mikrometer skiver og tine-mount vevsdelene på ledende MALDI glassplater (indium tinn oksid belagt lysbilder, Bruker Daltonics) eller MALDI target (Fig. 2A-C).
- Tørre seksjoner for 15 min under vakuum og lagre lysbilder ved -80 ° C til videre bruk. Vevsdelene bør analyseres i løpet av kortest mulig tid etter seksjonering, selv om oppbevares ved -80 ° C. Vi finner at MS signalkvaliteten vil bli merkbart redusert etter et år i oppbevaring. For å redusere oksidasjon av proteiner og peptider, kan luften i oppbevaringsboksen bli erstattet med en inert gass (f.eks argon eller nitrogen).
2. Matrix søknad
Matrisen søknadtrinn har en betydelig innvirkning på spekteret kvalitet og krever optimalisering av flere parametere avhengig av type vev samt analytten av interesse. Disse faktorene omfatter kjemiske parametere som type matrise, matrix konsentrasjon, pH, vev vasking og organiske modifikatorer samt instrumentale innstillinger, inkludert innskudd volum, lateral oppløsning og antall depositions 10 (Fig. 2D). For storskala eksperimenter, er det av stor betydning for å redusere variansen, for eksempel ved å bruke matrisen til alle seksjoner innen en dag, og av samme operatør. Selv om det er mange strategier for å søke matrise løsning som ved sublimasjon eller spray, den automatiserte avsetning av matriser av små matrise dråper, om 100-150 picoliter i størrelse, har vært brukt med hell for analyse av små proteiner og nevropeptider i ulike vev , inkludert hjernen § 9, 10,11, 12, 13.
- Avrime seksjoner i en dessicator for 1 tvår.
- Pass på at forsøket blir blendet av en annen person enn operatøren. Re-label alle prøvene.
- Vask seksjoner 1x i 70% etanol (EtOH, ved romtemperatur, RT) i 10 sek og to ganger i 95% EtOH (RT) for 10 sek. For store eksperimenter, utfører vask for alle glass glir sammen ved hjelp av en kyvette for å minimere variasjon.
- Tørk delene i en dessicator for 10 min.
- Vurdere vevsdelene under et mikroskop og se etter vev forvrengning, microtears og små sprekker som vil forringe MALDI MS kvalitet (Fig. 2D).
- Forbered frisk matrise løsning bestående av 50 mg / ml DHB i 50% metanol, 10% 150mm ammonium acetat (Amac) og 0,3% Trifluoroacetic syre (TFA) i vann.
- Matrix programmet er utført av diskrete dråpene deponering i et rektangulært mønster ved hjelp av en kjemisk blekkskriver (CHIP, Shimadzu). Det første trinnet er å optimalisere eksperimentelle parametre matrise søknad om neuropeptid analyse including antallet dråper per pass, antall pasninger. Dette eksperimentet er utført ved å bruke flere matrise matriser med forskjellige program parametre på samme vev delen, samtidig sørge for at hver matrise dekker tilsvarende områder av hjernen som corpus callosum, cortex, og striatum. Det samme eksperimentet må utføres hver gang parametere endres, inkludert forskjellige strukturer i hjernen, ulike matriser målretting spesifikke analytter, og dersom ulike matrise løsemidler er nødvendig for utvinning av spesifikke analytter.
- Skann glasset lysbildeholderen med vev seksjonen og justere holderen. Definer din array for matrise-programmet på vev seksjonen, og angi romlig oppløsning dvs. sted å få øye på avstand. Påfør matrise ved hjelp av optimaliserte protokollen på den kjemiske blekkskriver. For dette eksperimentet brukte vi en optimalisert protokoll for peptid avbildning med følgende utskrift parametere: 10 dråper (100 pl / drop), passerer 10 søknad og et sted å få øye på distance på 300 mikrometer.
- Scan endelig matrise oppdaget seksjoner og lagre bildet for registrering før MALDI datainnsamling (trinn 3,4).
- Oppbevar delene inntil videre bruk i en dessicator under vakuum.
