

Summary
Denne protokol giver forskerne en hurtig, indirekte metode til måling af TLR-afhængige NF-кb/AP-1 transkriptionsfaktor aktivitet i en murine makrofag cellelinje som reaktion på en række polymere overflader og adsorbede protein lag, der modellere biomaterialet implantat mikromiljø.
Abstract
Den vedholdende inflammatoriske vært respons på en implanteret biomaterial, kendt som den udenlandske krop reaktion, er en betydelig udfordring i udviklingen og gennemførelsen af biomedicinsk udstyr og vævs ingeniør konstruktioner. Makrofager, en medfødte immuncelle, er centrale aktører i den udenlandske krop reaktion, fordi de forbliver på implantatet stedet for levetiden af enheden, og er almindeligt undersøgt for at få en forståelse af denne skadelige vært svar. Mange biomaterialer forskere har vist, at adsorbede protein lag på implanterede materialer påvirke makrofag adfærd, og efterfølgende påvirke værten respons. Metoderne i dette papir beskriver en in vitro-model ved hjælp af adsorberede proteinlag indeholdende cellulære skade molekyler på polymer biomaterialet overflader til at vurdere makrofag respons. En NF-кb/AP-1 reporter makrofag cellelinje og den tilhørende kolorimetriske alkalisk fosfataseanalyse blev anvendt som en hurtig metode til indirekte at undersøge NF-кb/AP-1 transkriptionsfaktor aktivitet som reaktion på komplekse adsorptions proteinlag, der indeholder blodproteiner og skade relaterede molekylære mønstre, som en model af de komplekse adsorbede protein lag dannet på biomaterialet overflader in vivo.
Introduction
Den udenlandske krop reaktion (FBR) er en kronisk vært respons, der kan have en negativ indvirkning på ydeevnen af et implanteret materiale eller enhed (f. eks, Drug levering udstyr, biosensorer), gennem vedvarende frigivelse af inflammatoriske mediatorer og ved at hæmme integrationen mellem det implanterede materiale og det omgivende væv1. Dette medfødte immunrespons initieres af implantationsproceduren og er karakteriseret ved langvarig tilstedeværelse af medfødte immunceller og fibrøs kapsel dannelse omkring implantatet1. I forbindelse med materiale værts respons har makrofag-materiale interaktioner en betydelig indvirkning på udviklingen af værts respons og udvikling af en FBR1. Makrofager er en forskelligartet medfødte immuncelle population, rekrutteret til implantationsstedet enten fra væv-residente makrofag populationer eller fra blodet som monocyte-afledte makrofager. De begynder at akkumulere på implantationsstedet kort efter implantation, og inden for dage bliver den fremherskende cellepopulation i implantat mikromiljøet. Materiale-klædende makrofager, sammen med fremmedlegeme gigant celler (fbgc) dannet gennem makrofag fusion, kan fortsætte på den materielle overflade for levetiden af implantatet2,3. Derfor, makrofager anses for at være centrale aktører i den udenlandske krop respons på grund af deres roller orkestrering de karakteristiske trin af FBR: akut inflammatorisk respons, væv Remodeling, og dannelse af fibrotisk væv1.
Bompenge lignende receptorer (TLRs) er en familie af mønster genkendelses receptorer, der udtrykkes af mange immunceller, herunder makrofager, og har vist sig at spille en væsentlig rolle i inflammation og sårheling. Ud over det patogen afledte ligands er TLRs i stand til at binde endogene molekyler, kendt som skade relaterede molekylære mønstre (DAMPs), som frigives under celle nekrose og aktivere inflammatoriske signalerings veje, der resulterer i produktion af proinflammatoriske cytokiner4. Vi og andre har foreslået, at skader opstået under bløddels biomateriale implantations procedurer frigiver damps, som derefter adsorcerer til biomaterialet overflader ud over blodproteiner og modurer efterfølgende celle-materiale interaktioner5,6. Når makrofager interagerer med det adsorbede protein lag på et implantat, kan deres overflade-TLRs genkende adsorptions DAMPs og aktivere proinflammatoriske signalering kaskader, hvilket fører til NF-κB og AP-1 transkriptionsfaktor aktivering og produktion af proinflammatoriske cytokiner. Vi har tidligere vist, at murine makrofager har øget NF-κB/AP-1 aktivitet og tumor nekrose faktor α (TNF-α, proinflammatorisk cytokin) sekretion som reaktion på fugt-holdige adsorbede protein lag på en række polymere overflader sammenlignet med overflader med adsorbede serum eller plasma kun (dvs., ingen DAMPs til stede), og at dette svar er i vid udstrækning medieret af TLR2, mens TLR4 spiller en mindre rolle5.
NF-κb/AP-1 reporter makrofag Cell line (tabel over materialer), der anvendes i denne protokol er en bekvem metode til at måle relative NF-κb og AP-1 aktivitet i makrofager5,7,8. I kombination med TLR pathway hæmmere, denne cellelinje er et nyttigt redskab til at undersøge TLR aktivering og dens rolle i inflammation som reaktion på en række stimuli5,7,8. Den reporter celler er en modificeret mus makrofag-lignende cellelinje, der kan stabilt producere udskillet embryonale alkalisk fosfatase (SEAP) på NF-κB og AP-1 transkriptionen faktor aktivering9. Den kolorimetriske enzymatiske alkalisk fosfataseanalyse (tabel over materialer) kan derefter anvendes til at kvantificere relative mængder af SEAP-udtryk som et indirekte mål for NF-κb/AP-1-aktiviteten. Da NF-κB og AP-1 er nedstrøms for mange celle signalerings veje, kan neutraliserende antistoffer og hæmmere, der er rettet mod specifikke Tlr'er (f. eks. TLR2) eller TLR-adapter molekyler (f. eks. MyD88), bruges til at verificere rollen for en bestemt vej. Den metode, der er beskrevet i denne artikel, giver en enkel og hurtig tilgang til vurdering af bidraget fra TLR signalering i murine makrofag svar til en række polymere overflader med adsorberede proteinlag, der indeholder både blodproteiner og damps som en in vitro model af implanterede biomaterialer.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. klargøring af medier og reagens
- Forbered fibroblast medier. Kombiner 450 mL Dulbecco's modificerede Eagle medium (DMEM), 50 mL føtal bovint serum (FBS) og 5 mL penicillin/streptomycin. Opbevares ved 4 °C i op til 3 måneder.
- Forbered reporter makrofag Growth Media i 50 ml aliquoter. 45 mL DMEM, 5 mL FBS, 5 μg/mL Mycoplasma eliminations reagens (tabel over materialer) og 200 μg/ml phleomycin D1 (tabel over materialer). Opbevares ved 4 °C i op til 3 måneder.
- Udarbejd reporter makrofag assay-medier i 50 ml aliquoter. Kombiner 45 mL DMEM, 5 mL varme inaktiverede FB'ER (HI-FBS), 5 μg/mL Mycoplasma eliminations reagens og 200 μg/mL phleomycin D1. Opbevares ved 4 °C i op til 3 måneder.
2. belægning cellekultur overflader med poly (methylmethacrylat)
- Poly (methylmethacrylat) (PMMA) opløses i chloroform ved 20 mg/mL (f. eks. 100 mg PMMA i 5 mL chloroform) i et 20 mL glas scintillationshætte glas. Anbring en magnetisk omrørs stang i hætteglasset, og lad den røre i mindst 2 timer, indtil alle faste stoffer er opløst.
