

Summary
我们描述了收集未受精
Abstract
许多生物的基因定位到特定的空间和时间控制基因表达的亚细胞目的地。最近的研究表明,大多数转录本地化细胞和胚胎的非随机的位置。识别本地化基因的方法之一,是生化净化蜂窝结构的兴趣,并确定所有相关的成绩单。使用最近开发的高通量测序技术,它现在是直接识别所有相关的RNA的亚细胞结构。为了促进转录的识别,有必要的工作与完全基因组测序的生物体。一个有吸引力的亚细胞结构的生化净化系统是从青蛙的非洲爪蟾卵提取物。但是,X.蟾目前没有一个全基因组测序,这阻碍成绩单识别。在这篇文章中,我们描述了一种方法从相关的青蛙,X.产生卵提取物热带,具有全基因组测序。我们提供微管聚合,纯化及成绩单隔离。虽然这篇文章描述了一个特定的微管相关转录识别方法,我们相信,它会很容易地应用到其他亚细胞结构,将提供一个强大的本地化的RNA识别方法。
Introduction
空间和时间上的基因表达的控制是非常重要的所有单元格,构图1的早期胚胎的控制是特别重要的。实现空间控制基因表达特定目的地在细胞或胚胎的mRNA通过积极的本地化。非常大的,在许多类型的细胞中,( 例如,卵母细胞,胚胎,和神经元)mRNA的定位是用来限制蛋白表达的编码蛋 白质的作用在网站上。由于一个本地化的mRNA可以催化蛋白质生产的多轮它是更有效的本地化比本地化单个蛋白质分子的mRNA。上本地化的mRNA通常翻译压抑,直到他们到达他们的目的地,这进一步限制本地化的编码蛋 白2。除了RNA的定位,以控制胚胎图案的许多记录的情况下,一些研究已经证明了本地化的基因在网站上的编码的蛋白质的行动。突出的例子包括本地化的β-肌动蛋白3和ARP2 / 3的mRNA能动的成纤维细胞的领先优势和本地化的许多有丝分裂器的mRNA减数分裂和有丝分裂纺锤体5-7。
母体效应突变通过遗传筛选确定许多本地化的mRNA的经典例子,后来确定本地化的RNA编码。然而,最近的全基因组的研究已经开始本地化的RNA的范围提供更广泛的洞察。最近的果蝇胚胎原位杂交屏幕显示〜70%的基因有一个具体的定位,包括许多新颖的目的地8。从小鼠成纤维细胞伪足的纯化确定不同群体的定位的mRNA 9。从本组的工作减数分裂异种使用生化纯化的微管脓鸡蛋提取数以百计的mRNA,共纯化与主轴5,7。我们的工作表明,大多数微本地化基因编码的蛋白质在有丝分裂的控制功能,支持这个想法,到现场行动的编码蛋白质的mRNA本地化。此外,检测mRNA的亚细胞部分生化净化富集能力突出的力量,这种方法鉴定本地化的mRNA。
大多数本地化的RNA使用主动运输,无论是肌动蛋白和微管骨架上,实现运输到最终目的地10。为了更好地理解的程度和类型的RNA定位到特定的目的地,使用生化方法,它是需要有一个体外系统,可以复述骨架过程。学习骨架生物学最大的系统之一是卵提取物生产未受精的卵子从青蛙的爪蟾 。 十蟾卵提取物已用于几十年来研究各种各样的细胞骨架过程,已经作出了很大的贡献,我们理解的机制和分子控制细胞骨架装配和动态11。此外,X.非洲爪蟾卵提取物是适合进行大规模纯化的微管相关蛋白12,13和精心设计的方法,用于生产各种类型的卵提取物14-16。然而,对于基因组研究X的使用仍存在一些缺点非洲爪蟾作为一个模型系统。
