

Summary
מיחזור אנדוזומים הם חלק מהרשת הצינורית האנדוסומית. כאן אנו מציגים שיטה לכמת את הדינמיקה של מיחזור אנדוזומים באמצעות GFP-STX13 כסמן אברונים.
Abstract
מיחזור אנדוזומים (REs) הם אברונים צינוריים-ארסיים שנוצרו מאנדוזומים מוקדמים /מיון בכל סוגי התאים. אברונים אלה ממלאים תפקיד מפתח בביוגנזה של מלנוזומים, אברון הקשור ליזוזום המיוצר על ידי מלנוציטים. REs לספק את המטען ספציפי מלנוציט למלנוזומים מוקדמים במהלך היווצרותם. חסימה בדור של REs, נצפתה במספר מוטציות של תסמונת הרמנסקי-Pudlak, גורמת hypopigmentation של העור, השיער, והעין. לכן, לימוד הדינמיקה (מתייחס למספר ולאורך) של REs שימושי כדי להבין את הפונקציה של אברונים אלה במצבים נורמליים ומחלות. במחקר זה, אנו שואפים למדוד את הדינמיקה RE באמצעות תושב SNARE STX13.
Introduction
ביוסינטזה של פיגמנטים מלנין מתרחשת מלנוזומים, אברון ספציפי מלנוציטים (LRO) כי קיים יחד עם ליזוזומים קונבנציונליים. המערכת האנדוציטית ממלאת תפקיד מרכזי בביוגנזה של מלנוזומים, הנדרשת לצבע עור והגנה פוטו-הגנתית מפני קרינה מייננת1,2,3. במהלך תהליך זה, אנזימי סינתזה מלנין ממוינים על אנדוזומים מוקדמים / מיון ולאחר מכן מועברים מלנוזומים מוקדמים באמצעות אנדוזומים צינוריים או אופניים הנקראים אנדוזומים מיחזור (REs)4,5,6,7,8,9,10. פילוח והיתוך של אברונים אלה לווסת את ההתבגרות של מלנומוזומים פיגמנטיים פונקציונליים לחלוטין7,11,12,13,14. פגמים בהיווצרות של אברונים אלה או מיון מטען לאברונים אלה גורמים לבקנות oculocutaneous ופנוטיפים קליניים אחרים, שנצפו תסמונת הרמנסקי-Pudlak15,16.
כאן אנו מתארים טכניקה פשוטה המבוססת על מיקרוסקופיה כדי ללמוד ולנתח את REs. בשיטה זו, ניצלנו חלבון transmembrane, Qa-SNARE Syntaxin (STX)13 התב"ע המונה על מיחזור אנדוזומים17 ומחזורים בין מיון אנדוזומים ומלנוזומים במלנוציטים12,18. יתר על כן, מחיקה של N-terminal לא מובנה דומיין רגולטורי (כלומר SynN או STX13Δ129) מאפשרת SNARE להיתקע melanosomes, אשר מודד את מסלול הסחר קדימה לכיוון melanosome12. השתמשנו בסמן אנדוסומאלי ידוע מיחזור Rab GTPase (Rab)11 במחקרים שלנו14,19. הדמיית פלואורסצנטיות של החלבונים GFP-STX13WT, GFP-STX13Δ129, mCherry-Rab11 ו- TYRP1 במלנוציטים מסוג בר ואחריו כימות הלוקליזציה היחסית שלהם תספק את הטבע והדינמיקה של REs בנוסף למיקודם למלנוזומים. לכן, זוהי טכניקה פשוטה שניתן להשתמש בה כדי לדמיין ולמדוד את הדינמיקה של REs במלנוציטים.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
הפרוטוקול כרוך זריעת מלנוציטים ואחריו טרנספקטו של plasmids. צעדים נוספים כוללים קיבעון, חיסונים, הדמיה וניתוח של התאים כדי למדוד את האורך והמספר של REs. התיאור המפורט של הפרוטוקול ניתן להלן.
