


Summary
迅速
Abstract
体内活性筛选化合物可以用来作为第一步,以确定可能到1,2药物制剂开发的候选人。我们开发了一种新型的nanoinjection /电检测,使检测的神经电路介导的在果蝇 3,4逃生响应功能的生物活性化合物的调节作用。我们在体内实验中,使用果蝇巨纤维系统(GFS, 图1)允许不同类型的化合物,如小分子肽的筛选,只需要最小数量引起的效果。此外,果蝇的飞行服务队提供了一个潜在的分子靶点神经或肌肉的种类繁多。 (GFS)巨纤维突触电(峡路口)以及化学(胆碱)到外围的中间神经突触(PSI)和“Tergo Trochanteral肌肉神经元(TTMn)5 6。最后,神经肌肉接头(NMJ)的TTMn和DLMn跳(TTM)和飞行肌(DLM),是谷氨酸7-12。在这里,我们演示了如何注入化合物的纳升数量级,而获得巨人光纤系统,以及如何监察该电路的功能复合的影响的电生理细胞内记录。我们显示枸橼酸methyllycaconitine(MLA)的,胆碱受体拮抗剂,破坏PSI的DLMn连接,但不能连接或跳跃或飞行肌肉的功能NMJ的GF TTMn的的的特异性。
在这部影片开始之前,它是至关重要的,你仔细地观察和熟悉的朱庇特的视频名为“电生理记录,从四melan巨纤维通路“ogaster从奥古斯丁等 7,视频这里的目的是为这一现有技术的扩展。在这里,我们使用的细节,只对另外的的配对nanoinjections和监测技术的电生理记录方法和重点。
Protocol
1。电钻机设置
- 详细描述了为电钻机设置所需的设备由奥古斯丁等 。在该杂志14。为所需的电生理仪器的详细解释,请参阅本文。
- 加入了第六的微操作机器人,其持有的nanoinjector修改先前所描述的电钻机设置14。为方便应置于两者之间的刺激电极显微操作,在图2所示的动物的头。
- 在实验开始前,确保你有一个旋转轴与所有显微操作的舒适范围,所有电极以及注射微量(见步骤2),可以达到动物。
2。 nanoinjection集行动
- 设置的nanoinjection需要1 Nanoliter2000(世界Precisi仪器,萨拉索塔,佛罗里达州,美国)或类似的喷油器的类型,允许在纳升数量控制注射。
- 准备用玻璃微电极,电极拔拔80-100MΩ的电阻与喷油器提供的注射针头。
- 为顺利注射,它需要的微电极锥在45度角的11-17微米开幕( 图3)。
- 慢慢回填注射微量合成油使用Hamilton注射器由Nanoliter2000手册的指示,确保无气泡存在。
- 仔细确保上nanoinjector微量的和准备来载入排空多余的油由Nanoliter2000手册的指示,该化合物。
- 放置在微操作机器人的喷油器和加载的复合在Nanoliter2000手册的指示。确保微量的一角,在此期间不中断程序。
- 设置所需数量由Nanoliter2000手册的指示,在喷油器的控制盒注射纳升。请注意,总金额注入不应超过100 NL。我们发现,大批量的生理盐水对照解决方案可能会影响政府飞行服务队的电路功能。
- 注射器拔出从控制箱与注射本身的异常,因为在拍摄过程中采集的nanoinjector电源干扰与录音,这是作为背景噪声( 图4)可见,这是至关重要的。但是,不要断开电源控制箱,因为它会重新设置它。
3。 果蝇准备
- 麻醉与CO 2或冰2至6天的老苍蝇如前所述14,15。
- 一旦动,使用镊子动物转移到中小板无线TH软牙科用蜡采摘它从它的腿。请注意,雄性果蝇的重量约1.0毫克,重约1.2毫克,一个女的果蝇,因此复合体的重量比是在男性和女性的苍蝇不同。因此,建议使用只有一个性别实验。
- 正如先前所述14,15,精心安装飞背侧,确保胸部和头部被固定放置在身体周围的软牙科蜡。仔细传播的翅膀,让他们躺在胸部( 图2,C),垂直。苍蝇应安装用尽可能少的损害。
4。配对Nanoinjection /电
- 将其头部对刺激电极的电钻机上安装飞。
- 如前所述14,15(六钉与相应的刺激,地面和记录电极的动物igure 2)。除非需要,否则进入到TTM肌肉DLM和其他的地方之一记录电极。 DLM的肌肉位于胸部之间的前后背成中央毛发和苍蝇中线。过去一年肌肉靠近机翼附件,后部和前跨阿拉尔飞毛7之间。
