

Abstract
肝実質細胞のほかに、肝臓は非実質細胞(NPC)、すなわちクッパー細胞(KC)、肝臓内皮細胞(LEC)と肝星細胞(HSC)で構成されています。初代ヒト肝細胞(PHH)の二次元(2D)培養物は、依然として薬物代謝および肝毒性のin vitro試験のための「ゴールドスタンダード」とみなされます。それはPHHの2D単作が脱分化と機能の損失に苦しんでいることはよく知られています。最近、肝NPCは、肝臓(病態)生理学およびPHH機能の維持に中心的な役割を果たしていることが示されました。現在の研究は、2Dの単一栽培の限界を克服するための3D-と共培養モデルによるインビボ組織構造の再構築に焦点を当てています。以前に我々は、ヒト肝細胞を単離する方法を公開し、実験生物学および医学1における細胞培養での使用のために、これらの細胞の適合性を調べました。このテにおいて幅広い関心に基づいて、この記事の目的をchniqueすることは、この手法を簡単に再現できるようになります、ビデオ、などの肝細胞の単離プロセスのためのより詳細なプロトコルを提供することでした。
ヒト肝細胞は、二段階EGTA /コラゲナーゼP灌流技術により外科的介入のヒト肝組織サンプルから単離しました。 PHHは、50×gで最初の遠心分離によってNPCから分離しました。密度勾配遠心分離工程は、死細胞の除去のために使用しました。個々の肝細胞集団は、特定の細胞の性質、細胞選別手順を使用して濃縮されたNPCの画分から単離しました。 PHH分離のほかに、私たちはさらなる培養のためのKC、LECとHSCを分離することができました。
まとめると、提示されたプロトコルは、1つのドナー組織サンプルから高品質と量のPHHとNPCの単離を可能にします。精製された肝細胞集団へのアクセスは、人間の李のようなin vivoでの作成 が可能性があります版モデル。
Introduction
ヒト肝組織は非常に複雑であり、2つの異なる細胞エンティティ、実質細胞と非実質細胞(NPC)から成ります。肝実質細胞は肝細胞と胆管を含みます。肝細胞は、全肝細胞の60〜70%を表し、代謝、肝機能のほとんどを占め、 例えば、胆汁酸および因子合成、生体内変化とエネルギー代謝2,3を補完します。
より小さいNPC画分は、全肝細胞の30〜40%を構成します。 NPCは、異なる細胞集団、すなわち、クッパー細胞(KC)、肝内皮細胞(LEC)と肝星細胞(HSC)が挙げられます。この異種細胞画分は、肝臓の生理学的プロセスにおいて中心的な役割を果たしています。さらに、NPCは、肝硬変4などの慢性肝損傷、急性肝障害を仲介する、 例えば、薬剤誘発性肝障害(DILI)に参加するだけでなく、。
近年では、時間UMAN肝細胞はますます不可欠な研究開発薬物試験の、薬剤開発および肝疾患における新たな生化学的経路の同定になっています。 インビトロ試験のためにPHHの単一栽培は依然として「ゴールドスタンダード」として考えられている5。現在の同型肝臓モデルの主な制限は、数日4内の脱分化と肝細胞の機能の喪失です。 3次元(3D)培養技術の確立は、これらの制限は、4,6を補償することができることを示しています。しかし、現代の3D培養技術は、アクション7のすべての肝毒性のモードを表示することができません。既存のin vitroモデルで不足しているNPC集団は、in vivoでの状況にこの矛盾の可能性のある理由として議論されています。それにもpathophyにおける異なる肝細胞集団との間の細胞間コミュニケーションは、生理的恒常性において中心的な役割を果たしていることが示されましたsiologicは8を処理します 。したがって、科学的な関心は、NPCとそれらの細胞間相互作用にますます焦点を当てています。共培養および組織工学システムにおけるそれらの意図的な使用はできるだけin vivoでの状況に近いですインビトロ肝モデル8,9の高い需要のためのソリューションである可能性があります。
現在、主な課題は、PHHとNPCの明確に定義された部分が含まれている標準化されたヒト肝共培養モデルの開発です。その結果、非常に異種肝細胞の分離技術が必要とされ、それらは、純粋な細胞集団を得るために最適化されなければなりません。 PHH分離のための標準化されたプロトコルが10を存在するが、人間のNPCの標準化された単離は、まだ開発中です。ほとんどの公開されたNPCの分離プロトコルは、非ヒト細胞11,12を用いた実験に基づいています。ごく少数の出版物は、人間のNPCの分離プロセスを記述し、ほとんどがカバーのみ単一の細胞型11〜16の単離のための方法。細胞分離のために活用されている最も重要な細胞特性が大きさ、密度、付着挙動、および表面タンパク質の発現です。これらの特性に基づいて、我々は実験生物学と医学 1で以前に出版されたPHH、KC、LECとHSCを単離するために簡略化されたプロトコルを開発しました。そのため、この技術において幅広い関心が、この記事の目的は、より簡単な技術を再現することができますビデオを含む、肝細胞の単離プロセスのためのより詳細なプロトコルを提供することでした。プロトコルはまた、収量と生存能力の評価のためだけでなく、特定の免疫染色を用いて同定および純度評価のための品質管理方法を含みます。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
注:すべての細胞を、一次または二次肝腫瘍を有する部分的な肝切除後に残っ切除非腫瘍性ヒト肝臓組織から単離しました。 Universitätsmedizinベルリン - 患者のインフォームドコンセントをシャリテの倫理指針に従って得ました。
材料とソリューションの作製
- 分離プロセス中に細菌汚染を避けるために、事前にすべての機器および材料を滅菌します。
- 新たに調製された消化液を除いて、 表1及び2に記載の肝臓組織試料、肝細胞および非実質肝細胞の単離方法と初代ヒト肝細胞の培養の灌流のために必要な溶液を調製使用前に。全ての溶液は4℃で保存することができ、調製後4週間以内にそれらを使用することが推奨されます。
- 殺菌します0.22μmのボトルトップフィルターを用いて全ての溶液。
灌流機器の作製
- 図1Aに示すように、肝臓組織試料の灌流および消化のための機器を設定します。