1. זריעת מלנוציטים של עכברים על כיסויים שטופלו מראש
- מצפים את כיסויי הזכוכית בצלחת פטרי (כלומר, 4 - 5 בצלחת 35 מ"מ) עם מטריצת ממברנה במרתף בינונית (1:20 במדיום RPMI מלא: RPMI + 10% חום מומת FBS + 1x גלוטמין + תערובת אנטיביוטיקה 1x) ולייבש אותו במכסה המנוע תרבית הרקמות במשך 15 דקות. יש לשטוף את כיסויי הכיסוי פעם אחת עם 1x PBS לפני השימוש.
- שמור על מלנוציטים מסוג עכבר בר (melan-Ink4a-Arf-1 מ- C57BL/6J, a/a, Ink4a-Arf-/- עכברים, המתוארים ב-20 וזמינים בבנק תא הגנומיקה הפונקציונלי של Welcome Trust) בצלחת פטרי (כלומר, צלחת 35 או 60 מ"מ) בתוספת מדיום RPMI מלא.
- לשטוף את התאים פעמיים עם 1x PBS ולהוסיף 0.5 או 1 מ"ל של טריפסין-EDTA (0.25%) פתרון עבור תאים טריפסינים. לדגור על התאים ב 37 °C (5 °F) במשך 2 - 5 דקות עבור הניתוק מן צלחת פטרי.
- הוסף 1 - 2 מ"ל של מדיום RPMI מלא, להשעות ולאחר מכן להעביר את התאים לצינור צנטריפוגה.
- צנטריפוגה השעיית התא ב 4 °C (5 °F), 376 x g במשך 5 דקות ולאחר מכן resuspend הכדור ב 1x PBS.
- חזור על שלב הצנטריפוגה והגדל מחדש את התאים ב- 1 מ"ל של מדיום RPMI מלא.
- זרע את התאים על קרום המרתף כיסויים בציפוי בינוני ב 50-60% conהשפעה (כ 6 x 105 תאים על צלחת תרבית תאים 35 מ"מ המכיל 4 - 5 כיסויים). תמיד להוסיף 200 ננומטר של PMA (להוסיף 5 μL של מלאי עובד 40 μM phorbol 12-myristate 13-אצטט) להשעיית תא מצופה במדיום RPMI מלא.
- לאחר זריעה, לדגור על הצלחת ב 37 °C (50 °F) במשך 12-24 שעות.
2. טרנספקטציה של תאים עם פלסמידים STX13
- השתמש ריאגנטים הבאים: pEGFP-C1-STX13WT ו pEGFP-C1-STX13Δ129 (מתואר ב12). mCherry-Rab11, הייתה מתנה חביבה מגראסה רפוסו, מכון קירי, פריז (מתואר בהפניה19). נוגדן נגד TYRP1 מ-ATCC (TA99).
- לאחר 12 - 24 שעות של זריעה, לבצע חלוקה של התאים עם plasmids באמצעות ריאגנט transfection מבוסס שומנים בדם. עבור צלחת 35 מ"מ, לקחת 5 μL של ריאגנט transfection ב 250 μL של OPTI-MEM בינוני בצינור microcentrifuge ולקחת כ 200 ננוגרם של כל plasmid ב 250 μL של OPTI-MEM.
- לדגור על הצינורות המכילים DNA ואת ריאגנט transfection במשך 5 דקות. לערבב ללא pipetting חוזר (נפח כולל יהיה כ 500 μL). דגירה במשך 30 דקות בטמפרטורת החדר. בצע הקשה ידנית של הצינור כל 10 דקות במשך 30 דקות ב RT.
- במהלך הדגירה, לשטוף את התאים פעמיים עם 1x PBS, פעם אחת עם OPTI-MEM ולאחר מכן להוסיף 1 מ"ל של OPTI-MEM לתאים.
- לאחר 30 דקות של דגירה, להוסיף את תערובת ריאגנט-DNA transfection לתאים בצורה טיפה חכם על ידי כיסוי המנה.