3. MALDI MS datainnsamling og prosessering
MS analyse av nevropeptider blir utført på en MALDI flygetid instrument (Ultraflex II, Bruker Daltonics, Tyskland) opererer i reflektor-modus, bruker programvare assistert datainnsamling fra hver enkelt matrise sted 14. Derfor nøyaktig romlig undervisningen er avgjørende. Det er viktig at MALDI optimalisering, kjøp og spesielt målet registrering eksperimenter utføres av samme operatør som bør fortrinnsvis blindet de eksperimentelle grupper. I en storstilt eksperiment med flere glassplater kan de MALDI eksperimenter skal utføres av en operatør, mens en annen person er i drift den kjemiske blekkskriver.
- Legge glassplater i massespektrometer.
- Sjekk kalibrering av MALDI oppkjøpsmetoden ved hjelp av en lav molekylvekt standard kalibrering mix (Bruker Daltonics).
- Optimaliser oppkjøp parametere.
- For å optimalisere MS signal og for å unngå ablating matrisen fra nabolandene matrise innskudd, bør størrelsen på laser og optimal fokus på vev bestemmes.
- Laseren energi er satt for å sikre maksimal MS kvalitet fra så mange matrix innskudd som mulig uten å heve grunnlinjen, redusere peak oppløsning eller mette detektoren.
- Vurdere maksimalt antall bilder per matrise sted til bare støy blir oppdaget, ofte 1000-2000 skudd. Anslå antall bilder som bør akkumulert og antall skudd anskaffet før laseren posisjon innenfor en flekk bør endres. For å prøve hver matrise flekk jevnt, akkumuleres vi 600 bilder på 25 skudd trinn, for et totalt antall på 24 trinn ved hjelp av en random bevegelsesmønster, fra hver matrise deponering.
- Registrer skanning av alle flekket deler til motor koordinater i MALDI scenen ved hjelp av FlexImaging (v.2.0) 10 og utføre datainnsamling i batch-modus ved AutoXexuteBatchRunner.exe programvare.
- Behandle hver enkelt spektra ved hjelp av baseline subtraksjon (Convex skrog V3), glatting og ekstern kalibrering (valgfritt), etterfulgt av eksport som en ASCII-fil (*. Dat, *. Txt eller *. Csv-format). 15
4. Dataanalyse
Endelig dataanalyse består data post prosessering og data reduksjon av fokus bare på topp, fulgt av statistisk analyse.
- Som et første skritt ble MALDI IMS delene evalueres for overnormalization effekter. Dette kan enkelt oppnås ved å ansette datavisualisering verktøy som FlexImaging (Bruker Daltonics) eller BioMap (Novartis). Som et første skritt de samlede ion bildene er evaluated før total ion strøm (TIC) normalisering, etterfulgt av manuell inspeksjon av single ion distribusjon bilder av forskjellige fremtredende peptid topper. Se etter karakteristiske peak intensitet distribusjoner, og hvis de er knyttet til vev funksjoner (skader), spotting kvalitet eller normalisering effekter (Fig. 3).
- Avgrense områder av interesse (f.eks striatum) i henhold til histologiske funksjoner og eksportere tilsvarende spektra i ascii-format. Fortrinnsvis kan normalisering av spektra til den totale ion strømmen (TIC) utføres på dette stadiet.
- Importere ASCII-filer i en datahåndtering programvare som Origin (v.8.1, Originlab), MATLAB (MathWorks, Boston, MA, USA) eller R 16. Toppdeteksjon kan utføres ved hjelp peak søkeverktøy inkludert i programvaren, for eksempel "peak analyse" i Origin eller "mspeaks" i Matlab. Eksporter peaklists fra alle spektra som en enkelt fane tekstfil.
- For å avgjøre bin grenser for detektert peptidtopper, er Binning analyse utført ved hjelp av egnede verktøy (f.eks pbin 17) eller in-house skriftlig skript for MATLAB eller R. Her singelen tekstfil som inneholder alle peak plukket data er lastet inn i programvaren og parametrene for peak grensen bestemmelse er spesifisert for eksempel hvor ofte en topp bør være tilstede i spektra for å være relevant for eksperimentet. For eksempel inneholder eksperimentet 2 grupper av dyr, 5 dyr i hver gruppe, og 100 spektrene er samlet inn fra hvert dyr og region av interesse. Anta en topp er potensielt interessant hvis det er til stede i minst majoriteten av dyrene i en gruppe (3/5) og i minst halvparten av spektra av disse dyrene (3x50 = 150 spektra), vil dette gi en total prosentandel på 15% for 150 positive spektra av totalt 1000 (2x5x100) spektra. Bruke pbin verktøy, gir dette trinnet en enkelt binrange fil som inneholder alle bin bredder bestemmes fra de oppkjøpte data. For å bekrefte at bin grenserer hensiktsmessig, er det enkelt å visualisere de skuffer i Origin sammen med de opprinnelige spektra spor.