- 灌流および消化中に最適なコラゲナーゼPの活動を確保するために39℃に水浴温度を調整します。
肝臓組織試料の3灌流および消化(1.5時間)
- 切除した肝臓組織から無傷のグリソン鞘と組織サンプルを選択します。組織サンプルを切断すると、良好な可視隻と小型の切断面を得ることを試みます。輸送および灌流まで氷上で肝臓組織試料を処理することにより、暖かい虚血時間を避けてください。
- 無菌条件下で組織重量を取り、層流中でペトリ皿に肝臓組織試料を配置します。再から滅菌圧縮して組織試料の表面を清掃してください血液をmainingと私はすべてのカニューレが浸透したことを確認するために、1×灌流-Solutionを使用してカニューレセットをフラッシュします。
- いくつかのより大きな血管にカニューレのオリーブを修正するために組織接着剤を使用してください。肝臓組織試料の大きさ及び表面の血管の数に応じて、3~8カニューレで設定カニューレを使用します。灌流をテストし、漏れをチェックします。組織接着剤で、明確な1×灌流溶液Iを漏らすすべての血管を、閉じます。
- その穴のあいたフィルターディスク( 図1A)上のブフナー漏斗にカニューレ肝臓組織試料を置きます。
- 7.5ミリリットル/分であり、使用されるカニューレの数と肝組織の抵抗に応じて14.6ミリリットル/分の蠕動ポンプの流量を設定します。現在しかし遅い灌流があることを確認するために流量を毎回調整します。全血を流し出さないが、少なくとも20分されるまで、組織を灌流。良いperfusi地域で明るくなって組織を観察に。
注:いくつかのケースでは、プラスチッククランプを有するカニューレの一方をクランプするために必要であり得るか、または灌流を最適化するために、肝臓カプセルに対してスパチュラで静かに押して領域内の圧力を増加させます。光茶色がかった色に黄色の光への完全な色の変化は、良好な灌流を示しています。 - コラゲナーゼP( 表1)を含む消化液に灌流液を変更します。
- 消化段階のセットアップ( 図1A)を並べ替えます。そのため、最大15分間、 図1Bによれば消化-ソリューションの円形の流れを行います。
注:肝臓組織サンプルが十分に消化されるとき、すぐに灌流を停止することが重要です。良好な消化は、それがスパチュラで押されると、カプセルの変形を維持することによって評価されるような組織は、弾力性の兆候を示していない場合、観察することができます。
肝細胞の単離4(1時間)
- Tオフ蠕動ポンプをURNとガラス皿に肝臓組織試料を配置します。氷冷ストップソリューション( 表1)を用いて組織試料の外側を洗浄します。肝臓組織試料からカニューレを外します。カニューレが取り付けられていたエリアの中央に切開することにより、肝臓組織のサンプルを開くにはメスを使用してください。 Glisson'sカプセルが無傷のままで注意しておいてください。
- 組織試料の内部を洗浄した後、氷冷停止溶液で全体の組織サンプルをカバーしています。組織から細胞を解放するために穏やかに組織を振ります。
- 細胞懸濁液を収集し、50mlのプラスチックチューブに視線漏斗(ガーゼ湿布を敷いたプラスチック製の漏斗)を介して、それをフィルタリングします。 500 mlの最終容量が消費されるまで、肝臓組織試料に対して、より停止溶液を加えます。
- 遠心分離機50×gで、5分間、4℃で細胞懸濁液。後で非実質細胞の単離のために上清を収集します。 ( 図2のPBSで細胞ペレットを洗浄A)。
- 遠心分離機50×gで、5分間、4℃で再び細胞懸濁液。上清を収集し、肝細胞インキュベーション培地中で再懸濁ペレット( 表2、 図2B)。
- トリパンブルー染色を用いて得られた細胞懸濁液中の細胞数及び生存率を決定します。ノイバウアーカウントチャンバー内の生活と死細胞を数えます。以下の式を用いて細胞数、生存率およびPHHの収量を計算します。
収量(計数した細胞)を細胞懸濁液(ml)を計数した細胞のX希釈因子X量が10,000 X =
収量(肝細胞/(g肝臓組織試料))=(細胞懸濁液(ミリリットル)の収率(肝細胞/(mlの培地))×体積)/(肝臓組織試料の重量(g))
生存率(%)= 100%X(生細胞数)/(総細胞数)
肝細胞の5精製(1時間)
注:この精製工程が推奨され、生存能力場合70%未満です。
- 氷の上のすべての手順を実行します。密度勾配遠心5mlの密度勾配溶液を、15 mlのPBSを混合することにより25%の密度勾配を準備します。
- 両層の明確な分離が( 図2C)が達成されることを保証するために、25%の密度勾配層の上に慎重にゆっくりと肝細胞が豊富な細胞懸濁液のうち、合計で50澪セルの最大を入れてください。 1,250×gで、20分間、ブレーキなしで4°C( 図2D)で遠心分離し、遠心分離に慎重にチューブを入れてください。
- 残りの細胞懸濁液と間期における死細胞を吸引します。脂肪含有量に応じて、1つはまた、密度勾配液を吸引する可能性があります。
注:PHH低脂質含量を持つが密集し、ペレットと密度勾配を完全に吸引することができますを形成します。高脂質含量のPHHはより拡散してペレットを形成し、生存細胞の多くは、ペレット、上記密度勾配溶液中に残ることがあります。 - 50 XG、5分、4℃で再びPBSと遠心分離と肝細胞ペレットを再懸濁します。ペレットをプール、再びPBSで洗浄し、肝細胞インキュベーション培地中に精製されたPHH再懸濁します。ステップ4.6で説明したように細胞計数を行います。
肝細胞の6.栽培
- 例えば、ラット尾コラーゲン(コラーゲンタイプI)は、細胞外マトリックスでそれらをコーティングすることによってPHHの播種のための細胞培養皿を準備します。ラジャンらによって確立されたプロトコルに従って、ラット尾コラーゲンを準備します。17
- PBSで200:ラット尾コラーゲン原液1を希釈します。表面全体が覆われている培養皿への転送を100μl/ cm 2のラット尾コラーゲン溶液、世話。室温で20分間、細胞培養プラスチックをインキュベートします。残りのラット尾部コラーゲン溶液を吸引します。
