Attraction of Asian citrus psyllid (Diaphorina citri) adults to the odors of their natal host plant volatiles (citrus) is depicted in Figure 2A. Significantly (α < 0.05) more adults chose the arm of the T-maze olfactometer receiving odors from living citrus plants as compared with a blank (clean air) control.
An example of repulsion is shown in Figure 2B. In this case, the insects were exposed to one arm of the T-maze receiving citrus volatiles, while the second arm received volatiles from a citrus plant that was treated with the known repellent, dimethyl disulfide11. In this case, three types of behaviors were observed. The number of psyllids arrested at the release arm and not responding did not differ statistically from the number entering the arm receiving the odors from citrus plants alone (Figure 2B). However, no psyllids entered the arm containing the host plant volatile when co-presented with the repellent (Figure 2B).
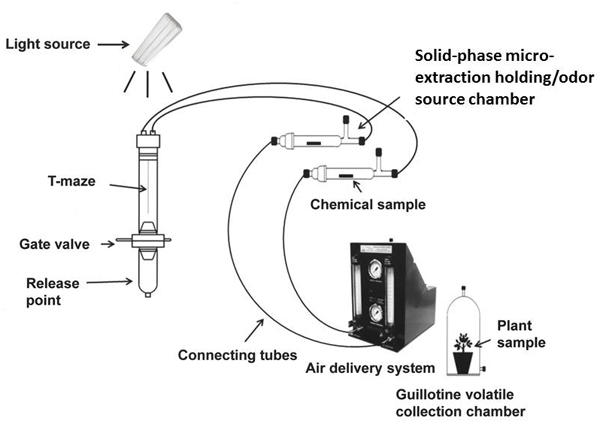
Figure 1. Schematic diagram of vertically oriented T-maze olfactometer joined to air delivery system and odor treatment release devices. The diagram is an adaptation of Figure 1 in Mann et al.10. A test subject is placed individually into the release chamber of the 2-port divided T-olfactometer. It moves towards the odors through a gate valve which allows controlled upward movement into the olfactometer and prevents the insect from crawling back into the release point area. The tube bifurcates 10 cm after the gate value. A positive response to an odor is recorded once the insect moves 0.5 cm into a specific side of the division. The guillotine volatile collection chamber may be used in place of a solid-phase micro-extraction holding/odor source chamber (SPMEC) as an odor source.

Figure 2. Response of Asian citrus psyllid (Diaphorina citri) adults to volatiles from citrus host plants versus clean air control (A) or citrus volatiles versus citrus volatiles co-released with a repellent (dimethyl disulfide) (B) in a vertically oriented T-maze olfactometer. Figure 2A: One-way analysis of variance (ANOVA) followed by Tukey’s HSD test (α < 0.05) was performed to compare the number of psyllid making a choice between either arm (2 treatments, n = 120). Columns indicated by different letters are significantly different from one another (α < 0.05). Figure 2B: One-way analysis of variance (ANOVA) followed by Tukey’s HSD test (α < 0.05) was performed to compare the number of psyllid making a choice between either arm or remaining in the release point (3 treatments, n = 120). Columns indicated by different letters are significantly different from one another (α < 0.05).