几十年来, 非洲爪蟾的青蛙是一个强大的系统研究的发展和细胞生物学,由于大量的卵母细胞的大小和强大的外部发展17。此外,卵提取物系统的发展,可以扼要重述许多细胞processe的S在试管中取得了这只青蛙,一个强大的实验模型。然而, 非洲爪蟾已经阻碍了缺乏一个完整的基因组序列,这已经放缓四倍体性质genome.In对比度,密切相关的物种, 热带爪蟾,在2010年18二倍体基因组被测序。虽然X。热带实验不听话为X非洲爪蟾17序列基因组的可用的一种具有吸引力的模型系统,以进行全基因组分析。
在这份报告中,我们描述了一种方法,使减数分裂II,抑制细胞生长因子(CSF)被捕提取从X热带 19。然后,我们介绍一个简单的方法,从这个提取纯化的微管和相关的RNA。的RNA可以被转换成适合使用最近开发的高通量测序技术测序的库。一旦库他们可以对齐的青蛙,以确定特定的mRNA相比总提取物的微管样品中富集的基因组测序。这提供了一个功能强大的方法检测微管目标mRNA定位在基因组范围内。在使用高通量测序和基因组测序除了能够检测本地化的mRNA,发现新的转录本,目前没有出现在公共数据库中的注释提供了可能性。
Protocol
1。一代X. 热带鸡蛋
从NASCO下令,所有在热带爪蟾的青蛙。我们的青蛙都装在一个水生栖息地循环水系统保持在27°C。水系统有很多选项,照顾十一些热带这只青蛙物种的的哈兰德与格兰杰实验室的网站上可以找到很好的通用信息( http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / )。我们的青蛙维持在由tankwater(0.4克Ciclid湖盐0.6克海盐,0.625克的NaHCO 3每公升的水,pH值7.0)20。这个配方的结果〜1800μS电导率,这是一个高盐度十热带 , 但是 ,我们已经发现,我们的青蛙氏茁壮成长环境和卵母细胞的质量得到改善。替代tankwater食谱中可以找到上面列出的资源一般十热带护理。
- 青蛙注射人类绒毛膜促性腺激素(HCG),连续三天,以刺激产蛋:首先,准备两种浓度的人绒毛膜促性腺激素的解决方案。的重悬10,000 U的冻干人绒毛膜促性腺激素1000 U / ml的终浓度为10毫升无菌去离子H 2 O粉。然后,淡化9毫升H 2 O 100 U / ml的终浓度为1毫升1000 U / ml的HCG解决方案。同时存储解决方案,在4°C
- 第1天,准备产蛋4-6青蛙注射HCG 2:00-3:00 PM之间。每个青蛙在背淋巴囊泄殖腔附近注入0.2毫升100 U / ml的HCG解决方案。青蛙快,在随后的两次注射量最小的青蛙废物目前产蛋期间,但是可选的。
- 第2天,注入相同的青蛙用0.2毫升100 U / ml的hCG的解决方案,下午2:00-3:00之间。