- 培養dishe上の肝細胞インキュベーション培地中の種子15×10 4肝細胞/ cm 2のラット尾コラーゲンでコーティングされたのです。少なくとも4時間、37℃、5%CO 2の加湿インキュベーター中で細胞を培養。 4時間後、肝細胞は、付着した培地を変更することができます。
- 実験に応じて調査を行います。 48時間の培養時間は、細胞が単離プロセスから回復することを可能にすることをお勧めします。
非実質肝細胞の7単離(1.5〜2時間)
- 遠心分離機残りの赤血球および肝細胞を排除するために72×gで、5分間、4℃で回収した上清(ステップ4.5および4.6)。プール上清および回2細胞ペレット得るためにそれらを遠心:残りKCの沈降のためにHSC、LEC、一部はKCと650×gでの沈降のための300×gで、5分、4°C、7分、4°Cを。
- プールペレットおよびHBSS中で再懸濁それらの両方。密度勾配centrifための密度勾配溶液とPBSを混合することにより25%と50%の濃度勾配を作製ugation(25%の密度勾配溶液5 mlの密度勾配溶液を、15 mlのPBS、50%の密度勾配溶液の調製:10mL密度勾配溶液と10mlのPBS、 図2を参照)。 50%の密度勾配溶液層の上に慎重に25%の密度勾配溶液を置き。
- 両層の明確な分離が達成されるようにして25%の密度勾配溶液層の上に慎重にゆっくりとNPCのサスペンションを置きます。
- 遠心ブレーキなしで1,800×gで、20分間、4℃での密度勾配上での細胞懸濁液( 図2.2)。
- 最上層から吸引し、死細胞と細胞の破片。 NPCは、25%と50%の密度勾配層( 図2)との間の相間に配置されています。 NPCを収集し、HBSSと遠心機で上記の二重遠心分離工程(ステップ7.2。)を適用し、細胞懸濁液を、それらを洗ってください。
クッパー細胞の8分離(アドヒアランス分離工程)(1時間)
- ステップ4.6で説明したようにNPC画分中のKCのための細胞数を実行します。 (懸濁液中のKCの外観について図3Bを参照してください)。遠心分離培地( 表2)を播種クッパー細胞における上記の二重遠心分離工程(ステップ7.2)と再懸濁NPCとNPC画分。
- 5×10 5 KC / cm 2の密度で、プラスチック製細胞培養容器のKCを含有する画分をシードします。 37℃、5%CO 2の加湿インキュベーター中で20分間KC培養物をインキュベートします。プライマリKCは時間の短い期間( 図2.3)内の細胞培養プラスチックに付着します。
- 主にLECとHSCからなる、ない付着NPCを含む上清を収集します。プールLEC(セクション9を参照)、HSCの後の分離のための上清(セクション10を参照)。 HBSSで接着KCを洗浄し、37℃、5%CO 2でクッパー細胞培養培地( 表2)でそれらを養います加湿インキュベーターで2。
内皮細胞の9分離(1.5時間)
- 遠心分離機300×gで、5分間、4℃で回収した上清(ステップ8.5)。 PBSでペレットを洗浄。 300×gで、5分で遠心分離した後、4℃の星細胞/内皮細胞の分離媒体中で細胞を再懸濁し、ステップ4.6で説明したように、残りのすべてのセルに対して細胞計数を行います。
- 免疫標識のためのMACS-KITからブロッキング溶液を20μlおよびCD31マイクロビーズの20μlを添加、1ミリリットル星状細胞/内皮細胞の分離媒体中1×10 7澪細胞を再懸濁し、4℃で15分間、得られた懸濁液をインキュベート℃の温度( 図2.4)。
- 磁気活性化細胞選別システムMACS( 図2.5)のためmanufacturer'sプロトコルに記載されているように、HSCは別にLEC。磁気保持CD31陽性LECを溶出し、星状Cでそれらを一時停止エル/内皮細胞培養培地( 表2)。
- ステップ4.6で説明したようにLECのための細胞計数を行います。ラット尾コラーゲンでコーティングした細胞培養容器で1.25×10 5細胞/ cm 2の密度でシードLEC(ステップ6.1参照)。加湿インキュベーター中、37℃で、5%CO 2で細胞を養います。
星細胞の10分離(0.5時間)
- 非標識のHSCは、MACS手順の間に分離カラムを通過します。 (ステップ9.5、 図2.5を参照)HSC画分を収集します。ステップ4.6で説明したように細胞計数を行います。
- 星細胞/内皮細胞培地( 表2)(ステップ6.1参照)ラット尾コラーゲンでコーティングした細胞培養容器中で5×10 4細胞/ cm 2の密度でHSCを播種し、37℃、5%でそれらを養います加湿インキュベーター中のCO 2。
表1: 灌流および分離ソリューション。
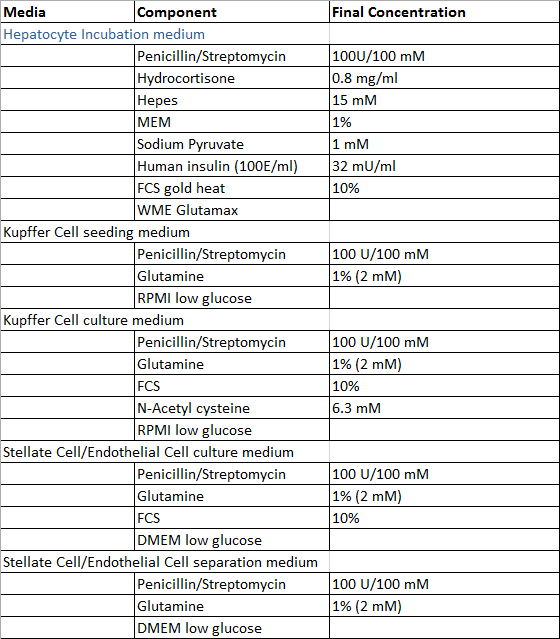
表2: 文化との分離媒体。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
接着特性およびMACSの使用と組み合わせクリーンアップ手順として密度勾配遠心分離を用いて実質および非実質画分への分離は、成功したPHHとNPCの分離につながります。 PHHとNPCは、高品質と量で単離することができる。 図1は、肝灌流および消化のための機器の代表的なセットアップを示しています。プロテアーゼのタンパク質分解活性を減少させるために、リターンにコラゲナーゼP活性を安定化させるために溶液II - 10%FCSを灌流を含むコラゲナーゼPを添加しました。結果としてNPCの分離のために必要なより長い消化時間がPHHの生存率に負の影響なしに適用することができます。

図1: 灌流消化セットアップ最初の灌流工程は、オードで行われますrは、残留血液を除去する組織を温めると1xの灌流溶液I(PI)(A)を用いて、細胞-細胞-接続を溶解するためのCa 2+を除去しました 。消化溶液(DG)の再循環は、灌流工程II(B)中に、肝臓組織の消化のために行われます。