- Peak-området integrering kan redusere varians som er viktig for statistisk analyse. Vi bruker en in-house skrevet manus til R for å beregne arealet under kurven mellom topp grenser fastsatt i trinn 4. Integrerte peak områdene er importert til MS Excel (v.2007) og statistisk analyse ved hjelp av ikke-parametriske uparede test med SAM-verktøyet er utført 18.
5. Peptid identifikasjon
Sekvens verifikasjon av de observerte peptid identiteter er avgjørende for å konkludere biologisk relevans. Den mest nøyaktige metoden inkluderer sann top-down bestemmelse direkte av vev ved hjelp av peptid fragmentering ved hjelp av tandem massespektrometri (MS / MS), selv om høye peptid konsentrasjoner er nødvendig for denne typen analyser 12,13. For lave rikelig peptider eller flere peptider med closEM / z verdier (± 0,5%), er på vev analyse svekket og av vev analyse med peptidomic strategi benyttes som inkluderer ekstraksjon, separasjon og MS basert identifisering av endogene nevropeptider. For forsøket presenteres her, var det sentrale fokuset på opioid peptid oppdagelse, som er en særlig utfordring fordi disse peptidene er ganske lav rikelig sammenlignet med andre nevropeptider i spektra. Videre disse peptidene er heller polare som gjør dem relativt hydrofile og vanskelig å beholde med felles peptid utvinning og separasjonsteknikker .. Derfor har vi brukt en tidligere rapportert protokoll for vev utvinning og opioid peptid prefractionation i kombinasjon med standard LC-MS/MS basert peptid identifikasjon 9,19.
- Samle koronale deler av målgruppen strukturer av interesse (nucleus accumbens, NAC, caudatus putamen, CPU). Monter frossen rottehjernen i en cryostat mikrotomen og fjerne omliggende hjerne materiale (CORtex, septum, corpus callosum) med en skalpell. Samle seksjoner (30 mikrometer, n = 50) av dissekert NAC og CPU og tøvær montere NAC og CPU deler av avsnittene om forskjellige glassplater.
- Pakk peptider off vev ved å tilsette 100 mL 5% ACN/0.1% TFA, inkuberes i to minutter og samle i Eppendorf lav proteinbinding rør. Gjenta dette trinnet to ganger.
- Utføre peptid prefractionation ved hjelp av sterk kationebytter kromatografi med trinnvis (n = 4) eluering ved økt ionisk styrke 19. Tørk ned prøvene under vakuum ved hjelp av en speedvac konsentrator.
- Analyser peptid fraksjoner ved hjelp av nanoflow C18 reversert fase væskekromatografi (1100, Agilent Technologies, Santa Clara, CA) tilkobles electrospray tandem massespektrometri (LC-MS/MS). MS eksperimenter ble utført på en hybrid lineær iontrap / Fourier transform ion syklotronen resonans (FTICR) instrument (LTQ FT 7T, Thermo Scientific, Waltham, MA,) Peptide fulscan spektra (m / z 150-2000) Ble anskaffet med FTICR analysatoren med høy masse oppløsning (100 000) etterfulgt av påfølgende fragmentering av de 5 mest intense peptid toppene i iontrap ved hjelp av kollisjon indusert dissosiasjon (CID) 9.
- Peptidsekvens identifikasjon utføres av database matching og kan suppleres med de-novo sekvensering analyse. For database søk blir kommersielt tilgjengelige søkemotorer (Mascot, XTandem eller Protein prospector) ansatt 20. Søkene er vanligvis utføres mot databaser som inneholder sekvenser av kjente eller antatte nevropeptider og sekvenser av neuropeptid forløper proteiner 21.