- 注射微量含有的化合物的三个单眼位于头后部内侧,但不注入尚未( 图2,C),中心对齐。
- 复方注射之前获得的GF基线记录 过去一年和GF,DLM巨人光纤系统(政府飞行服务队, 图1)通过刺激大脑的途径。这样做,激活10列车10刺激(40-50毫伏)在100赫兹之间的列车14,15( 图4)延迟1秒的持续时间为0.03毫秒每个巨纤维(GFS)。野生型飞SH乌尔德是能够遵循这种刺激率DLM的和TTM途径之一到一。丢弃飞DLM和GF GF的过去一年的途径,不遵循100 Hz刺激在1:1的比例。
- 切换到连续刺激的GF在1 Hz的单脉冲( 图4)。
- 快速插入控制盒注射器。即使背景噪音会干扰1 Hz的刺激录音,不停止。
- 小心地插入到略低于飞cuticule的头壳微量注射,注入飞淋巴化合物所需的金额,同时保持1 Hz的刺激( 图4)。由于飞的开放式循环系统,整个神经系统会受到的化合物,在几秒钟内。虽然一个特定的注射部位是不能提供化合物进入淋巴的关键,我们发现单眼的地区,这是本地化ð在头壳最背侧,是一个方便的网站,允许一个简单的注射化合物的分布迅速,甚至导致内侧。
- 立即从注射部位注射微量拔下喷油器从控制箱,1 Hz的快刺激,同时继续注射后1分钟( 图4)。
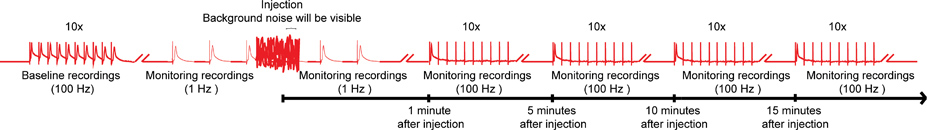
- 为了揭示政府飞行服务队的化合物更微妙的影响,强调在100赫兹的巨纤维(GFS)10 10刺激的列车与列车之间的延迟1秒。继续测试这个范例中,每5分钟至15分钟( 图4)的GF通路的功能。然而,较短的时间间隔或更长的监测期间也有可能。
- 为了检验该化合物是否具有在神经肌肉接头(NMJs)政府飞行服务队的作用,并可能缩小化合物的影响,继续激活运动神经元方向TLY由胸刺激。对于这一点,从眼睛的刺激电极,取代他们的胸部前两侧10 10刺激的列车,以刺激运动神经元,在100赫兹。
注:电在视频显示的痕迹不相符的纯染料注射的效果。
5。代表结果
拮抗剂对PSI DLM巨人光纤系统突触的影响
methyllycaconitine柠檬酸(MLA)是胆碱受体拮抗剂,是α7胆碱受体亚型的具体。 PSI的以DLMn GF-DLM的通路中的突触是正常功能Dα7胆碱受体亚型的依赖,同时去除Dα7胆碱受体亚型基因的GF过去一年通路5,6没有影响。为了证明我们检测的灵敏度和特异性,我们注入工作重点在不同浓度(0,0.02,0.04,0。08,注入0.12毫微克/毫克,46 NL)到动物的头(N = 10%复合处理; N = 15生理盐水治疗)。只雄蝇(野生型基因型野生10E),和化合物的效果监测,共注射后15分钟。
图5描述了注射前获得的基线的工作重点和生理盐水对照溶液注射后获得的录音和之间的差异。我们发现,注入的工作重点在GF-DLM的途径无法遵循通过刺激大脑中的政府飞行服务队一到100赫兹,而GF过去一年途径不受影响。 ( 图5,顶部和中间的痕迹,t-检验生理盐水对照组[1纳克/毫克和工作重点,在每个时间点不同浓度之间进行,除非数据非参数正态性和方差相等测试,否则,我们使用1 Mann-Whitney秩和检验。* P <0.001)。然而,一到一个Response的DLM的观察时,直接刺激运动神经元( 图5,底部跟踪),表明DLM的和TTM NMJ功能不影响工作重点。工作重点出现注射后达到其最大作用1分钟,0.04,0.08和0.12毫微克/毫克注射工作重点,因为没有进一步的显着变化,在接下来的15分钟的测试期间所指出的。此外,该化合物达到0.08毫微克/毫克的最大作用,因为更强的反应并没有观察到高剂量的0.12毫微克/毫克。

图1。系统(GFS)巨纤维纤维的巨系统图。(政府飞行服务队,以红色显示)巨人纤维突触电(峡路口)以及化学(胆碱)到外围形成突触中间神经(PSI,以绿色显示) Tergo Trochanteral肌肉神经元(TTMn,以黄色显示)5。在PS我要DLMn(背纵肌的神经元,以蓝色显示)连接是依赖于Dα7胆碱受体亚型6。最后到跳(TTM,紫色所示)和飞行肌的TTMn和DLMn器(DLM,紫色所示)神经肌肉接头(NMJ)是谷氨酸。
