

Summary
Den cytotoxiska aktiviteten hos fenantridin PJ-34 i cancerceller som genomgår mitos dokumenterades i realtid genom levande konfokal avbildning. PJ-34 utrotas human bröstcancer MDA-MB-231 celler, som hyser extra-centrosomes i mitos. Till skillnad från vanlig bifokala mitos, var extra-centrosomes inte klustrade i de två spindeln poler i närvaro av PJ-34.
Abstract
Fenantrenderivaten agerar som potenta PARP1 hämmare förhindrade bifokala klustring av övertaliga centrosomes i flera centrosomal humana cancerceller i mitos. Den fenantridin PJ-34 var den mest potenta molekylen. Declustering av extra-centrosomes orsakar mitotic misslyckande och celldöd i flera centrosomal celler. De flesta fasta humana cancrar har hög förekomst av extra-centrosomes. Aktiviteten av PJ-34 dokumenterades i realtid genom konfokal avbildning av levande humana bröstcancer MDA-MB-231-celler transfekterade med vektorer som kodar för fluorescerande γ-tubulin, som är rikligt i de centrosomes och för fluorescerande histon H2b närvarande i kromosomerna. Avvikande kromosomer arrangemang och de-klustrade γ-tubulin brännpunkter representerar declustered centrosomes upptäcktes i de transfekterade MDA-MB-231-celler efter behandling med PJ-34. Un-klustrade extra-centrosomes i de två spindel polerna föregick deras celldöd. Dessa resultat kopplade for första gången nyligen upptäckt exklusiva cytotoxiska aktiviteten hos PJ-34 i humana cancerceller med extra-centrosomes de-klustring i mitos, och mitotic misslyckande leder till celldöd. Enligt tidigare fynd som observerats med konfokal avbildning av fasta celler, PJ-34 uteslutande utrotas cancerceller med multi-centrosomes utan att försämra normala celler genomgår mitos med två centrosomes och bi-focal spindlar. Denna cytotoxiska aktivitet av PJ-34 delades inte av andra potenta PARP1 hämmare, och observerades i PARP1 bristfälliga MEF härbärgerar extracentrosomes, vilket tyder på dess självständighet av PARP1 inhibition. Live konfokala imaging erbjuds ett användbart verktyg för att identifiera nya molekyler utrota celler under mitos.
Introduction
Fenantren härstammar PARP1 hämmare, inklusive PJ-34, var avsedda att skydda vilande celler från apoptotiska celldöd inducerad av energikrävande PARP1 medierad DNA-reparation enligt stressbetingelser (stroke eller hjärtinfarkt) 1. Men nyligen upptäckte vi att PJ-34, två gånger högre koncentration än den inducerande PARP1 inhibition, uteslutande kan orsaka celldöd i humana cancerceller 2,3. Den snabbare proliferationen av cellen var, var den effektivare utrotning av cellerna. Den cytotoxiska aktiviteten av PJ-34 tillskrevs extra-centrosomes de-klustring i mitos 2. Många humana cancerceller hamnen multicentrosomes 4,5. Inkubation av humana bröstcancerceller MDA-MB-231 som hyser övertaliga centrosomes, med 20 ^ M PJ-34 effektivt utrotas dessa celler inom 72-96 h utan att försämra vilande celler eller några godartade prolifererande celler härbärgerande två centrosomes i mitos
Bipolär centrosomen montering är avgörande för bipolär spindel formation i mitos 4,5. Därför har celler med mer än två centrosomes utvecklat en knappast förstådd molekylär mekanism är kluster sina extra centrosomes vid två poler 4-9. Underlåtenhet av bipolär montering av sina centrosomes kan orsaka multipolär förvrängd spindlar och avvikande kromosomer segregation som hejdar cellen-cykeln i G2 / M gripandet, och leder till celldöd tillskrivs mitotic misslyckande 4,5. De molekylära mekanismerna bakom extra-centrosomes de-klustring är intensivt undersökt <sup> 10. Att förstå denna död mekanism kommer att exklusivt utrota cancerceller utan att skada friska vävnader 5,10.
Således, föreningar som aktiverar mitotic död katastrof cell erbjuder en ny form av ett selektivt cancerterapi, som kan vara effektiva i ett brett spektrum av humana fasta cancers.Our resultat tyder på att konfokal avbildning kan användas för att identifiera molekyler som påverkar extra centrosomes klustring i mitos 2,3, vilket gör dessa föreningar cancer inriktning läkemedelskandidater.