6. Representative Resultater
MALDI avbildning massespektrometri av striatale banene vevsdelene så forberedt i henhold til protokollen som er beskrevet her resulterte i påvisning av mer enn 1000 topper tilsvarende ca 300 monoisotopic molekylære arter (gjennomsnitt spektra visti fig. 1). Datavisualisering for fremtredende molekylær ion toppene ble oppnådd ved hjelp av Flex bildebehandling og viste karakteristiske peak intensitet distribusjoner som er godt i tråd med anatomiske trekk (Fig.3). Et ytterligere trekk ved MALDI IMS er dens relative god reproduserbarhet. I dette eksperimentet, var den samlede koeffisient av varians for de maksimale intensitet av alle registrerte molekylære arter 30%, men mange topper vises svært lav variasjon og høy reproduserbarhet innen behandlingsgruppene (fig 4). De relative peak intensitet data fra fire ulike regioner av interesse, inkludert dorsolateral og dorsomedial del av både lesioned og intakt striata ble utsatt for statistisk analyse. For å justere for flere sammenligninger samtidig, ble den statistiske analysen utføres ved hjelp av parametrisk testing med SAM-verktøy 18. De mest fremtredende endringene ble funnet i dorsolateral delen av dopamin-denerveres, Parkinsonrelaterte striatum. Her si gnificant endringer i mellom de ulike behandlingsgruppene ble observert for to dynorphin peptider, dynorphin B og alpha-neoendorphin (Fig.5). I detalj, ble en relativ økning av både dynorphin peak intensitet med 50-60% observert i høy dyskinetic dyr sammenlignet med lave dyskinetic dyr og lesjon kontroller (p <0,05, F (2, 15) = 12,8 DynB, F = 5,7 aNeo; fig. 5).

. Figur 1 Gjennomsnittlig MS spor innhentet fra to nærstående regioner i striatum; caudatus putamen (CPU) og nucleus accumbens (NAC). De to regionene vise ulike MS profiler med noen molekylære arter unikt uttrykt i en region, eller på ulike peak intensitet (innsats, m / z 2028). Den romlige fordelingen mønster av hver topp kan visualiseres ved hjelp av spesialisert bildebehandlingsprogrammer (nederst panel).
2.jpg "/>
Figur 2 (A) Hjernen er montert på en cryostat chuck ved hjelp av en integreringsrettigheter medier (OTC, pil)., Er omsorg tatt at OTC ikke forurense området av hjernen til å være seksjonert ettersom OTC årsaken ion undertrykkelse av peptider. (B, C) tynne snitt (≈ 12 mikrometer tykkelse) er tin-montert på MALDI kompatible glassplater og tørket i noen sekunder for å unngå frost skader som sett i C. (D) Microtears kan være vanskelig å oppdage med det blotte øye , men svekke MALDI matrise krystallisering og utslette MALDI MS signal. Det samme seksjonen farget med cresyl fiolett avslører microtears og sprekker (nederst til høyre mikrofotografiet).

Figur 3. Det første trinnet i data evalueringen er å visualisere flere topper over massen området analyseres (AI). Her ble striatale banene deler fra 9 mus avbildes med MALDI MS. Visualisering av den gjennomsnittlige totale jegpå dagens vil avdekke områder med påfallende høye eller lave ion intensiteter (piler). Disse områdene kan bli berørt av over-eller under-normalisering effekter og forvrenge dataanalyse kompromiss resultatene. Dårlig anatomiske definisjon av peak distribusjoner avdekke seksjoner med generelt lav topp signal-til-støy, for eksempel § § 3 og 9, topper F gjennom I.

Figur 4. MS reproduserbarhet mellom behandlingsgruppene kan vurderes ved å beregne gjennomsnittlig MS kurven og standard feil for hver m / z-verdien (innstikk, m / z 722 og 1749). God reproduserbarhet sikrer gyldig statistisk analyse.