- לדגור על התאים ב 37 °C (6 °F) במשך 6 שעות. שאפו את המדיום OPTI-MEM עם ריאגנט טרנס-זיהום והוסיפו מדיום RPMI מלא בתוספת 200 ננומטר של PMA.
- לדגור על התאים ב 37 °C (55 °F) עבור 48 שעות.
3. קיבעון התאים
הערה: ההליך הבא מבוצע מחוץ למכסה המנוע של תרבית הרקמות.
- לאחר 48 שעות של transfection, לשטוף את התאים פעמיים עם 1x PBS ולאחר מכן לתקן את התאים עם 3% פורמלדהיד (מוכן טרי 1x PBS) במשך 30 דקות.
- לאחר קיבעון, לשטוף את התאים פעמיים עם 1x PBS ולאחסן את כיסויים ב 1x PBS עד לשימוש נוסף. לחלופין, תאים יכולים להיות מותקנים על שקופיות זכוכית (ראה להלן) או מאוחסנים ב 4 °C (70 °F).
4. חיסונים של התאים
- מכינים תא לח: מניחים חתיכת סרט פרפין חתוכה על נייר סינון לח בצלחת פטרי, מכוסה בנייר אלומיניום.
- הכן 25 μL של פתרון נוגדנים ראשוני (0.2% סאפונין ב 1x PBS, 0.1% BSA ב 1x PBS ו 0.02% נתרן אזיד ב 1x PBS). הוסף נוגדן בדילול של 1: 200. הוסף פתרון זה כטיפה על סרט פרפין בחדר לח.
- הרימו בזהירות את כיסוי המכסה במלקחיים, הפוך אותו על טיפה זו של תמיסה מכתימה נוגדנים ראשונית ולאחר מכן לכסות את המכסה של התא לח. דגירה בטמפרטורת החדר במשך 30 דקות.
- באופן דומה, להכין את פתרון הנוגדנים המשני בדילול של 1:500 ולהניח אותו על סרט פרפין ליד כיסוי בתא לח. להכתמת הגרעין, הוסף DAPI (1:20,000 עד 1:30,000) לפתרון.
- בעזרת מלקחיים, הרימו בזהירות את כיסוי הכיסוי מתמיסת הנוגדנים העיקרית וטובלים אותו שלוש פעמים ב-1x PBS (בכוס זכוכית).
- הקש על כיסוי על נייר טישו כדי להסיר את PBS עודף על כיסוי. מניחים אותו על תמיסת כתמי נוגדנים משנית בתא הלח ולא חושפים אותו לאור בשל נוכחותם של נוגדנים מתויגים פלואורסצנטית בתמיסה.
- לדגור על כיסוי שוב במשך 30 דקות בטמפרטורת החדר. אנא שימו לב תמיד לצד כיסוי הכולל את התאים לאורך שלבים אלה.
- לאחר הדגירה, בזהירות להרים את כיסוי מפתרון הנוגדנים המשני ולאחר מכן לטבול אותו שלוש פעמים ב 1x PBS. יתר על כן, הקש על כיסוי על נייר טישו כדי להסיר את PBS עודף על כיסוי.
- מניחים 12 μL של ריאגנט הרכבה פלואורומאונט-G על מגלשת זכוכית ומניחים בזהירות את כיסויי הצבע (הפונים לכיוון הזכוכית) על ריאגנט הרכבה. הפוך את שקופית הזכוכית על נייר טישו ולאחר מכן לחץ בעדינות.
5. מיקרוסקופיית פלואורסצנטיות של התאים
- צלם את התאים המוכתמים תחת מסנני שדה בהיר (BF) ופלואורסצנטיות (IF) באמצעות מיקרוסקופ פלואורסצנטי הפוך המצויד במצלמת CCD באמצעות מטרה אפיכרומטית 60x (שמן) או כל מיקרוסקופ אחר עם תצורה דומה.
6. כימות חפיפה בין החלבונים המותאמים לשפות רה ומלנוזומים:
הערה: השלבים הבאים מתבצעים לכימות מקדם החפיפה של מנדר בין החלבונים באמצעות תוכנת פיג'י (להורדה חופשית מהקישור: https://imagej.net/software/fiji/). השתמש בתמונת TIFF עם ערוצים מרובים.