Vi har dokumenterat den cytotoxiska aktiviteten hos fenantridin PJ-34 genom att skanna fasta och levande humana cancerceller (med hög förekomst av extra-centrosomes i mitos) kontra normala celler. En steg-för-steg beskrivning av imaging metoder som används för att identifiera den cytotoxiska aktiviteten av PJ-34 i humana cancerceller ingår nedan.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Ett. Cell Culture Framställning
MDA-MB-231-celler köptes från ATCC (American Type Culture Collection) och lagrades i flytande kväve.
- Seed 10 6 MDA-MB-231-celler i 92 mm diameter petriskål i 10 ml komplett medium innehållande Dulbeco Modified Eagle Medium (DMEM), 10% hästserum, 1% L-glutamin och 1% Penstrep-Amfotericin B. Låt celler att proliferera till ca 80 till 100% konfluens.
- Ta bort odlingsmedium från skålen och släng.
- Tvätta cellskiktet kortvarigt med 0,25% (vikt / volym) trypsin-EDTA-lösning för att avlägsna alla spår av serum.
- Tillsätt 2,0 ml trypsin-EDTA-lösning för att dela och observera celler genom inverterat mikroskop tills cellager sprids (vanligen inom 5 till 15 minuter).
- Lägg 18 ml komplett odlingsmedium och försiktigt aspirera cellerna med pipett. Överför till en tub.
- Centrifugera cellsuspensionen vid 1200 rpm.
- Återsuspendera cellpelleten i 24 ml culture medium.
- Tillsätt 2 ml av cellsuspension till 35 botten mm glas rätter (ca 25% sammanflödet) och plats i inkubatorn (5% CO2, 37 ° C).
- Lösningar:
- Komplett Medium för celler proliferation: DMEM med 10% FBS, 1% antibiotika (100 enheter / ml penicillin G, 100 pg / ml streptomycin, Pen-Strep-amfo lösning) och 2 mM L-glutamin.
- Trypsin-EDTA-lösning, innehållande 0,25% trypsin-EDTA.
- Rätter:
92 mm petriskål.
35 mm diameter poly-D-lysin belagda glas botten kultur rätter.
2. Beredning av celler för Live Confocal Imaging
- Seed 2 x 10 5 MDA-MB-231 celler i kultur glas botten rätter i 2 ml komplett medium som nämns i avsnitt 1. När cellodling når sammanflödet av 60-70% (ca 3-4 x 10 5 celler per skål), fortsätt med transfektion.
- Transfektera cellerna med två plasmider ENCODning Fusionsproteinerna γ-Tubulin-GFP (för fluorescerande detektering av centrosomes) och Histone-RED (H2b-RED, för fluorescerande detektering av kromosomer) med liposomala transfektionsreagens Jet-PI, efter tillverkning protokollet. Kortfattat, blanda 2 pg från varje plasmid i ett rör med 100 | il NaCl (150 mM). Blanda transfektionsreagens (100 | il) med 100 | il NaCl (150 mM) i ett andra rör, och inkubera 5 min vid rumstemperatur (RT). Sedan kombinera de två lösningarna, blanda (med milt vortex) och spinn-ned. Inkubera under 30 min vid RT.
- Under inkubationen av transfektionsblandningen, tvätta cellerna en gång med PBS och ersätta cellmediet med 2 ml varmt DMEM utan några tillägg (37 ° C).
- Tillsätt försiktigt transfektionsblandningen till cellerna i DMEM och sedan återvända cellerna till inkubatorn (37 ° C, 5% CO2) under 8 timmar.
- Efter 8 h av inkubation ersätta DMEM med 2 ml komplett medium och inkubera cellerna i inkubator under 24tim.
- 24 timmar efter transfektion, byt mediet av cellerna med 2 ml komplett medium innehållande 20 iM PJ-34.
- Inkubera cellerna i ytterligare 18 h (37 ° C, 5% CO2).
- Ämne cellerna att leva konfokala avbildning under minst 16 timmar i bildbehandling kammaren hålla cellerna vid 5% CO2 och 37 ° C.