図2: 完全PHHとNPC分離プロセスの単純化した略図で 実験生物学と医学の許可を得てファイファーら 1、2014から変更されました。。。まず、肝臓組織のサンプルは、灌流および二段階EGTA /コラゲナーゼP灌流法(A)によって消化されます。得られた細胞懸濁液をsupern(より小さいNPC-画分から大きくPHH画分(ペレット)を分離するために、50×gで、5分間、4℃(B)で最初に遠心分離しますatant)。 70%以下PHH生存率の場合には、実行可能なPHH分率は死んだが、チューブの底にPHHの沈降をもたらす1,250×gで、20分間、4℃(C)での密度勾配遠心分離によって濃縮することができます細胞/細胞残屑は、密度勾配層(D)の上部に配置されています。 1)300×gで、5分間、4℃、2)650×gで7分間、4°C:最初の遠心分離(1)の回収した上清は、2つのステップを使用して遠心分離します。最初の遠心分離後、KCは、部分的に上清中に位置しています。この文脈において、第2の分離工程が必要です。得られた細胞ペレットをプールし、HBSS中に再懸濁させます。その後、細胞懸濁液を慎重に二層(25%/ 50%)の密度勾配の上に積層されています。層状密度勾配管を1,800×gで、20分間、4℃で遠心分離する(2)。 25%の密度勾配層の上死細胞は廃棄されます。 25%の間期と50%Dとの間に位置するNPCensity傾斜層を収集し、プールします。 NPC画分はコーティングされていない細胞培養プラスチック上に播種されます。 20分のインキュベーション時間を用いて(密着分離工程)KCは他の肝細胞集団から分離される(3)。 LECとHSCは、MACSキットを使用することによって分離されています。したがって、上清に回収残りの肝細胞を300×gで、5分間、4℃で遠心分離し、CD31結合マイクロビーズ(4)で標識されます。唯一のCD31陰性HSCは、MACS分離カラム(5)を渡します。 CD31陽性LEC列に固執します。最後に、カラムを磁気デバイスから削除され、CD31陽性のLECは、カラムから溶出される(5)。 この図の拡大版をご覧になるにはこちらをクリックしてください。
孤立したPHHは14.2×10 6±6.6×10 6生PHH / g肝臓組織の収量と生存率を示し、約760.6±4.2%( 表3 1)。顕微鏡で目に見える特徴は、脂肪滴と1の間と4核、( 図3A)との組み合わせで、典型的な大細胞質体積でした。セルサイズは、懸濁液中〜30μmの20との間を変化します。
KCはNPC画分の中で最も一般的な細胞型でした。私たちは、92.8±3.5%( 表3 1)の生存率は約1.9×10 6±0.2×10 6生KC / g肝臓組織を単離しました。 KCは、表面上の低細胞質/核比、典型的な微絨毛( 図3B)と非常に小さなセル(約5μm)です。
最後に、我々は残りのCD31陰性HSCからCD31陽性LECを分離するためMACS分離技術を使用していました。 LECの収量は約2.7×10 5±0.1×10 5生存可能LEC / gでLIVましたER組織と達成生存率は95.6±2.8%( 表3 1)でした。識別基準は、短い培養時間( 図3G)の後に、複数のgranulaの複数形と細胞懸濁液中に約10μmの大きさ( 図3C)と同様に特徴的な紡錘形です。
分離工程は、89.6±3.8%( 表3 1)の生存率は約4.7×10 5±0.2×10 5生存可能なHSC / g肝臓(N = 8)HSCの収量が得られました。顕微鏡的に識別可能な特徴は、約20μmの大きさや脂肪滴( 図3D)の様々な量の典型的な粒状化出現しました。
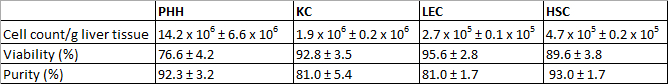
表3: 孤立PHHとNの収量、生存率および純度PC。三つの異なるドナーを評価しました。データは平均±SDとして与えられます。この表は、ファイファーら 1、2014年以前に出版されたと実験生物学や医学の許可を得て転載されています。
細胞培養の純度の識別と決意するために、すべての単離された細胞画分は、細胞型特異的抗原に対する抗体で処理しました。細胞は、蛍光二次抗体で処理し、免疫蛍光顕微鏡法によって調べました。純度はヘキスト染色により視覚化し、総細胞数との関係で正の蛍光染色された細胞を計数することによって決定しました。
24後培養PHHの時間特性多角形状としばしば倍数( 図3E)を示しました。 PHHは、CK 18( 図3I)が陽性であったと92.3±3.2%の純度を示し( 表3 1)。
KCは、細胞培養プラスチック表面に20分以内に接着しました。著名な丸い細胞核を持つ24時間小円形細胞のインキュベーション時間の後( 図3F)が観察されました。表面タンパク質CD68は、KC( 図3J)を同定するために使用しました。 CD68陽性細胞の純度は81.0±5.4%( 表3 1)に達しました。
NPCの分離中にCD31のラベリングを用いたMACS分離にもかかわらず、CD31で孤立したLECを染色することが可能でした。したがって、単離および培養LECは、同定および純度を決意するためにCD31で染色しました。さらにLECは、間葉細胞マーカーのビメンチン( 図3K)のための免疫反応性を示しました。私たちは、ポジティブ染色された細胞( 表3 1)の約81.0±1.7%を観察しました。 その典型的な著名な脂肪滴とHSC( 図3H)は、GFAP( 図3L)のための免疫蛍光染色によってマークされていました。 HSCの純度は93.0±1.7%( 表3 1)でした。
すべての細胞画分は、他のNPCマーカーで対比しました。すべての細胞画分は、他の肝臓特異的細胞型の少数を含有するが、肝細胞マーカーCK18及び胆管マーカーCK19陰性でした。

図3: 人間の実質と懸濁液中の非実質肝細胞のおよび付着後の形態左の列(A - D)を直接分離工程の後に異なる孤立した肝細胞集団を示しています。位相差顕微鏡ビューで:PHH(A)、KC(B)、LEC(C)、およびHSC(D)。真ん中の列(E - H)単離されたの画像を提示し、培養(位相差顕微鏡)の24時間後PHH(E)、KC(F)、LEC(G)とHSC(H)を培養しました。異なる細胞画分の免疫蛍光ベースの特徴付けは、最後の列に示されている:PHHは肝細胞マーカーCK18(I、単離後24時間)に陽性シグナルを示し、KCは、マーカーCD68(J、単離後24時間)が陽性でしたLECは、ビメンチン(K、単離後72時間)とHSCのための陽性シグナルは、GFAP(L、単離後72時間)に陽性であることを示しました。細胞核はヘキストで染色しました。倍率:400X。 EXPERの許可を得てファイファーら 1、2014から変更されましたimental生物学と医学。 この図の拡大版をご覧になるにはこちらをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