- פתח את התמונה הגולמית. עבור אל האפשרות תמונה, בחר צבע | פצל ערוצים והשתמש בשני הערוצים לניתוח.
- פתח את תוסף JACoP באפשרות תוסף.
- הגדר את הסף עבור שני הערוצים כך שכל הנקודות הבהירות נבחרו והרקע יבוטל.
- עבור אל האפשרות ניתוח, בחר מקדמי M1 ו- M2 לקבלת מקדם החפיפה של מנדר.
- לחץ על האפשרות נתח בתוסף JACoP וראה את התוצאה המציגה את מקדם החפיפה של מנדר.
7. כימות המספר והאורך הצינוריים של אנדוזומים:
הערה: השלבים הבאים מתבצעים לכימות המספר והאורך של הצינורות באמצעות תוכנת פיג'י.
- פתחו את האפשרות 'עבור לתמונה' של התמונה, בחרו 'צבע |' פצל ערוצים והשתמש בערוץ הרצוי לניתוח.
- עבור שוב לתמונה , בחר הקלד | המר לתמונת 8 סיביות.
- לאחר מכן עבור אל תוספים, בחר ניתוח | צינורות. הגדר את ערך סיגמא ב- 0.1075 . לחץ על בסדר.
- עבור שוב לתמונה , בחר הקלד | המר לתמונת 8 סיביות.
- עבור אל | תמונה כוונון | Threshold (השתמש באותם ערכי סף עבור כל התמונות). תמונות צריכות להיות בעוצמה שווה בקירב).
- עבור אל עיבוד | | בינארי המר למסיכה.
- עבור אל תהליך, בחר בינארי ולאחר מכן בחר שלד.
- עבור אל ניתוח, בחר שלד ובחר ניתוח שלד. בתוצאה ובפלט | בחר (א) חשב את הנתיב הקצר ביותר | (ב) להציג מידע מפורט | (ג) שלד תצוגה שכותרתו. לחץ על בסדר.
הערה: התוצאה תיפתח בתבנית המנוולת בכרטיסיות. העמודה של אורך הענף הממוצע מציגה את אורך כל הצינורות השונים בתא שנבחר (הגדר קנה מידה לקבלת ערכים במיקרומטרים). - לקבלת מספר הצינורות בתא, עבור אל ניתוח ובחר באפשרות נתח חלקיקים . לחץ על בסדר.
הערה: בתוצאות שהושגו, עמודת הספירה מציגה את מספר הצינורות בתא מסוים זה. - שמור את הנתונים וניתח.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
כימות של לוקליזציה מוטנטית STX13Δ129 למלנוזומים
מיקרוסקופיה אימונופלואורסצנטית של STX13 במלנוציטים מסוג בר עכברים הראתה GFP-STX13WT מקומי כמבנים ריסקוליים וצינוריים ו- GFP-STX13Δ129 המותאמים לשפות מקומיים כמבנים דמויי טבעת בנוסף למשטח התא (איור 1A). יתר על כן, GFP-STX13Δ129 דמוי טבעת תאית הראה colocalization עם חלבון מלנום TYRP1 (איור 1A) ומלנוזומים בתמונה בהירה (נתונים שאינם מוצגים)12. כפי שמוצג קודם לכן, קבוצה של GFP-STX13WT בדפוס יתר נצפתה במלאנוסומים12. כדי למדוד את הלוקליזציה היחסית של GFP-STX13WT ו- GFP-STX13Δ129 למלנוזומים, השתמשנו בפיג'י וניתחנו עם תוסף JACoP. מקדם החפיפה (MOC) של מנדר הנמדד בין GFP-STX13Δ129 עם TYRP1 גבוה בכ-1.5 לעומת GFP-STX13WT עם TYRP1 (איור 1B). באופן מעניין, TYRP1 הראה 2.9 פי כמה ערכי MOC גבוהים יותר עם GFP-STX13Δ129 בהשוואה ל- GFP-STX13WT (איור 1B). נתונים אלה מצביעים על כך שהלוקליזציה של GFP-STX13Δ129 למלנוזומים גבוהה יחסית בהשוואה ל- GFP-STX13WT במצב יציב.