- 第3天,注入相同的青蛙用0.2毫升1000 U / ml的人绒毛膜促性腺激素的解决方案,在上午7:00-10:00之间。青蛙下蛋:填写6夸脱的塑料桶用新鲜tankwater,青蛙和地方在黑暗中,在25°C注射后,将开始产蛋4小时后,将完成7小时。青蛙下蛋的环境中维持在最低25°C。
- 提取解决方案和设备准备之前收集鸡蛋。
20X MMR:100毫米HEPES,pH值7.8; 2毫米EDTA pH值7.8; 2 M氯化钠,氯化钾40毫米,20毫米氯化镁 ; 40毫米氯化钙2。高压灭菌器,并储存于室温。准备1升1X MMR之前提取准备。
10X XB:100毫米HEPES,pH7.7; 10毫米MgCl 2 1毫米氯化钙 ,氯化钾1 M 500毫米蔗糖。在4°C的高压灭菌器和存储准备1升的1X预算外J乌斯提取准备前。 Dejelly解决方案:准备250毫升3%半胱氨酸溶液在去离子H 2 O和pH值7.8-8.0用10 N NaOH。刚准备提取准备前。
CSF-XB:取200毫升1X预算外,加2毫升0.5米EGTA pH值7.7和200微升1 M氯化镁2。刚准备提取准备前。
CSF-XB +:取50毫升CSF-XB,并加入50微升的LPC(10毫克/毫升的每支股票的亮肽素,胃蛋白酶抑制剂,胰凝乳DMSO)。加入50微升素D(10毫克/毫升DMSO)。刚准备提取准备前。
准备一个0.2%的明胶溶液去离子H 2 O,微波溶解和过滤消毒。在室温下保存。
预订2贝克曼2×½英寸超速管。
准备两个15毫升的玻璃圆底离心管,每0.5毫升H 2 O,以减轻超速离心机管。
请火抛光的玻璃巴斯德移液器。快结束5¾英寸的玻璃吸管,露出一个大开放,暴露于火焰平滑新暴露的枪头。
- 准备的500毫升的玻璃烧杯中,用于存储通过旋流周围0.2%的明胶溶液涂布烧杯壁鸡蛋。丢弃使用后的明胶溶液的烧杯中。
- 收集鸡蛋从塑料桶用于铺设6-7小时,第三次注射后第3天。如果需要,轻轻挤压每个青蛙一次得到任何剩余的鸡蛋。与的新鲜tankwater和转移到500毫升的玻璃烧杯中,涂有0.2%的明胶溶液洗涤一次蛋。
2。从X提取物的制备热带蛋
提取物的制备的所有步骤可以在室温下进行,约25°C。在整个洗涤,重要的是要保持蛋Ş淹没在液体中,让他们保持湿润。暴露在空气中能导致鸡蛋逃避细胞周期阻滞或溶解。
- 倒出多tankwater尽可能同时保留足够的液体,以保持湿鸡蛋。含蛋的一侧倾斜的烧杯中,烧杯中的壁慢慢加入约300毫升1X MMR,使物理搅拌的鸡蛋被最小化。让鸡蛋定居,然后倾出上清液含杂物。 十热带蛋拉丝,在该步骤中,,所以去除激活的蛋后完成dejellying。重复共三个1X MMR洗涤。
- Dejelly鸡蛋。倒出尽可能多的可能的和添加一半的dejelly解决方案,孕产妇死亡率。漩涡持续约5分钟。溶果冻大衣将上清液中可见,一两分钟后。倒出关闭添加其余dejelly解决方案。继续不断的漩涡,直到鸡蛋包非常紧密地和所有东方植物极(极白色颜料)向底部的菜。快速倒出尽可能尽可能多的dejelly的解决方案。一旦dejellied蛋,他们是非常敏感的机械操作。