注:绿PSI的连接,电气和化学。然而,没有任何反应(缺乏差距路口)shakB突变,可以记录从DLM的政府飞行服务队的刺激后,在大脑中,这表明在没有电气连接的化学成分是不足以唤起动作电位PSI的5,16-18。 GF是因为PSI的连接缝隙连接依赖,这一数字仅显示为简单起见突触间隙连接。

图2。
显微操作小号ET-。
- 一个修改先前公布的协议14设置使用适合配对队同时nanoinjections录音注射显微操作。安装的飞行准备是面向对实验者的苍蝇头。注射微操作机器人(#1)被放置在前面的实验者之间的两个机器人钨刺激电极(2#和3#)。两个玻璃记录电极(#4和#5)显微操作,分别放在左边和右边。钨接地电极(#6)的微操作机器人被放置在后面最远的左侧(如图所示),或在右侧。
- 一位接近认为,从顶部的各种电极和注射微量的安排。
- 一个正确安装D。黑电极和注射微量注射刺穿。注意动物的身体其胸部水平和它的翅膀上传播出去。蜡牢固地缠它的身体,防止动物移动。此外,接地电极(#6,在腹部),记录电极的玻璃(#4和#5,在胸部,由黑暗的轮廓突出),(#2和#3,在每只眼睛刺激电极)被刺穿的地方,如前面所述14。微量注射液(#1)正确对齐三个单眼(圆圈)的中心。在这方面,应放在插入注射微量。

图3。斜面注射微量。适当斜面微量的图显示在这里。电极的开放应在45度角斜面,并有11至17微米之间的开放。一个适当的斜面注射微量是至关重要的顺利注入与最小ð豪悦国际来飞。

图4。总体方案的nanoinjection /电协议。一个代表图的整体方案,为nanoinjection /电协议。基线记录获得通过刺激巨纤维在100赫兹(GFS)10 10刺激(这里只显示了一列火车)列车启动。注射前,开始1 Hz的刺激一秒。在注射时间(喷油器插在控制箱),你会观察显着的背景噪音,但是,不要停止录音。注射后(从控制盒中拔出注射器),持续约1分钟为1 Hz的刺激。最后,继续强调政府飞行服务队10 10刺激列车在100赫兹,并继续测试这个范例中,每5分钟至15分钟的GF通路的功能。注:录音瓦特ERE操纵创建的总体方案,并不代表获得一个特定的结果。不向规模化,并非所有的痕迹。 点击图片放大 。

图5。政府飞行服务队的工作重点。
- 一个图形描述的影响α7胆碱受体拮抗剂Methyllycaconitine柠檬酸对果蝇GF-DLM的途径(MLA),在不同浓度(0,0.02,0.04,0.08,0.12毫微克/毫克N = 10%的复合治疗。 N = 15)生理盐水治疗。注射后只有一分钟,被视为一个重要和立竿见影的效果与0.04毫微克/毫克的工作重点。疗效显着,也看到与在0.8ng/mg和100Hz的刺激,政府飞行服务队的工作重点0.12ng/mg。盐水控制和0.02毫微克/毫克的工作重点之间无显着差异。此外,没有改变我1分钟后注射后,在测试的时间(15分钟),列印效果。生理盐水对照组(0毫微克/毫克)和工作重点,在每个时间点不同浓度之间进行t检验。报告为平均值+ / - 扫描电镜(SEM),* P <0.001水平。
- DLM的反应的样品在100 Hz刺激的痕迹。上部曲线显示工作重点后,广发刺激大脑中注射前的肌肉反应。注意能够应对在100赫兹到每个刺激肌肉。中东跟踪显示DLM的答复后,工作重点注射液(0.12毫微克/毫克)。请注意,肌肉是无法应对每个刺激在100赫兹到一。 (星号)。底部跟踪显示相同的准备后,在胸部的运动神经元的直接刺激(0.12毫微克/毫克)的DLM的反应。 DLM的回应胸部的刺激在100赫兹到,大脑的刺激与反应的失败可以归因于胆碱能PSI - DLMn连接的。
- 图形描述的工作重点不同浓度(0,0.02,0.04,0.08,0.12毫微克/毫克)的GF过去一年途径的影响。被视为在任何时间点盐水(1毫微克/毫克)和复方注射之间没有显着的影响。生理盐水对照组(1毫微克/毫克)和不同浓度的工作重点,在每个时间点之间进行t检验,* P <0.001。