כימות לוקליזציה של STX13WT לאנדוזומים של מיחזור
מיקרוסקופיה חיסונית של GFP-STX13WT הראתה קולוקליזציה עם חלבון אנדוסומאלי מיחזור ידוע Rab11 (שבא לידי ביטוי בשם mCherry-Rab11) (איור 2A,B). MOC נמדד בין GFP-STX13WT עם mCherry-Rab11 הוא כ 1.4 פיפולים גבוה יותר בהשוואה mCherry-Rab11 עם GFP-STX13WT (איור 2B). כדי למדוד את המספר והאורך של צינורות אנדוסומאליים חיוביים GFP-STX13WT, השתמשנו בתוכנת פיג'י כמתואר בסעיף הפרוטוקול. mCherry-Rab11 משמשת כשליטה חיובית בניסויים (איור 2). מלאנוציטים שהודבקו ב-GFP-STX13WT הראו מספר גבוה יותר של צינורות לתא בהשוואה לתאים המבטאים mCherry-Rab11 (איור 2C גרף עליון, השווה את בר A עם בר B). עם זאת, מספרי הצינורות מצטמצמים בביטוי משותף של GFP-STX13WT ו-mCherry-Rab11 בתאים (איור 2C גרף עליון, השוואת בר A עם C ובר B עם D). באופן מעניין, אורך הצינור הממוצע (מיקרומטר) הן עבור GFP-STX13WT והן עבור mCherry-Rab11 דומה זה לזה בתאים המבטאים בנפרד או יחד (איור 2C גרף תחתון). יחד, נתונים אלה מראים כי GFP-STX13WT לוקליזציה REs דומה Rab11.
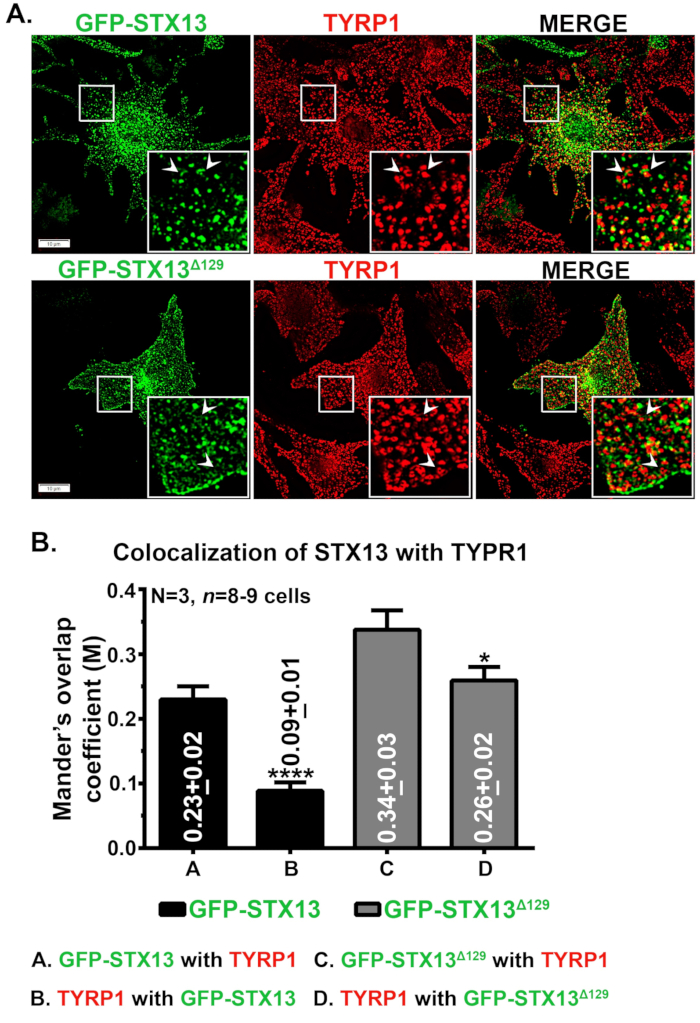
איור 1: לוקליזציה של GFP-STX13WT ו- GFP-STX13Δ129 למלנוזומים במלנוציטים מסוג בר. (A) מלנוציטים מסוג מלן-דיו4a-Arf-1 הודבקו עם GFP-STX13WT ו- GFP-STX13Δ129. התאים תוקנו, הוכתמו בנוגדן נגד TYRP1 ולאחר מכן נותחו על ידי מיקרוסקופיה פלואורסצנטית. ערכות הן תצוגה מוגדלת של האזורים בקופסה הלבנה. החצים מצביעים על לוקליזציה של GFP-STX13WTto REs ו- GFP-STX13Δ129 למלנוזומים. מוטות קנה מידה, 10 מיקרומטר. (ב) כימות של colocalization בין STX13 ו- TYRP1. מקדם החפיפה (M) של מנדר בין GFP-STX13WT או GFP-STX13Δ129 עם TYRP1 ולהיפך מיוצג (ממוצע ± S..M E.) בנפרד בעלילה. N = 3. *p ≤0.05 ו****p ≤0.0001. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