Forsigtig: Chloroform er skadelig ved indånding. Sørg for at bruge opløsningsmiddel i en stinkhætte, mens du bærer PVA handsker. - Pipetten 400 μL PMMA-opløsning på midten af et borosilikat glas mikroskop-slide i et spin-Coater, og drej ved 3000 rpm i 2 min. Forbered antallet af dias, der kræves til analysen, samt 3 − 5 ekstra til måling af vandkontaktvinkel. Opbevar slides i en ren boks (sprøjtet og udslettet med 70% ethanol) til fremtidig brug.
Bemærk: Spin belægning bruges ofte til at deponere en tynd, ensartet belægning på en flad overflade. Et spin Coater roterer et substrat ved høje hastigheder ved hjælp af centrifugalkraft for at sprede belægnings opløsningen over overfladen.- Mål vand kontaktvinkel på to tilfældige positioner på overfladen af ekstra coatede slides (dvs., ikke de slides, der anvendes til cellekultur) med en goniometer for at sikre glasoverfladen var helt belagt med polymer.
Bemærk: Kun vand med den højeste renhed (f. eks. glas tredobbelt destilleret) skal anvendes til måling af vand kontakt vinkler.
- Mål vand kontaktvinkel på to tilfældige positioner på overfladen af ekstra coatede slides (dvs., ikke de slides, der anvendes til cellekultur) med en goniometer for at sikre glasoverfladen var helt belagt med polymer.
- I et biologisk sikkerhedsskab (BSC) Fastgør 8-kammer klæbrig brønde til PMMA belagt-slides ved hjælp af sterile pincet og efter aseptisk teknik. Tryk fast på toppen af de klæbrig brønde for at sikre, at de er stærkt fastgjort. Inkuber gliderne med vedlagte klæbrig brønde ved 37 °C natten over for at sikre forseglingen.
- Test forseglingen af de klæbrig brønde ved at tilføje 200 μL cellekultur kvalitet (endotoxin-fri) vand til hver brønd. Inkuber ved stuetemperatur (RT) i 60 min, og sørg for, at der ikke er lækage, før du fortsætter. Aspirer vandet, er omhyggelig med ikke at forstyrre PMMA belægning.
- Udfør endotoksin-fri vand skyller ved at tilsætte 300 μL endotoxin-frit vand til hver brønd og inkuberet i 1 time (tre gange), 12 timer og 24 timer før brug for at fjerne eventuelle resterende opløsningsmidler.
- Test endotoksin koncentrationen af de slides, der skal anvendes til cellekultur. Inkuber 200 μl endotoksin-frit reagens vand (tabel over materialer) i en brønd på hver slide i 1 time. mål endotoksin koncentrationen i ekstrakten ved hjælp af en endepunkts kromogen endotoksin-analyse (tabel over materialer).
Bemærk: Følgende protokol er specifik for endotoksin-analysesættet, der er anført i tabellen over materialer. - Brug kun vand og hjælpematerialer (dvs. pipettespidser, mikrocentrifuge glas og brønd plader), der er certificeret pyrofri (dvs. endotoksinfri) til dette arbejde. Desuden bør alle glasvarer, der anvendes til fremstilling af de polymer belagte overflader, aftørres ved hjælp af tør varmesterilisering (250 °C i 30 min) før brug10. Måling af endotoksin i ekstraktet opløsning, som beskrevet her, kan resultere i en undervurdering af endotoksin på materialets overflade11,12. Det anbefales derfor, at der ved udvikling af en polymer coating-protokol udføres en endotoksin-analyse reaktion (dvs. trin 2.5.4 − 2.5.6 for testprøver [reagens vand] eller spids kontrol) direkte i brønde, der indeholder den coatede prøve for at sikre, at der ikke uforvarende tilføres nogen kilder til endotoksin i systemet under belægningsprocessen.
- Alle testprøver (dvs. ekstrakter) og reagenser til endotoksin-analyse anvendes til rt. Rekonstituer kromogen reagens i analyse buffer og endotoksin standard i reagens vand, gør det muligt at opløse i 5 minutter og forsigtigt hvirvle inden brug. Dæk alle flasker med paraffin folie, når de ikke er i brug.
- Opret en 5 − 8-punkts standard fortyndings kurve for endotoksin standard, der spænder fra den nedre til den øvre grænse for analysen ved at udføre en seriel fortynding af endotoksin standarden i reagens vand.
- For at kontrollere for forbedring eller hæmning af endotoksin analysen i testprøverne, skal der forberedes en positiv kontrol (også kaldet en Spike Control eller spidse prøve) ved at fortynde en kendt mængde endotoksin i ubrugt test prøveopløsning.
Bemærk: Koncentrationen af den positive kontrol bør være den samme koncentration som en standard i midten af standardkurven. Hvis den inddrevne mængde af endotoksin Spike (dvs. koncentrationen af den positive kontrol minus koncentrationen af den uspikede prøve) er inden for 50 − 200% af den nominelle koncentration af endotoksin spike, kan ekstraktionsopløsningen anses for ikke signifikant at forstyrre analysen. - Tilsæt 50 μL standarder, prøver eller Spike kontroller til hver brønd af en 96-brønd plade i duplikat eller tre eksemplarer. Brug reagens vand som en negativ kontrol.
- Tilsæt 50 μL kromogen reagens til hver brønd. Tilsæt hurtigt reagenset til alle brønde. Brug en timer til at registrere den tid, det tager at tilføje reagens til alle brønde. Dæk pladen med en klæbe forsegling og Inkuber ved 37 °C (Inkubationstiden er meget afhængig og angivet på analysecertifikatet, der indgår i det kromogene reagens Kit). Alternativt kan du tjekke pladen hver 15 min. under inkubationen, indtil der observeres farveændringer i alle standard brønde.
- Efter inkubation tilsættes 25 μL 50% eddikesyre til hver brønd (endelig koncentration på 10% eddikesyre pr. brønd) for at standse reaktionen. Tilsæt eddikesyre i samme rækkefølge, som det kromogene reagens blev tilsat. Læs pladens absorbans ved hjælp af en plade læser ved 405 nm. Aspirér væske og kassér pladen.
Bemærk: Tilsætning af eddikesyre bør tage samme tid at tilføje til hver brønd, som det kromogene reagens tog (± 30 s).
- Ultraviolet (UV) Steriliser diasene i 30 minutter før cellekultur eksperimenter.
3. belægning af cellekultur overflader med Polydimethylsiloxan
- Bland Polydimethylsiloxan (PDMS) elastomer i et 10:1 vægt forhold (Base: Hærdningsmiddel). I et biologisk sikkerhedskabinet pipetteres ca. 10 mL Polydimethylsiloxan base i et sterilt rør. Afvejes røret, og tilsæt langsomt hærdnings midlet, indtil 10% er blevet tilsat.
Forsigtig: Brug PDMS-reagenser i et godt ventileret område, og undgå øjenkontakt ved at bære sikkerhedsbriller. - Bland elastomer grundigt ved omrøring med en steril serologisk pipettespids og ved pipettering op og ned. Tilsæt ca. 200 μL af opløsningen til hver brønd på en 48-brønd plade. VIP brønd pladen langsomt for at sikre fuldstændig dækning af brønde med elastomer opløsning.
- Placer brønd pladen med elastomer i en vakuum ovn, der er sat til 50 cmHg, 40 °C. Fjern låget og dækslet med en enkeltlags serviet for at forhindre, at andre rester falder i brøndene. Lad det inkubere i mindst 48 h.
- Bekræft brøndene er helt belagt via visuel inspektion. Sørg for, at elastomer er helbredt ved forsigtigt at prodding med en steril pipettespids, før du fjerner den.