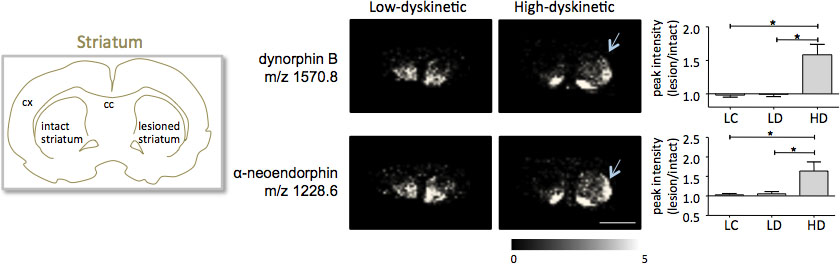
Figur 5. Dynorphin B og alpha-neoendorphin peak intensitet er betydelig økt i 6-OHDA-Lesioned, Parkinsonrelaterte, striatum av high-dyskinetic dyr (HD, piler) sammenlignet med lav-dyskinetic (LD) og lesjon kontrollgruppen (LC). Peptid peak intensitet uttrykt som gjennomsnittlig fold-endring av intakte siden ± SEM (lesjon / intakt side). * P <0.05; cx cortex, cc corpus callosum. Scale bar 5 mm.
Discussion
Det er flere fordeler ved å ansette MALDI bildebehandling massespektrometri i studiet av nevropeptider. En objektiv analyse av MS data kan avsløre at bare bestemte hjerne kjerner, eller som i de resultatene som presenteres her hvor kun den dorsolateral delen av striatum er forbundet med en viss patofysiologisk tilstand. Ved å beholde den romlige informasjonen er det da mulig å omdefinere deler av interesse å utføre statistisk analyse med høyere følsomhet og lavere variasjon sammenlignet med analyse av hele hjernen seksjoner eller ved hjelp av tradisjonelle peptidomics studier på peptid ekstrakter. I tillegg er det viktig å innse MALDI IMS lett kan oppdage tidligere ukjente posttranslasjonelle modifikasjoner, men strukturelle analyser må følge for å fastslå de eksakte aminosyreposisjonene som er modifisert.
Vanlige fallgruver i visualisere MALDI IMS data omfatter kartlegging maksimal peak intensitet til en lineær optisk skala fra blACK (0%) til farge (100%) for hver seksjon i den eksperimentelle serien (figur 3), i stedet for å kartlegge alle seksjonene til en felles absolutt skala der 100% er det maksimale topp intensitet av alle seksjonene (figur 5) . Den siste metoden tillater sammenligning av gruppe data og visualisering av forskjeller mellom behandlingsgruppene.
En stor hindring i MALDI IMS analyse er tildelingen av peptider til spesifikke masse topper. On-vev tandem-massespektrometri er noen ganger mulig, men ofte viser seg ganske vanskelig 13,14. Vi finner at en mer tradisjonell tilnærming inkludert en preparativ fraksjonering på sterk kationebytter kromatografi, etterfulgt av endrede fase LC-MS/MS kan brukes for å kunne sekvens mange nevropeptider og spesielt opioide peptider. Det er fortsatt ikke uvanlig å oppnå god kvalitet MS / MS spektre som ikke samsvarer med noen database oppføringer ved hjelp av vanlige søkemotorer som maskot. I disse tilfellene de novo-sekvensering for hånd er det only alternativet. Den ultimate bevis på topp identitet kan fås ved MALDI IMS av vevsdelene fra den aktuelle knockout mus, men dette er ikke alltid tilgjengelig eller mulig. Et alternativ er å validere resultatene av en diametralt annen metode, for eksempel ved vestlig immunoblotting eller immunhistokjemi. Dette kan ofte inkludere oppdra antistoffer og en betydelig mengde arbeid validere de nye antistoffer.
Den generelle strategien som presenteres i denne protokollen er optimalisert for store neuropeptid MALDI IMS eksperimenter inkludert flere seksjoner og eksperimentelle betingelser. Protokollen er optimalisert spesielt for opioide peptider, og vil ha stor betydning i fremtidige studier, som sysselsatte i ulike forskningsfelt, inkludert mekanismer underliggende smerte og den endogene respons på legemidler av avhengighet.
Disclosures
Forfatterne har ikke noe å avsløre.
Acknowledgments
Vi takker Hanna Warner for å bidra dataene for figur 3 og professor Jonas Bergquist for verdifulle innspill. Den svenske Forskningsrådet (Grant 522-2006-6416 (MA), 521-2007-5407 (MA), The Åke Wiberg stiftelse (MA, JH), Det kongelige svenske vitenskapsakademiet (MA, JH), og den svenske Chemical Society (JH) er takknemlig anerkjent for økonomisk støtte.