איור 2: לוקליזציה של GFP-STX13WT למיחזור אנדוזומים במלנוציטים מסוג בר. (A) מלנוציטים מסוג מלן-דיו4a-Arf-1 הועברו ל-GFP-STX13WT ו-mCherry-Rab11. התאים תוקנו ונותחו על ידי מיקרוסקופיית פלואורסצנטיות. ערכות הן תצוגה מוגדלת של האזורים בקופסה הלבנה. החצים מצביעים על לוקליזציה של GFP-STX13WT לתאים mCherry-Rab11-חיוביים. מוטות קנה מידה, 10 מיקרומטר. (ב) כימות של colocalization בין STX13 ו Rab11. מקדם החפיפה של מנדר (M) בין GFP-STX13WT עם mCherry-Rab11 ולהיפך מיוצג (ממוצע ± S.E.M.) בנפרד בעלילה. N = 3. עמ' ≤0.001. ג. כימות מספר ואורך (ב- μm) של STX13- או ר.ס.ז' חיובי רב 11. המספר הממוצע של צינורות / תא ואורך הצינור הממוצע של GFP-STX13WT ו mCherry-Rab11 מיוצגים (ממוצע ± S..M E.E.) בנפרד בעלילה. N = 3. שים לב כי התאים מועברים עם GFP-STX13WT ו mCherry-Rab11 ב (C) ו (D). אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
מיחזור אנדוזומים הם קבוצה של אברונים אנדוציטיים, והם מתווכים את מיחזור המטען למשטח התא בכל סוגי התאים21,22,23,24,25. בסוגי תאים מיוחדים כגון מלנוציטים, אברונים אלה מסיטים חלקית את נתיב הסחר שלהם לכיוון המלנוזומים עבור הביוגנזה שלהם3,16,26. יתר על כן, REs נוצרים לאחר מיון מטען על אנדוזומים מוקדמים / מיון ומורפולוגית מופיעים כמבנים צינוריים-ריסקולים. הדינמיקה שלהם (מספר ואורך) תלויה במספר גורמים תאיים, כולל חלבונים מוטוריים21,22,23,24,25,27,28. מחקרים מעטים השתמשו Rab11 כסמן לתייג את REs במלנוציטים14,19. בעוד שאינם מלנוציטים, STX13 ו- KIF13A שימשו בנוסף ל- Rab11 כדי לדמיין ולאפיין REs13,17,19. במחקר זה השתמשנו ב-STX13 כסמן לתיוג ה-REs במלנוציטים ואחריו השוואה עם צינורות חיוביים של Rab11 (איור 2). באופן מעניין, מחיקת מסוף N ב- STX13 (GFP-STX13Δ129) גורמת להקצאה שגויה של ה- SNARE למלנוזומים (איור 1). לפיכך, לוקליזציה של GFP-STX13WT ו- GFP-STX13Δ129 במלנוציטים לדמיין את REs ומלנוזומים, בהתאמה. חלבונים אלה יכולים לשמש סמני אברונים בהתאמה למחקרי לוקליזציה של מדינה יציבה.