- Tilsæt 300 μL 70% ethanol (lavet med absolut ethanol og endotoksin-fri vand), og Inkuber ved RT i 1 time. Fjern ethanol og Udfør endotoxin-fri vand skyller ved at tilsætte 300 μL endotoxin-frit vand til hver brønd og inkubere i 1 h (tre gange), 12 h , og 24 timer før brug for at fjerne eventuelle resterende opløsningsmidler.
- Inkuber 200 μl endotoksin-frit vand i tre brønde af hver plade i 1 time. mål endotoksin koncentrationen af vand ekstrakterne ved hjælp af en endepunkts kromogen endotoksin-analyse (trin 2.5.1 − 2.5.6).
4. belægning cellekultur overflader med fluorholdig poly (tetrafluorethylen)
- En 1 mg/mL opløsning af fluorholdig poly (tetrafluorethylen) (fPTFE) (f. eks. tilsættes 10 mg fPTFE til 10 mL fluorholdig opløsningsmiddel [tabel over materialer]) i et 20 ml glas scintillationshætte glas. Anbring en magnetisk omrørs stang i hætteglasset, og lad den røre i mindst 24 timer, indtil alle faste stoffer er opløst.
- Tilsæt ca. 150 μL polymer opløsningen til hver brønd af en polystyren 48-brønd plade (dvs. ikke vævskultur behandlet). VIP brønd pladen langsomt for at sikre fuldstændig dækning af alle brønde med polymeropløsning. Udskift låget.
- For at sikre effektiv fPTFE-belægning af brønde skal glas dæksedler være belagt med fPTFE og anvendes til måling af vandkontaktvinkel (trin 4.3.1). Placer dæksedler inde i brøndene på en 24-brønd plade. Tilsæt ca. 400 μL polymer opløsningen til hver brønd, der indeholder en dækseddel. Skub dæksedlerne ned med sterile pincet, så de er helt dækket af polymer opløsningen, og dæk brønd pladen med et låg.
- Placer brønd pladen med polymeropløsning og/eller dæksedler i en vakuum ovn, der er sat til 50 cmHg, 40 °C. Fjern låget og dækslet med en enkeltlags serviet for at forhindre, at andre rester falder i brøndene. Lad det inkubere i mindst 48 h.
- Mål vand kontakt vinklen på fPTFE-belagte dæksedler med goniometer for at sikre effektiv belægning.
Bemærk: Kun vand med den højeste renhed (f. eks. glas tredobbelt destilleret) skal anvendes til måling af vand kontakt vinkler.
- Mål vand kontakt vinklen på fPTFE-belagte dæksedler med goniometer for at sikre effektiv belægning.
- Tilsæt 300 μL 70% ethanol (lavet med absolut ethanol og endotoksin-fri vand), og Inkuber ved RT i 1 time. Fjern ethanol og Udfør endotoxin-fri vand skyller ved at tilsætte 300 μL endotoxin-frit vand til hver brønd og inkubere i 1 h (tre gange), 12 h , og 24 timer før brug for at fjerne eventuelle resterende opløsningsmidler.
- Inkuber 200 μl endotoksin-frit vand i tre brønde af hver plade i 1 time. mål endotoksin koncentrationen af vand ekstrakter ved hjælp af en endepunkts kromogen endotoksin-analyse (trin 2.5.1 − 2.5.6).
- UV sterilisere brønden plader i 30 min før cellekultur eksperimenter.
5. gør Lysat fra 3T3 celler
- Vokse 3T3 celler i flere T150 kolber til 70% sammenløbet. Til at frigøre celler, aspirere medier, vaske overflade med 5 mL PBS og aspirere PBS. Tilsæt 5 mL animalsk oprindelse-fri, rekombinant celle dissociations enzym (tabel over materialer) og Inkubér ved 37 °c i 3 − 5 minutter.
- Fjern cellerne ved forsigtigt at vippe kolben frem og tilbage. 5 mL PBS tilsættes for at neutralisere det rekombinante enzym, der anvendes til celle dissociation. De løsrevne celler overføres fra kolberne til et centrifugeglas og blandes via pipettering. Udføre en levende celletal ved hjælp af en hemocytometer og cellelevedygtighed farvestof.
Bemærk: Et celle dissociations enzym, der kan neutraliseres ved fortynding i PBS, blev udvalgt for at undgå, at der blev indført serum baserede proteiner i lyspræparatet. Hvis trypsin anvendes til at adskille cellerne, skal det neutraliseres med en serum-indeholdende opløsning, og der skal udføres en yderligere PBS-vask for at reducere mængden af serumproteiner, som overføres til lysnings præparatet. - Cellerne centrifugeres ved 200 x g i 5 min. Aspirér supernatanten, og gensuspender cellerne i original volumen (dvs. 10 ml x antal kolber) af PBS for at vaske eventuelle resterende medier. Gentag.
- Centrifuger cellerne igen ved 200 x g i 5 minutter og Aspirér supernatanten. Tilsæt mængden af PBS, der kræves for at opnå en endelig celle koncentration på 1 x 106 celler/ml. Celle opløsningen placeres i en-80 °C-fryser, indtil prøven er helt frosset (mindst 2 timer).
- Tø celle opløsning i et 37 °C vandbad. Når den er helt optøet, placeres opløsningen tilbage i-80 °C-fryseren, indtil den er helt frosset. Gentag for i alt 3 fryse-tø cyklusser.
- Foretag en mikrobicinchonininsyre (BCA) assay på celle lysatet ved en række fortyndinger (f. eks. 1/100, 1/200, 1/500, 1/1000) for at bestemme proteinkoncentrationen. Celle lysatet fortyndes til en proteinkoncentration på 468,75 μg/mL, aliquot og opbevares ved-80 °C til fremtidig brug.
Bemærk: Endelig proteinkoncentration i en 48-brønd plade er 125 μg/cm2 (baseret på overfladearealet af en brønd, 0,75 cm2). - Udfør en Western blot for at vurdere tilstedeværelsen af DAMPs i lysat (f. eks. varmechok protein 60 [HSP60], høj mobilitet gruppeboks 1 [HMGB1]) ved at indlæse 40 − 60 μg lysat protein i loading buffer på en 1,5 mm tyk 10% polyacrylamid gel og følg standard Western blot Procedurer.
6. vurdering af effekten af Adsorbede Proteinlag og bompenge lignende receptorer på NF-κB aktivitet af makrofager
Bemærk: For en skematisk af den eksperimentelle workflow og plade layout, se figur 1a og supplerende figur 1, hhv.
- Vokse reporter makrofager i en passende størrelse kolbe til 70% sammenløbet. Indsug medier, vask overflade med PBS, og Aspirer PBS. Tilsæt det rekombinante celle dissociations enzym, og Inkuber ved 37 °C i 8 min.
- Frigør cellerne ved at trykke hårdt på kolbens sider. Inaktivere det rekombinante celle dissociations enzym ved at tilføje et tilsvarende volumen af vækstmedier (indeholdende 10% FBS). Udføre en levende celletal ved hjælp af en hemocytometer og cellelevedygtighed farvestof.
Bemærk: Forventet levedygtighed for indberetteren makrofager efter en 8 min inkubation i celle dissociations enzymet er 90%. - Centrifuge celler ved 200 x g i 5 min. Aspirér supernatanten og resuspension i den oprindelige mængde PBS til at vaske celler. Centrifugeres igen og resuspenderes cellerne ved 7,3 x 105 celler/ml i analyse medier (indeholdende varme INAKTIVEREDE fb'er).