- 在100 Hz刺激的TTM反应的样品的痕迹。上部曲线显示前工作重点注射后,刺激大脑中与广发激活肌肉的反应。请注意,是能够响应在100 Hz到所有刺激肌肉。中东跟踪显示后的工作重点注射液(0.12毫微克/毫克)的DLM反应。从TTM肌肉刺激大脑中的绿的反应保持一至一。底部的跟踪显示,过去一年的准备在胸部的运动神经元(0.12毫微克/毫克)100 Hz刺激的反应。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
这里介绍的nanoinjection /电生物活性,可快速筛选的化合物在果蝇的神经系统。这是一个小说在体内的技术,需要少量的化合物,在一个特点的神经电路引起的各种分子靶点的作用。这种方法可以用来测试不同的化合物的生物活性,从未知毒素市售药物制剂。
在这里,我们证明了我们的检测功能,使用工作重点,其中有巨人光纤系统(GFS)的果蝇( 图5)效果。我们发现,它选择性地破坏了GF DLM的途径,但不是绿过去一年途径。直接通过表明,绿DLM的途径的缺陷是由于在神经肌肉接头(NMJ)的功能障碍,但它是与对立effec一致胸刺激激活运动神经元T在Dα7胆碱受体亚型的工作重点,提出在PSI-DLMn突触( 图1)。虽然绿TTMn连接被证明是胆碱,它是未知的,是否Dα7胆碱受体亚基在这个突触。此外,胆碱乙酰(茶)基因或Dα7的胆碱受体亚型基因的情况下(Dα7)的基因,不破坏的GF TTMn连接的功能,因为同时存在电气交界5,6,17,19, 20,这使得途径不太可能受影响的工作重点。
复方注射后,溶液应立即浸入整个神经系统的动物,由于其开放式循环系统21。如果注入正常,该化合物通常在几秒钟内达到胸部和腹部,但可以均匀的分散到一分钟。但是,如果不正确的淋巴注入化合物(即注射微量太德EP进入脑组织),然后观察整个动物的分散较慢。而染料可用于实行适当的注射技术,如视频所示,它是不建议合作注入进行测试,因为它可能会改变化合物的性质,因此其生物活性的化合物与蓝色食用色素。此外,由于大部分作为溶剂使用的解决方案是明确的颜色(生理盐水,二甲基亚砜等),这是很难看到的化合物,不论是否是从注射针头喷出。因此,当溶解特定化合物,它是重要的,以确保它完全进入到解决方案;否则未溶解的颗粒会迅速堵塞注射针尖,防止任何流体弹射。此外,虽然整个淋巴立即化合物分散可能,在中枢神经系统达到的目标,以及达到它的最大剂量,可能需要更长的时间基础上的化合物的化学性质,如大小和polaritŸ,和其渗透苍蝇的血脑屏障。22这样的能力,它是重要的监测几分钟后注射未知化合物的潜在影响,因为不同的化合物,可以有发病影响的时间,这可能会随着时间的推移增加在某些情况下的变化。化合物,完全阻止神经元的功能强大和直接的影响已经可以看出,在1 Hz触发反应,而政府飞行服务队,在更高的频率(100赫兹)的刺激是用来检测由于低剂量或更微妙的影响化合物的效力。如果没有影响化合物注射后观察,也可以是由于无论是小的药物剂量或化合物分子的具体目标是不存在政府飞行服务队。
此外,使用这里的生物活性新化合物(如芋螺毒素)作为筛检工具时,重要的是要注意,检测中发现的分子靶点限制政府飞行服务队的苍蝇。虽然该法本身不允许的实际注入化合物分子的目标定位,它允许收窄内政府飞行服务队的潜在目标。可以做额外的测试,如补丁钳上神经或肌肉或与果蝇突变体的遗传相互作用的研究,以确定这些化合物的具体目标。最后,所提出的录制协议的目的是检测政府飞行服务队的作用拮抗作用。然而,可以很容易地调整录音协议,由被动的化合物,而不是测试,如果政府飞行服务队是不是能够应对可靠当电路被实验者刺激诱导反应的监测,监察斗殴影响。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
没有利益冲突的声明。
Acknowledgments
我们要承认在马里实验室和Godenschwege的实验室,特别是艾琳米泽成员,征求意见,并帮助这个协议。这项工作是由国立神经疾病研究所和中风授予R21NS06637 FM和TAG; AB公司是由国家科学基金会奖号码082925,URM的资助:为未来的研究人员结合生物学。