- 小心地添加XB的鸡蛋。在第一次XB洗,去除去除溶解,浮肿,鸡蛋白,pseudocleavage鸡蛋逃脱CSF逮捕。活性炭X。热带蛋往往定居在顶部中心,所以使用塑料移液管,拉这些了。同时去除块皮肤和青蛙废物。洗净鸡蛋共三次〜300毫升1X XB溶液,轻轻摇动鸡蛋洗之间,并允许他们定居在烧杯底部。像以前一样,倾析各洗涤溶液尽可能同时保持蛋湿。
- CSF-XB,倒出鸡蛋洗净两次。
- 添加CSF-XB +鸡蛋。使用明胶处理火抛光的巴斯德吸管,转让CSF-XB +鸡蛋超离心管,照顾不公开电子GGS空气。 15毫升的玻璃离心管中的水垫内部发生。
- 自旋的鸡蛋在临床离心机在200×g离心1分钟,增加速度,以800×g离心和旋转持续30秒。
- 使用吸气删除尽可能多的缓冲,尽可能从鸡蛋。他们应该是在上面几乎是干的。快速移动蛋的SORVALL RC-6离心机配备了一个HB-6转子(或同等学历),并旋转17,000 XG 15分钟,在20°C。
- 取下黄色的细胞质层的颜料层和脂质层之间使用连接到1 ml注射器的18号针。穿刺侧管和慢慢拉动注射器管,取得细胞质提取物层。避免尽可能色素颗粒。
- 细胞质转移到新的超速离心管。这是正常的提取物在这一步出现略显混浊。 15毫升的玻璃离心管中与水垫内发生。再次离心17,000 XG 10分钟,在20°C重复分机18号针头raction。
- 细胞质转移到1.5 ml离心管。估价提取物的体积,并稀释到萃取液中的细胞松弛素D和LPC 1:1,000。拌匀,用1毫升的枪头,上下吹打很多次都没有引入气泡。从一个健康的青蛙殖民地是一个典型的产量约300-500微升提取物/青蛙。为了保持最大的活性,这是需要存储的提取物,在室温(20-25℃)进行实验操作。
3。从X净化紫杉醇稳定的微管热带提取物
- 紫杉醇加入100-200微升的等分试样提取物的终浓度为10μM,并在室温下孵育30分钟。对于对照反应,治疗等体积的提取物与的微管destablilzing的药诺考达唑(10μM)。预订100μl的未经处理的提取物进行分析。
- 稀释的药物治疗提取10卷BRB-80(80毫米的管道pH为6.8,1毫米氯化镁2,1毫米EGTA)+ 30%甘油。组装14毫升圆底聚丙烯管含10毫升BRB-80 + 60%甘油垫。使用大口径的枪头,层提取药物治疗反应轻轻顶部BRB-80 + 60%甘油垫。以17,000 xg离心10分钟,20℃时在Sorvall RC-6离心机配备了一个HB-6转子(或同等学历)和管适配器。
- 吸出上清液包含unsedimented的提取物,洗两次去离子H 2 O接口吸出剩余的缓冲量,慢慢地,不打扰凝胶样颗粒含有微管,微管相关蛋白,微管相关紫杉醇处理过的样品RNA在照顾。诺考达唑处理的样品不包含有形的物质。悬浮颗粒在1毫升的TRIzol和制造商的说明进行分离RNA。 Untreated提取物(最多100微升)可以直接重新悬浮在1毫升TRIzol试剂。
- 现在市售试剂盒准备转录图书馆RNA-seq的适合的。可购买这些通过http://www.illumina.com/ http://www.454.com/ 。