- Separat cellesuspension i 3 forskellige rør: TLR4 inhibitor, anti-TLR2, og ubehandlet. Inkuber celler med 1 μg/mL TLR4 hæmmer i 60 min ved RT eller med 50 μg/mL anti-TLR2 i 30 min ved RT.
- Tilsæt 200 μL lysat, 10% FBS, 10% kommerciel muse plasma (tabel over materialer) eller en blanding af proteinopløsninger til en 48-brønd plade (eller tilsvarende), og lad proteinet adsorbere ved 37 °c i det ønskede tidsrum (dvs. 30 min, 60 min eller 24 timer). Aspirér proteinopløsninger fra brønde ved hjælp af en frisk Pasteur-pipette til hver proteinopløsning, og vask overfladerne med 250 μL PBS i 5 min. aspirat PBS. Gentag for i alt 3 skyller.
Bemærk: Dette trin skal muligvis startes tidligere i protokollen afhængigt af den ønskede adsorptions tid. Juster protokollen i overensstemmelse hermed. - Efter inkubationstiden med TLR4-hæmmer eller anti-TLR2, pipette cellerne til at resuspendere. Tilsæt 200 μL celle opløsning til hver brønd.
- For TLR2 positiv kontroltilstand tilsættes Pam3CSK4 til en endelig koncentration på 150 ng/mL. For TLR4 positiv kontroltilstand tilsættes lipopolysaccharid (LPS) til en endelig koncentration på 1,5 μg/mL. Inkubatceller ved 37 °C i 20 timer.
- Prøve 20 μL supernatanten fra hver brønd og plade i duplikat til en 96-brønd plade. Medtag tre brønde af 20 μL analyse medier som baggrundskontrol. Tilsæt 200 μL SEAP reporter assay reagens til hver brønd. Dæk pladen med en klæbe forsegling og Inkuber i 2,5 h ved 37 °C.
Bemærk: Inkubationstiden kan variere afhængigt af forsøgs forholdene og bør optimeres for en stærk forskel i absorbans mellem positive og negative kontrol brønde.- Resten af supernatanten overføres til et 1,5 mL rør (pr. brønd). Centrifugeres ved 1.000 x g i 10 minutter for at pille snavs. Supernatanten overføres til et nyt 1,5 mL rør og opbevares ved-80 °C. Analysér supernatanten for tilstedeværelsen af proinflammatoriske cytokiner (f. eks. TNF-α, interleukin 6) via enzym relateret immunosorbent assay (ELISA).
- Fjern selvklæbende pladeforsegling. Læs pladens absorbans ved hjælp af en plade læser ved 635 nm. Aspirér væske og kassér pladen.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Rengørings metoderne til de polymer belagte overflader blev testet for at sikre, at belægningen ikke forstyrrede, hvilket ville blive opfattet som en ændring i vandkontakt vinklen til en ubestrøget glas dækseddel (figur 2). Iblødsætning PMMA-belagt mikroskop slides i 70% ethanol for 1 h blev fundet for at fjerne PMMA belægning (figur 2, venstre panel), sandsynligvis på grund af opløseligheden af pmma i 80 WT% ethanol13, derfor PMMA-belagte overflader blev rengjort ved hjælp af 30 min UV-sterilisering alene. Koncentrationen af PMMA til belægning blev optimeret tidligere5. En 1 h 70% ethanol Soak blev brugt til at rense PDMS, og UV sterilisering blev forsømt, da UV-lys kan forårsage kæde saks og påvirke overfladen befugtning egenskaber af PDMS14. Både 70% ethanol Soak og UV-sterilisering påvirkede ikke vandkontakt vinklen på fPTFE-belagte dæksedler (figur 2, højre panel), og derfor blev de to metoder i træk anvendt til at rense fptfe-belægninger. Metoden til fPTFE-belægning blev tidligere beskrevet af Grainger Group15.
En Western blot blev udført på 3T3 lysatet for at sikre, at FUGTIGE arter var til stede i den komplekse molekylære blanding. Resultaterne viste, at både HMGB1 og HSP60, to veldokumenterede damps16,17, var til stede i lysatet (figur 1B). Adsorptions af TLR-ligander fra lysatet til polymer overfladerne blev bekræftet af dyrkning reporter Macro Fages (ubehandlet, TLR2 neutraliseret eller TLR4 hæmmet) for 20 h på protein-adsorberede polymer overflader (dvs. vævskultur behandlet polystyren [TCPS], PMMA, PDMS, fptfe), og derefter indirekte vurdere NF-кb/AP-1 aktivitet baseret på SEAP produktion ved hjælp af en enzymatisk analyse (figur 1C og figur 3). Desuden havde indberetteren makrofager signifikant øget NF-кB/AP-1 aktivitet på adsorbet lysat sammenlignet med adsorbbede FBS eller plasma og ingen præ-adsorberede protein (medier) (figur 4). TLR-ligander syntetisk triacyleret lipopeptid (Pam3CSK4, TLR2 ligand) og lipopolysaccharid (LPS, TLR4 ligand) blev inkluderet som positive kontroller for at bekræfte antistoffet eller inhibitorer, og analysen fungerede korrekt. TLR2 neutralisering havde en mærkbart stærkere reduktion i NF-кB/AP-1 respons af reporter makrofager til adsorberende lysat sammenlignet med TLR4 hæmning. Så godt, små mængder af lysat fortyndet i serum (baseret på total protein) induceret signifikant øget NF-кB/AP-1 respons sammenlignet med serum alene, med den laveste effektive fortynding afhængig af polymer overfladen (figur 5). Disse resultater viser styrken af de adsorbede lysat-afledte molekyler på inducerende TLR-afhængige NF-кB/AP-1 aktivitet i reporter makrofager på en række polymere overflader.

Figur 1: metoder og resultater for alkalisk fosfataseassay af NF-кB/AP-1 reporter makrofager på TCP'ER, PMMA, PDMS og fPTFE. A) diagram over arbejdsgangen for indberetteren makrofag alkalisk fosfataseanalyse. B) Western blot af lysat, som bekræfter tilstedeværelsen af FUGTIGE arter HMGB1 og HSP60, med β-actin som læsse kontrol. C) NF-КB/AP-1-aktivitet (repræsenteret ved absorbans) af reporter-makrofager, som dyrkes på medier (negativ kontrol) 10% FBS, Lysate og PAM3CSK4 (TLR2 ligand, positiv kontrol) for 20 h. data viser resultaterne af et eksperiment og er repræsentative for resultater fra mindst 2 separate eksperimenter, vist som middel ± standardafvigelse (SD). Hvert eksperiment anvendte n = 3 separate brønde pr. tilstand, og hver brønd blev belagt i to eksemplarer til den enzymatiske analyse. Analyseret ved hjælp af en-vejs ANOVA og Tukey post-hoc test. p < 0,001. Dette tal er blevet tilpasset med tilladelse fra McKiel og Fitzpatrick5. Copyright 2018 det amerikanske kemikalie selskab. Venligst klik her for at se en større version af dette tal.

Figur 2: optimering af rengøringsmetoder for PMMA-og fPTFE-belagte overflader, vurderet ved hjælp af vand kontaktvinkel (WCA). Målingerne blev foretaget på 2 separate pletter på mindst 3 dæksedler. Data vises som middelværdien ± SD. analyseret ved hjælp af en-vejs ANOVA og Tukey post-hoc test. * p < 0,05. Dette tal er blevet tilpasset med tilladelse fra McKiel og Fitzpatrick5. Copyright 2018 det amerikanske kemikalie selskab. Venligst klik her for at se en større version af dette tal.