- Parallellt undersöker transfektion effektivitet 36 timmar efter transfektion med fluorescerande mikroskopi som följer:
- Seed 2 x 10 5 MDA-MB-231-celler i 6-brunnars platta innehållande ett täckglas per brunn i 2 ml komplett medium.
- Transfektera cellerna som nämns i avsnitten 2,2-2,5.
- 36 h efter transfektion fixera de transfekterade cellerna monterade på ett täckglas genom inkubation i kall metanol: aceton (01:01)-lösning, 7 min, -20 ° C.
- Sug fixeringen lösningen och låt täckglas med de monterade cellerna att torka i ett dragskåp.
- Ansök Förläng reagens Guld antifade med DAPI och låt the täckglas för att torka i mörker under 6 timmar.
- Undersök glider under fluorescerande mikroskop och beräkna procentandelen av de transfekterade cellerna (röda och gröna signaler) från den totala populationen av celler (DNA färgning av DAPI). Den önskade transfektion andel är ca 20-40% när 100-200 celler räknas.
Tre. Tekniska parametrar för Live Konfokala inställningar Imaging Scanner
- ScanMode XYZT, Pinhole [luftig] 1,00; Zoom 3.5, Upplösning 8 bitar, laser DPSS 561 nm, Argon, synlig laser 488 nm, laser He / Ne synliga 633 nm, Mål HCX PL APO CS 63X 1.40 OIL UV, numerisk bländare 1,4; Scan speed 700 Hz, brytningsindex 1,52.
- Bild 3-D presentation framställdes genom Imaris bildbehandlingsprogram 7.0.
4. Confocal avbildning av Mitos i fixerade celler
- Seed 2 x 10 5 MDA-MB-231-bröstcancerceller (ATCC), normala mus embryonala fibroblaster (MEF), eller PARP1 bristfällig MEF (PARP - / -, framställd av Dr Francoise Dantzer) på täckglas i 6-brunnar i 2 ml komplett medium. Täckglas tvättades med 96% etanol, följt av tvätt med sterilt DD vatten, torkades under 2 timmar, och placerades i varje brunn i 6-brunnars skål.
- Lägg PJ-34 (10-30 M) till mediet och inkubera cellerna för den tid som krävs (oftast upp till 96 timmar).
- Tvätta täckglasen en gång med PBS (fosfatbuffrad saltlösning), och fixera cellerna med användning inkubation i iskall metanol: aceton (01:01)-lösning, 7 min, -20 ° C.
- Aspirera fixeringslösning och låt täckglasen att torka i dragskåp (vid detta skede kan täckglasen förvaras i -20 ° C under flera veckor).
- Tvätta täckglasen en gång med PBST (PBS kompletterad med 0,1% Tween-20) för att permeabilisera cellmembranen och blockera cellerna med 10% NDS (Normal Donkey Serum) i PBST ("blockeringslösning") under 1 h vid RT.
- Inkubera permeabiliserade fixerade celler med primär antibodies under 2 timmar vid RT (för spindlar och centrosomes missfärgning). Antikropparna späds ut i den blockerande lösningen som följer: anti-α tubulin (1:250 utspädning) och anti-γ tubulin (1:200 utspädning). Primära antikroppar tillämpas enligt följande: gäller 100 l (i en droppe) i en blandning av antikroppar i blockerande lösningen för varje täckglas på 6-brunnar lock (locket är upp och ner). Försiktigt satte täckglas på antikroppar droppe, sådda celler inför drop. Inkubera täckglasen vetter antikropparna under 2 h vid rumstemperatur.
- Placera täckglasen tillbaka i brunnarna och tvätta cellerna tre gånger med PBST. Använd sedan samma procedur som beskrivs i 4.6 för märkning av celler på täckglasen med fluorescerande sekundära antikroppar. Inkubera cellerna på täckglasen med de sekundära antikropparna för 1 hr, RT, i mörker. Antikropparna späds ut i den blockerande lösningen som följer: Alexa Fluor 488 (1:1,000 utspädning, grönt) och Alexa Fluor 568 (1:1,000 utspädning;röd).
- Montera täckglas med Förläng Guld antifade reagens med DAPI (för kromosomer färgning) och inkubera över natten vid rumstemperatur i mörker för att torka.
- Undersök täckglas med konfokalmikroskopi.