公開されたプロトコルは、同時にヒト肝組織の同じサンプルから高品質と純度で、純粋なPHHやNPC、すなわちKC、HSCおよびLECを分離する技術が記載されています。 (ダムらによってレビュー。)ヒト組織を用いて行ったものの細胞集団18-20と単離手順のいずれか一方のみをカバーする肝細胞の単離を扱う出版物の大半はまれである21。ヒトの肝臓への動物組織( 例えば、ラット肝臓)で確立された方法の適応は、動物およびヒトの細胞集団との間のセル特性のいくつかの違いが明らかになりました。別の肝細胞集団を覆う肝細胞単離法の確立が実質および非実質細胞の分離を組み合わせることにより最適化された結果を得るために必要な消化時間の差に重要なステップであることを明らかにしました。この課題に直面して、我々は異なる技術を組み合わせた肝細胞単離プロトコルを開発し、適応しました私たちのPHH単離プロトコール10 SE。
溶液IIを含むコラゲナーゼPの10%FCSを添加することによって、我々は、プロテアーゼのタンパク質分解活性を低下させることができました。この変更は、NPCの高い数値を得るために必要な長い消化時間を、許可されました。その結果、我々は良い品質のNPCとして、高量でも同様PHHを単離することができました。成功した肝細胞の単離は、最初の組織の質に強く依存します。ドナーデータおよび既往歴、細胞の質と量に影響を与えることができます。我々の経験から、ドナー特異的因子との相関がないとも非常に経験豊富なスタッフが、失敗した細胞の単離に直面することができます。ドナー組織の品質が臨界点であるため、組織の品質を損なう可能性のあるすべての外部ソースは、最小化されなければなりません。
最も重要な要因は、組織トンの輸送中に外科的介入と冷たい虚血時間後に暖かい虚血時間です研究室O。さらに、細菌感染症のための任意のソースは避けるべきです。時には器官自体は、胆汁管疾患の場合には、例えば、細菌汚染を含むことができることに留意しなければなりません。単離手順中の重要なステップは、灌流時間および密度勾配遠心分離工程をカバーします。最初の灌流工程は、20〜30分を持続させるべき。短い灌流時間は、得られる細胞懸濁液中の細胞塊の発生が得られた細胞 - 細胞接触の不完全な剥離をもたらし得ます。長時間の最初の灌流は、細胞の生存率を低下させ、原因の Ca 2+枯渇細胞ストレスを誘発します。
酵素組織消化のために行われる第2灌流のステップは、右の時点でタンパク質分解反応を停止に関する決定を行うための最適な消化の程度を決定するために、いくつかの経験が必要です。短い消化時間は、ストレスと細胞の損傷をセルに低利回りと長期の消化時間につながります。我々の経験から、消化中の未消化の組織と損傷組織との間の時間枠は、多くの場合、1-3分のウィンドウ内にあります。不完全な灌流は、カニューレをピンチオフするためのクランプを使用して別の領域に組織内の圧力をシフトすることによって対抗することができます。さらに、組織サンプルに向かってへらでソフトな圧力は、組織灌流の変化につながります。組織が圧縮され内圧が上昇します。その後、血管にも圧縮されて、それらの半径が小さくなります。半径の減少と、抵抗が増大(ハーゲン・ポアズイユの法則)と灌流液は、最小の抵抗と灌流他の分野の方法を好みます。 Baccaraniおよび共同研究者によれば、我々は、線維症または肝硬変の組織は、細胞生存率22を低下につながる長い消化時間を必要とすることを観察しました。このような理由から、我々は、肝線維症または肝硬変患者の組織を回避することをお勧めします。
準備と勾配のうち密度勾配の取り扱いだけでなく、収穫細胞は(ステップ5を参照し、7)も重要な段階をカバーしています。異なる密度勾配層と細胞懸濁液は、鋭いinterphasesの作成をゆっくりと慎重な方法で転送されなければなりません。さらに、特に細胞を回収しながら、取り扱い中に勾配を損傷する危険性が常にあります。 NPC分離の間の密着分離工程は、全てのNPCの分画の後の収率および純度のために重要です。付着していないNPCとKCの純度の細胞数を増加させるために追加の洗浄ステップが役立つことがあります。この段階の洗浄溶液をさらにNPCの分離のために上清と共にプールします。細菌による汚染を避けるために、真菌またはウイルス厳密な無菌状態が23を確保しなければなりません。
必要な細胞集団に依存して、プロトコルは、特定のステップをスキップすることによって変更して調整することができます。例えば、唯一のKCがある場合NPCの沈降のための二重遠心分離工程は、2回目の遠心分離ステップに低減することが可能とHSCとLECの分離のための手順を落とすことができる必要がありました。潅流時間、遠心分離のためのg力は、組織および細胞質に依存して変化させることができます。線維性組織は、したがって、慎重に組織の弾力性を制御する必要があり、長時間の消化時間を必要とします。脂肪肝細胞における脂肪滴の蓄積は、細胞密度を減少させ、したがって、沈降特性を変化させます。我々の観察によれば、高いPHH量が必要とされるPHHの脂質含量に応じPHH分離中のg力を調整するのに有用であり得ます。これは、最初の遠心分離工程の任意の変化が負質および量の観点でNPCの分離に影響を与えることに留意しなければなりません。これまでのところ、我々は50 XG(低脂肪含有量の肝細胞)および150 XG(高脂肪含有量の肝細胞)との間でg力をお勧めします。さらに、脂肪HEPAtocytesは、密度勾配遠心分離の後に以下のコンパクト細胞ペレットを形成する傾向があり、さらに、細胞採取は、ステップ5.5に記載されるように変更されなければなりません。
単離手順をスピードアップするためにいくつかのステップを同時に行うことができます。単離肝細胞の精製と並行して、例えば二人目は、NPC分離で始めることができます。さらに密度勾配溶液を予め調製することができます。以上の二人が存在する場合でも、複数のステップを同時に行うことができます。