Figur 3: TLR-medieret NF-кB/AP-1 aktivitet (repræsenteret ved absorbans) af reporter makrofager dyrket på 10% FBS (Control), lysate, og positiv kontrol for 20 h. A) påvirkning af TLR2 neutralisering på journalistens makrofag respons på adsorbet lysat. Positiv kontrol er Pam (Pam3CSK4, TLR2 ligand). B) påvirkning fra TLR4 hæmning af journalistens makrofag respons på adsorberende lysat. Positiv kontrol er LPS (TLR4 ligand). Data viser resultaterne af et eksperiment og er repræsentative for resultater fra mindst 2 separate eksperimenter, vist som middelværdien ± SD. Hvert eksperiment anvendte n = 3 separate brønde pr. tilstand, og hver brønd blev belagt i to eksemplarer til den enzymatiske analyse. Analyseret ved hjælp af en-vejs ANOVA og Tukey post-hoc test. * * p < 0,01, * * * p < 0,001. Dette tal er blevet tilpasset med tilladelse fra McKiel og Fitzpatrick5. Copyright 2018 det amerikanske kemikalie selskab. Venligst klik her for at se en større version af dette tal.

Figur 4: NF-кB/AP-1 aktivitet (repræsenteret ved absorbans) af reporter makrofager kulturperler på medier (negativ kontrol), 30 min og 24 h adsorbede protein lag, og Pam3CSK4 (positiv kontrol) på TCPS for 20 h. Data kombineres fra 3 separate eksperimenter og vises som middel ± SD. Hvert eksperiment anvendte n = 3 separate brønde pr. tilstand, og hver brønd blev belagt i to eksemplarer for den enzymatiske analyse (dvs. n = 9 ikke-uafhængige cellekultur brønde og n = 18 ikke-uafhængige enzymatiske analyse brønde). Analyseret ved hjælp af en-vejs ANOVA og Tukey post-hoc test. p < 0,001. Venligst klik her for at se en større version af dette tal.
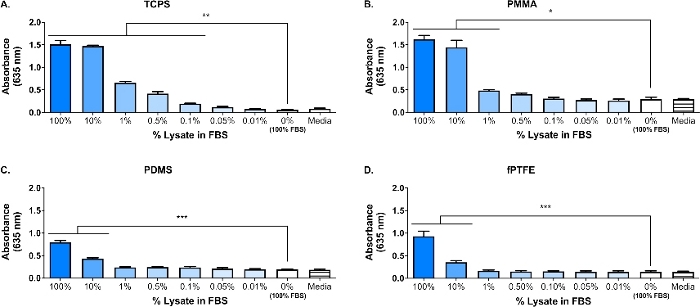
Figur 5: reporter makrofag NF-кb/AP-1 aktivitet (repræsenteret ved absorbans) efter 20 h som reaktion på fortyndinger af lysat i FBS (totalt protein = 280 μg/brønd) adsorberes til polymer overflader i 30 min. (A) tcp'er. B) PMMA. (C) PDMS. D) fptfe. Data viser resultaterne af et eksperiment og er repræsentative for resultater fra mindst 2 separate eksperimenter, vist som middelværdien ± SD. Hvert eksperiment anvendte n = 3 separate brønde pr. tilstand, og hver brønd blev belagt i to eksemplarer til den enzymatiske analyse. Analyseret ved hjælp af en-vejs ANOVA og Tukey post-hoc test. * p < 0,05, * * p < 0,01, * * * p < 0,001. Dette tal er blevet tilpasset med tilladelse fra McKiel og Fitzpatrick5. Copyright 2018 det amerikanske kemikalie selskab. Venligst klik her for at se en større version af dette tal.

Supplerende figur 1: eksempel layouts, der anvendes til NF-кb/AP-1 reporter makrofag cellekultur assay i 8-kammer og 48-brønd plade formater. Venligst klik her for at se en større version af dette tal.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Et primært fokus i vores laboratorium er værts reaktionen på solide biomateriale bløddels implantater, og især hvordan de cellulære skader, der påløber under implantationsproceduren, påvirker værts reaktionen. Det arbejde, der præsenteres her beskriver indledende eksperimenter ved hjælp af en reporter makrofag cellelinje og in vitro-genereret fugtig-holdige cellulære lysat, at undersøge indflydelsen af molekyler frigivet under cellulære skader (dvs., fra implantatet kirurgi) på makrofag reaktioner på biomaterialer. Fibroblast Cell lysat blev brugt til at modellere cellulære skader og frigivelse af damps på grund af biomaterialet placering. Fibroblaster blev valgt til at skabe lysatet på grund af forekomsten af fibroblaster i blødt væv, samt deres evne til at udskiller en række ekstracellulære matrix (ECM) proteiner, herunder fibronektin18. Freeze-tø cykling blev valgt som metoden til lysis til fremstilling af både intracellulære og ECM-afledte DAMPs, svarende til, hvad der ville være til stede i implantat miljøet. Proteasehæmmere blev ikke brugt til at gøre dette lysat. Mens ukontrolleret cellelyse som fryse-tø-cykling kan resultere i frigivelse af proteaser, der kan forringe DAMPs, vil disse enzymer sandsynligvis også være til stede i biomaterialet-implantat miljøet, når cellerne beskadiges under implantationsproceduren. Tilstedeværelsen af DAMPs i den komplekse molekylære blanding af lysatet blev bekræftet af Western blot (figur 1B; HMGB1 og HSP60) og SEAP reporter-analyse (figur 1C; NF-кB/AP-1 aktivitet som respons på adsorbet lysat). Vi har også udført assays, hvor lysat blev fortyndet i FBS baseret på total proteinkoncentration og adsorbet på cellekultur overflader (figur 5) for bedre at afspejle kompleksiteten af implantat miljøet, da det vil indeholde en overflod af blodproteiner samt damps6. Reporter makrofag NF-кb/AP-1 aktivitet forblev signifikant forøget på adsorberede lag fra lysat fortyndet i FBS, og den laveste fortynding for at opnå signifikant aktivering var overflade afhængig, spænder fra 0,1% (TCPS) til 10% (PDMS og fptfe).
Polymererne PMMA, PDMS og PTFE blev valgt til dette arbejde, fordi de er ikke-nedbrydelige og har været anvendt i udstrakt grad i litteraturen til at vurdere protein adsorptions og makrofag respons på biomaterialer19,20,21,22,23,24,25. Tcp'er blev også anvendt til sammenligning, da det er et fælles substrat, der anvendes til in vitro-makrofag og TLR-signalering-arbejde21,26,27,28. De materialer, der anvendes i vores arbejde, er repræsentative eksempler på ikke-nedbrydelig, solid biomaterialer. Men mange andre materialer kunne bruges med denne model, forudsat at materialet kan være belagt på cellekultur plader eller mikroskop slides og korrekt dekontamineres. NF-κb/AP-1 reporter makrofag Cell line blev valgt til denne in vitro model, fordi det muliggør hurtig, indirekte måling af NF-κb/AP-1 aktivitet gennem NF-κb/AP-1 inducerbar udtryk af SEAP. NF-κB/AP-1 reporter makrofager kræver brug af phleomycin D1 i dyrkningsmediet som et selektivt antibiotikum for at sikre, at kun celler med NF-кB/AP-1 inducerbare SEAP-genet er til stede29. For alkalisk fosfataseassay er det afgørende at bruge HI-FBS i cellekultur mediet for at undgå potentielle falske positive resultater genereret af alkalisk fosfataser, der findes i serum. Vores forskning til dato tyder på, at FBS-adsorbede overflader ikke genererer et detekterbart falsk positivt resultat, sandsynligvis fordi serum molekyler er stærkt adsorbteres til kultur overfladen og ikke frigives i supernatanten. Kultur tidspunktet for indberetteren af makrofager (20 timer), analyse af assay-tidspunkt (2,5 h) og absorbans-læse bølgelængde (635 nm) for alkalisk fosfataseanalyse blev optimeret med dette system for at sikre robuste og reproducerbare målinger under alle forhold.