Materials
| Name | Company | Catalog Number | Comments |
| Recording glass electrodes: borosilicate glass capillaries | World Precision Instruments, Inc. | 1B100F-4 | 1.0mm OD, 0.58mm ID |
| Stimulator | Grass Technologies | Model S48 | |
| Amplifier | Getting Instruments, Inc. | Model 5A | |
| Data acquisition Software: Digidata | Molecular Devices | Model 1440A | |
| Data collection software: pCLAMP | Molecular Devices | Version 10 | |
| Stereomicroscope with fiber optic microscope ring illuminator | AmScope | SM-4T Model HL250-AR | |
| Dissecting scope for mounting | AmScope | SM-2TZ | |
| Kite Manual Micromanipulator & Tilting Base | World Precision Instruments, Inc. | Model # M3301 Kite: Model # KITE-M3-L | |
| Drosophila melanogaster Wild 10E genotype (wild type strain) | Bloomington Stock center | Stock # 3892 | |
| Vertical pipette puller | David Kopf Instruments | Model 700c | |
| Injection glass micropipettes: Borosilicate glass capillaries | World Precision Instruments, Inc. | Catalogue # 4878 | 1.14mm OD, 0.5mm ID |
| Silicon oil | Fisher Scientific | Catalogue # S159-500 | |
| Beveler | Sutter Instrument Co. | K.T. Brown Type Model # BV-10 | |
| Nanoliter2000 | World Precision Instruments, Inc. | Catalogue # B203XVY | |
| Blue food coloring | McCormick & Co. | N/A | Ingredients: Water, Propylene Glycol, FD&C Blue 1, and 0.1% Propylparaben (preservative). |
| Methyllycaconitine citrate (MLA) | Tocris Bioscience | Catalogue # 1029 | |
| Plastic wax sticks | Hygenic Corporation (Akron Ohio USA) |
References
- Koehn, F. E., Carter, G. T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 4, 206-220 (2005).
- Miljanich, G. P. Ziconotide: neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 11, 3029-3040 (2004).