実験生物学と医学 1で以前に公開されたように、他のヒト肝細胞単離プロトコルと比較して我々の結果は、同等以上の細胞の収量と生存率を示しています。 KC分離Alabrabaと共同研究者は、に匹敵する約98〜13%の生存率と組み合わせる2.3×10 6生KC / g肝臓組織の収率で単離結果を実証するために私たちのKC結果(細胞数:1.9×10 6生KC / g肝臓組織、生存度約93%)。公開されたLEC分離データの大部分は、全体の臓器15,24からの単離を記載しています。ゲルラッハと共同研究者だけでなく、10 3、10 6細胞/臓器15,24の間Lalorや同僚単離された細胞の数。これらのデータは、組織サンプルから単離、細胞に直接比較することはできません。しかし、私たちのプロトコルを使用して、我々は臓器全体に外挿したときにはるかに大きいです2.7×10 5生存可能LEC / g肝臓組織のLECのための収量を示しました。 HSCは、約4.7×10 5生存HSC / g肝臓組織と生存率は約90%の収率で単離しました。フリードマンと共同研究者によって公開された既存の結果は、半分より低い細胞収量(2.3×10 5 HSC /グラム肝臓)を示したが、同様の純度(91%)14。私たちのプロトコルに関しては、低い細胞収量が悪く灌流および低による消化または1x灌流-Solutioの無循環によって引き起こされる可能性がnはIおよび組織内の消化・ソリューション。さらに、組織内の気泡は、1×灌流溶液Iと消化・ソリューションの組織内循環を乱すことができます。これらの場合において、灌流は、単一のカニューレをクランプおよび/または組織試料を押圧するためのスパチュラを用いて気泡の灌流圧と除去を増加させることによって改善することができます。悪い生存率はほとんどの場合、細胞ストレスの結果です。長時間の虚血時間は、膜タンパク質のの Ca 2+枯渇し、タンパク質分解による損傷は、細胞膜のブレブによって可視細胞の損傷にリンクされています。我々の観察から、非常に剪断応力に対して敏感と、ほとんどの場合、これらの細胞は、単離手順の間に死亡します。要約すると、実質および非実質肝細胞の正常な単離および分離が重要なステップは、右の時間枠内で行われ、分注ステップを注意深く、一般に、細胞単離のための時間を行い、剥離のされている必要がhouldは21できるだけ短くします。説明されたプロトコルの欠点は、分離条件( 例えば 、灌流時間は)完全に標準化することはできませんが、組織の品質に個別に適合させなければならないということです。また、得られた細胞集団の収率および純度は、組織の品質および消化の結果に依存して変化することができます。
我々は最近、この方法1により単離された各NPCの細胞型の機能解析と組み合わせた栽培条件の影響を実証する研究を発表しました。可能性は、単離すると肝臓の別々の異なる細胞集団は、革新的なヒト肝細胞共培養および組織工学のin vitro肝モデルを作成することができます。よく2Dモノ培養におけるPHH栽培が脱分化し、典型的な細胞機能7の喪失につながることが知られています。この理由は、生体内組織アルキを模倣することが必要ですin vitroでの肝臓モデル内テクチャ。 Kostadinovaおよび共同研究者(2013 8,9)だけでなく、メスナーおよび共同研究者(2013 8,9)は 、正常肝毒性効果を検出するための機能的な共培養肝モデルを確立しました。しかし、NPCは特徴付けられていないし、特定の機能は、これらのシステムでは調査されませんでした。
したがって、更なる研究は、共培養中のNPCの長期生存に関する調査、彼らの固有の特性との相互作用に焦点を当てるべきです。そのような研究のために、それはまた、胆管の単離のためのプロトコルを確立することは興味深いかもしれません。ネイティブの肝臓に含まれるすべての細胞型を含む機能のin vitro共培養の実現には、ヒト肝モデルのようなin vivoでの方向へのさらなるステップである可能性があります。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
0315741:私たちは、この研究は、教育研究のドイツ連邦省(BMBF)プロジェクト仮想肝臓でサポートされていた、図1の作成を支援するための嘉李劉に感謝したいと思います。
Materials
| Name | Company | Catalog Number | Comments |
| General Equipment | |||
| PIPETBOY | Eppendorf | ||
| pipettes | Eppendorf | ||
| microscope | Carl Zeiss | ||
| microscope | Olympus | ||
| CO2-incubator | Binder | ||
| Lamin Air | Heraeus | ||
| Centrifuge Varifuge 3.0R | Heraeus | ||
| Urine Beaker | Sarstedt | 2041101 | |
| perfusor syringe 50 ml | B.Braun | 12F0482022 | |
| Bottle Top Filter | Nalgene | 1058787 | |
| Falcon 50 ml Polypropylene Conical Tube | BD Biosciences | 352070 | |
| Falcon 15 ml Polypropylene Conical Tube | BD Biosciences | 352096 | |
| Tissue Culture plate | BD Biosciences | 533047 | 24 well |
| serological pipettes | BD Biosciences | 357525, 357551, 357543 | 25 ml, 10 ml, 5 ml |
| pipette tips | SARSTEDT | 0220/2278014, 0005/2242011, 0817/2222011 | 100 µl, 200 µl, 1,000 µl |