Fem. Cellviabilitet Uppmätt av ATP-produktion
ATP-produktion mäts med ett luminiscent ATP detektionsanalys kit.
- Frö av cellerna i 96-brunnars platta, cirka 20.000 celler i 800 | il medium i varje brunn. Tre blankbrunnar bör användas för bestämning av bakgrunden luminiscens av mediet.
- Bered ATP standardspädningsserier från ungefär 10 um till 100 pm och hålla på is.
- Tillsätt 50 l av detergent till varje brunn och skaka plattan under 5 min i orbitalskakare, 700 rpm.
- Bered varje flaska av "frystorkad substrat" med 5 ml av "substrat buffert" i satsen.
- Tillsätt 50 l av den rekonstituerade substratlösning till brunnarna, och skakaplattan i 5 min på orbitalskakare, 700 rpm.
- Förvara plattan i mörker under 10 minuter.
- Mät Luminiscensläsare för varje brunn genom ELISA mikroplattläsare.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 är en stabil vattenlöslig fenantridin 1 (figur 1). Våra tidigare resultat avslöjade celldöd och de-klustrade extra centrosomes i flera typer av fasta multi-centrosomal cancerceller som behandlades med PJ-34. Däremot var normala celler som förökar sig inte nedskrivna 2,3. Centrosomes identifierades genom dubbel märkning med antikroppar riktade mot centrine1 och γ-tubulin i det fasta extra-centrosomal celler 2.
Här var den cytotoxiska aktiviteten av PJ-34 dokumenteras i dessa levande extra-centrosomal celler vid realtid genom att använda levande konfokalmikroskopi. Live human bröstcancer MDA-MB-231-celler, som har en hög förekomst (> 50%) av extra-centrosomes 4,5, har genomsökts efter minst 16 timmar efter konfokal avbildning fokuserad på transfekterade celler med γ-tubulin-GFP ( fluorescerande märkning av γ-tubulin foci 2) och med histon H2b-RED (fluorescerande labeling av kromosomer). Sex till tio levande transfekterade celler avsöktes parallellt i varje försök. Dubbel immunomärkning av γ-tubulin foci i de transfekterade cellerna med centrin1 var tekniskt omöjligt.
Spritt γ-tubulin foci och avvikande kromosomer arrangemang sällan upptäcks i slumpmässigt utvalda obehandlade MDA-MB-231 celler i mitos. Bifokala klustring av γ-tubulin foci, representerande extra-centrosomes bifokala klustring, dokumenterades i de flesta levande obehandlade MDA-MB-231 celler (Figur 2), var däremot un-klustrade centrosomes och avvikande arrangemang av kromosomer detekterades i levande transfekterade MDA-MB-231-celler inkuberade med PJ-34 (20 pM), och mitos i dessa celler avslutades genom celldöd (Figur 3). Dessa celler inkuberades med PJ-34 för 18 till 24 tim innan skanning och för ytterligare 16 h under skanning (Figur 3). Den realtid dokumentation av cell death under mitos stöder en tidigare definierad positivt samband mellan antal humana maligna celler med multipolär spindlar i mitos och andelen celldöd i celler som inkuberats med PJ-34 (20 ^ M) 2.
PJ-34 verkar som en potent inhibitor PARP1 1. Vi undersökte därför möjligheten att PARP1 hämning orsakar celldöd i samband med mitotic misslyckande. Till skillnad från levande avbildning, avbildning av fasta MDA-MB-231-celler aktiveras undersökning av en stor population av celler i cellkulturer, vilket möjliggör statistisk analys av effekterna av PARP1 hämmare i en mängd humancancercellinjer. Aktiviteten av PJ-34 jämfördes med aktiviteten hos andra potenta, icke-fenantren PARP1 hämmare i normal eller PARP1 fattiga celler (dvs. normal och PARP1 (- / -) möss embryonala fibroblaster (MEF)) (Figur 4). PARP1 bristfälliga MEF hamnen multi-centrosomes i mitos, men de är intet tumörceller 11. Dessa celler framställdes av Dr Francoise Dantzer, Strasbourg, Frankrike.