מחקרים הראו כי מספר חלבונים כגון TYRP1, TYR (טירוזינאז), VAMP7, OCA2, Rab32/38 למקומים את המלנוזומים בנוסף לאברונים תאיים אחרים3,29. המחקרים שלנו הראו כי מוטציית STX13 שנמחקה ממסוף N (GFP-STX13Δ129) לוקליזציה למלנוזומים וקרום פלזמה. אנו משערים כי GFP-STX13Δ129 יכול לשמש ככתב כדי ללמוד את הסחר מ REs אל פני השטח התא LRO. לעומת זאת, GFP-STX13WT לוקליזציה REs דומה Rab11. המחקרים שלנו המחישו כי GFP-STX13WT יכול לשמש גם לסימון REs במלנוציטים. אנו צופים כי GFP-STX13WT עשוי להיות סמן RE טוב יותר מאשר Rab11 מאז ביטוי יתר Rab11 משנה את הדינמיקה אנדוסומלית. בסך הכל, GFP-STX13WT משמש כסמן RE פוטנציאלי כדי ללמוד את הדינמיקה שלהם במצב יציב.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
המחברים מצהירים כי אין להם ניגוד אינטרסים.
Acknowledgments
עבודה זו נתמכה על ידי המחלקה לביוטכנולוגיה (BT/PR32489/BRB/10/1786/2019 ל-SRGS); ועדת מחקר מדע והנדסה (CRG/2019/000281 ל-SRGS); DBT-NBACD (BT/HRD-NBA-NWB/38/2019-20 ל-SRGS) ותוכנית השותפות IISc-DBT (ל-SRGS). התשתית במחלקה נתמכה על ידי DST-FIST, DBT ו- UGC. AMB נתמך על ידי DBT-JRF (DBT/2015/IISc/NJ-02).
Materials
| Name | Company | Catalog Number | Comments |
| anti-TYRP1 antibody (TA99) | ATCC | HB-8704 | |
| Fluoromount-G | Southern Biotech | 0100-01 | |
| Lipofectamine 2000 | ThermoFisher Scientific | 11668-500 | |
| Matrigel matrix | BD Biosciences | 356231 | |
| OPTI-MEM | ThermoFisher Scientific | 022600-050 | |
| Phorbol 12-myristate 13-acetate | Sigma-Aldrich | P8139 | |
| RPMI Medium 1640 | ThermoFisher Scientific | 31800-022 |
References
- Dell'Angelica, E. C. The building BLOC(k)s of lysosomes and related organelles. Current Opinion in Cell Biology. 16 (4), 458-464 (2004).
- Raposo, G., Marks, M. S. Melanosomes--dark organelles enlighten endosomal membrane transport. Nature Reviews in Molecular Cell Biology. 8 (10), 786-797 (2007).
- Ohbayashi, N., Fukuda, M. Recent advances in understanding the molecular basis of melanogenesis in melanocytes. F1000Research. 9, F1000 Faculty Rev-608 (2020).
- Theos, A. C., et al. Functions of adaptor protein (AP)-3 and AP-1 in tyrosinase sorting from endosomes to melanosomes. Molecular Biology of the Cell. 16 (11), 5356-5372 (2005).
- Di Pietro, S. M., et al. BLOC-1 interacts with BLOC-2 and the AP-3 complex to facilitate protein trafficking on endosomes. Molecular Biology of the Cell. 17 (9), 4027-4038 (2006).
- Setty, S. R., et al. BLOC-1 is required for cargo-specific sorting from vacuolar early endosomes toward lysosome-related organelles. Molecular Biology of the Cell. 18 (3), 768-780 (2007).