| Name | Company | Catalog Number | Comments |
| Isolation Equipment | |||
| water bath | Lauda | ||
| peristaltic pump | Carl Roth | ||
| circulation thermostat | Lauda | ||
| pH meter | Schott | ||
| fine scales | Sartorius | ||
| stand | |||
| Büchner funnel | Haldenwanger | ||
| plastic funnel | |||
| silicone tube | |||
| cannulae with olive tips | |||
| glass dish | |||
| forceps | |||
| scalpel | Feather | 12068760 | |
| Neubauer counting chamber | Optic Labor | ||
| cell lifter | Costar | ||
| Surgical Drape | Charité Universitätsmedizin Berlin | A2013027 | |
| compress | Fuhrmann | 40013331 | |
| sterile surgical gloves | Gammex PF | 1203441104 | |
| Tissue glue | B. Braun | 1050052 | |
| glass bottle | VWR | ||
| Collagenase P | Roche | 13349524 | |
| Percoll Separating Solution | Biochrom | L6145 | Density 1.124 g/ml |
| Hank’s BSS | PAA | H00911-3938 | |
| Dulbecco’s PBS | PAA | H15 - 002 | without Mg/Ca |
| Ampuwa | Plastipur | 13CKP151 | |
| Albumin | Sigma-Aldrich | A7906 | |
| NaCl | Merck | 1,064,041,000 | |
| KCl | Merck | 49,361,000 | |
| Hepes Pufferan | Roth | 133196836 | |
| EDTA | Sigma | E-5134 | |
| Name | Company | Catalog Number | Comments |
| Media Equipment | |||
| DMEM | PAA | E15-005 | Low Glucose (1 g/L) (without L-Glutamine) |
| HEPES Buffer Solution 1 M | GIBCO | 1135546 | |
| L-Glutamine | GIBCO | 25030-024 | 200 mM |
| MEM NEAA | GIBCO | 11140-035 | |
| penicillin/streptomycin | GIBCO | 15140-122 | |
| RPMI 1640 | PAA | E15 - 039 | without L-Glutamine |
| Sodium Pyruvate | GIBCO | 1137663 | 100 mM |
| Trypan Blue Solution | Sigma-Aldrich | T8154 | 0.4% |
| William’s E | GIBCO | 32551-020 | |
| with GlutaMAX™ | |||
| EGTA | Sigma-Aldrich | 03780-50G | |
| Fortecortin | Merck | 49367 | 8 mg/2 ml |
| Human-Insulin | Lilly | HI0210 | 100 I.E./ml |
| N-Acetyl cysteine | Sigma-Aldrich | A9165-5G | |
| Fetal calf serum (FCS) | PAA | A15-101 | |
| Name | Company | Catalog Number | Comments |
| Equipment for Immunostainings | |||
| CD 68 | R&D Systems, USA | monoclonal | |
| CK 19 | Santa Cruz | D2309 | polyclonal |
| CK18 | Santa Cruz | K2105 | monoclonal |
| Vimentin | Santa Cruz | monoclonal | |
| GFAP | Sigma Aldrich | monoclonal | |
| Triton X-100 | Sigma Aldrich | 23.472-9 | |
| Goat anti-Mouse IgG1-PE | Santa Cruz | C0712 | |
| Goat anti-rabbit IgG-FITC | Santa Cruz | L0412 | |
| Methanol | J.T.Baker | 1104509006 | |
| Formaldehyde 4% | Herbeta Arzneimittel | 200-001-8 | |
| Bovine serum albumin (BSA) | Sigma Aldrich | A7906-100G |
References
- Pfeiffer, E., et al. Isolation, characterization, and cultivation of human hepatocytes and non-parenchymal liver cells. Exp Biol Med. , Maywood. (2014).