Fast normalt och PARP1 (- / -) MEF var immunolabeled för α-och γ-tubulin som märkt deras spindlar och centrosomes, respektive, som rapporterats tidigare två. Några av de undersökta cellkulturerna behandlades med PJ-34 eller andra potenta, icke-fenantren PARP1 hämmare, inklusive ABT-888 och AG01469, som inhiberar den enzymatiska aktiviteten hos PARP1, och BSI-201, kan en förening som tydligen dämpar PARP1 bindning till nicked DNA 12-14. Ingen av de testade PARP1 inhibitorer nedsatt normal MEF vid koncentrationer inhibera PARP1 aktivitet (Figur 4). Däremot PJ-34 dosberoende orsakas un-klustring av γ-tubulin foci, snedvridning av spindlar och celldöd i PARP1 (- / -) MEF (figur 4A och B). Detta observerades inte i normal MEF behandlats med PJ-34 (Figure 4B) eller i PARP1 (- / -) MEF behandlades med icke-phenenthrene PARP1 hämmare ABT-888 eller AG014699 (figur 4C). Det bör noteras att PJ-34 i koncentrationer som överstiger 20 iM gjorde försämrar normal MEF, trots normal MEF var mer resistent till PJ-34 aktivitet än PARP1 (- / -) MEF.
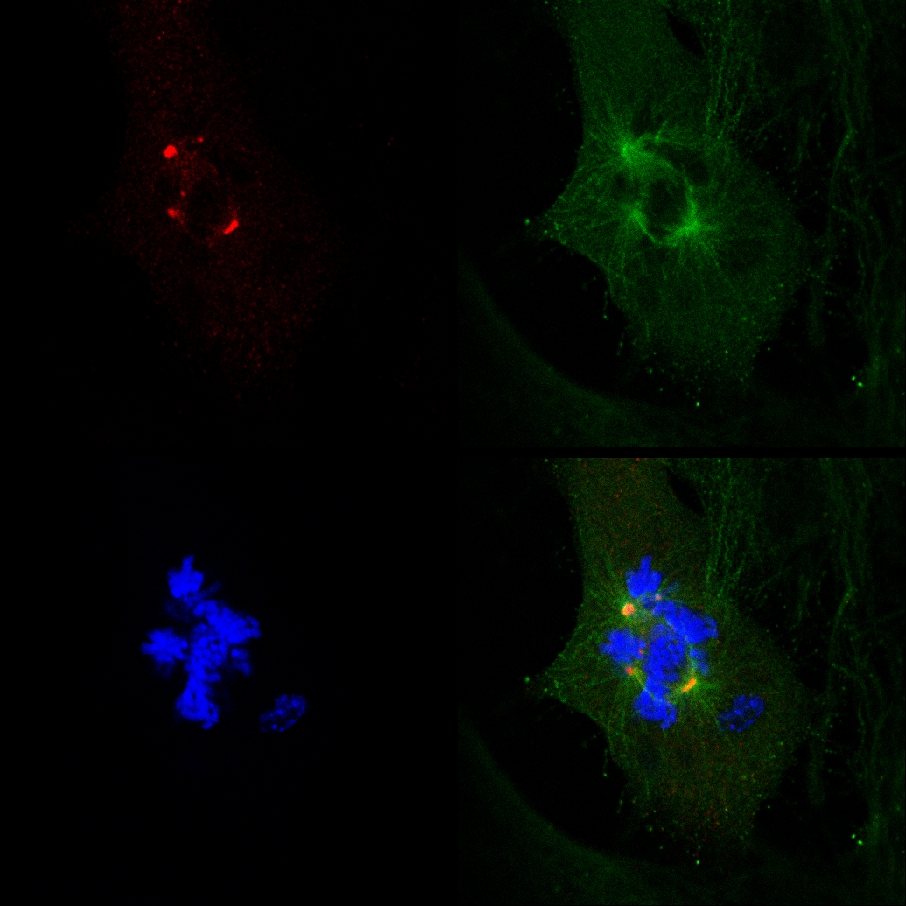
Det faktum att PJ-34 utrotat PARP1 (- / -) MEF trots deras PARP1 brist, och sambandet mellan bildandet av multi-focal spindlar och cell utrotning i PARP1 (- / -) MEF inkuberas med PJ-34 vid koncentrationer högre än de som krävs för PARP1 inhibition, inte var förenlig med en kausal koppling mellan extra-centrosomes de-kluster i PARP1 (- / -) MEF och PARP1 inhibition (Figur 4A). Den cytotoxiska aktiviteten av PJ-34 i PARP1 (- / -) MEF kan förklaras bättre av sin verksamhet som en extra centrosomes de-klustring medel i flera centrosomal celler 2 ( Kompletterande information "/> Figur 3. En live konfokala imaging dokumentation av celldöd i en slumpmässigt vald MDA-MB-231 celler i mitos med un klustrade-γ-tubulin märkta extra-centrosomes. MDA-MB-231-celler inkuberades med PJ-34 under 24 h före scanning och under 16 h av levande konfokal avbildning. PJ-34 applicerades 24 timmar efter transfektion med vektorer som uttrycker γ-tubulin-GFP (grönt, γ-tubulin foci och centrosomes) och H2b-RED (röd, kromosomer). 