Representative Results
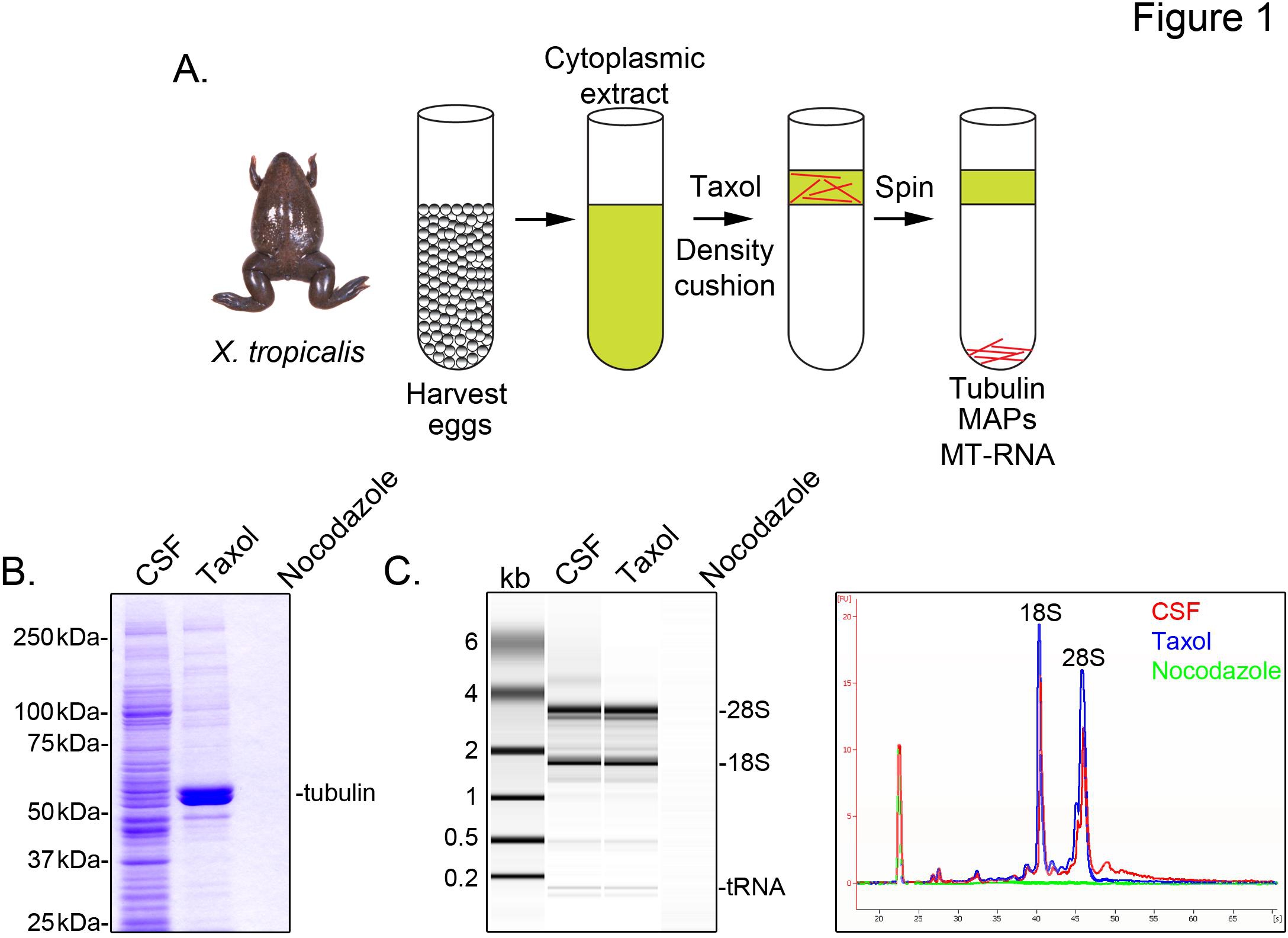
要识别X.与微管 有关的热带成绩单,我们准备胞浆提取物,未受精的卵子在中期减数分裂II(CSF)被捕。治疗允许这种提取物与紫杉酚稳定的微管的形成,通过甘油胶垫( 图1A),可以通过沉淀纯化。考马斯亮蓝凝胶分析证实,在这些准备工作( 图1B)收回的α/β-微管蛋白沉积物在紫杉酚依赖性,并代表主要蛋白质种类。较低级别的其他蛋白质也存在于紫杉酚的颗粒,但不与微管解聚药物诺考达唑治疗的制剂,表明紫杉醇的馏分具体联营公司(MAP的)与微管蛋白的。
安捷伦生物分析仪是用来检查一般RNA组成的,在所有的十热带提取部位( 图1C X种tRNA 蟾卵提取5,21。凝胶突起A线的痕迹显示mRNA的信号是显着地较低的微管含紫杉酚颗粒,最显着的,这表明该区域的迁移,28S rRNA基因的一个子集mRNA的cosediment与X中的微管热带 。以这种方式分离的RNA是适用于RNA,以下实验中使用市售试剂。

图1。 MT-RNA纯化RNA-SEQ(A)净化计 划,以隔离MT-RNA。收获卵女十卓picalis青蛙。细胞质提取物在制备后,加入紫杉醇诱导微管聚合。微管和MT-RNA纯化沉淀,通过甘油坐垫(B)考凝胶电泳分析蛋白(A)中描述的方案。总CSF提取物相比,蛋白质沉淀的存在紫杉醇或噻氨酯哒唑(C)生物分析仪凝胶电泳分析RNA分离使用(A)中描述的方案。 :核糖核酸,孤立的CSF中提取紫杉醇或诺考达唑的存在下相比,RNA沉淀。示凝胶投影线的痕迹。 转载夏普, 等人 (2011)的许可。 点击这里查看大图 。
Discussion
在这份报告中,我们描述了一个简单的方法来产生CSF被捕的卵提取物从X 19 热带和使用这种提取物研究微管相关的RNA 7。从X CSF-被捕卵提取物生产的基本步骤热带是为X所用的相同蟾几个关键差异。与X工作中最具挑战性的方面之一热带蛙获得足够的高品质鸡蛋提取物微核或纺锤体组装活动堪比十蟾卵提取物。为了达到最佳的产卵条件,同时防止打滑减数分裂II细胞周期阻滞,注射激素之间的时间间隔为X热带短于用于X。非洲爪蟾 ,定时从第三注射hCG的开始产卵也大为缩短。与X从注射hCG到t 蟾的时机他开始产蛋,它是便捷,高效的鸡蛋一夜之间变成被解雇缓冲。然而,由于铺设与X之间更短的时间注射hCG和蛋热带 ,经常需要手动表达从青蛙的卵。从两个不同的蛙卵提取物的另一个显着区别是一步dejellying。与X非洲爪蟾卵是如此之大,很容易确定果冻大衣观察密切鸡蛋间隔烧杯中溶解。开始的dejellying反应,卵开始收拾更加密集。然而, 十热带鸡蛋要小得多,它可以是相当困难的鸡蛋包装密度单独确定果冻大衣时已经解散。我们已经发现,最可靠的方法来确定果冻大衣溶解时是监视的动物(黑色)和植物(白)的磁极取向。当所有的植物极大利耳鼻喉科向烧杯底部的果冻大衣足够的继续进行,提取物已被删除。最后,而X。蟾卵提取物可以存放在凉爽的温度(4-12℃),我们观察到,关键是要保持十热带卵提取物在室温下(20-25℃)在制备过程中和实验操作保存生化活性。由于在易用性方面的差异,我们更倾向于使用X。非洲爪蟾蛙卵提取物的生产。然而,对于实验中需要或促进一个有机体具有的基因组测序,X.热带是一个很好的替代系统。