En Initial protein adsorptions tidspunkt på 30 min blev valgt til dette arbejde på grund af dets almindelige anvendelse i protein adsorptions litteratur (figur 1C)30,31,32,33,34. Men vi har også udforsket længere adsorptions tider (dvs. 60 min og 24 timer, figur 4) for bedre at repræsentere det adsorbede protein lag, som makrofager ville interagere med in vivo, hvilket sandsynligvis vil forekomme 4 − 24 timer efter implantation1. Det er blevet postuleret, at størstedelen af protein adsorptions og udveksling forekommer i den første 60 min eksponering for en overflade26,35,36, derfor en 60 min adsorptions tid kan være en mere relevant tidspunkt. Vi har også bevæget sig fra at bruge FBS som en negativ kontrol for tilstedeværelsen af DAMPs i det adsorbede protein lag til kommerciel mus plasma. Begrundelsen for at bruge plasma i stedet for serum er, at plasmaproteiner er kendt for at spille en betydelig rolle i protein adsorptions og makrofag respons1, og at plasma giver en bedre repræsentation af proteinerne i sårmiljøet. Plasma, der anvendes i protein adsorptions forsøg, er almindeligvis tilberedt som en 1 − 10% fortynding26,36,37, hvilket motiverede vores brug af 10% plasma. Humant plasma er almindeligt anvendt26,36, da det er lettere at opnå i store mængder og mere klinisk relevant, sammenlignet med mus plasma. Men, vi valgte at bruge kommerciel mus plasma for i denne model til at holde arterne af proteinopløsninger i overensstemmelse med den af de reporter celler.
Brugen af journalisten makrofag Cell line indførte nogle begrænsninger i studiet. Først, ved hjælp af en murine leukæmi makrofag cellelinje har iboende begrænsninger, som fænotype og adfærd kan variere fra primære makrofag kulturer. Selv om denne begrænsning vil blive behandlet i det fremtidige arbejde ved hjælp af primære makrofager, blev den forældre lige makrofag cellelinje vist at nøje efterligne muse knoglemarvs afledte makrofager med hensyn til deres celleoverflade receptorer og respons på mikrobielle ligander for TLRs 2, 3 og 438. Desuden gav NF-κB/AP-1 reporter makrofager lignende resultater som respons på HMGB1 og LPS stimulation sammenlignet med peritonealdialyse primær murine makrofager39. Det skal bemærkes, at NF-κB/AP-1 reporter makrofager, og deres forældre stamme, ikke udtrykker TLR540. Forskere har vist, at HMGB1 var i stand til at aktivere NF-κb transkriptionsfaktorer via TLR5 signalering veje i hek-293 celler stabilt transficeret med human TLR541. Derfor blev bidraget af HMGB1-TLR5 signalering til den samlede NF-κB aktivitet på lysatbelagte overflader forsømt i denne model. Derudover udtrykker journalistens makrofager og deres forældre stamme ikke ASC-adapter proteinet, og danner derfor ikke de fleste typer af inflammasomer og kan ikke behandle inaktive IL-1β eller inaktive IL-18 til deres modne formularer42. Derfor, den model, vi har brugt ikke tegner sig for bidraget af ASC-afhængige inflammasome aktivitet og efterfølgende autokrin Il-1β og Il-18 signalering i makrofag respons på lysat-adsorbede overflader. Derfor er denne analyse tænkt som en foreløbig undersøgelse af TLR-afhængig NF-κB aktivering, og efterfølgende forskning ved hjælp af primære makrofager anbefales at give en mere komplet og repræsentativ forståelse af makrofag aktivering og fænotype på materielle overflader af interesse.
Den alkaliske fosfataseanalyse måler indirekte NF-кB/AP-1-aktiviteten af indberetteren makrofager. Men, der er mange signalerings veje andre end TLRs, der involverer NF-кB/AP-1 (f. eks interleukin-1 receptor [IL-1R]43 og tumor nekrose faktor receptor [tnfr]44). Derfor var det nødvendigt at vurdere bidraget fra TLR2 og TLR4 signalering i den øgede NF-кB/AP-1 respons på lysate-adsorberende overflader ved hjælp af inhibering assays (figur 3). Begrundelsen for at vælge disse to overflade TLRs var, at mindst 23 DAMPs, der har vist sig at signalere gennem TLR2 og TLR445, herunder de velkarakteriserede HMGB1, og begge receptorer er udtrykt på cellens overflade og kan interagere direkte med biomaterialet overflade6. TLR2-og TLR4-hæmnings assays viste, at når TLR2 eller TLR4 signalering blev blokeret, var NF-кB/AP-1-respons fra indberetteren af makrofager til adsorberende lysat reduceret, hvilket indikerer, at begge veje er involveret. Men, der var en mærkbart større reduktion i NF-кB/AP-1 aktivitet, når TLR2 signalering blev neutraliseret, hvilket tyder på, at TLR2 kan spille en primær rolle i reaktionen fra reporter makrofager at adsorbede lysate. Vi erkender, at der kan være nogle off-Target hæmning med TLR signalering pathway neutralisere antistoffer og hæmmere. Et neutraliserende antistof blev brugt til at hæmme TLR2 pathway, da der ikke var kommercielt tilgængelige TLR2 inhibitor molekyler på tidspunktet for dette arbejde.
De metoder, der præsenteres her, bruger lysate, som en kompleks kilde til DAMPs, og NF-κB/AP-1 reporter makrofager som en in vitro-model for makrofag respons på DAMPs og andre proteiner adsorbede til polymere biomaterialer (figur 1). Vi forventer, at vores protokol kan bruges til hurtigt at analysere NF-κB/AP-1 svar og upstream TLR signalering af reporter makrofager til en række forskellige materialer (herunder nedbrydelige materialer, porøse stilladser eller hydrogels) og adsorbede protein lag (figur 3). Men brugen af porøse materialer og silicagelrogeler vil introducere kompleksitet i systemet, da det kan være udfordrende at skelne mellem adsorbede molekyler og indgroede molekyler. Vi forventer også, at denne protokol let kan tilpasses til at undersøge bidraget fra andre signalering pathway opstrøms for NF-кb/AP-1 (f. eks C-type lektin receptorer46 og nucleotid-bindende oligomerisering domæne (NOD)-lignende receptorer47) med de relevante hæmmere. Desuden kan NF-κb/AP-1 respons af reporter makrofager sammenlignes mellem forskellige materialer, forudsat respons er normaliseret til baseline celle aktivitet (dvs. celler i medier på hver overflade med ingen præ-adsorbede protein) og alle materialer har målbart endotoksin niveauer.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har intet at afsløre.
Acknowledgments
Forfatterne taknemmeligt anerkender operationel finansiering fra canadiske institutter for sundhedsforskning projekt (PTJ 162251), Queen's University Senatet rådgivende forskningsudvalg og infrastruktur støtte fra det canadiske fundament for innovation John Evans Leadership Fund (projekt 34137) og ministeriet for forskning og innovation Ontario forskningsfond (projekt 34137). L.A.M. blev støttet af en Queen's University R. Samuel McLaughlin Fellowship, et naturvidenskab og ingeniørvidenskab forskning Rådet af Canada Canadian Graduate Scholarship Master's Award og en Ontario Graduate Scholarship. Forfatterne vil gerne takke Dr. Myron Vis for hans generøse gave af NF-κb/AP-1 reporter makrofag Cell line og DRS. Michael blennerhassett og Sandra lourenssen for brugen af deres gel Imaging System og plade læser.