- Layer, R. T., Wagstaff, J. D., White, H. S. Conantokins: peptide antagonists of NMDA receptors. Curr. Med. Chem. 11, 3073-3084 (2004).
- Lewis, R. J. Conotoxins as selective inhibitors of neuronal ion channels, receptors and transporters. IUBMB Life. 56, 89-93 (2004).
- Allen, M. J., Godenschwege, T. A., Tanouye, M. A., Phelan, P. Making an escape: development and function of the Drosophila giant fibre system. Semin. Cell Dev. Biol. 17, 31-41 (2006).
- Fayyazuddin, A., Zaheer, M. A., Hiesinger, P. R., Bellen, H. J. The nicotinic acetylcholine receptor Dalpha7 is required for an escape behavior in Drosophila. PLoS biology. 4, e63 (2006).
- Jan, L. Y., Jan, Y. N. L-glutamate as an excitatory transmitter at the Drosophila larval neuromuscular junction. The Journal of physiology. 262, 215-236 (1976).
- Usherwood, P. N., Machili, P., Leaf, G. L-Glutamate at insect excitatory nerve-muscle synapses. Nature. 219, 1169-1172 (1968).
- Marrus, S. B., Portman, S. L., Allen, M. J., Moffat, K. G., DiAntonio, A. Differential localization of glutamate receptor subunits at the Drosophila neuromuscular junction. The Journal of neuroscience : the official journal of the Society for Neuroscience. 24, 1406-1415 (2004).
- Petersen, S. A., Fetter, R. D., Noordermeer, J. N., Goodman, C. S., DiAntonio, A. Genetic analysis of glutamate receptors in Drosophila reveals a retrograde signal regulating presynaptic transmitter release. Neuron. 19, 1237-1248 (1997).
- Qin, G. Four different subunits are essential for expressing the synaptic glutamate receptor at neuromuscular junctions of Drosophila. The Journal of neuroscience : the official journal of the Society for Neuroscience. 25, 3209-3218 (2005).
- Schuster, C. M. Molecular cloning of an invertebrate glutamate receptor subunit expressed in Drosophila muscle. Science. 254, 112-114 (1991).
- Tanouye, M. A., Wyman, R. J. Motor outputs of giant nerve fiber in Drosophila. Journal of. 44, 405-421 (1980).
- Augustin, H., Allen, M. J., Partridge, L. Electrophysiological Recordings from the Giant Fiber Pathway of D. melanogaster. J. Vis. Exp. (47), e2412 (2011).
- Allen, M. J., Godenschwege, T. Drosophila Neurobiology. Zhang, B., Freeman, M. R., Waddell, S. , Cold Spring Harbor Laboratory Press. 215-224 (2010).
- Blagburn, J. M., Alexopoulos, H., Davies, J. A., Bacon, J. P. Null mutation in shaking-B eliminates electrical, but not chemical, synapses in the Drosophila giant fiber system: a structural study. J. Comp. Neurol. 404, 449-458 (1999).
- Thomas, J. B., Wyman, R. J. Mutations altering synaptic connectivity between identified neurons in Drosophila. J. Neurosci. 4, 530-538 (1984).
- Baird, D. H., Schalet, A. P., Wyman, R. J. The Passover locus in Drosophila melanogaster: complex complementation and different effects on the giant fiber neural pathway. Genetics. 126, 1045-1059 (1990).
- Gorczyca, M., Hall, J. C. Identification of a cholinergic synapse in the giant fiber pathway of Drosophila using conditional mutations of acetylcholine synthesis. J. Neurogenet. 1, 289-313 (1984).
- Allen, M. J., Murphey, R. K. The chemical component of the mixed GF-TTMn synapse in Drosophila melanogaster uses acetylcholine as its neurotransmitter. The European journal of neuroscience. 26, 439-445 (2007).
- Mejia, M. A novel approach for in vivo screening of toxins using the Drosophila Giant Fiber circuit. Toxicon. 56, 1398-1407 (2010).
- Stork, T. Organization and function of the blood-brain barrier in Drosophila. J. Neurosci. 28, 587-597 (2008).

{kind=link}