Figur 1. Den fenantridin PJ-34: N-(6-oxo-5 ,6-dihydro-fenantridin-2-yl)-N, N-dimetyl-acetamid. 
Figur 2. Bi-fokus klustring av extra-centrosomes i en slumpmässigt vald levande MDA-MB-231 celler i mitos. A. Övre panel: Märkta centrosomes i en slumpmässigt utvald levande MDA-MB-231-cell transfekterad med γ-tubulin-GFP Nedre panelen:. Kromosom re-arrangemang under mitos i en slumpmässigt vald MDA-MB-231-cell transfekterad med histon H2b-RED. B. bifokala mitos med klustrade extra-centrosomes identifierats i en slumpmässigt vald kultmätt MDA-MB-231-cell. Cellerna transfekterades med både γ-tubulin-GFP (märkning γ-tubulin foci, grön) och histon H2b-röd (märkning kromosomer, röd). 48 h efter transfektion exponerades cellerna till en levande konfokal avbildning under 16 timmar. Sex celler skannas parallellt i varje experiment. Fyra olika experiment utfördes. Se även kompletterande information. Klicka här för att visa en större bild . 
Figur 3. Extra-centrosomes de-klustring föregås celldöd i levande MDA-MB-231-celler som behandlats med PJ-34. Ett slumpmässigt utvalda levande MDA-MB-231 celler i mitos med spridda centrosomes (1 st ram på vänster) avslutades genom celldöd ( 2: a och 3: e ramar). Denna cell valdes slumpmässigt i en cellkultur inkuberades under 24 timmar med PJ-34 (20 pM) anbringas 24 h efter transfektion med vektorer som uttrycker γ-tubulin-GFP (märkning γ-tubulin foci inklusive centrosomes, grönt) och histon H2b-RED ( märkning kromosomer, röd). Cellen igenom efter 16 timmar av levande konfokal avbildning. Sex celler skannas parallellt i varje experiment. Tre olika experiment utfördes. Se även kompletterande information. Klicka här för att visa en större bild . 
Figur 4. En cytotoxiska aktiviteten hos PJ-34 i PARP1 (- / -) möss embryonala fibroblaster. A. (vänster) Andelen Multifokal spindlar beräknas i normal (svart linje) och Parp1 - / - (grå linje) MEF, inkuberades under 48 timmar med PJ-34 vid de angivna koncentrationerna. Procentandelen multipolär spindlar beräknades av 20 totalt spindlar upptäcktes i 3 olika experiment. (Right) Minskad cellöverlevnaden upptäckts i cellkulturer som inkuberas i 72 timmar med PJ-34 (20 M) i förhållande till överlevnaden av kontroll obehandlade celler (normal (svart linje) och Parp1 - / - (grå linje) MEF). Cellöverlevnad analyserades genom cellernas ATP-produktion (protokoll 5). De medelvärden av fyra mätningar för varje cellinje i tre olika experiment presenteras B. Spindlar i slumpmässigt utvalda fixerad normal och Parp1 -. / - MEF i mitos, obehandlade (kontroll) eller inkuberades med PJ-34 under 48 h vid den indikerade koncentrationer. PJ-34 orsakade multipolär spindlar. Cellerna fixerades, permeabiliserades och immunolabeled för α-och γ-tubulin (grön märkning av spindlar och röd märkning av centrosomes, respektive). Kromosomer märktes med DAPI reagens (blå). . Representativa resultat av 3 olika experiment C. Potent icke-fenantren PARP1 hämmare inte påverkade centrosomes klustring i PARP1 (- / -) MEF. Spindlar av slumpmässigt utvalda normala och Parp1 - / - MEF presenteras, obehandlade MEF (kontroll) eller MEF behandlades under 48 h med icke-fenantren PARP-hämmare, AG01469 (20 | iM) eller ABT888 (20 | iM). Kromosomer är märkta med DAPI reagens (blå). Liknande resultat erhölls i 3 olika experiment. Klicka här för att visa en större bild .