Materials
| Name | Company | Catalog Number | Comments |
| Cell culture reagents | |||
| anti-mouse/human CD282 (TLR2) | Biolegend | 121802 | |
| CLI-095 (TLR4 inhibitor) | Invivogen | TLRL-CLI95 | |
| C57 complement plasma K2 EDTA 10ml, innovative grade US origin | InnovativeResearch | IGMSC57-K2 EDTA-Compl-10ml | Mouse plasma |
| Dulbecco's modified eagle medium (DMEM) | Sigma Aldrich | D6429-500ML | |
| Dulbecco's phosphate buffered saline (DPBS) | Fisher Scientific | 14190250 | No calcium, no magnesium |
| Fetal bovine serum (FBS), research grade | Wisent | 98150 | |
| LPS-EK | Invivogen | TLRL-EKLPS | Lipopolysaccharide from Escherichia coli K12 |
| NIH/3T3 fibroblasts | ATCC | CRL-1658 | |
| Pam3CSK4 | Invivogen | tlrl-pms | Synthetic triacylated lipopeptide - TLR1/2 ligand |
| Penicillin/streptomycin | Sigma Aldrich | P4333-100ML | |
| Plasmocin | Invivogen | ANT-MPP | Mycoplasma elimination reagent |
| RAW-Blue cells | Invivogen | raw-sp | NF-κB/AP-1 reporter macrophage cell line |
| Trypan blue solution, 0.4% | Fisher Scientific | 15250061 | |
| TrypLE express enzyme (1X) | Fisher Scientific | 12604021 | animal origin-free recombinant cell dissociation enzyme |
| Zeocin | Invivogen | ANT-ZN-1 | |
| Kits and assays | |||
| ELISA precoated plates, mouse IL-6 | Biolegend | B213022 | |
| ELISA precoated plates, mouse TNF-α | Biolegend | B220233 | |
| Endotoxin (Escherichia coli) - Control standard endotoxin (CSE) | Associates of Cape Cope Inc. | E0005-5 | Endotoxin for standard curve in chromogenic endotoxin assay |
| LAL water, 100 mL | Associates of Cape Cope Inc. | WP1001 | Used with chromogenic endotoxin assay |
| Micro BCA protein assay | Fisher Scientific | PI23235 | |
| Limulus amebocyte lysate (LAL) Pyrochrome endotoxin test kit | Associates of Cape Cope Inc. | C1500-5 | Chromogenic endotoxin assay reagent |
| QUANTI-Blue alkaline phosphatase detection medium | Invivogen | rep-qb2 | Alkaline phosphatase assay to indirectly measure NF-κB/AP-1 activity |
| Polymeric coating reagents | |||
| Chloroform, anhydrous | Sigma Aldrich | 288306-1L | |
| Ethyl alcohol anhydrous | Commercial Alcohols | P006EAAN | Sigma: Reagent alcohol, anhydrous, 676829-1L |
| Straight tapered fine tip forceps | Fisher Scientific | 16-100-113 | |
| Fluorinert FC-40 solvent | Sigma Aldrich | F9755-100ML | Fluorinated solvent for fPTFE |
| Cell culture grade water (endotoxin-free) | Fisher Scientific | SH30529LS | |
| Poly(methyl methacrylate) (PMMA) | Sigma Aldrich | 182230-25G | |
| Sylgard 184 elastomer kit | Fisher Scientific | 50822180 | |
| Teflon-AF (fPTFE) | Sigma Aldrich | 469610-1G | Poly[4,5-difluoro-2,2-bis(trifluoromethyl)-1,3-dioxole-co-tetrafluoroethylene] |
| Consumables | |||
| Adhesive plate seals | Fisher Scientific | AB-0580 | |
| Axygen microtubes, 1.5 mL | Fisher Scientific | 14-222-155 | |
| Borosilicate glass scintillation vials, with white polypropylene caps | Fisher Scientific | 03-337-4 | |
| Clear PS 48-well plate | Fisher Scientific | 08-772-52 | |
| Clear TCPS 96-well plate | Fisher Scientific | 08-772-2C | |
| Clear TCPS 48-well plate | Fisher Scientific | 08-772-1C | |
| Cover glasses, circles | Fisher Scientific | 12-545-81 | |
| Falcon tissue culture treated flasks, T25 | Fisher Scientific | 10-126-10 | |
| sticky-Slide 8 Well | Ibidi | 80828 | |
| Superfrost microscope slides | Fisher Scientific | 12-550-15 | |
| Tissue culture treated flasks, T150 | Fisher Scientific | 08-772-48 |
References
- Anderson, J. M., Rodriguez, A., Chang, D. T. Foreign body reaction to biomaterials. Seminars in Immunology. 20 (2), 86-100 (2008).
- Anderson, J. M., Miller, K. M.
- Collier, T. O., Anderson, J. M. Protein and surface effects on monocyte and macrophage adhesion, maturation, and survival. Journal of Biomedical Materials Research. 60 (3), 487-496 (2002).
- Bianchi, M. E. DAMPs, PAMPs and alarmins: all we need to know about danger. Journal of Leukocyte Biology. 81 (1), 1-5 (2007).
- McKiel, L. A., Fitzpatrick, L. E. Toll-like Receptor 2-Dependent NF-κB/AP-1 Activation by Damage-Associated Molecular Patterns Adsorbed on Polymeric Surfaces. ACS Biomaterials Science & Engineering. 4 (11), 3792-3801 (2018).
- Babensee, J. E. Interaction of dendritic cells with biomaterials. Seminars in Immunology. 20 (2), 101-108 (2008).
- Sintes, J., Romero, X., de Salort, J., Terhorst, C., Engel, P. Mouse CD84 is a pan-leukocyte cell-surface molecule that modulates LPS-induced cytokine secretion by macrophages. Journal of Leukocyte Biology. 88 (4), 687-697 (2010).
- Tom, J. K., Mancini, R. J., Esser-Kahn, A. P. Covalent modification of cell surfaces with TLR agonists improves and directs immune stimulation. Chemical Communications. 49 (83), 9618-9620 (2013).
- Abdulkhalek, S., et al. Neu1 sialidase and matrix metalloproteinase-9 cross-talk is essential for toll-like receptor activation and cellular signaling. Journal of Biological Chemistry. 286 (42), 36532-36549 (2011).
- Gorbet, M. B., Sefton, M. V.
- Xing, Z., Pabst, M. J., Hasty, K. A., Smith, R. A. Accumulation of LPS by polyethylene particles decreases bone attachment to implants. Journal of Orthopaedic Research. 24 (5), 959-966 (2006).
- Ding, H., et al. Comparison of the cytotoxic and inflammatory responses of titanium particles with different methods for endotoxin removal in RAW264.7 macrophages. Journal of Materials Science: Materials in Medicine. 23 (4), 1055-1062 (2012).
- Hoogenboom, R., Becer, C. R., Guerrero-Sanchez, C., Hoeppener, S., Schubert, U. S. Solubility and thermoresponsiveness of PMMA in alcohol-water solvent mixtures. Australian Journal of Chemistry. 63 (8), 1173-1178 (2010).