References
- Obeso, J. A., Olanow, C. W., Nutt, J. G. Levodopa motor complications in Parkinson's disease. Trends Neurosci. 23, S2-S7 (2000).
- Caprioli, R. M., Farmer, T. B., Gile, J. Molecular imaging of biological samples: localization of peptides and proteins using MALDI-TOF MS. MALDI-TOF MS. Anal. Chem. 69, 4751-4760 (1997).
- Obeso, J. A. The evolution and origin of motor complications in Parkinson's disease. Neurology. 55, S13-S20 (2000).
- O, W. H. Noncommunicable Diseases and Mental Health Cluster, Noncommunicable Disease Prevention and Health Promotion Department, Ageing and Life Course. Active Ageing: A Policy framework. , (2002).
- Schapira, A. H. Movement disorders: advances in cause and treatment. Lancet Neurology. , 6-7 (2010).
- Obeso, J. A., Rodriguez-Oroz, M. C., Rodriguez, M., DeLong, M. R., Olanow, C. W. Pathophysiology of levodopa-induced dyskinesias in Parkinson's disease: problems with the current model. Ann. Neurol. 47, S22-S32 (2000).
- Cenci, M. A., Lee, C. S., Bjorklund, A. L-DOPA-induced dyskinesia in the rat is associated with striatal overexpression of prodynorphin- and glutamic acid decarboxylase mRNA. Eur. J. Neurosci. 10, 2694-2706 (1998).
- Andersson, M., Hilbertson, A., Cenci, M. A. Striatal fosB expression is causally linked with l-DOPA-induced abnormal involuntary movements and the associated upregulation of striatal prodynorphin mRNA in a rat model of Parkinson's disease. Neurobiol Dis. 6, 461-474 (1999).
- Hanrieder, J. Alterations of striatal neuropeptides revealed by imaging mass spectrometry. Molecular & Cellular Proteomics. , (2011).
- Cornett, D. S., Reyzer, M. L., Chaurand, P., Caprioli, R. M. MALDI imaging mass spectrometry: molecular snapshots of biochemical systems. Nat. Methods. 4, 828-833 (2007).
- Ljungdahl, Imaging Mass Spectrometry Reveals Elevated Nigral Levels of Dynorphin Neuropeptides in L-DOPA-Induced Dyskinesia in Rat Model of Parkinson's Disease. PLoS ONE. 6, e25653 (2011).
- Groseclose, M. R., Andersson, M., Hardesty, W. M., Caprioli, R. M. Identification of proteins directly from tissue: in situ tryptic digestions coupled with imaging mass spectrometry. J. Mass. Spectrom. 42, 254-262 (2007).
- Andersson, M., Groseclose, M. R., Deutch, A. Y., Caprioli, R. M. Imaging mass spectrometry of proteins and peptides: 3D volume reconstruction. Nat. Methods. 5, 101-108 (2008).
- Deininger, S. -O. Imaging Mass Spectrometry. Setou, M. , Springer. Japan. 199-208 (2010).
- Norris, J. L. Processing MALDI Mass Spectra to Improve Mass Spectral Direct Tissue Analysis. Int. J. Mass. Spectrom. 260, 212-221 (2007).
- Ihaka, R., Gentleman, R. R. A Language for Data Analysis and Graphics. Journal of Computational and Graphical Statistics. 5, 299-314 (1996).
- Mass Spectrometry Binning Software GAB. , Vanderbilt Center for Quantitative Sciences. Nashville, TN. Available from: http://www.vicc.org/biostatistics/software.php (2012).
- Tusher, V. G., Tibshirani, R., Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. U.S.A. 98, 5116-5121 (2001).
- Bergstrom, L., Christensson, I., Folkesson, R., Stenstrom, B., Terenius, L. An ion exchange chromatography and radioimmunoassay procedure for measuring opioid peptides and substance P. Life. Sci. 33, 1613-1619 (1983).
- Falth, M. Neuropeptidomics strategies for specific and sensitive identification of endogenous peptides. Mol. Cell. Proteomics. 6, 1188-1197 (2007).
- Falth, M. SwePep, a database designed for endogenous peptides and mass spectrometry. Mol. Cell. Proteomics. 5, 998-1005 (2006).