Klicka här för att visa en större bild . 
Klicka här för att visa en större bild .
Figur 2B. Kompletterande figur. Ett slumpmässigt utvalda MDA-MB-231 celler i anafas med bi-focal klustrade γ-tubulin märkt extra centrosomes scannades under 16 timmar med levande konfokal avbildning, 48 timmar efter att transfekteras med vektorer som uttrycker γ-tubulin-GFP och H2b-RED (märkning γ-tubulin fokus och centrosomes i de fixerade celler (grön) och H2b histone märkning av kromosomerna (röda), respektive).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Live konfokal avbildning tillgänglig i realtid dokumentation av den cytotoxiska effekten av PJ-34 med levande multi-centrosomal celler under mitos (Figur 3 och Kompletterande information). Detta var den första levande dokumentation tillskriva cytotoxicitet av PJ-34 i humana cancerceller till extra centrosomes de-klustring och celldöd, vilket tyder på induktion av Mitotisk Catastrophe celldöd av PJ-34 5-9. Däremot var bi-focal klustring av super-numerary centrosomes observerats i levande obehandlade MDA-MB-231 celler som genomgår normal mitos med bi-Focali klustrade extra centrosomes (Figur 2, tilläggsinformation).
Enligt dessa resultat, lever konfokal avbildning av transfekterade celler kan vara användbart för att upptäcka translokation av proteiner inblandade i extra-centrosomes klustring i transfekterade celler under mitos 5,9,15,16. Identifiering av proteiner som påverkas av PJ-34 i multi-CentrosOMAL celler kan ge några ledtrådar för att förstå döden mekanismer som aktiveras av extra-centrosomes de-klustring.
Fördelen med levande konfokal avbildning i att ge information i realtid under mitos i levande celler, är också bunden till flera begränsningar. Antalet celler skannas per försök är begränsad. Därför är chansen att upptäcka celler i mitos är låga, och flera upprepade försök krävs för en realtids-dokumentation av mitos i skannade cellerna. Dessutom är framgång starkt beroende av hög effektivitet för transfektion av celler med vektorer som uttrycker märkta proteiner. Således, trots att tillförlitliga, är levande konfokal avbildning tidskrävande och kräver mycket erfarna hårda arbetare.
I jämförelse, immunocytokemi och konfokal avbildning av fixerade celler möjliggör undersökning av ett stort antal celler per försök, som krävs för en tillförlitlig statistisk analys. Vi använde denna metod för att jämföraeffekter av PJ-34 i mitos av normala godartade celler till dess effekter på cancerceller som genomgår mitos med övertaliga centrosomes 2,3. Likaså var denna metod som används för att jämföra effekterna av PJ-34 till den icke-fenantren PARP1 hämmare i PARP (- / -) hyser extra-centrosomes (celler med hög förekomst av multi-centrosomes 11) (Figur 4) MEF.
Sammanfattningsvis indikerar våra resultat fördelen att kombinera de mest värdefulla realtidsinformation tillhandahålls av konfokal avbildning av levande celler i mitos (figur 2 och 3) med cytokemi och konfokal analys av fixerade celler. En kombination av dessa metoder kan vara användbara för att identifiera den cytotoxiska aktiviteten av små molekyler som, liksom PJ-34, rikta särskilda mekanismer avgörande för cellens överlevnad. Den unika beroende av många mänskliga cancerceller på extra-centrosomes bipolära klustring för deras spridning och överlevdeal gör PJ-34 en möjlig kandidat för cancerterapi.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna förklarar att de inte har några konkurrerande ekonomiska intressen.
Acknowledgments
Finansieringskällor denna forskning: en gemensam fond av Tel Aviv University tekniköverföring företaget, RAMOT och Sheba-Medical Center (M. CA och SI.), ICRF - israelisk Cancer Research Foundation (M. CA.) Och Israel Science Foundation ( SI).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}