- Efimenko, K., Wallace, W. E., Genzer, J. Surface modification of Sylgard-184 poly(dimethyl siloxane) networks by ultraviolet and ultraviolet/ozone treatment. Journal of Colloid and Interface Science. 254 (2), 306-315 (2002).
- Godek, M. L., Sampson, J. A., Duchsherer, N. L., McElwee, Q., Grainger, D. W. Rho GTPase protein expression and activation in murine monocytes/macrophages is not modulated by model biomaterial surfaces in serum-containing in vitro cultures. Journal of Biomaterials Science. Polymer Edition. 17 (10), 1141-1158 (2006).
- Park, J. S., et al. Involvement of Toll-like Receptors 2 and 4 in Cellular Activation by High Mobility Group Box 1 Protein. Journal of Biological Chemistry. 279 (9), 7370-7377 (2004).
- Ohashi, K., Burkart, V., Flohé, S., Kolb, H. Cutting Edge: Heat Shock Protein 60 Is a Putative Endogenous Ligand of the Toll-Like Receptor-4 Complex. The Journal of Immunology. 164 (2), 558-561 (2000).
- Wong, T., McGrath, J. A., Navsaria, H. The role of fibroblasts in tissue engineering and regeneration. British Journal of Dermatology. 156 (6), 1149-1155 (2007).
- van Wachem, P. B., et al. The influence of protein adsorption on interactions of cultured human endothelial cells with polymers. Journal of Biomedical Materials Research. 21 (6), 701-718 (1987).
- Miller, K. M., Anderson, J. M. Human monocyte/macrophage activation and interleukin 1 generation by biomedical polymers. Journal of Biomedical Materials Research. 22 (8), 713-731 (1988).
- Bonfield, T. L., Colton, E., Anderson, J. M. Plasma protein adsorbed biomedical polymers: Activation of human monocytes and induction of interleukin 1. Journal of Biomedical Materials Research. 23 (6), 535-548 (1989).
- González, O., Smith, R. L., Goodman, S. B. Effect of size, concentration, surface area, and volume of polymethylmethacrylate particles on human macrophages in vitro. Journal of Biomedical Materials Research. 30 (4), 463-473 (1996).
- Anderson, J. M., et al. Protein adsorption and macrophage activation on polydimethylsiloxane and silicone rubber. Journal of Biomaterials Science. Polymer Edition. 7 (2), 159-169 (1995).
- Lord, M. S., Foss, M., Besenbacher, F. Influence of nanoscale surface topography on protein adsorption and cellular response. Nano Today. 5 (1), 66-78 (2010).
- Chen, S., et al. Characterization of topographical effects on macrophage behavior in a foreign body response model. Biomaterials. 31 (13), 3479-3491 (2010).
- Shen, M., Horbett, T. A. The effects of surface chemistry and adsorbed proteins on monocyte/macrophage adhesion to chemically modified polystyrene surfaces. Journal of Biomedical Materials Research. 57 (3), 336-345 (2001).
- Love, R. J., Jones, K. S. The recognition of biomaterials: Pattern recognition of medical polymers and their adsorbed biomolecules. Journal of Biomedical Materials Research Part A. 101 (9), 2740-2752 (2013).
- McNally, A. K., Anderson, J. M. Phenotypic expression in human monocyte-derived interleukin-4-induced foreign body giant cells and macrophages in vitro: Dependence on material surface properties. Journal of Biomedical Materials Research Part A. 103 (4), 1380-1390 (2015).
- Gambhir, V., et al. The TLR2 agonists lipoteichoic acid and Pam3CSK4 induce greater pro-inflammatory responses than inactivated Mycobacterium butyricum. Cellular Immunology. 280 (1), 101-107 (2012).
- Suzuki, O., Yagishita, H., Yamazaki, M., Aoba, T. Adsorption of Bovine Serum Albumin onto Octacalcium Phosphate and its Hydrolyzates. Cells and Materials. 5 (1), 45-54 (1995).
- Johnston, R. L., Spalton, D. J., Hussain, A., Marshall, J. In vitro protein adsorption to 2 intraocular lens materials. Journal of Cataract and Refractive Surgery. 25 (8), 1109-1115 (1999).
- Jin, J., Jiang, W., Yin, J., Ji, X., Stagnaro, P. Plasma proteins adsorption mechanism on polyethylene-grafted poly(ethylene glycol) surface by quartz crystal microbalance with dissipation. Langmuir. 29 (22), 6624-6633 (2013).
- Swartzlander, M. D., et al. Linking the foreign body response and protein adsorption to PEG-based hydrogels using proteomics. Biomaterials. 41, 26-36 (2015).
- Chamberlain, M. D., et al. Unbiased phosphoproteomic method identifies the initial effects of a methacrylic acid copolymer on macrophages. Proceedings of the National Academy of Sciences. 112 (34), 10673-10678 (2015).
- Dillman, W. J., Miller, I. F. On the adsorption of serum proteins on polymer membrane surfaces. Journal of Colloid And Interface Science. 44 (2), 221-241 (1973).
- Ishihara, K., Ziats, N. P., Tierney, B. P., Nakabayashi, N., Anderson, J. M. Protein adsorption from human plasma is reduced on phospholipid polymers. Journal of Biomedical Materials Research. 25 (11), 1397-1407 (1991).
- Warkentin, P., Wälivaara, B., Lundström, I., Tengvall, P. Differential surface binding of albumin, immunoglobulin G and fibrinogen. Biomaterials. 15 (10), 786-795 (1994).
- Berghaus, L. J., et al. Innate immune responses of primary murine macrophage-lineage cells and RAW 264.7 cells to ligands of Toll-like receptors 2, 3, and 4. Comparative Immunology, Microbiology and Infectious Diseases. 33 (5), 443-454 (2010).
- Zhang, Y., Karki, R., Igwe, O. J. Toll-like receptor 4 signaling: A common pathway for interactions between prooxidants and extracellular disulfide high mobility group box 1 (HMGB1) protein-coupled activation. Biochemical Pharmacology. 98 (1), 132-143 (2015).
- Mizel, S. B., Honko, A. N., Moors, M. A., Smith, P. S., West, A. P. Induction of macrophage nitric oxide production by Gram-negative flagellin involves signaling via heteromeric Toll-like receptor 5/Toll-like receptor 4 complexes. Journal of Immunology. 170 (12), 6217-6223 (2003).
- Das, N., et al. HMGB1 Activates Proinflammatory Signaling via TLR5 Leading to Allodynia. Cell Reports. 17 (4), 1128-1140 (2016).
- Pelegrin, P., Barroso-Gutierrez, C., Surprenant, A. P2X7 Receptor Differentially Couples to Distinct Release Pathways for IL-1β in Mouse Macrophage. The Journal of Immunology. 180 (11), 7147-7157 (2008).
- Tak, P. P., Firestein, G. S. NF-κB: A key role in inflammatory diseases. Journal of Clinical Investigation. 107 (1), 7-11 (2001).
- Ashkenazi, A., Dixit, V. M. Death receptors: signaling and modulation. Science. 281 (5381), 1305-1308 (1998).
- Erridge, C. Endogenous ligands of TLR2 and TLR4: agonists or assistants. Journal of Leukocyte Biology. 87 (6), 989-999 (2010).
- Feng, Y., et al. A macrophage-activating, injectable hydrogel to sequester endogenous growth factors for in situ angiogenesis. Biomaterials. 134, 128-142 (2017).
- Lonez, C., et al. Cationic lipid nanocarriers activate Toll-like receptor 2 and NLRP3 inflammasome pathways. Nanomedicine: Nanotechnology, Biology, and Medicine. 10 (4), 775-782 (2014).