- Delevoye, C., et al. AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis. Journal of Cell Biology. 187 (2), 247-264 (2009).
- Bultema, J. J., Ambrosio, A. L., Burek, C. L., Di Pietro, S. M. BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles. Journal of Biological Chemistry. 287 (23), 19550-19563 (2012).
- Sitaram, A., et al. Differential recognition of a dileucine-based sorting signal by AP-1 and AP-3 reveals a requirement for both BLOC-1 and AP-3 in delivery of OCA2 to melanosomes. Molecular Biology of the Cell. 23 (16), 3178-3192 (2012).
- Nag, S., et al. Rab4A organizes endosomal domains for sorting cargo to lysosome-related organelles. Journal of Cell Science. 131 (18), (2018).
- Dennis, M. K., et al. BLOC-2 targets recycling endosomal tubules to melanosomes for cargo delivery. Journal of Cell Biology. 209 (4), 563-577 (2015).
- Jani, R. A., Purushothaman, L. K., Rani, S., Bergam, P., Setty, S. R. STX13 regulates cargo delivery from recycling endosomes during melanosome biogenesis. Journal Cell Science. 128 (17), 3263-3276 (2015).
- Shakya, S., et al. Rab22A recruits BLOC-1 and BLOC-2 to promote the biogenesis of recycling endosomes. EMBO Reports. 19 (12), 45918 (2018).
- Bowman, S. L., et al. A BLOC-1-AP-3 super-complex sorts a cis-SNARE complex into endosome-derived tubular transport carriers. Journal of Cell Biology. 220 (7), 202005173 (2021).
- Wei, M. L. Hermansky-Pudlak syndrome: a disease of protein trafficking and organelle function. Pigment Cell Research. 19 (1), 19-42 (2006).
- Bowman, S. L., Bi-Karchin, J., Le, L., Marks, M. S. The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic. 20 (6), 404-435 (2019).
- Prekeris, R., Klumperman, J., Chen, Y. A., Scheller, R. H. Syntaxin 13 mediates cycling of plasma membrane proteins via tubulovesicular recycling endosomes. Journal of Cell Biology. 143 (4), 957-971 (1998).
- Mahanty, S., et al. Rab9A is required for delivery of cargo from recycling endosomes to melanosomes. Pigment Cell Melanoma Research. 29 (1), 43-59 (2016).
- Delevoye, C., et al. Recycling endosome tubule morphogenesis from sorting endosomes requires the kinesin motor KIF13A. Cell Reports. 6 (3), 445-454 (2014).
- Ha, L., et al. ARF functions as a melanoma tumor suppressor by inducing p53-independent senescence. Proceedings of the National Academy of Science U. S. A. 104 (26), 10968-10973 (2007).
- Soldati, T., Schliwa, M. Powering membrane traffic in endocytosis and recycling. Nature Reviews Molecular Cell Biology. 7 (12), 897-908 (2006).
- Grant, B. D., Donaldson, J. G. Pathways and mechanisms of endocytic recycling. Nature Reviews Molecular Cell Biology. 10 (9), 597-608 (2009).
- Hsu, V. W., Prekeris, R.
- Taguchi, T.
- Goldenring, J. R.
- Delevoye, C., Marks, M. S., Raposo, G. Lysosome-related organelles as functional adaptations of the endolysosomal system. Current Opinion in Cell Biology. 59, 147-158 (2019).
- Hsu, V. W., Bai, M., Li, J. Getting active: protein sorting in endocytic recycling. Nature Reviews in Molecular Cell Biology. 13 (5), 323-328 (2012).
- Desfougeres, Y., D'Agostino, M., Mayer, A. A modular tethering complex for endosomal recycling. Nature Cell Biology. 17 (5), 540-541 (2015).
- Le, L., Sires-Campos, J., Raposo, G., Delevoye, C., Marks, M. S. Melanosome biogenesis in the pigmentation of mammalian skin. Integrated Computational Biology. 61 (4), 1517-1545 (2021).