- Si-Tayeb, K., Lemaigre, F. P., Duncan, S. A.
- Alpini, G., Phillips, J. O., Vroman, B., LaRusso, N. F. Recent advances in the isolation of liver cells. Hepatology. 20 (2), 494-514 (1994).
- Godoy, P., et al. Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME. Arch Toxicol. 87 (8), 1315-1530 (2013).
- Gómez-Lechón, M. J., Castell, J. V., Donato, M. T. Hepatocytes--the choice to investigate drug metabolism and toxicity in man: in vitro variability as a reflection of in vivo. Chem Biol Interact. 168 (1), 30-50 (2007).
- Ginai, M., et al. The use of bioreactors as in vitro models in pharmaceutical research. Drug Discov Today. 18 (19-20), 922-935 (2013).
- Schyschka, L., et al. Hepatic 3D cultures but not 2D cultures preserve specific transporter activity for acetaminophen-induced hepatotoxicity. Arch Toxicol. 87 (8), 1581-1593 (2013).
- Kostadinova, R., et al. A long-term three dimensional liver co-culture system for improved prediction of clinically relevant drug-induced hepatotoxicity. Toxicol Appl Pharmacol. 268 (1), 1-16 (2013).
- Messner, S., Agarkova, I., Moritz, W., Kelm, J. M. Multi-cell type human liver microtissues for hepatotoxicity testing. Arch Toxicol. 87 (1), 209-213 (2013).
- Nussler, A. K., Nussler, N. C., Merk, V., Brulport, M., Schormann, W., Yao, P., Hengstler, J. G. The Holy Grail of Hepatocyte Culturing and Therapeutic Use. Strategies in Regenerative Medicine. Santin, M. , Springer. New York. 1-38 (2009).
- Friedman, S. L., Roll, F. J. Isolation and culture of hepatic lipocytes, Kupffer cells, and sinusoidal endothelial cells by density gradient centrifugation with Stractan. Anal Biochem. 161 (1), 207-218 (1987).
- Knook, D. L., Blansjaar, N., Sleyster, E. C. Isolation and characterization of Kupffer and endothelial cells from the rat liver. Exp Cell Res. 109 (2), 317-329 (1977).
- Alabraba, E. B., et al. A new approach to isolation and culture of human Kupffer cells. J Immunol Methods. 326 (1-2), 139-144 (2007).
- Friedman, S. L., et al. Isolated hepatic lipocytes and Kupffer cells from normal human liver: morphological and functional characteristics in primary culture. Hepatology. 15 (2), 234-243 (1992).
- Lalor, P. F., Lai, W. K., Curbishley, S. M., Shetty, S., Adams, D. H. Human hepatic sinusoidal endothelial cells can be distinguished by expression of phenotypic markers related to their specialised functions in vivo. World J Gastroenterol. 12 (34), 5429-5439 (2006).
- Lee, S. M., Schelcher, C., Demmel, M., Hauner, M., Thasler, W. E. Isolation of human hepatocytes by a two-step collagenase perfusion procedure. J Vis Exp. (79), (2013).
- Rajan, N., Habermehl, J., Coté, M. F., Doillon, C. J., Mantovani, D. Preparation of ready-to-use, storable and reconstituted type I collagen from rat tail tendon for tissue engineering applications. Nat Protoc. 1 (6), 2753-2758 (2006).
- Chang, W., et al. Isolation and culture of hepatic stellate cells from mouse liver. Acta Biochim Biophys Sin (Shanghai). 46 (4), 291-298 (2014).
- Zeng, W. Q., et al. A new method to isolate and culture rat kupffer cells. PLoS One. 8 (8), e70832 (2013).
- Tokairin, T., et al. A highly specific isolation of rat sinusoidal endothelial cells by the immunomagnetic bead method using SE-1 monoclonal antibody. J Hepatol. 36 (6), 725-733 (2002).
- Damm, G., et al. Human parenchymal and non-parenchymal liver cell isolation, culture and characterization. Hepatology International. 7, 915-958 (2013).
- Baccarani, U., et al. Isolation of human hepatocytes from livers rejected for liver transplantation on a national basis: results of a 2-year experience. Liver Transpl. 9 (5), 506-512 (2003).
- Shen, L., Hillebrand, A., Wang, D. Q., Liu, M. Isolation and primary culture of rat hepatic cells. J Vis Exp. (64), (2012).
- Gerlach, J. C., et al. Large-scale isolation of sinusoidal endothelial cells from pig and human liver. J Surg Res. 100 (1), 39-45 (2001).