本报告中所述的方法,我们已经使用作为微管稳定剂紫杉醇诱导微管聚合。我们选择这种方法,因为紫杉醇是一个强大的微管稳定剂,有利于大规模分离纯化的微管。该方法的第在我们所描述的,也可能会提高,通过比较与微管聚合方法替代使用的微管相关蛋白和RNA。替代方案可能包括聚合GTP引发聚合(经典的技术),22或使用RAN-GTP微管聚合模仿引起微管染色驱动主轴组件23。最后,使用纯化的精子细胞核诱导微管的聚合反应将是最接近的模拟类型的微管在有丝分裂过程中成核(中心体,染色质,和着丝粒介导的)。这些微核的替代来源的缺点是成核剂是不那么容易紫杉醇,他们没有核或稳定微管,高效紫杉醇。因此,这些方法将更加难以用于大规模的纯化。比较多个不同类型的微管成核剂的优点是,它可能是未能识别蛋白质和/或RNA的特定微管成核每条通路。
我们在这里所描述的方法需要利用两栖动物细胞质提取物。然而,这种方法可以扩展到从其他生物体中提取系统使用。有丝分裂提取物已被描述的同步的人体组织培养细胞24,忠实地复述微管装配的许多方面。我们已经成功地使用这些提取物的识别微管相关的RNA的HeLa细胞5。类似的微管纯化方案已描述了许多不同的生物25,26,虽然尚未审查的微管相关的RNA。这里描述的方法可用于与任何的有机体可以制作成核微管的能力的胞质提取物的浓缩。
最后,虽然方法,我们这里讨论隶微管相关蛋白和RNA的净化,这种方法可以推广到其他亚细胞结构。虽然大多数本地化的mRNA尚未确定采用生化方法在DNA和RNA测序技术的最新研究进展使这种方法的一个有吸引力的方法来确定本地化的RNA。在这种方法中,任何感兴趣结构的亚细胞或胚胎子可被分离或纯化。则相关联的蛋白质和RNA,可以在基因组范围内确定。的RNA可以进行比较,以总的细胞或胚胎,找出丰富的本地化的RNA的RNA含量。此方法可用于全蛋(动物和植物分离,类似的方法,确定了第一个局部的RNA在爪蟾 27日 ),肌动蛋白相关的RNA,ER相关的RNA,线粒体相关的RNA,或任何亚细胞结构,可以完好相关的RNA进行纯化。基于微管相关RNA我们的工作,我们预测,这将是一个很好的方法来发现新功能的蛋白质,在一个给定的位置。此外,标识的位置和程度,所有的本地化的RNA,将提供洞察细胞和胚胎如何使用mRNA的定位控制基因表达。
Disclosures
没有利益冲突的声明。
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D.
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al.
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}